“determinación de aceites en algas marinas como...
TRANSCRIPT

CONSEJO NACIONAL DE CIENCIA Y TECNOLOGIA -CONCYT-
SECRETARIA NACIONAL DE CIENCIA Y TECNOLOGIA -SENACYT-
FONDO NACIONAL DE CIENCIA Y TECNOLOGIA -FONACYT-
FACULTAD DE CIENCIAS QUÍMICAS Y FARMACIA
UNIVERSIDAD DE SAN CARLOS DE GUATEMALA -USAC-
INFORME FINAL
“Determinación de aceites en algas marinas como potencial
materia prima para la producción de Biodiesel”
PROYECTO FODECYT No. 43-2007
Lic.Erick Giovanni Estrada Palencia
Investigador Principal
GUATEMALA, OCTUBRE DEL 2009.

2

3
AGRADECIMIENTOS
La realización de este trabajo, ha sido posible gracias al apoyo financiero dentro del Fondo
Nacional de Ciencia y Tecnología, -FONACYT-, otorgado por La Secretaría Nacional de
Ciencia y Tecnología -SENACYT- y al Consejo Nacional de Ciencia y Tecnología -
CONCYT-.

4
OTROS AGRADECIMIENTOS
A la empresa Guatebiodiesel por proveer las instalaciones físicas para el montaje de
laboratorio y ejecución de la parte experimental del proyecto, por proveer asesoría respecto
del uso de equipo y elaboración de biodigestores y proveer insumos para los análisis
químicos respectivos.
A la Escuela de Química de la Facultad de Ciencias Químicas y Farmacia, especialmente al
Departamento de Química Orgánica, Lic. Diana Pinagel por facilitar los permisos
necesarios para ejecutar la parte administrativa del proyecto y facilitar el espacio físico en
los diferentes laboratorios.
A la Licda. Karin Herrera del Laboratorio Microbiológico de Referencia (LAMIR) de la
Escuela de Química Biológica, por permitirnos el uso de equipo en dicho laboratorio. A la
Licda. María del Carmen Bran de la misma escuela, por proveernos de ciertos servicios,
utilizando su equipo de laboratorio. A la Licda. Norma Gil de Castillo y al Ing. Agr. Pedro
Julio García del Centro de Estudios del Mar y Acuicultura (CEMA) por proveernos del
espacio físico para establecer cultivos de microalgas.

5
Tabla de Contenido
PARTE I ..................................................................................................................... 10 I.1 INTRODUCCIÓN .................................................................................................... 10 I.2 PLANTEAMIENTO DEL PROBLEMA ................................................................. 11 I.2.1 Antecedentes ......................................................................................................... 11 I.2.2 Justificación .......................................................................................................... 11
I.3 OBJETIVOS ............................................................................................................. 13 I.3.1 General .................................................................................................................. 13 I.3.2 Específicos ............................................................................................................ 13 I.4 METODOLOGIA ..................................................................................................... 14 I.4.1 El Método ............................................................................................................. 14
I.4.1.1 Sitio de Muestreo ........................................................................................ 14 I.4.1.2 Colecta de muestras de fitoplancton ........................................................... 14
I.4.1.3 Identificación de microalgas ....................................................................... 16 I.4.1.4 Preparación y tratamiento del agua de mar ................................................. 16
I.4.1.5 Preparación de las soluciones madre ........................................................... 16 I.4.1.5.1 Extracto de suelo para medio Plymouth-erdschreiber ......................... 17
I.4.1.5.2 Vitaminas ............................................................................................. 18 I.4.1.6 Preparación de soluciones de enriquecimiento ........................................... 18 I.4.1.6.1 Medio Plymouth Erdschreiber ............................................................. 18
I.4.1.6.2 Medio f/2 de Guillard .......................................................................... 18 I.4.1.6.3 Medio f/2 de Guillard sin silicato ........................................................ 19
I.4.1.6.4 Hierro a concentración de f/2 ............................................................... 19 I.4.1.6.5 Fosfato a concentración de f/2 ............................................................. 19
I.4.1.6.6 Nitrato a concentración de f/2 .............................................................. 19 I.4.1.6.7 Medios de cultivo preparados .............................................................. 19
I.4.1.7 Inoculación de las soluciones de enriquecimiento ...................................... 20 I.4.1.8 Monitoreo y observación de los cultivos. ................................................... 20 I.4.1.9 Preparación de medio de cultivo sólido ...................................................... 20
I.4.1.10 Aislamiento por micromanipulación ........................................................... 21
I.4.1.11 Monitoreo de los cultivos establecidos por aislamiento por
micromanipulación ..................................................................................... 22 I.4.1.12 Purificación de cultivos obtenidos mediante aislamiento por
micromanipulación. .................................................................................... 22
I.4.1.13 Propagación de cultivos exitosos del aislamiento por micromanipulación 22 I.4.1.14 Cultivo en caja de Petri de cultivos monoespecíficos. ................................ 23 I.4.1.15 Preparación de inoculo de cultivos xénicos y axénicos .............................. 23
I.4.1.16 Producción de biomasa de microalgas ........................................................ 23 I.4.1.17 Cosecha de la biomasa de microalgas ......................................................... 23 I.4.1.18 Extracción de aceites de microalgas ........................................................... 23 I.4.2 La Técnica Estadística .......................................................................................... 24 I.4.2.1 Análisis de Agrupamiento Jerárquico ......................................................... 24
I.4.3 Los Instrumentos utilizados .................................................................................. 24 I.4.3.1 Material ....................................................................................................... 24 I.4.3.1.1 Cristalería ............................................................................................. 24

6
I.4.3.1.2 Reactivos .............................................................................................. 25 I.4.3.1.3 Otros materiales de laboratorio ............................................................ 25 I.4.3.1.4 Materiales eléctricos ............................................................................ 26
I.4.3.1.5 Otros materiales ................................................................................... 26 I.4.3.1.6 Medios de transporte ............................................................................ 27 I.4.3.1.7 Equipo .................................................................................................. 27
PARTE II ……………………………………………………………………...28
II.1 MARCO TEÓRICO ....................................................................................................... 28 II.1.1 Algas marinas ..................................................................................................... 28 II.1.1.1 Características generales ............................................................................. 28 II.1.1.2 Clase Bacillariophyceae (del Phylum Heterokontophyta). ......................... 28
II.1.1.2.1 Pared Celular ...................................................................................... 28 II.1.1.2.2 División Celular y la Formación de una Nueva Pared Celular .......... 29 II.1.1.2.3 Mobilidad ........................................................................................... 30
II.1.1.2.4 Productos de Almacenamiento ........................................................... 30
II.1.1.2.5 Ecología .............................................................................................. 30 II.1.1.2.6 Clasificación ....................................................................................... 31 II.1.1.3 Phylum Chlorophyta (Algas verdes). .......................................................... 31
II.1.1.3.1 Estructura Celular ............................................................................... 31 II.1.1.3.2 Fototaxismo y manchas oculares ........................................................ 32
II.1.1.3.3 Reproducción Asexual ........................................................................ 32 II.1.1.3.4 Reproducción Sexual .......................................................................... 32 II.1.1.3.5 Clasificación ....................................................................................... 33
II.1.1.4 Clase Cyanophyceae (Algas verde-azules) ................................................. 34
II.1.1.4.1 Morfología .......................................................................................... 34 II.1.1.4.2 Pared Celular y Deslizamiento ........................................................... 34 II.1.1.4.3 Vainas ................................................................................................. 34
II.1.1.4.4 Pigmentos y fotosíntesis ..................................................................... 35 II.1.1.4.5 Reproducción Asexual ........................................................................ 35
II.1.1.4.6 Crecimiento y Metabolismo ............................................................... 36 II.1.1.4.7 Ecología .............................................................................................. 36
II.1.1.4.8 Clasificación ....................................................................................... 36 II.1.1.5 Orden Dynophyta ........................................................................................ 36 II.1.1.5.1 Clasificación ....................................................................................... 37 II.1.1.6 Clase Chrysophyceae (del Phylum Heterokontophyta) .............................. 37
II.1.1.7 Estudios de Algas marinas en Guatemala ................................................... 38 II.1.2 Cultivos de microalgas ....................................................................................... 39 II.1.2.1 Cultivos de Enriquecimiento ....................................................................... 39
II.1.2.2 Dinámica del crecimiento de microalgas .................................................... 40 II.1.3 Biodiesel a partir de Aceites en microalgas ....................................................... 40 II.1.3.1 Aceites vegetales ......................................................................................... 40 II.1.3.2 Biocombustibles y Biodiesel ....................................................................... 41 II.1.3.3 Microalgas como productores de aceites .................................................... 41
II.1.3.4 Producción de biomasa de Microalgas ........................................................ 43 II.1.3.5 Estanques de Carril ..................................................................................... 44 II.1.3.6 Fotobioreactores .......................................................................................... 44

7
II.1.3.7 Comparación entre estanques de carril y fotobioreactores tubulares .......... 46
PARTE III ...........................................................................................................47 III.1 Resultados.................................................................................................................47
III.2 Discusión de Resultados ...................................................................................53
.....III.2.1 Diversidad de microalgas en los sitios muestreados........................................53
.....III.2.2 Aislamiento por micromanipulación....................................................................55
.....III.2.3 Cultivos establecidos a partir de líneas de reproducción de soluciones de
enriquecimiento .........................................................................................55
.....III.2.4 Purificación de especies aisladas .......................................................................55
.....III.2.5 Proporciones de inoculo para la producción de biomasa ..................................56
.....III.2.6 Rendimiento en la producción de aceites por especie de microalga..................56
PARTE IV ..........................................................................................................................57 IV.1 Conclusiones ..............................................................................................................57
IV.2 Recomendaciones ...................................................................................................59
IV.3 Referencias Bibliográficas .......................................................................................60
IV.4 Anexos .....................................................................................................................67
.....IV.4.1Micrografías de las especies de microalgas........................................................68
..........IV.4.1.1 Orden Bacillariophyta.................................................................................68
..........IV.4.1.2 Phylum Chlorophyta .................................................................................80
..........IV.4.1.3 Clase Cyanophyceae .................................................................................83
..........IV.4.1.4 Orden Dynophyta ......................................................................................83
..........IV.4.1.5 Otros Taxa ....................................................................................................86
PARTE V ........................................................................................................................87
V.1 Informe Financiero ......................................................................................................87
V.2 Cronograma de Actividades ........................................................................................88

8
RESUMEN
El presente estudio se realizó con el propósito de explorar la diversidad de especies de
microalgas (Diatomeas (Bacillariophyceae), clorofíceas (Chlorophyta), dinoflagelados
(Dynophyta) y flagelados marinos) y macroalgas marinas de la costa atlántica
guatemalteca, con la finalidad de obtener cultivos puros de algunas especies y determinar la
cantidad de aceites en su biomasa. A partir de estos resultados es posible proponer especies
potenciales para el cultivo masivo y la producción de energía limpia por medio de la
producción de Biodiesel. Se realizaron las colectas en 12 puntos de muestreo, de los cuales
se comparó la diversidad biológica de 5, los que también fueron sujetos a tratamientos de
cultivos de enriquecimiento. Se identificó un total de 44 especies, 32 en los sitios
comparados y 12 en los que no se compararon. Se aislaron 14 especies de microalgas, entre
ellas, Odontella aurita, Melosira nummuloides, Scenedesmus sp., Nannochloris sp.,
Entomoneis sp., incluyendo Pediastrum duplex y Botryococcus braunii, dos especies de
agua dulce. Todas se cultivaron en tubo de ensayo con medio de cultivo líquido, y otras se
lograron purificar para su cultivo en caja de Petri. Se establecieron cultivos en frasco de
vidrio para la producción de biomasa de las especies aisladas.
A partir de la biomasa seca obtenida de las especies cultivadas se realizaron extractos
utilizando el equipo de extracción Sohxlet. De estos resultados se propusieron algunas
especies para utilizarlas con la finalidad de proponerlas como potenciales fuentes de
biomasa para la producción de Biodiesel. Las especies Scenedesmus sp., Nannochloris sp. y
Navicula sp.1 presentaron los porcentajes más altos de aceites en biomasa seca.
Actualmente no existen trabajos bien documentados al respecto de microalgas del país,
tampoco se cuenta con un cepario de referencia, para realizar trabajos de investigación. Con
este trabajo se desea llenar esos vacios de información, así como comprobar si algunas de
las especies de microalgas podrían tener el potencial para la producción de aceites que
pueden ser utilizados como materia prima para la elaboración de biodiesel. Los resultados
obtenidos constituyen una fuente de información con la que no se contaba, ya que se
reportan más trabajos dedicados al estudio de macroalgas por parte de investigadores en las
universidades de Guatemala. Esta labor aporta una base de datos que contribuye al
conocimiento de la diversidad de microalgas del Atlántico guatemalteco, así como un
cepario que puede ser punto de partida para futuros proyectos de investigación. Además, es
un incentivo al sector industrial para aplicar la tecnología del cultivo de microalgas en
Guatemala, estando localizado en una región de alta incidencia solar y por ende, alta
productividad.

9
ABSTRACT
The following research was performed with the purpose to explore the diversity of
microalgae (diatoms (Bacillariophyta), green algae (Chlorophyta) and marine flagellates)
and macroalgae in the Atlantic coast of Guatemala, with the finality of obtaining pure
cultures of some species of microalgae and determine the amount of oil in the dry biomass.
The results made it possible to propose several potential species suitable for mass culture as
the principal process in the production of clean fuels in the form of biodiesel. Microalgae
samples where collected in 12 sampling sites, out of which 5 where compared for their
biological diversity and where also subjected to enrichment culture treatments. A total of 44
species of microalgae where identified, 32 in the compared sites, and 12 more in the ones
that were not compared. 14 species were isolated, to mention, Odontella aurita, Melosira
nummuloides, Scenedesmus sp., Nannochloris sp., Entomoneis sp., including Pediastrum
duplex and Botryococcus braunii, two fresh water species. They were all cultivated in test
tube with liquid culture media, and others where purified for its culture in Petri dishes.
Cultures were established in larger glass containers to produce biomass of the isolated
species.
From the dried biomass oils where extracted using the Sohxlet extraction apparatus. From
the results several species where proposed as potential sources of biomass for the
production of biomass. The species Scenedesmus sp., Nannochloris sp., and Navicula sp.1
showed the higher percentages of oil in the dry biomass.
At the present time there are no available well documented researches regarding microalgae
in Guatemala, neither is there a reference strain collection, appropriate for research. It is
intended with this work to fill information gaps, as well as knowing if some of the collected
and isolated species of microalgae prove to be useful for the production of oils as the basis
to produce biodiesel. The obtained results constitute a source of information which was not
available before. There is more research dedicated to macroalgae by researchers in the
universities of Guatemala. This work provides a data base, contributing to the knowledge of
microalgae diversity in the Atlantic coast of Guatemala, as well as a strain collection that
can serve as a starting point for future research. Furthermore, it may serve as an incentive
for the industry to start applying microalgae culture technology in Guatemala. Being
located in a region of high solar incidence, it is appropriate for providing high yields.

10
PARTE I
I.1 INTRODUCCIÓN
La siguiente investigación surgió a partir de la observación de la problemática ecológica y
energética mundial. La inestabilidad en los precios de los combustibles fósiles ha
conducido a diferentes entidades a fomentar la investigación y desarrollo de combustibles
de menor impacto ambiental que no dependan de fuentes conflictivas y poco estables. Este
es el caso de los biocombustibles, entre los cuales podemos mencionar el bioetanol, biogás,
biodiesel, hidrógeno, entre otros. En Guatemala el bioetanol es el combustible que más se
produce como derivado de la producción de la industria del azúcar. En el caso del biodiesel,
hay pocas plantas establecidas exitosamente en el país, las cuales tienen una baja
producción, comparada con la demanda nacional. La fuente de materia prima que estos
utilizan es, principalmente, el aceite utilizado en la industria alimenticia. Este aceite suele
ser escaso y de elevado costo. Por esto, se pretende con este proyecto, proponer fuentes
alternativas de aceite para la producción de biodiesel que no compitan con las cosechas
destinadas al consumo alimenticio de los seres humanos.
Para lograr este amplio objetivo inicialmente se realizan colectas en varios sitios de
muestreo en la costa Atlántico de Guatemala, desde Livingston hasta Punta de Manabique.
Se evalúa la diversidad de microalgas por sitio de muestreo y se compara la diversidad y
riqueza biológica entre estos. Se practican los procedimientos de cultivo de enriquecimiento
y aislamiento por micromanipulación para obtener cultivos monoespecíficos de especies de
microalgas. Se establecen cultivos en caja de Petri por medio de estriado con el propósito
de obtener colonias puras y lograr cultivos puros, libres de bacterias.
Los cultivos monoespecíficos en medio líquido en tubo de ensayo se utilizaron como
inoculo para medio de cultivo en frasco de 500 ml, a partir de los cuales se produjo
biomasa. Se acumuló la biomasa seca de las especies evaluadas y se realizó la extracción de
los aceites con el equipo Sohxlet utilizando hexano como solvente. Se obtuvieron
resultados alentadores por lo que se procedió a darle continuidad al mismo a nivel de planta
piloto, en un nuevo proyecto que actualmente se ejecuta con la intención de sintetizar
biodiesel a partir de los aceites extraídos a partir de las cepas que se trabajaron en este
proyecto. El cepario se mantuvo en constante mantenimiento para proveernos de inoculo
fresco y puro.

11
I.2 PLANTEAMIENTO DEL PROBLEMA
I.2.2 Antecedentes
En los últimos 30 años, se han registrado los 17 ó 18 años más cálidos del siglo. Dichas
condiciones han producido un cambio muy fuerte de las concentraciones de dióxido de
carbono en la atmósfera, relacionando directamente al calentamiento global y al efecto
invernadero. La emisión de gases a la atmósfera por la combustión de compuestos
orgánicos-fósiles, combustibles y productos secundarios de procesos industriales (dióxido
de carbono, el óxido nitroso y el metano) han contribuido con lo anterior y por lo tanto con
el deterioro del ambiente. El mundo industrializado ha conseguido que la concentración de
estos gases haya aumentado un 30% desde el siglo pasado, cuando, sin la actuación
humana, la naturaleza se encargaba de equilibrar las emisiones naturales. El impacto
potencial es enorme, con predicciones de falta de agua potable, grandes cambios en las
condiciones para la producción de alimentos y un aumento en los índices de mortalidad
debido a inundaciones, tormentas, sequías y olas de calor. En definitiva, el cambio
climático no es un fenómeno sólo ambiental sino de profundas consecuencias económicas y
sociales. Los países más pobres, que están peor preparados para enfrentar cambios rápidos,
serán los que sufrirán las peores consecuencias (Dix et al., 1999).
Diversos tratados y protocolos han impulsado el desarrollo de nuevas tecnologías con la
finalidad de reducir los efectos negativos en la atmósfera impulsando una nueva tendencia
hacia prácticas amigables con el ambiente. Tratados tales como: El Protocolo de Kyoto y la
Convención Marco de las Naciones Unidas sobre el Cambio Climático. (Dix et al., 1999 y
naciones Unidas, 1998), dentro de los cuales se encuentra la generación de medidas
alternativas de energía tal como el biodiesel, entre otros. Producción de este a través de
residuos vegetales, son una alternativa energética impulsada por ser fuentes renovables de
energía sumado al incremento en el precio del petróleo y su agotamiento (Romero, 2005).
Se consultaron varia fuentes bibliográficas para la elaboración de este trabajo de
investigación pero en Guatemala al menos en la Universidad de San Carlos de Guatemala, e
instituciones relacionadas con el temo no se han realizado estudios de microalgas pero si de
macroalgas como tema de investigación y elaboración de un libro elaborado en la Facultad
de Ciencias Químicas y Farmacia.
I.2.3 Justificación
Este trabajo surge de la inquietud de Guatebiodiesel, que es una empresa que se dedica a la
producción e investigación de biocombustibles en Guatemala, para los productores de
biodiesel en Guatemala es un problema muy grande la poca disponibilidad de materia
prima para la elaboración del mismo, la cual consiste en aceites fijos, que provienen en su
mayoría de cultivos de girasol, soya, palma, entre otros, el problema es que el costo de los
mismos es muy elevado, lo que hace poco rentable esta materia prima para la producción,
además que se fomenta el consumo y cultivo de estas plantas, con lo que se pone en riesgo
la biodiversidad y se compite a la vez con el uso primordial de estos aceites que es el de
consumo humano.

12
En India, Alemania, Estados Unidos, Canadá y Brasil, por mencionar algunos son pioneros
en la producción de biodiesel, pero ellos disponen de recursos económicos y grandes
extensiones de tierra las cuales se acondicionan para este propósito.
Guatemala no cuenta con esa disponibilidad de recursos económicos para invertir en la
investigación y desarrollo de proyectos que permitan la siembra de cultivos extensivos. Y
además no cuenta con disponibilidad de tierras con una extensión representativa que haga
económicamente viable su desarrollo.
Por lo que se buscan fuentes alternativas para la obtención de materia prima, abundante,
que no requiera de grandes extensiones de tierra, que no fomente el monocultivo y ponga
en riesgo la disponibilidad y diversidad de alimentos para consumo humano, y se sea
rentable su extracción a gran escala.

13
I.3 OBJETIVOS
I.3.1 General
Identificar las algas marinas de la costa atlántica guatemalteca y determinar la
presencia y concentración de aceites esenciales de estas.
I.3.2 Específicos
1. Identificar taxonómicamente las algas (microscópicas y macroscópicas)
predominantes de la costa atlántica guatemalteca.
2. Establecer el método de cultivo exsitu para la reproducción de las algas marinas
(microscópicas y macroscópicas) predominantes.
3. Comparar la presencia y concentración de aceites esenciales en los diferentes grupos
de algas macroscópicas y microscópicas de la costa atlántica guatemalteca
4. Determinar el método óptimo para la extracción de aceites esenciales en los
diferentes grupos de algas macroscópicas y microscópicas de la costa atlántica
guatemalteca.

14
I.4 METODOLOGIA
I.4.1 El Método
I.4.1.1 Sitio de Muestreo
Litoral Atlántico de Guatemala, Izabal:
- Latitud: 15° 44’16’’
- Longitud 88° 35’ 30’’
- Altitud promedio: 0 msnm
- Temperatura media: 27°C
- Temperatura mínima promedio anual: 22°C
- Temperatura máxima promedio anual: 33°C
- Precipitación anual: 3,700 mm/plg2
- Descripción del lugar: Izabal y su litoral Atlántico está localizado dentro de la zona
de vida de Bosque muy húmedo tropical (Holdridge, 1967), con temperaturas promedio
altas y fuertes lluvias. Río Dulce, que tiene 36 km de largo y desemboca en el Mar Caribe,
forma, a poca distancia del lago, un golfete que tiene 61,8 km² de área.
I.4.1.2 Colecta de muestras de fitoplancton
Se realizaron varias giras a Izabal para colectar muestras de fitoplancton a lo largo
de las costas del Mar Caribe guatemalteco. Se seleccionaron puntos de muestreo
distanciados, registrándose la localización geográfica de los sitios de colecta con un
geoposicionador (GPS). Las muestras se obtuvieron mediante el barrido de una red de
fitoplancton con una malla de 50 μm durante unos 5 minutos justo bajo la superficie del
agua. Para cada sitio se colectaron dos muestras, una para realizar la fase experimental y la
otra para observación bajo el microscopio e identificación. Cada muestra se almacenó en un
bote de plástico, la primera conservada en una hielera con hielo, y la segunda fijada con
lugol. En la primera gira se utilizó una sonda multiparámetros para medir los parámetros
del agua en cada sitio de muestreo. En la Tabla 1 se muestran los sitios con sus nombres y
coordenadas geográficas. La Figura 1 muestra los sitios de colecta localizados utilizando el
programa Google Earth TM
. Cinco de las muestras de cultivo se utilizaron para realizar
cultivos de enriquecimiento y hacer una comparación de la biodiversidad entre los sitios de
muestreo, tal como se describe más adelante.

15
Tabla 1. Coordenadas geográficas de los puntos muestreados.
Sitio de Muestreo Coordenadas hexagesimales
Punto 1 Punta Herrería 15°48’17.00”N, 88°42’59.00”O
Punto 2 Punta de Palma 15°45’29.00”N, 88°38’41.00”O
Punto 3 Ensenada pequeña antes de Puerto Barrios 15°44’39.00”N, 88°37’58.00”O
Punto 4 Puerto Barrios 15°43’59.00”N, 88°36’28.00”O
Punto 5 Antes de Bahía La Graciosa 15°47’28.00”N, 88°35’16.00”O
Punto 6 Bahía La Graciosa 15°51’23.00”N, 88°33’44.00”O
Punto 7 Salida de Bahía La Graciosa 15°53’34.00”N, 88°32’60.00”O
Punto 8 Camino a Punta de Manabique 15°54’46.00”N, 88°34’1.00”O
Punto 10 Livingston (Desembocadura de Río Dulce) 15°49’21.00”N, 88°44’54.00”O
Punto 12 CECON Río Dulce 15°39’25.00”N, 88°59’59.00”O
Fuente: Proyecto Fodecyt 43-2007.
Figura 1. Mapa generado en Google EarthTM
mostrando los puntos de muestreo en la
costa del Mar Caribe de Guatemala.
Fuente: Proyecto Fodecyt 43-2007.

16
I.4.1.3 Identificación de microalgas
Se identificaron las especies de microalgas utilizando libros de taxonomía de microalgas
(Tomas, 1997) y sitios web con fotografías especializados en taxonomía (Sitio Web CCAP,
2008). Las especies que no se lograron identificar se tomaron como morfoespecies por
motivos prácticos. Además, se guardó un registro fotográfico digital de todas las especies.
I.4.1.4 Preparación y tratamiento del agua de mar
El siguiente tratamiento está basado en Andersen, 2005. El agua de mar se colectó en sitios
lejanos de la costa en recipientes de plástico de 55 Lts previamente lavados con HCl al 10%
por varios días y luego bien desaguados. Se transportó al laboratorio y se le dio el siguiente
tratamiento:
- Prefiltración: el agua se filtró al vacío a través de un embudo Buchner con filtros
de papel Whatman. Esto garantizó la eliminación de partículas grandes y exceso de
partículas pequeñas que pudieran dificultar la siguiente filtración.
- Filtración: se utilizaron filtros Whatman GF/F (fibra de vidrio) con poros de 1µm
con el equipo de filtración al vacío. El agua prefiltrada se pasó por este sistema para
obtener un agua libre de partículas de materia orgánica. Esto se hace para eliminar
microalgas, protozoos, zooplancton, hongos, otros microorganismos y partículas contenidas
en el agua mayores a 1μm.
- Almacenamiento: el agua filtrada se almacenó en recipientes plásticos, alejada de
la luz y el calor del exterior hasta el momento en que se utilizó.
- Esterilización: el agua filtrada se esterilizó en autoclave en recipientes de vidrio
durante 15 minutos a 15 psi.
I.4.1.5 Preparación de las soluciones madre
Se prepararon soluciones madre de los macronutrientes (Ver Tabla 2), micronutrientes (Ver
Tabla 3) y vitaminas (Ver Tabla 4). Estas se utilizaron para preparar grandes cantidades de
soluciones de enriquecimiento y los medios de cultivo Plymouth Erdschreiber (Tomkins et
al, 1995) y f/2 (Guillard, 1975). Se esterilizaron en autoclave y se almacenaron en botes
plásticos opacos herméticos, en refrigeración.

17
Tabla 2. Concentraciones de reactivos utilizadas para la preparación de soluciones madre
de macronutrientes.
Compuesto Solución Madre (g/L dH2O) Medio de Cultivo
NaNO3 75.0 g/L dH2O f/2 de Guillard
NaNO3 200.0 g/L dH2O Plymouth Erdschreiber
NaH2PO4 · H2O 5.0 g/L dH2O f/2 de Guillard
Na2HPO4 · 12H2O 20.0 g/L dH20 Plymouth Erdschreiber
Na2SiO3 · 9H2O 30.0 g/L dH2O f/2 de Guillard
Fuente: Proyecto Fodecyt 43-2007.
Estas soluciones se esterilizaron en autoclave luego de su preparación, y se utilizaron en la
preparación de los medios de cultivo bajo condiciones estériles.
Tabla 3. Concentraciones de reactivos utilizadas para la preparación de soluciones madre
de micronutrientes.
Compuesto Solución madre (g/L dH2O) Cantidad
MnCl2 · 4H2O 180.0g/L dH2O 1 ml
ZnSO4 · 7H2O 22.0g/L dH2O 1 ml
CoCl2 · 6H2O 10.0g/L dH2O 1 ml
CuSO4 · 5H2O 9.8g/L dH2O 1 ml
Na2MoO4 · 2H2O 6.3g/L dH2O 1 ml
FeCl3 · 6H2O ---------- 3.15 gr
Na2EDTA · 2H2O ---------- 4.36 gr
Fuente: Proyecto Fodecyt 43-2007.
A 950ml de dH2O se le agrega 1 ml de cada una de estas soluciones y se le disuelven 3.15
gr de FeCl3 · 6H2O y 4.36 gr de Na2EDTA · 2H2O para preparar una solución de metales
traza.
I.4.1.5.1 Extracto de suelo para medio Plymouth-
Erdschreiber
Se cernió la tierra en un cedazo de malla de 1 cm, se esparció y se quitaron a mano los
invertebrados y restos de materia orgánica. Se dejó secar al aire para luego cernirla en un
cedazo de malla entre 2 y 4 mm. Para la preparación del extracto se mezcló una parte (en
volumen) de esta tierra con dos partes de agua destilada y se esterilizó en autoclave a 15 psi
por 2 horas. Se dejó enfriar, se decantó el sobrenadante y se pasó por un filtro Whatman
No.1. Luego se guardó en recipientes para preparar tandas de medio de cultivo y se volvió a
esterilizar en autoclave durante 15 min (Tomkins et al, 1995).

18
I.4.1.5.2 Vitaminas
Para preparar la solución de vitaminas se agregó 1 ml de la solución madre de
cianocobalamina, 1 ml de la solución madre de tiamina y 200 mg de tiamina HCl (Vitamina
B1) a 950 ml de dH2O. Se aforó a 1000 ml y esterilizó por filtración. Fue necesario su
almacenamiento bajo congelación para evitar la desnaturalización (Anderson et al, 2005).
Tabla 4. Concentraciones de vitaminas utilizadas para la preparación de soluciones
madre.
Compuesto Cantidad Solución Madre
Cianocobalamina (Vitamina B 12) 1 ml 1.0 g/L dH2O
Biotina (Vitamina H) 1 ml 1.0 g/L dH2O
Tiamina · HCl (Vitamina B1) 200 mg --------------------
Fuente: Proyecto Fodecyt 43-2007.
I.4.1.5.3 Preparación de soluciones de enriquecimiento
Para el cultivo de las muestras de fitoplancton se prepararon las 6 soluciones de
enriquecimiento descritas a continuación.
I.4.1.5.4 Medio Plymouth Erdschreiber
Se preparó una solución de agua de mar al 95% combinando 902.5 ml de agua de mar
filtrada con 47.5 ml de agua destilada. Esta solución y las demás se esterilizaron en
autoclave a 15 psi durante 15 min. Luego se le agregó en condiciones asépticas 1 ml de la
solución de NaNO3, 1 ml de la solución de Na2HPO4 · 12H2O y 50 ml del extracto de suelo
(Tompkins et al, 1995).
I.4.1.5.5 Medio f/2 de Guillard
Se agregaron a 950 ml de agua de mar filtrada los componentes presentados en la Tabla 5.
Tabla 5. Volúmenes necesarios de soluciones madre para la preparación del medio de
cultivo f/2 de Guillard.
Cantidad Compuesto Solución Madre
1 ml NaNO3 75 g/L dH2O
1 ml NaH2PO4 · H2O 5 g/L dH2O
1 ml Na2SiO3 · 9H2O 30 g/L dH2O
1 ml Solución de metales traza f/2
0.5 ml Solución de vitaminas f/2
Fuente: Proyecto Fodecyt 43-2007.

19
Se aforó a 1 Lt con agua de mar filtrada y se esterilizó en autoclave. Es preferible si la
solución de vitaminas se agrega en condiciones asépticas posteriormente a la esterilización
(Guillard, 1975).
I.4.1.5.6 Medio f/2 de Guillard sin silicato
Se preparó el medio f/2 omitiendo la solución de Na2SiO3.
I.4.1.5.7 Hierro a concentración de f/2
Se agregó el FeCl3 · 6H2O y Na2EDTA · 2H2O en las mismas cantidades utilizadas en el
medio f/2, omitiendo todos los demás nutrientes. Luego se esterilizó en el autoclave.
I.4.1.5.8 Fosfato a concentración de f/2
Se agregó 1 ml de la solución madre de NaH2PO4 por litro de agua de mar filtrada.
Posteriormente se esterilizó en autoclave.
I.4.1.5.9 Nitrato a concentración de f/2
Se agregó únicamente 1 ml de la solución de NaNO3 a 75 g/L dH2O por litro de agua de
mar filtrada, esterilizándose posteriormente.
I.4.1.5.10 Medios de cultivo preparados
Se prepararon estas 6 soluciones de enriquecimiento para establecer los cultivos a partir de
las 5 muestras del mar. Se esterilizaron en autoclave. La siguiente tabla muestra la
denominación que se utilizó para rotular e identificar los tubos de ensayo con cada
tratamiento.
Tabla 6. Denominación utilizada para los medios de cultivo.
Denominación A B C D E F
Medio de
cultivo
F/2 de Guillard
con silicato
F/2 de Guillard
sin silicato
Plymoyuth-
erdschreiber
Fe-EDTA a
concentración
F/2
Nitrato a
concentración
F/2
Fosfato a
concentración
F/2
Fuente: Proyecto Fodecyt 43-2007.
Estos se sirvieron en un total de 120 tubos de ensayo estériles con tapadera de rosca: 20 con
15 ml de cada solución de enriquecimiento.

20
I.4.1.6 Inoculación de las soluciones de enriquecimiento
Utilizando pipetas volumétricas se inocularon con 1 ml de las 5 muestras de fitoplancton
(Muestras 3, 4, 5, 7 y 10) los 120 tubos de ensayo con las 6 soluciones de enriquecimiento.
Estos tubos se rotularon y se colocaron en un tramo de la estantería con luz superior a un
fotoperiodo de 12 horas.
I.4.1.7 Monitoreo y observación de los cultivos.
Los cultivos se monitorearon tres días alternos a la semana para observar los cambios y la
apariencia. Entre la tercera y cuarta semana de crecimiento se llevaron a cabo las
observaciones de los cultivos. Para esto se agitó primero cada tubo, desprendiendo las
células adheridas a la pared del tubo y homogenizando el cultivo. Se montaron tres gotas de
muestra sobre un portaobjetos, cubriendo cada una y se observaron en el microscopio de
luz a un aumento de 400X en la mayoría de los casos. Se anotaron los resultados de las
abundancias relativas de cada especie encontrada en cada tubo y se tabularon.
I.4.1.8 Preparación de medio de cultivo sólido
Se preparó medio de cultivo f/2 de Guillard con agar a concentración de 10gr/Lt
(semisólido) de la siguiente manera:
- Se preparó 1 Lt de medio de cultivo f/2 de Guillard con silicato de la manera
explicada previamente, aforando en un balón de 1000 ml.
- Se colocó un beaker de 1,000 ml sobre una hornilla eléctrica con agitador
magnético.
- Se agregaron 700 ml del medio de cultivo al beaker y comenzó a calentar.
- Se pesaron 10 gr de agar-agar en una balanza y vertieron dentro del beaker.
- Se inició la agitación y subió el calor a lo máximo controlando el momento en que
inició la ebullición para disolver el agar.
- Al iniciar la ebullición se agregaron los 300 ml restantes de medio de cultivo
líquido.
- Se sirvió equitativamente el contenido del beaker en tres erlenmeyers de 500 ml y
se taparon con papel aluminio o papel kraft.
- Se introdujeron a la autoclave junto con pipetas volumétricas de 1 ml, tubos de
ensayo con tapadera de rosca y otro material necesario, y esterilizaron en autoclave durante
15 minutos.
- Se vació la autoclave cuando bajó la presión.
- Se preparó la cámara de flujo laminar para servir el medio estéril.
- Se desinfectó
- Se introdujeron las cajas de Petri desechables y las pipetas
- Se introdujo el medio de cultivo estéril en erlenmeyers
- Se introdujo la solución estéril de vitaminas
- Se encendió la luz ultravioleta durante 20 minutos el ventilador.

21
- Se agregaron 0.5 ml de solución de vitaminas por cada litro de medio de cultivo,
agitando suavemente para evitar la formación de burbujas.
- Cuando el medio de cultivo se entibió a temperatura cómoda al tacto se sirvió en las
cajas de Petri y en tubos de ensayo con tapadera.
- Se colocaron las cajas de Petri en torres y empacaron en su bolsa original. Los tubos
se colocaron acostados con una leve inclinación para que solidificarse en esa posición.
- Se guardaron en refrigeración al haber solidificado el medio.
I.4.1.9 Aislamiento por micromanipulación
Se preparó la cámara de flujo laminar de la siguiente manera:
- Se desinfectaron todas las superficies con etanol al 70%.
- Se introdujo el microscopio invertido y se desinfectó del mismo modo.
- Se introdujeron cajas de Petri con medio de cultivo estéril, tubos de ensayo con
medio de cultivo líquido y sólido.
- Se colocaron portaobjetos, mechero de alcohol o incinerador eléctrico, pipetas
Pasteur estériles, pipeteadores, tubos capilares con la punta reducida, manguera
Tygon de 1mm de diámetro y 4 pies de longitud, beaker de 1,000 ml., papel
Parafilm, marcador permanente.
- Se encendió el ventilador y la luz ultravioleta durante 20 minutos antes de
comenzar.
- Se prepararon dos personas, una como asistente y otra para el aislamiento, con bata
limpia, manos lavadas minuciosamente y desinfectadas con etanol al 70%.
- Se escogieron los tubos de cultivos de enriquecimiento y las muestras del mar que
se utilizarían para la micromanipulación.
- Se conectó un capilar reducido a la manguera Tygon y se colocó alrededor del
cuello. Se flameó con cuidado el capilar para esterilizar.
- Se montó una muestra de cultivo de microalgas sobre un portaobjetos, sin colocar
cubreobjetos, sobre la platina del microscopio y se inició la búsqueda de
especímenes saludables de la especie de microalga deseada. En esta fase se apagó el
ventilador de flujo debido a que la vibración afecta en la observación y
micromanipulación.
- Se flameó un portaobjetos, y al enfriarse se colocaron 4 gotas separadas de medio de
cultivo estéril utilizando una pipeta Pasteur. El portaobjetos se puso sobre una
superficie libre del microscopio cerca de la platina.
- Se eligió un espécimen y se succionó con el capilar de la siguiente manera:
Se colocó el extremo libre de la manguera entre los labios, tapando el
orificio con la lengua.
Se tomó con una mano el extremo del capilar y se acercó a la
muestra, observando a una magnificación de 40X.
Al estar el capilar justo sobre el espécimen se introdujo la punta fina
del capilar, colocándola sobre la célula, y destapando el otro extremo
de la manguera se succionó por capilaridad. En caso que no la
absorbiera, se vaciaba el capilar y se volvía a intentar. En ocasiones
era necesario succionar con la boca.

22
- Se expulsaba el contenido del capilar dentro de la primera gota de medio de cultivo
estéril en el otro portaobjetos, colocando este sobre la platina.
- Fue posible aislar varias células (tandas) antes de proceder a limpiarlas, o limpiar
una por una.
- Para limpiar una célula (o cadena de células) se succionó de la misma manera la
célula aislada y se pasó por tres gotas de medio de cultivo estéril. Es aconsejable
flamear el capilar antes de succionar una célula.
- Habiendo succionado la célula de la última gota se conectó el ventilador.
- Se inoculó la célula sobre medio de cultivo sólido en caja de Petri o tubo de ensayo,
o en medio de cultivo líquido (f/2 o f/10 de Guillard) en tubo de ensayo.
- Si era en caja de Petri se podían inocular hasta 6 células, sellando con Parafilm.
- Los recipientes de cultivo se colocaron bajo iluminación en las estanterías a un
fotoperiodo de 12 horas, los tubos de ensayo en gradillas.
Los aislamientos se realizaron a partir de las muestras originales y de cultivos de
enriquecimiento que presentaran las especies deseadas.
I.4.1.10 Monitoreo de los cultivos establecidos por aislamiento por
micromanipulación
Los cultivos en tubo de ensayo así obtenidos se monitorearon periódicamente a partir de la
primera semana de su inoculación. Este consistió en la observación semanal de tubos bajo
el microscopio invertido, buscando evidencia de propagación de las células inoculadas. Se
apartaron los cultivos exitosos y mantuvieron en observación los tubos sin crecimiento. Los
cultivos contaminados por especies de microalgas generalistas (generalmente Navicula sp.)
o cianobacterias, y los que no presentaron crecimiento, se descartaron. Los tubos
descartados se reutilizaban luego de lavarse y esterilizarse.
I.4.1.11 Purificación de cultivos obtenidos mediante aislamiento
por micromanipulación.
En ocasiones los cultivos obtenidos consistían en dos o (raramente) tres especies, por lo que
se requirieron posteriores intentos de aislamiento por micromanipulación. Se realizó esto
para separar especies deseadas de especies generalistas que eran difíciles de eliminar en el
primer aislamiento.
I.4.1.12 Propagación de cultivos exitosos del aislamiento por
micromanipulación
A medida que se lograban establecer cultivos monoespecíficos (de una sola especie de
microalga), estos se propagaban constantemente en tubos de ensayo. De esta manera se
mantuvieron viables las líneas de reproducción y obtuvo una dotación constante de cultivos
aptos para ser utilizados como inoculo para cantidades mayores de medio de cultivo en la
producción de biomasa.

23
I.4.1.13 Cultivo en caja de Petri de cultivos monoespecíficos.
A partir de cultivos monoespecíficos en tubo de ensayo se realizaron cultivos en caja de
Petri usando el método de estriado y por goteo. Las cajas se sellaron con Parafilm TM
y se
colocaron bajo iluminación artificial a un fotoperiodo de 12 horas con la superficie del
medio de cultivo hacia arriba. Las colonias puras de microalgas obtenidas en medio de
cultivo sólido se inocularon nuevamente en tubo de ensayo con medio líquido, obteniendo
así cultivos axénicos (cultivos de una sola especie).
I.4.1.14 Preparación de inoculo de cultivos xénicos y axénicos
Los cultivos monoespecíficos en tubo de ensayo (xénicos o axénicos) se utilizaron como
inoculo para medio de cultivo líquido f/2 de Guillard en frascos de 500 ml. Estas
inoculaciones se realizaban dentro de la cámara de flujo laminar para reducir la
contaminación. Los frascos se colocaron bajo luz fluorescente a fotoperiodo de 12 horas y
aireación constante utilizando una bomba de acuario.
I.4.1.15 Producción de biomasa de microalgas
A partir de dos semanas de crecimiento los cultivos en frasco de 500 ml podían utilizar
como inoculo, cada uno para 4 botes de 1Lt con medio de cultivo líquido f/2 de Guillard.
Los botes inoculados se colocaban bajo el mismo sistema de cultivo. De dos a tres semanas
después se procedía a cosechar la biomasa producida.
I.4.1.16 Cosecha de la biomasa de microalgas
La biomasa de microalgas precipita en los medios de cultivo, por lo que solo basta con
dejar los botes en reposo y luego extraer con una pipeta y pipeteador las microalgas
sedimentadas. Se prepara previamente un embudo Buchner con un filtro Whatman GF/F
(fibra de vidrio) con poros de 1µm. La biomasa cosechada se va colocando en el embudo y
se deja filtrar por gravedad. Se lava dos veces con agua destilada dentro del mismo embudo
y se deja hasta eliminar la mayor cantidad de agua. Se colecta la biomasa con una pequeña
espátula de metal y se coloca en una porcelana o vidrio de reloj para secarla. Se coloca
sobre una lámpara de luz fluorescente para dejar secar durante la noche.
I.4.1.17 Extracción de aceites de microalgas
Para realizar la extracción de los aceites de biomasa de microalgas secas se siguieron los
siguientes pasos.
- Se armó el equipo Sohxlet, que consiste en un balón de fondo redondo, adaptador
Sohxlet y un condensador.

24
- Se pesaron 10 gr de biomasa de microalgas secas, se colocaron en un dedal y se
introdujeron dentro del adaptador Sohxlet.
- Se agregó éter de petróleo (u otro solvente orgánico apolar) al sistema (al balón y al
contenedor del Sohxlet).
- Se calentó gradualmente en baño María, manteniendo en ebullición y reflujo durante
4 horas aproximadamente.
- Al finalizar se eliminó el solvente por evaporación y se dejó enfriar.
- Se colectó el aceite extraído en un vial de vidrio previamente tarado y se pesó en
una balanza analítica.
- Se calculó el porcentaje de rendimiento para cada especie de microalga.
I.4.2 La Técnica Estadística
I.4.2.1 Análisis de Agrupamiento Jerárquico
Se realizó un análisis de agrupamiento jerárquico utilizando el programa estadístico SPSS
15.00. Se compararon los cinco sitios de muestreo seleccionados (3, 4, 5, 7 y 10) según la
presencia o ausencia de las especies, utilizándose el índice de similitud de Jacard. Se
ingresaron los datos en la hoja del programa a partir de los datos en Microsoft Excel.
Este análisis utiliza los datos de presencia y ausencia de las especies de microalgas
colectadas en cada sitio de muestreo por separado. Compara la similitud de cada sitio con
todos los otros sitios, siendo el valor más alto la comparación con el mismo sitio. A partir
de esto, elabora un dendrograma que relaciona los sitios según su similitud. Los sitios más
similares en su diversidad están más cercanos, y los que difieren en mayor grado quedan
más lejanos en el gráfico. Así, se crean grupos por similitud y se distinguen los sitios que
difieren más.
I.4.3 Los Instrumentos utilizados
I.4.3.1 Material
I.4.3.1.1 Cristalería
Ampolla de decantación
Balones aforados de 250 ml y 1Lt
Beakers (de 50 ml a 1Lt)
Cajas de Petri
Cubreobjetos
Embudos Buchner de 47mm y 95mm
Equipo Sohxlet para extracción
Erlenmeyers de 500 ml y 1Lt
Frascos ámbar de 250 a 1 Lt
Frascos de 500 ml

25
Kitasatos de 1 Lt
Pipetas de 1 a 10 ml
Pipetas Pasteur
Portaobjetos
Probetas de 100 ml y 1 Lt
Tubo Pyrex de 5mm de diámetro
Tubos capilares
Tubos de ensayo con tapadera de rosca
Varillas de agitación
Vidrios de reloj
I.4.3.1.2 Reactivos
Agar-agar
Agua destilada
Biotina
Cianocobalamina
CoCl2 · 6H2O
CuSO4 · 5H2O
Etanol 95%
Éter de petróleo
FeCl3 · 6H2O
H2O2
Lugol
MnCl2 · 4H2O
Na2EDTA · 2H2O
Na2MoO4 · 2H2O
Na2SiO3 · 9H2O
NaH2PO4 · H2O
NaNO3
Tiamina
ZnSO4 · 7H2O
I.4.3.1.3 Otros materiales de laboratorio
Aceite de inmersión
Agitadores magnéticos
Asas en argolla
Asas en punta
Cajas de Petri desechables estériles
Filtros de membrana de celulosa de 0.7 µm
Filtros Whatman de papel

26
Filtros Whatman GF/F de poro de 1µm
Gradillas para tubos de ensayo
Manguera Tygon de 1mm diámetro interior
Mechero de alcohol
Papel limpialentes
ParafilmTM
Pipeteadores
Pisetas
Succionadores para pipetas Pasteur
I.4.3.1.4 Materiales eléctricos
Alambre paralelo calibre 12
Espigas
Regletas
Temporizador
Tomacorrientes
Tubos fluorescentes 40W blanco cálido No.12
I.4.3.1.5 Otros materiales
Alambre galvanizado delgado
Algodón
Bolsas plásticas grandes
Botes plásticos blancos de 500ml
Botes plásticos de 1Lt
Cepillos para lavar cristalería
Cortador de alambre
Desarmadores
Estanterías plásticas
Hielera
Hielo
Jabón
Manguera para acuario
Marcador permanente
Masking tape
Papel aluminio
Papel mayordomo
T’s para bomba de acuario
Tijeras grandes
Válvulas de 3 y 5 salidas para bomba de acuario

27
I.4.3.1.6 Medios de transporte
Camionetilla o Pickup diesel
Lancha con motor de 175 caballos de fuerza
I.4.3.1.7 Equipo
Autoclave
Balanza analítica
Bomba de vacío
Bombas aireadoras para acuario
Cámara de Flujo Laminar
Cámara fotográfica digital
Conductímetro – salinómetro
GPS
Hornilla con agitador magnético
Microscópico óptico Olympus 40X a 1000X
Microscopio invertido Olympus 40X a 400X
Red de plancton
Refrigerador
Sonda Multi-parámetros

PARTE II
II.1 MARCO TEÓRICO
II.1.1 Algas marinas
II.1.1.1 Características generales
Las comunidades de algas marinas están constituidas por organismos micro y
macroscópicos. Las más abundantes y las mayores concentraciones de lípidos se
almacenan en las algas microscópicas, las cuales se agrupan en varios grupos de los cuales
los siguientes son de alta importancia:
II.1.1.2 Clase Bacillariophyceae (del Phylum Heterokontophyta).
Es probable que este grupo haya evolucionado a partir de un miembro escamoso de las
Chrysophyceae o Bolidophyceae (Guillou et al., 1999). Las diatomeas son algas
unicelulares y, en ciertos casos, coloniales, que se encuentran casi en cualquier hábitat
acuático como autótrofos fotosintéticos de vida libre, heterótrofos incoloros o simbiontes
fotosintéticos (Schmaljohann et al, 1978). Se pueden encontrar como plancton o perifiton,
formando películas café verdoso sobre sustratos como rocas o plantas acuáticas. Las células
están rodeadas por una pared celular rígida similar a una caja de dos piezas, formada por
sílice (SiO2), llamada frústula. Los cloroplastos contienen clorofilas a, c1, y c2 y
fucoxantina, la cual provee a las células su característico color café dorado (Lee, 2008).
La frústula formada por dos partes rodea el protoplasma que contiene un núcleo más o
menos central suspendido por un sistema de hebras protoplasmáticas. Los cloroplastos
ocupan la mayor parte de la célula, a veces como dos plástidos parietales aunque en
ocasiones como numerosos plástidos discoides (Lee, 2008).
II.1.1.2.1 Pared Celular
La frústula consiste en dos piezas casi iguales, una de menor tamaño que casa con la mayor
como una caja de Petri. La externa es llamada epiteca y la interna, hipoteca. Cada teca se
compone de dos partes, la valva y la banda conectora, adherida al borde de la valva. Las
dos bandas conectoras forman la faja. El margen de las bandas se une por medio de una
película pectinacea. Si esta se destruye, la valva y las bandas se separan. El material silíceo
de la frústula es colocado en patrones regulares que producen una pared ornamentada. La
ornamentación de las diatomeas puede ser dividid en cuatro tipos básicos: 1) Céntrica y
radial, donde la estructura es ordenada en relación a un punto central; 2) Trelisoide, donde

29
se arregla uniformemente sobre la superficie sin relación a un punto o línea; 3) Gonoide,
donde la estructura está dominada por ángulos; 4) Penada, donde la estructura está
simétricamente ordenada a ambos lados de una línea central (Lee, 2008). Algunas
diatomeas penadas tienen un sistema de rafe, formado por un rafe una apertura longitudinal
en la teca), dividido en dos partes por un nódulo central. Cada mitad del rafe termina en un
ensanchamiento de la pared llamado nódulo polar. La ornamentación en las diatomeas
penadas es bilateralmente simétrica alrededor del rafe. En las diatomeas penadas que no
poseen un sistema de rafe, existe un área ornamentada que surge del centro de la valva, a lo
que se le llama pseudorafe. También existen otros dos tipos de perforaciones de la pared en
las diatomeas, el poro simple y el lóculo o areola. El poro consiste en un agujero simple con
una pared silicificada homogénea que en ocasiones está reforzada por costillas y costae. El
lóculo consiste en una cámara hexagonal en la pared, que está separada de otros lóculos por
medio de espaciadores verticales, que en ocasiones poseen poros para permitir la
comunicación entre lóculos. Esto da a la valva la semejanza de un panal (Lee, 2008). Varias
especies de diatomeas poseen poros especiales que secretan mucilago. La superficie de una
valva puede tener extensiones, llamadas procesos, cuya principal función parece ser
mantener el contacto entre células contiguas y asistir en la formación de colonias. La
frústula se compone de cuarcita, que también puede contener pequeñas cantidades de
aluminio, magnesio, hierro o titanio. El componente inorgánico de la frústula está cubierto
por un componente orgánico o “piel”, compuesto de aminoácidos y azúcares.
II.1.1.2.2 División Celular y la Formación de una Nueva
Pared Celular
El método asexual normal de reproducción es cuando una célula se divide en dos, las
valvas de la célula parental se convierten en las epitecas de las células hijas, estas
produciendo una nueva hipoteca. A través de las divisiones celulares el tamaño de las
células hijas va disminuyendo, así como la proporción de ancho a altura y la morfología de
la célula. Las diatomeas requieren silicio de manera definitiva, como ácido silícico
(Si(OH)4) no disociado. La disminución de este en aguas marinas por el afloramiento de
diatomeas limita el subsiguiente crecimiento de las mismas. Previo a la división celular, la
célula se alarga, separando la epiteca de la hipoteca, y el núcleo se divide. Luego que el
protoplasma se ha dividido en dos por la invaginación del plasmalema, los aparatos de
golgi producen unas vesículas traslúcidas que se reúnen bajo el plasmalema. Estas vesículas
se fusionan para formar la silicalema o membrana de la vesícula de deposición de sílice. La
vesícula gradualmente se expande y asume la forma de una nueva valva. La vesícula de
deposición de sílice determina la forma de la frústula silicificada (Lee, 2008). Luego de
formarse la epiteca e hipoteca, las vesículas de Golgi se fusionan para formar el silicalema
de la banda de la faja. Esta banda es depositada afuera de la célula en el área entre la
hipovalva y la epivalva cuando la deposición se ha completado (Dawson, 1973).

30
Las diatomeas producen cinco tipos de agregaciones mucilaginosas: tubos, almohadillas,
tallos, fibrillas y películas adherentes. Una alta cantidad del carbono fijado es utilizado en
la formación de mucílagos extracelulares (de Brouwer et al, 2002).
II.1.1.2.3 Mobilidad
Algunas diatomeas se pueden deslizar sobre la superficie de un sustrato, dejando una huella
de mucílago. Esta característica es restringida a las diatomeas penadas con rafe y a las
diatomeas céntricas con procesos labiados. Este movimiento se caracteriza por cambios
bruscos de velocidad. Las diatomeas que se adhieren a un sustrato y son móviles sobre el
sustrato tienen las ventajas de poder mantenerse fijas en agua en movimiento, evitar ser
enterradas al poder moverse hacia arriba en los sedimentos, movilizarse para colonizar
áreas vacías y, moverse a áreas con más luz y/o nutrientes (Medlin et al., 1986).
II.1.1.2.4 Productos de Almacenamiento
El producto de almacenaje en las diatomeas es la crisolaminarina, que se encuentra en las
vesículas de la célula. Es similar a la laminarina de las Phaeophyceae (Granum et al, 2001).
Contienen 4α-metil esteroles, como el 4-desmetilesterol y colesterol, que son útiles como
indicadores de diatomeas en la columna de agua y los sedimentos orgánicos (Mansour et al,
1999).
II.1.1.2.5 Ecología
Las diatomeas forman el principal componente de la flora marina en aguas abiertas, y una
parte significativa de la flora de agua dulce. Las diatomeas adheridas se encuentran como
superficies ligosas cafés en varios tipos de sustratos, así como crecimientos acolchonados
cafés debidos al crecimiento abundante de diatomeas epífitas. Las especies penadas se
encuentran en números similares en el mar y en agua dulce, mientras que las céntricas y
gonoides son predominantemente marinas. En términos generales, mientras más frío es el
ambiente marino, mayor es la población de diatomeas. Las poblaciones en las costas son
mayores que en mar abierto, aunque en relación a la diversidad de especies sea lo contrario.
Las aguas costeras reciben mayor cantidad de nutrientes de los bordes del continente (Lee,
2008). Para mantenerse en la columna de agua las diatomeas oceánicas poseen una vacuola
que, por medio de un mecanismo iónico, mantiene una densidad ligeramente menor que la
del agua salada. La flotabilidad también puede ser atribuida a una mayor área en proporción
al volumen; para casos como Chaetoceros, que produce largas setas; Coscinodiscus, con
células discoides; Fragilaria, formando listones; o Rhizosolenia, con formas alargadas.
Variaciones diarias de la flotabilidad permiten a las células subir y bajar en la columna de
agua, aprovechando nutrientes a diferentes profundidades.

31
II.1.1.2.6 Clasificación
Estas parecen haberse originado hace unos 200 millones de años en el Pérmico Tardío a
partir de un miembro escamoso de las Chrysophyceae o Bolidophyceae. Dos escamas
evolucionaron en valvas, mientras que otras escamas se transformaron en las bandas de la
faja. Las diatomeas fósiles más antiguas son céntricas y se encuentran en el registro fósil
del Cretácico Temprano. Las diatomeas penadas se registran hasta el Cretácico Tardío. Las
primeras diatomeas penadas no poseían un rafe, el cual apareció hasta el Eoceno medio.
Las Bacillariophyceae se pueden dividir en dos órdenes (Lee, 2008):
- Biddulphiales: Ornamentación radial o gonoide, muchos cloroplastos, sin rafe, forma
esporas de latencia, espermatozoides móviles con un flagelo, reproducción sexual
oogámica.
- Bacillariales: Ornamentación penada o trelisoide, uno o dos cloroplastos, rafes presentes,
sin espermatozoides flagelados, reproducción sexual por conjugación.
II.1.1.3 Phylum Chlorophyta (Algas verdes).
Las Chlorophyceae, o algas verdes, poseen clorofilas a y b, y forman almidón con el
cloroplasto, usualmente en asociación con un pirenoide. Las clorofíceas difieren del resto
de las algas eucariotas en que forman sustancias de reserva en el cloroplasto, en vez de
dentro del citoplasma. No existe un retículo endoplasmático alrededor de los cloroplastos
(Lee, 2008). Las algas verdes son principalmente de agua dulce, solamente un 10% siendo
marinas (Smith, 1995). Los órdenes Caulerpales, Dasycladales y Siphonocladales son
predominantemente marinos. Ulotrichales y Coleochaetales predominan en aguas dulces.
Oedogoniales y Zygnematales son exclusivamente de agua dulce. Las regiones marinas
tropicales y subtropicales del mundo tienden a compartir especies, mientras que en aguas
frías, entre los polos, hay una grande diferencia entre las especies. Las aguas más cálidas
cercanas al ecuador han servido como una barrera geográfica para la evolución de nuevas
especies y géneros. Las Chlorophyceae son las segundas más abundantes, especialmente en
aguas continentales. Estas pueden ser unicelulares o coloniales. La mayor cantidad de
reservas es el almidón y algunos aceites.
II.1.1.3.1 Estructura Celular
En las Chlorophyta no se forman filamentos microtubulares en los flagelos. En algunos
géneros el aparato de Golgi produce escamas o cerdas fibrilares. Las paredes celulares están
compuestas principalmente de celulosa, aunque los xilanos o mananos sustituyen a la
celulosa en las Caulerpales (Huizing et al, 1979). Las algas primitivas en las

32
Prasinophyceae poseen escamas extracelulares, o una pared derivada de escamas
intercaladas, compuesta de polisacáridos acídicos (Becker et al, 1996). En las Volvocales
las paredes están compuestas de glicoproteínas (Goodenough et al., 1985). Los pigmentos
cloroplásticos son similares a los de las plantas superiores. Utilizan clorofila a y b. El
carotenoide principal es la luteína. Los géneros sifonáceos, así como los unicelulares
Tetraselmis y Mesostigma, son las únicas algas verdes que contienen sifonoxantina y
sifoneína (Yoshi et al., 2003). Los carotenoides se acumulan en condiciones de deficiencias
de nitrógeno, alta radiación solar o alta salinidad. Tal es caso de Dunaliella, la cual acumula
β-caroteno entre los tilacoides, y Haematococcus, que acumula astaxantina en glóbulos
lipídicos afuera del cloroplasto (Lee, 2008). Los mecanismos metabólicos por los cuales
producen el almidón son muy similares a los que utilizan las plantas superiores. Algunos
grupos poseen vacuolas contráctiles, que tienen la función de bombear exceso de agua al
medio y evacuar desechos.
II.1.1.3.2 Fototaxismo y manchas oculares
Existen dos tipos de fototaxismo en las clorofíceas: movimiento por flagelos o por
secreción de mucílago. La mayoría de células flageladas que muestran fototaxismo poseen
una mancha ocular o estigma. Este siempre se encuentra en el cloroplasto, y consiste en una
o varias capas de diminutas gotas de lípidos, de color rojo-naranja a causa de los
carotenoides. La respuesta fototáxica varía con la intensidad de la luz. Se ha observado que
organismos que presentan fototaxismo positivo a moderadas intensidades de luz exhiben un
fototaxismo negativo a intensidades muy altas de luz. La temperatura también afecta la
respuesta fototáxica (Lee, 2008).
II.1.1.3.3 Reproducción Asexual
Hay varios tipos de reproducción asexual, el más simple es la fragmentación de colonias en
dos o más partes, cada una convirtiéndose en una nueva colonia. La zoosporogénesis ocurre
comúnmente, inducida por cambios en el ambiente del alga. Las aplanosporas no poseen
flagelos y tienen una pared adicional a la pared celular original. Las autosporas son
aplanosporas con la misma forma que la célula parental, se producen usualmente en
múltiplos de dos en la célula madre. Un cenobio es una colonia con un número definido de
células ordenado de manera específica. Al madurar un cenobio ocurre un agrandamiento
pero sin más división celular (Lee, 2008).
II.1.1.3.4 Reproducción Sexual
Esta puede ser isogámica, anisogámica u oogámica, con la línea general de evolución en la
misma dirección. Los gametos suelen ser células especializadas y no vegetativas. Si la
especie es isogámica o anisogámica, los gametos no se suelen formar en células

33
especializadas. En especies oogámicas, los gametos se forman normalmente en
gametangios especializados. La mayoría de las Chlorophyta forman gametos móviles
flagelados (zoogametos). En las Zygnematales (Chlorophyta) se forman aplanogametos o
gametos ameboides. En algunas Chlorophyta la gametogénesis es inducida por cambios
ambientales, mientras que en otras, la presencia de dos cepas sexualmente diferentes es
necesaria (Lee, 2008).
II.1.1.3.5 Clasificación
Las clasificación de las cuatro clases importantes de Chlorophyceae fue realizada en base a
características ultraestructurales. Esta clasificación fue corroborada posteriormente
utilizando genética molecular (Lee, 2008). A continuación sus características:
- Prasinophyceae: Flagelados escamosos o desnudos con husos interzonales que persisten
durante la citokinesis; algas verdes primitivas; algunas de las cuales originaron a las otras
clases de Chlorophyta.
- Charophyceae: Células móviles asimétricas; dos flagelos adheridos a la célula en
posición lateral; raíz flagelar consiste en una banda ancha de microtúbulos y una segunda
raíz microtubular más pequeña; puede haber una estructura de capas múltiples; no hay
rizoplasto; escamas son comunes fuera de las células móviles; el fragmoplasto produce
nuevas paredes después de la división; suele carecer de manchas oculares; predominantes
en agua dulce; produce un zigoto en dormancia tras la reproducción sexual; la meiosis
procede a la germinación del cigoto.
- Ulvophyceae: Flagelos adheridos al extremo anterior de la célula; células móviles con
simetría externa casi radial; en ocasiones hay un rizoplasto; no hay una estructura de capas
múltiples; escamas pueden estar presentes en las células móviles; la pared celular se
produce por un pliegue durante la división celular; son comunes las manchas oculares;
predominantemente marinas; es común la alternancia de generaciones.
- Chlorophyceae: Células móviles con simetría externa casi radial; flagelos adheridos al
extremo anterior de la célula; a veces hay rizoplasto; no hay una estructura de capas
múltiples; la teca es común en células móviles; el huso interzonal colapsa en la telofase; el
ficoplasto produce una nueva pared celular en la división celular; manchas oculares son
comunes; predominan en agua dulce; el cigoto atraviesa un período de dormancia; la
meiosis ocurre cuando el cigoto germina.

34
II.1.1.4 Clase Cyanophyceae (Algas verde-azules)
Muy cercanas a las bacterias en estructura y organización, estas algas juegan un papel
importante en la fijación del nitrógeno en la atmósfera. Las cianofíceas o algas verde-azules
con también llamadas cianobacterias. Este nombre hace alusión a la mayor relación con las
bacterias procariotas que con las algas eucariotas. Se cree que estas evolucionaron hace
unos 2.7 billones de años. Las cianobacterias contienen clorofila a (a veces también
clorofila b o d), ficobiliproteínas, glicógeno como sustancia de reserva, y paredes celulares
que contienen aminoazúcares y aminoácidos (Lee, 2008).
II.1.1.4.1 Morfología
La morfología más simple entre las cianobacterias es la unicelular de vida libre, o envuelta
en una vaina mucilaginosa. La evolución subsecuente resultó en la formación de una fila de
células llamada tricoma. Cuando el tricoma está rodeado por una vaina, se le llama
filamento. Es posible tener más de un tricoma en un filamento. El talo más complejo es el
filamento ramificado. Este puede ser uniseriado (una sola fila de células) o multiseriado
(Lee, 2008).
II.1.1.4.2 Pared Celular y Deslizamiento
La pared celular de una cianobacteria es básicamente igual a la de una bacteria Gram-
negativa. Se encuentra una capa de peptidoglucano afuera de la membrana celular. Este es
un polímero enorme compuesto de dos derivados de un azúcar, N-acetilglucosamina y
ácido N-acetilmurámico, y algunos aminoácidos. Afuera del peptidoglucano se encuentra
un espacio periplásmico, probablemente ocupado por una red laxa de fibras de
peptidoglucano. Una membrana externa rodea al espacio periplásmico. Algunas
cianobacterias pueden deslizarse sobre una superficie sólida aún cuando no existe un
órgano visible responsable para el movimiento ni un cambio distintivo de la forma del
organismo. Es un movimiento lento paralelo al eje de la célula y es ocasionalmente
interrumpido por un movimiento en dirección opuesta. El deslizamiento es acompañado por
la producción constante de mucílago, que es dejado como una huella. Algunas
cianobacterias pueden rotar cuando se deslizan. Otras especies utilizan otro mecanismo de
movimiento llamado “twitching” (retorcijón), producido por pili que se proyectan sobre la
superficie de las células (Lee, 2008).
II.1.1.4.3 Vainas
Es común encontrar vainas compuestas de mucílago y una pequeña proporción de celulosa
en las cianobacterias. Esta vaina protege a la célula de la desecación. Las vainas suelen
tener coloración. Son rojas en algas de suelos altamente ácidos, y azules en suelos básicos.

35
En hábitats de alta salinidad, las vainas son amarillas y cafés, especialmente cuando las
algas se secan. La escasez de CO2 reduce la producción de la vaina, mientras que el exceso
de carbono fijado induce a la formación de esta (Otero et al., 2004).
II.1.1.4.4 Pigmentos y fotosíntesis
Los principales componentes del sistema fotosintético de las cianobacterias son la clorofila
a en la membrana tilacoidal, y las ficobiliproteínas, que son cromoproteínas solubles en
agua estructuradas en agregados macromoleculares (ficobilisomas) adheridos a la superficie
externa de las membranas tilacoidales. Poseen carotenoides como la equineona, la
mixoxantofila, y altas proporciones de β-caroteno (Goodwin, 1974). Los tilacoides parecen
haberse originado a partir de invaginaciones del plasmalema. Muchas cianobacterias
realizan fotosíntesis bajo condiciones aeróbicas y anaeróbicas. En aerobiosis, los electrones
para el fotosistema I se derivan del fotosistema II. Bajo anaerobiosis, en presencia de azufre
(en forma de H2S), los electrones se derivan de la reducción del azufre. Estas
cianobacterias son anaerobios fototróficos facultativos, y ocupan un nicho importante en
ecosistemas acuáticos (Padan, 1979). La fotosíntesis en muchas cianobacterias es
estimulada por una disminución de la concentración de oxígeno, el oxígeno compitiendo
con el dióxido de carbono por la enzima ribulosa-1, 5-bisfosfato carboxilasa/oxigenasa
(Stewart et al., 1970). Este fenómeno parece reflejar una adaptación a la ausencia de
oxígeno libre en la atmósfera del Precámbrico, cuando las cianobacterias aparecieron. La
actividad de las cianobacterias liberó oxígeno a la atmósfera, creando una capa protectora
de ozono. Este evento permitió la evolución de organismos más sensibles a la radiación.
II.1.1.4.5 Reproducción Asexual
La reproducción asexual se lleva a cabo con la formación de hormogonia, baeocitos o
fragmentación de colonias. Los hormogonios son característicos de las verdaderas
cianobacterias filamentosas. Son pequeños segmentos de tricoma que se desprenden del
filamento parental para alejarse por deslizamiento, eventualmente desarrollándose en un
filamento separado. Se distinguen del filamento vegetativo por su movilidad, el menor
tamaño de sus células y la ausencia de heterocistos (Meeks et al., 2002). El factor que inicia
la diferenciación en hormogonia es algún cambio en los parámetros ambientales, tal como
la disminución o aumento de algún nutriente o la variación en la cantidad de luz. Los
beocitos son endosporas que son formadas por algunas cianobacterias cocoides. El
protoplasma se divide varias veces en distintos planos sin algún crecimiento entre las
divisiones. Los beocitos resultan más pequeños que la célula original. Son similares a las
endosporas bacterianas (Lee, 2008).

36
II.1.1.4.6 Crecimiento y Metabolismo
Existen tres tipos nutricionales entre las cianobacterias; los quimioheterótrofos facultativos,
que pueden crecer en la oscuridad usando una fuente orgánica de carbono, y en la luz;
fotótrofos obligados, que solo crecen en la luz en un medio inorgánico; y fotoheterótrofos,
que pueden utilizar compuestos orgánicos como fuente de carbono solamente en la luz
(Stanier, 1973).
II.1.1.4.7 Ecología
En ambientes marinos, las cianobacterias de la zona litoral se encuentran como una película
negra incrustada sobre las rocas en el límite superior de la marca de la marea alta. La
mayoría de las cianobacterias en la zona litoral fijan nitrógeno, lo que contribuye a la
productividad de las orillas rocosas y los arrecifes de coral (Mague et al., 1975). A mar
abierto se encuentran principalmente diminutas cianobacterias cocoides como
Synechococcus. Por su pequeño tamaño pueden absorber nutrientes con mayor facilidad
que algas de mayor tamaño, lo que les hace posible crecer en aguas pobres en nutrientes.
También hay especies de cianobacterias filamentosas, como Trichodesmium, que crecen en
mar abierto, formando grandes masas flotantes (Lee, 2008).
II.1.1.4.8 Clasificación
La clase Cyanophyceae es clasificada en tres órdenes (Lee, 2008):
- Chroococcales: Células individuales o laxamente cohesionadas en colonias gelatinosas
irregulares. Incluye los géneros Microcystis, Synechococcus y Synechocystis.
- Oscillatoriales: Cianobacterias filamentosas. Incluye los géneros Oscillatoria,
Trichodesmium, Phormidium, Lyngbya, Hydrocoleus y Spirulina.
- Nostocales: Cianobacterias filamentosas con heterocistos. Incluye los géneros Nostoc,
Anabaena, Aulosira, Calothrix y Rivularia.
II.1.1.5 Phylum Dynophyta
Estos son organismos importantes del plancton en aguas marinas y continentales, aunque
son mucho más diversos en hábitats marinos. En general, son menos importantes en aguas
polares frías que en aguas más cálidas. El grupo de los Dinophysales es un grupo tropical.
Un dinoflagelado móvil típico consiste de un epicono y un hipocono dividido por una faja
transversa o cíngulo. El epicono e hipocono están normalmente divididos en un número de
placas tecales, cuyo número exacto y disposición son característicos de un género en

37
particular. Existe un sulcus longitudinal que se extiende perpendicular a la faja. Los
flagelos longitudinales y transversales emergen a través de las placas tecales en el área en
donde se une la faja y el sulcus. El flagelo longitudinal se proyecta afuera de la célula,
mientras que el flagelo transverso es ondulado y se encuentra cercanamente unido a la faja.
Las células pueden ser fotosintéticas o incoloras y heterotróficas. Los organismos
fotosintéticos tienen cloroplastos rodeados por una membrana de retículo endoplasmático
cloroplástico, que no es continuo con la membrana externa y el envoltorio del núcleo. Las
clorofilas a y c2 se encuentran en los cloroplastos, siendo la peridinina y neoperidinina los
principales carotenoides. La sustancia de reserva es el almidón, similar al de las plantas
superiores, y se encuentra en el citoplasma. Puede existir una mancha ocular. El núcleo
tiene sus cromosomas permanentemente condensadas, a lo que se le llama núcleo
dinocariótico o mesocariótico (Lee, 2008).
Algunas especies de dinoflagelados son responsables por intoxicaciones y muerte en peces
y en humanos. También pueden producir luz por un proceso llamado bioluminescencia,
causado por la reacción enzimática de la luciferasa sobre la luciferina (Lee, 2008).
II.1.1.5.1 Clasificación
Existe una única clase dentro del phylum Dinophyta, la clase Dinophyceae. Se consideran
cuatro órdenes (Lee, 2008):
- Prorocentrales: Pared celular dividida verticalmente en dos mitades; no hay faja; dos
flagelos originados en el ápice celular.
- Dinophysiales: Pared celular dividida verticalmente en dos mitades; células con
extensiones elaboradas de la teca.
- Peridiniales: Células móviles con un epicone e hipocono separados por una faja; teca
relativamente gruesa.
- Gymnodiniales: Células móviles con un epicono e hipocono separado por una faja; teca
delgada o reducida a vesículas vacías.
II.1.1.6 Clase Chrysophyceae (del Phylum Heterokontophyta)
Las Chrysophyceae se distinguen químicamente por poseer clorofilas a, c1, y c2 y
estructuralmente por dos flagelos insertados a la célula perpendiculares uno con el otro, un
fotoreceptor en el flagelo corto y una mancha ocular en la porción anterior del cloroplasto,
vacuolas contráctiles en la porción anterior de la célula, retículo endoplasmático
cloroplástico, y escamas silicificadas radialmente o biradialmente simétricas. La

38
crisolaminarina es la sustancia de almacenamiento. Muchos miembros de la clase producen
estatosporas cubiertas por una pared silicificada con un poro terminal. La mayoría de
especies en las Chrysophyceae son de agua dulce, encontrándose en aguas bajas en calcio
(Lee, 2008).
Este grupo de algas es muy similar a las diatomeas. Contienen un sistema complejo de
pigmentación. Estas existen primordialmente en aguas continentales. Estas producen
aceites naturales y carbohidratos como sustancias de reserva.
Las algas microscópicas son las formas más primitivas de plantas. Durante el procesos de
fotosíntesis en microalgas es similar al de las plantas superiores, estas son más eficientes
transformadoras de energía solar debido a su estructura celular sencilla, lo cual, sumado a
tener mayor acceso a CO2 y nutrientes por estar suspendidas en el agua. De esta manera
las microalgas son capaces de producir 30 veces más la cantidad de aceites por unidad de
área de tierra comparada con cosechas de semillas oleaginosas terrestres (Sheehan et al,
1998).
II.1.1.7 Estudios de Algas marinas en Guatemala
Los estudios actuales se han enfocado al estudio de la especie de Hydrilla verticillata,
calidad del agua en diferentes puntos como los conducidos por la Autoridad para el Manejo
Sostenible de la Cuenca del Lago de Izabal-AMASURLI, proyectos de investigación del
Departamento de Análisis Inorgánico de la Universidad de San Carlos y varias tesis de
grado las cuales se han enfocado en los estudios de peces en el área (Dix et al, 1999; Bol,
2002 y Herrera, 1999). Los estudios marinos para las costas guatemaltecas se han
enfocado a estudios de pesquerías, ecología marina (moluscos, arrecifes coralinos, pastos
marinos y mamíferos marinos (Cazali, 1998; Arrivillaga et al, 2004; Fonseca et al, 2003;
Prado, 1990 y Torrentera et al 1989). Un estudio en el Océano Atlántico guatemalteco
reportó la presencia de 20 especies pertenecientes a 15 géneros de algas verdes
(Chlorophyta) (Cuevas, 1984).
El Centro de Estudios del Mar y Acuicultura (CEMA) de la Universidad de San Carlos de
Guatemala ha establecido un laboratorio para el aislamiento y cultivo de microalgas. El
enfoque básico de este laboratorio es el cultivo de especies con aplicaciones en la
acuicultura, como la producción de alimento vivo (fitoplancton y zooplancton) para la
crianza de peces, moluscos y crustáceos. Se han realizado estudios sobre diversidad de
microalgas marinas en las costas los océanos Pacífico y Atlántico, comportamiento de la
marea roja (afloramiento de dinoflagelados), entre otros, pero no se ha realizado
investigación en relación a el uso de la biomasa de microalgas para la producción de
biocombustibles (Carrillo, com. Pers.) La investigación en esta rama se encuentra en una

39
fase de iniciación en el país, y aún requiere ser evaluada biológica, económica, ecológica y
socialmente previo a ser implementada una tecnología de producción en masa.
II.1.2 Cultivos de microalgas
II.1.2.1 Cultivos de Enriquecimiento
Los cultivos de enriquecimiento han sido usados desde hace mucho tiempo como un paso
preliminar para el aislamiento de células individuales de microalgas. Estos se establecen al
agregar nutrientes a una muestra natural, que la enriquecen para que ocurra el crecimiento
de microalgas. Las sustancias comunes de enriquecimiento incluyen medios de cultivo,
extracto de suelo, o macronutrientes (nitrato, fosfato), pero también se pueden usar
micronutrientes (metales traza). El extracto de suelo es uno de los más simples y exitosos,
siempre y cuando la tierra sea de buena calidad. En algunos casos se agregan sustancias
orgánicas para aislar algunos tipos especiales de microalgas, pero la cantidad debe ser baja,
pues estas sustancias pueden promover el crecimiento bacteriano. El excesivo crecimiento
de bacterias puede volver el medio anóxico o tóxico, aniquilando a las algas (Andersen et
al, 2005).
Las muestras naturales suelen ser deficientes en uno o más nutrientes, pero las algas
sobreviven en la naturaleza, porque la actividad bacteriana, el forrajeo y la muerte de
organismos reciclan los nutrientes. Cuando ya se ha colectado una muestra, este reciclaje se
reduce o se altera, lo que puede causar la muerte a la especie buscada. Un leve
enriquecimiento puede ser útil cuando se realizan largas expediciones marinas, siempre que
se incuben en cámaras de cultivo dentro del barco. Los cultivos se revisan periódicamente
para examinar si la especie deseada se encuentra en crecimiento, haciéndose abundante. Si
es así, se podrán aislar células con mayor facilidad. Cuando las especies deseadas
responden favorablemente al enriquecimiento se facilita el aislamiento y se aumenta el
éxito (Andersen et al, 2005).
Generalmente se usa el nitrato, fosfato o un extracto de suelo como la sustancia de
enriquecimiento. Sin embargo, esta puede ser una mezcla de varias, o incluso un elemento
traza. Lo que suele resultar es que un organismo diferente domina en cada uno de los
enriquecimientos realizados. Algunos organismos que no se observan en la muestra inicial
aparecen en unos 7 días después del enriquecimiento (Andersen et al, 2005).

40
II.1.2.2 Dinámica del crecimiento de microalgas
El crecimiento de un cultivo axénico de microalgas se caracteriza por cinco fases:
1. Fase de inducción: en esta fase hay un lento aumento en la densidad celular, toma
bastante tiempo cuando se ha transferido un cultivo desde una caja de Petri a un medio
líquido. Los cultivos realizados con algas en crecimiento exponencial presentan una fase de
inducción corta. Este aletargamiento en el crecimiento es atribuido a la adaptación
fisiológica del metabolismo celular al crecimiento, tal como el aumento en los niveles de
enzimas y metabolitos que intervienen en la división celular y fijación de carbono (Lavens
et al, 1996).
2. Fase exponencial: en esta fase, la densidad celular aumenta como una función del tiempo
(t) de acuerdo a la función logarítmica:
Ct=C0emt
,
donde Ct y C0 son las concentraciones celulares en el tiempo t y 0, respectivamente, y m es
la tasa específica de crecimiento, la cual depende de la especie, intensidad lumínica y
temperatura (Lavens et al, 1996).
3. Fase de declinación del crecimiento: esta se alcanza cuando los nutrientes, iluminación,
pH, dióxido de carbono u otros factores físicos empiezan a limitar en crecimiento (Lavens
et al, 1996).
4. Fase estacionaria: aquí se equilibra la tasa de crecimiento con los factores limitantes,
resultando en una densidad celular relativamente constante (Lavens et al, 1996).
5. Fase de muerte: la calidad del agua se deteriora y los nutrientes se agotan, no pudiendo
sustentar el crecimiento. Disminuye rápidamente la densidad celular, llevando al colapso
del cultivo (Lavens et al, 1996).
II.1.3 Biodiesel a partir de Aceites en microalgas
II.1.3.1 Aceites vegetales
En las plantas se pueden hallar dos tipos de aceites, los esenciales y los fijos, que se
diferencian por su función dentro de la planta, sus propiedades fisicoquímicas y por la
cantidad en la que se encuentran dentro de la misma.

41
El contenido de ácido graso (%), de los aceites mayormente conocido es el siguiente:
Mirístico (C14:0) 1.1; Palmítico (C16:0) 42.9; Esteárico (C18:0) 4.6; Oleico (C18:1) 39.3;
Linoléico (C18:2) 10.7 (Romero, 2005).
II.1.3.2 Biocombustibles y Biodiesel
Son biocombustibles aquellos productos químicos orgánicos obtenidos del procesamiento
de biomasa y que son utilizados como combustibles. En algunos casos, como la leña, la
biomasa es en si misma el combustible. El término biomasa se refiere a toda la vegetación
del planeta y a muchos productos y subproductos que provienen de ella, como por ejemplo
aceites, residuos agrícolas o algunos desechos municipales e industriales y es la fuente más
antigua de energía renovable conocida. Los biocombustibles poseen grandes ventajas
respecto a los combustibles derivados del petróleo, entre ellas se pueden mencionar:
1. Proporcionan una fuente de energía renovable y, por lo tanto, inagotable.
2. Disminuyen en forma notable las emisiones de CO2, de los hidrocarburos no quemados y
de materia sólida y elimina los sulfatos por completo.
3. Revitaliza la economía rural, generando empleo y valor agregado a las cosechas.
El biodiesel es una mezcla de esteres que pueden producirse a partir de diferentes tipos de
aceites vegetales, grasas animales o aceites residuales de la industria de los alimentos,
haciéndolos reaccionar con metanol u otro alcohol de baja masa molecular, obteniendo
glicerina como producto secundario (Weiss, 1983). Las dos reacciones mediante las cuales
se puede obtener biodiesel son la de transesterificación y la de esterificación.
II.1.3.3 Microalgas como productores de aceites
Los aceites de semillas vegetales actualmente proveen una pequeña fracción de la demanda
de combustibles líquidos por parte del sistema de transporte. La fotosíntesis microbiana ha
sido poco explotada hasta el momento, y recibido poca atención. El potencial de las
microalgas para producir biocombustibles ya es ampliamente reconocida. Las microalgas
poseen ciertas particularidades ventajosas sobre las plantas superiores para la producción de
biocombustibles: a) la productividad de aceites por unidad de área puede exceder
grandemente a los mejores cultivos agrícolas; b) necesitan menos agua que los cultivos
terrestres; c) las microalgas pueden cultivarse en agua marina o salobre en terrenos no
cultivables y no compiten por los recursos con la agricultura convencional; d) la producción
de biomasa de microalgas puede combinarse con la fijación de dióxido de carbono de
desecho; e) los fertilizantes para el cultivo de microalgas pueden obtenerse de aguas
residuales; f) el cultivo de algas no necesita pesticidas o herbicidas; g) el residuo de la
biomasa luego de la extracción del aceite puede utilizarse como alimento para animales,
fertilizante, o fermentado para producir etanol o metano; h) la composición bioquímica de

42
la biomasa puede modificarse al variar las condiciones del crecimiento, aumentando así la
producción de aceites (Rodolfi, L.2008).
Sin embargo también existen limitaciones significativas relacionadas con la tecnología para
su cultivo. Entre estas: a) es necesario escoger y cultivar cepas altamente productivas; b) la
dificultad de mantener las especies seleccionadas en cultivos al exterior; c) hay pocas
plantas comerciales en operación y limitada disponibilidad de datos en cultivos a gran
escala; d) la necesidad de altas cantidades de energía necesarias para bombear el agua,
transferir el dióxido de carbono, mezclar la suspensión del cultivo, cosechar y secar la
biomasa algal. A pesar de esto se ve necesario realizar el esfuerzo para sobrepasar estas
limitaciones, tomando en cuenta que la transición de combustibles fósiles a combustibles
renovables es necesaria e inevitable (Rodolfi, L.2008).
Si se reemplazara todo el combustible utilizado para transporte que se consume en Estados
Unidos con biodiesel, se requeriría 0.53 billones de m3 de biodiesel anualmente bajo la
actual tasa de consumo. Los cultivos para aceite, el aceite de desecho de cocina y la grasa
animal no podrían satisfacer esta demanda. Solamente para cumplir con la mitad de los
requerimientos en combustible diesel para transporte se necesitarían áreas demasiado
grandes de cultivos oleaginosos, siendo esto ecológicamente no sostenible. Es claro que los
cultivos oleaginosos no pueden contribuir de manera significativa para reemplazar
combustibles líquidos derivados del petróleo en el futuro próximo. Esta situación cambia
dramáticamente cuando se evalúa el potencial de las microalgas para producir biodiesel.
Entre el 1% y el 3% del total de áreas cultivables en los Estados Unidos sería suficiente
para producir la biomasa de microalgas para satisfacer el 50% de la demanda de
combustibles para el transporte (Chisti, 2007).
En base a las investigaciones, las microalgas parecen ser la única fuente de biodiesel que
podría reemplazar al biodiesel fósil. A diferencia de otros cultivos oleaginosos, las
microalgas crecen extremadamente rápido y muchas son altas en contenido de aceite. Las
microalgas comúnmente doblan su biomasa en 24 horas. Durante la fase de crecimiento
exponencial este puede llegar tan rápido como en 3.5 horas. El contenido de aceite puede
exceder el 80% en peso de la biomasa seca (Metting, 1996). Niveles de aceite entre el 20%
y el 50% pueden ser bastante comunes. La productividad por unidad de tiempo depende en
la tasa de crecimiento del alga y el contenido de aceite en el alga. Para la producción de
biodiesel se necesitan especies de microalgas altamente productivas (Chisti, 2007).
Dependiendo de la especie, las microalgas pueden producir diferentes tipos de lípidos,
hidrocarburos y otros aceites complejos. No todos los aceites son apropiados para hacer
biodiesel, pero los aceites útiles son bastante comunes. El uso de microalgas para biodiesel
no compite con la producción de alimento humano, forraje, u otros productos obtenidos de
cultivos agrícolas (Chisti, 2007). También existen microorganismos heterótrofos que

43
producen aceites, con el inconveniente que el carbono que consumen para su desarrollo y
crecimiento proviene de las plantas (Ratledge y Wynn, 2002). Así, la producción de aceites
de algas requiere la habilidad de producir grandes cantidades de biomasa rica en aceite a
bajo costo (Chisti, 2007).
II.1.3.4 Producción de biomasa de Microalgas
La producción de biomasa de microalgas es generalmente más costosa que los cultivos
agrícolas. La fotosíntesis requiere luz, dióxido de carbono, agua y sales inorgánicas. La
temperatura debe oscilar entre los 20 y 30°C. Para reducir los costos, la producción de
biodiesel debe depender en la luz solar disponible, a pesar de las variaciones diarias y
estacionales de los niveles de luz. El medio de cultivo debe proveer los elementos
inorgánicos que constituyen a la célula algal. Los elementos esenciales incluyen nitrógeno,
fósforo, hierro y, en algunos casos, silicio. Los requerimientos nutricionales mínimos
pueden ser estimados utilizando la fórmula molecular aproximada de la biomasa
microalgal, CO0.48H1.83N0.11P0.01. Esta fórmula se basa en los datos presentados por
Grobbelaar (2004). Los nutrientes tal como fósforo deben ser proveídos en exceso
significativo debido a que los fosfatos producen complejos con los iones metálicos, por lo
que no todo el fósforo agregado es biodisponible. El agua de mar suplementada con
fertilizantes comerciales con nitrato y fósforo, y algunos micronutrientes, es un modo
común de cultivar microalgas marinas (Molina Grima et al, 1999). Los medios de cultivo
son generalmente económicos.
La biomasa contiene aproximadamente el 50% de carbono en peso seco. Este carbono se
deriva comúnmente del dióxido de carbono. La producción de 100 toneladas de biomasa
algal fija cerca de 183 toneladas de dióxido de carbono. Este debe ser introducido al medio
de manera continua durante las horas de luz. Si la alimentación de CO2 es controlada en
respuesta a sensores del pH, se minimiza la pérdida de dióxido de carbono y variaciones de
pH. La producción de biodiesel puede ser acoplada a la emisión de dióxido de carbono
producida en plantas de energía eléctrica que queman combustibles fósiles. Este carbono
está disponible sin costo (Sawayama et al., 1995). La producción de microalgas a gran
escala utiliza cultivos continuos durante las horas de luz. Bajo este método de operación, el
medio de cultivo fresco es alimentado a una tasa constante, y la misma cantidad de cultivo
es retirada continuamente (Molina Grima et al., 1999). Tanto como el 25% de la biomasa
producida durante el día puede perderse durante la noche debido a la respiración. El grado
de pérdida depende del nivel de iluminación, la temperatura de crecimiento y la
temperatura nocturna (Chisti, 2007). Los métodos prácticos para la producción a gran
escala son los estanques de carril y los fotobioreactores tubulares (Molina Grima et al.,
1999).

44
II.1.3.5 Estanques de Carril
Un estanque de carril está fabricado por un canal de recirculación continua de unos 30 cm
de profundidad. El mezclado y circulación son producidos por una rueda de paletas. El flujo
es guiado en las curvas por deflectores colocados en el canal. Los canales se construyen de
concreto o tierra compactada, y pueden estar cubiertos de plástico blanco. Durante el día, el
cultivo se alimenta frente a la rueda de paletas, donde el flujo inicia. El caldo es cosechado
detrás de la rueda de paletas, al haber completado la circulación. El flujo se mantiene todo
el tiempo para prevenir la sedimentación (Chisti, 2007). Este tipo de cultivo se ha utilizado
desde los años 50’s.
En los canales el enfriamiento se logra por evaporación. La temperatura fluctúa entre el día
y la noche, y estacionalmente. La pérdida de agua por evaporación puede ser significativa.
Esto, y la menor eficiencia en el aprovechamiento del dióxido de carbono, los hace menos
eficientes que los fotobioreactores. La productividad es afectada por la contaminación por
microalgas no deseadas y microorganismos que se alimentan de las algas. La concentración
de biomasa se mantiene baja porque los canales son mezclados pobremente y no se puede
mantener una zona ópticamente oscura (Terry, 1985). Los estanques de carril se consideran
más económicos que los fotobioreactores por su bajo costo de construcción y operación.
Sin embargo, estos tienen una baja productividad de biomasa en comparación a los
fotobioreactores.
II.1.3.6 Fotobioreactores
A diferencia de los estanques, los fotobioreactores permiten el cultivo de especies puras por
períodos prolongados. Los fotobioreactores han sido exitosamente utilizados para la
producción de grandes cantidades de biomasa de microalgas (Molina Grima et al., 1999).
Un fotobioreactor tubular consiste en una serie de tubos rectos transparentes usualmente
hechos de plástico o vidrio. En estos tubos es conde se recibe la luz solar. Estos tubos son,
generalmente, de 10 cm de diámetro o menores. Se limita este diámetro debido a que la luz
no penetra muy profundamente en el cultivo denso, lo cual es necesario para garantizar una
alta productividad de biomasa en el fotobioreactor. El cultivo de microalgas es circulado
desde un tanque de reserva hacia el colector solar y de regreso al tanque. Este funciona
como un cultivo continuo (Chisti, 2007).
El colector solar es orientado para maximizar la captura de luz. En una disposición típica,
los tubos solares se colocan paralelos uno con el otro y planos sobre el suelo. También se
pueden colocar horizontalmente como un cerco para maximizar el número de tubos que se
pueden acomodar en un área. Los tubos siempre son orientados de norte a sur. El suelo bajo
el colector solar es regularmente pintado de blanco o cubierto con plástico blanco para
aumentar la reflectividad, incrementando la luz recibida por los tubos. En vez de estar sobre
el suelo, los tubos pueden ser flexibles y enrollarse alrededor de un marco de soporte para

45
formar un fotobioreactor tubular espiral (Chisti, 2007). La iluminación artificial puede ser
utilizada, pero es costosa en comparación a la iluminación natural. Sin embargo, en la
producción de algunos productos de alto valor, se ha utilizado a para la producción de
biomasa a gran escala (Pulz, 2001).
La sedimentación en los tubos se previene manteniendo un flujo turbulento alto. El flujo se
produce mediante una bomba mecánica o una bomba de compresión de aire. Las bombas
mecánicas pueden dañar la biomasa pero son fáciles de diseñar, instalar y operar. Las
bombas de compresión de aire se han utilizado con éxito. Estas requieren una constante
provisión de aire para operar. Los fotobioreactores deben ser limpiados y desinfectados
periódicamente, lo cual se logra mediante mecanismos automáticos instalados (Chisti,
1999).
La fotosíntesis genera oxígeno, cuya concentración puede ser bastante alta, e inhibir la
fotosíntesis. Además, una alta concentración del oxígeno disuelto en combinación con
intensa iluminación produce un daño fotooxidativo a las células. Para prevenir la inhibición
y el daño celular el máximo tolerable de oxígeno disuelto no debe exceder el 400% de valor
de saturación del aire. Para lograr esto el cultivo debe regresar periódicamente a una zona
de desgasificación que es burbujeada con aire para eliminar el oxígeno acumulado. El
cultivo debe regresar a los tubos colectores de luz libre de burbujas de gas. Debido a que la
zona de desgasificación está pobremente iluminada su volumen debe mantenerse pequeño
en relación al volumen del colector solar (Chisti, 2007).
A medida que el cultivo se mueve a lo largo del fotobioreactor, el pH aumenta a causa del
consumo de dióxido de carbono. Este es alimentado en la zona de desgasificación en
respuesta a un controlador del pH. También se requieren puntos adicionales de inyección de
dióxido de carbono a intervalos a lo largo de los tubos, para prevenir la limitación de
carbono y el aumento excesivo del pH (Molina Grima et al., 1999).
Los fotobioreactores requieren enfriamiento durante las horas de luz. El control de la
temperatura durante la noche también es beneficioso. La pérdida de biomasa durante la
noche puede reducirse al disminuir la temperatura en la noche. Los fotobioreactores
tubulares de exterior pueden enfriarse efectivamente y de manera poco costosa utilizando
intercambiadores de calor. Estos pueden colocarse en la columna de desgasificación o
dentro de los tubos. En climas calientes se pueden mojar los tubos por fuera para enfriar por
evaporación (Tredici, 1999). En algunos casos se han usado invernaderos con temperatura
controlada, pero esto aumenta demasiado los costos para la producción de biodiesel (Pulz,
2001).

46
II.1.3.7 Comparación entre estanques de carril y fotobioreactores
tubulares
Los fotobioreactores proveen una mayor producción de aceite por hectárea que los
estanques. Esto se debe a que la productividad volumétrica de biomasa en los
fotobioreactores es 13 veces mayor en comparación a los estanques. Ambos métodos de
producción son técnicamente asequibles. Se han utilizado áreas de 5,681 m2 para
fotobioreactores y de 7,828 m2 para estanques con una producción anual de biomasa de
100,000 Kg (Chisti, 2007). La obtención de la biomasa del cultivo es necesaria para la
extracción del aceite. Esta se recupera fácilmente por filtración, centrifugación u otros
métodos (Molina Grima et al., 2003). El costo de este proceso puede ser alto, siendo
bastante menor cuando el cultivo proviene de fotobioreactores que cuando proviene de
estanques. Esto se debe a que los cultivos son más densos, hasta unas 30 veces más que los
cultivos obtenidos de estanques de carril (Chisti, 2007).

47
PARTE III
III.1 RESULTADOS
Tabla 7. Especies encontradas en los cinco puntos de muestreo seleccionados combinadas
con las que se observaron en los 6 tratamientos de cultivos de enriquecimiento para cada
muestra. Las especies no identificadas fueron catalogadas como morfoespecies. Para una
descripción de los sitios de muestreo ver Tabla 1.
Especie Punto
3
Punto
4
Punto
5
Punto
7
Punto
10
Ameboide X
Bacteriastrum furcatum X X X X X
Botryococcus braunii X
Melosira nummuloides X X X X
Ceratium furca X X X
Ceratium fusus X X X X
Ceratium trichoceros X X
Ceratium tripos X X X X
Chaetoceros decipiens X X X X X
Scenedesmus sp. X X X X
Coscinodiscus centralis X X X X X
Diatomea verde X X X
Euglenofícea X X
Fragilaria sp.1 (agujas cortas) X X
Fragilaria sp.2 (agujas largas) X X X X
ME 12 Chlorophyta filamentosa X
Staurastrum sp. X X
Entomoneis sp. X
ME 8 Dinoflagelado X X X
ME 9 X
ME10 Diatomea anclada X X
ME13 Clorofícea (tren) X X
Microcystis sp. X X X
Navicula sp. (dulces) X X X
Navicula sp. (arrocitos) X X X X
Nitzschia longissima X X
Pediastrum duplex X X
Pleurosigma sp. X
Protoperidinium sp. X X X
Rhizosolenia sp. X X X X
Synechococcus sp. X X X X X
Thalassionema nitzschioides X X X
Total = 32 20 20 19 18 14
Fuente: Proyecto Fodecyt 43-2007.

48
Tabla 8. Especies encontradas en todos los muestreos. Estas son las especies observadas
en las muestras de los sitios comparados y los no comparados.
Especies de los puntos comparados Especies de los otros sitios muestreados
Ameboide Amphora sp.
Bacteriastrum furcatum Coscinodiscus centralis
Botryococcus braunii Coscinodiscus granii
Melosira nummuloides Cyclotella sp.
Ceratium furca Nannochloris sp.
Ceratium fusus Odontella aurita
Ceratium trichoceros Odontella longicruris
Ceratium tripos Pseudonitzschia sp.
Chaetoceros decipiens Tabellaria sp.
Scenedesmus sp. Tetraselmis sp.
Coscinodiscus centralis Thalassionema sp.
Diatomea verde Thalassiosira sp.
Euglenofícea
Fragilaria sp.1 (agujas cortas)
Fragilaria sp.2 (agujas largas)
ME 12 Chlorophyta filamentosa
Staurastrum sp.
Entomoneis sp.
ME 8 Dinoflagelado
ME 9
ME10 Diatomea anclada
ME13 Clorofícea (tren)
Microcystis sp.
Navicula sp.1 (arrocitos)
Navicula sp. 2 (dulces)
Nitzschia longissima
Pediastrum duplex
Pleurosigma sp.
Protoperidinium sp.
Rhizosolenia sp.
Synechococcus sp.
Thalassionema nitzschioides
32 especies 12 especies
Total: 44 especies
Fuente: Proyecto Fodecyt 43-2007.
Los puntos de muestreo comparados en relación a su biodiversidad y tratamientos de
cultivo de enriquecimiento corresponden a los sitios 3, 4, 5, 7 y 10, y los no comparados
son los sitios 1, 2, 6, 8 y 12. Las muestras de estos últimos solo fueron analizadas para
conocer su diversidad y realizar algunos aislamientos sin la aplicación de soluciones de
enriquecimiento.
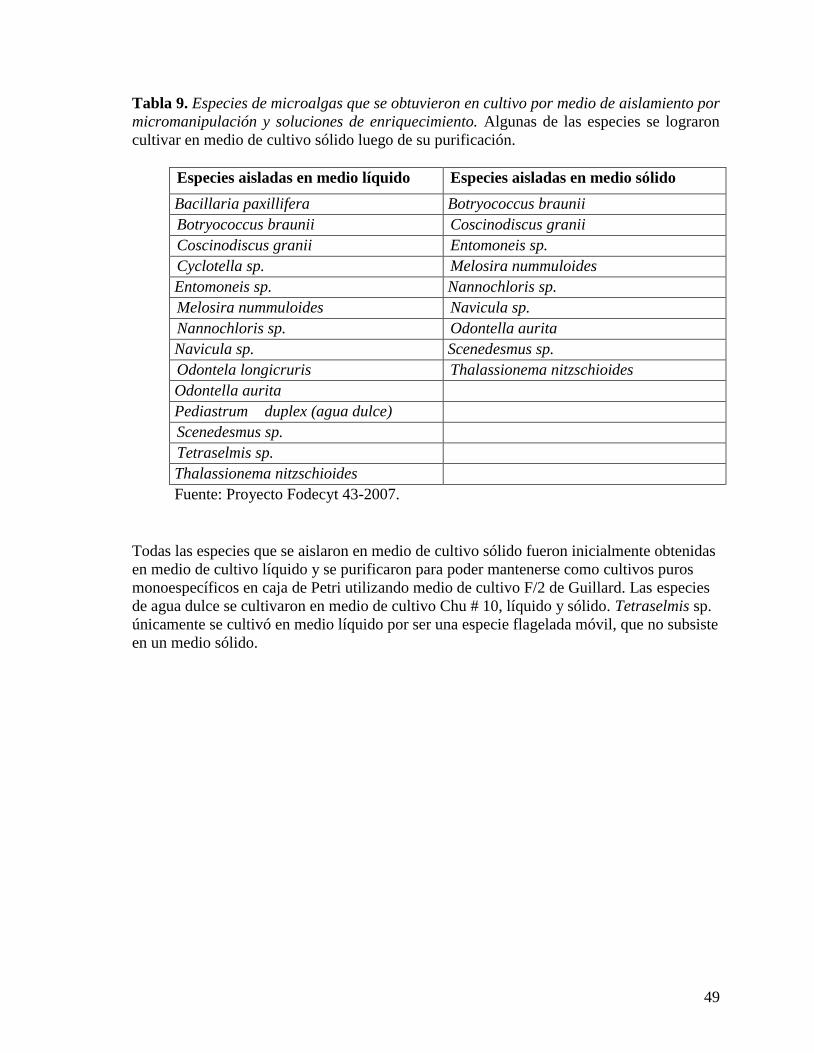
49
Tabla 9. Especies de microalgas que se obtuvieron en cultivo por medio de aislamiento por
micromanipulación y soluciones de enriquecimiento. Algunas de las especies se lograron
cultivar en medio de cultivo sólido luego de su purificación.
Especies aisladas en medio líquido Especies aisladas en medio sólido
Bacillaria paxillifera Botryococcus braunii
Botryococcus braunii Coscinodiscus granii
Coscinodiscus granii Entomoneis sp.
Cyclotella sp. Melosira nummuloides
Entomoneis sp. Nannochloris sp.
Melosira nummuloides Navicula sp.
Nannochloris sp. Odontella aurita
Navicula sp. Scenedesmus sp.
Odontela longicruris Thalassionema nitzschioides
Odontella aurita
Pediastrum duplex (agua dulce)
Scenedesmus sp.
Tetraselmis sp.
Thalassionema nitzschioides
Fuente: Proyecto Fodecyt 43-2007.
Todas las especies que se aislaron en medio de cultivo sólido fueron inicialmente obtenidas
en medio de cultivo líquido y se purificaron para poder mantenerse como cultivos puros
monoespecíficos en caja de Petri utilizando medio de cultivo F/2 de Guillard. Las especies
de agua dulce se cultivaron en medio de cultivo Chu # 10, líquido y sólido. Tetraselmis sp.
únicamente se cultivó en medio líquido por ser una especie flagelada móvil, que no subsiste
en un medio sólido.

50
Tabla No. 10. Especies más abundantes en cada tratamiento. Aquí se nombran las
especies más abundantes en los cultivos de enriquecimiento según su tratamiento. El
numeral representa el punto de muestreo y la literal el tratamiento. Tratamientos: A=F/2 de
Guillard con silicato; B= F/2 de Guillard sin silicato; C=Plymouth-erdschreiber; D=Fe-
EDTA a concentración F/2; E= Nitrato a concentración F/2; F=Fosfato a concentración F/2.
Tratamiento Especie
3A Navicula sp.1 (arrocitos)
3B Scenedesmus sp., Ceratium spp.
3C Navicula sp.1 (arrocitos), Navicula sp.2(dulcitos)
3D Navicula sp.1 (arrocitos)
3E Navicula sp.1 (arrocitos)
3F Synechococcus sp., Navicula sp.1 (arrocitos)
4A Navicula sp.1 (arrocitos)
4B Navicula sp.1 (arrocitos)
4C Melosira nummuloides, Navicula sp.1 (arrocitos)
4D Nitzschia longissima, Navicula sp.1 (arrocitos)
4E Entomoneis sp.
4F ME8 Dinoflagelado
5A Navicula sp.1 (arrocitos)
5B Melosira nummuloides
5C ME5 (Diatomea verde), Synechococcus sp.
5D Navicula sp.1(dulces), Scenedesmus sp.
5E Scenedesmus sp.
5F Navicula sp.1 (arrocitos)
7A Navicula sp.1 (arrocitos), Synechococcus, Fragilaria, Navicula sp.
7B ME10 Diatomea anclada, Navicula sp.2, Synechococcus sp. Fragilaria sp. 2
7C Fragilaria sp.2, Synechococcus sp., Navicula sp.1, Diatomea verde
7D Diatomea verde, Navicula sp.1 (arrocitos), Fragilaria sp. 2
7E Fragilaria sp. 2, Diatomea verde, Navicula sp.1 (arrocitos), Scenedesmus sp.
7F Navicula sp.1 (arrocitos)
10A Navicula sp.1 (arrocitos)
10B Navicula sp.1 (arrocitos)
10C Navicula sp.1 (arrocitos)
10D Navicula sp.1 (arrocitos)
10E Navicula sp.1 (arrocitos), Navicula sp. (dulcitos)
10F Navicula sp.1 (arrocitos)
Fuente: Proyecto Fodecyt 43-2007.

51
Se observa que la especie más común es Navicula sp.1, a excepción de la muestra 7 y de
algunos tratamientos para otras muestras. Estas excepciones fueron útiles para poder aislar
especies que no se pudieron aislar a partir de las muestras originales. Esta especie de
Navicula es altamente generalista, evitando que se desarrollen especies más especialistas,
por lo que es considerada como contaminación.
Figura 2. Análisis de agrupamiento jerárquico con datos de presencia-ausencia de las
especies por cada punto de muestreo, incluyendo las observadas en cultivo.
Fuente: Proyecto Fodecyt 43-2007.
El gráfico muestra la similitud que existe entre cada uno de los puntos de muestreo según
las especies de microalgas encontradas en cada punto, en donde M# corresponde a cada
punto de muestreo y la escala numérica (0 a 25) muestra un grado descendente de similitud
entre cada punto de muestreo. El grado de similitud está dado por el punto de intersección
entre dos líneas.
Este muestra que los sitios de muestreo 3 y 4 tienen la mayor similitud en diversidad de
especies de microalgas, formando un grupo con el sitio 5 de menor similitud. Los sitios 7 y
10 comparten cierta similitud entre sí, aunque menor que los sitios del grupo anterior. El
grupo de los sitios 7 y 10 difiere totalmente del grupo de 3, 4 y 5.

52
Tabla 11. Porcentaje de aceite de las especies cultivadas.
Especies aisladas en medio líquido Porcentaje de aceite presente
Bacillaria paxillifera 0-5%
Botryococcus braunii No determinado
Coscinodiscus granii 0%
Cyclotella sp. 3%
Entomoneis sp. 2%
Melosira nummuloides 12%
Nannochloris sp. 17%
Navicula sp.1 16%
Odontela longicruris 8%
Odontella aurita 11%
Pediastrum duplex (agua dulce) 0%
Scenedesmus sp. 15%
Tetraselmis sp. 5.5%
Thalassionema nitzschioides No determinado
Fuente: Proyecto Fodecyt 43-2007.
Se pueden observar porcentajes de extracción de aceite que se encuentran en el rango del
0% al 17% en biomasa seca. Hay que hacer énfasis en que, para obtener un porcentaje más
preciso, sería apropiado refinar el aceite para eliminar impurezas de naturaleza polar y
apolar que pudieran estar contenidas en los extractos. Las especies que presentaron mayor
porcentaje de aceites fueron Nannochloris sp., Odontella aurita y Navicula sp. y
Scenedesmus sp. Es importante mencionar que la obtención de cultivos puros favorecería a
mantener los porcentajes de rendimiento altos y constantes.

53
III.2 DISCUSIÓN DE RESULTADOS
III.2.1 Diversidad de microalgas en los sitios muestreados
La figura 2 muestra un análisis de agrupamiento generado a partir de los datos de
presencia-ausencia de las especies para los puntos de muestreo 3, 4, 5, 7 y 10. Para este
análisis se incluyeron las especies que se observaron en los cultivos de enriquecimiento
además de las que se encontraron en las muestras originales (Ver Tabla 7). Nos da
evidencia de la diferencia en la diversidad de microalgas entre los puntos de muestreo. Los
puntos más similares fueron los puntos 3 y 4 (localizados en costas opuestas de la entrada a
la Bahía de Puerto Barrios), los que presentaron las siguientes 17 especies en común:
Bacteriastrum furcatum, Melosira nummuloides, Ceratium furca, Ceratium fusus, Ceratium
trichoceros, Ceratium tripos, Chaetoceros decipiens, Scenedesmus sp., Coscinodiscus
centralis, Fragilaria sp.1 (agujas cortas), Euglenofícea, ME 8 Dinoflagelado, Navicula sp.
(arrocitos), Protoperidinium sp., Rhizosolenia sp., Synechococcus sp., Thalassionema
nitzschioides (Ver Tabla 7). El punto 5 se agrupó con estos dos puntos, con 14 especies en
común entre los tres, a saber, Bacteriastrum furcatum, Melosira nummuloides, Ceratium
furca, Ceratium fusus, Ceratium tripos, Chaetoceros decipiens, Scenedesmus sp.,
Coscinodiscus centralis, ME 8 Dinoflagelado, Navicula sp. (arrocitos), Protoperidinium
sp., Rhizosolenia sp., Synechococcus sp., Thalassionema nitzschioides. Estos fueron los
puntos geográficamente más cercanos entre sí. Los puntos 7 y 10 mostraron la menor
similitud con el grupo de 3, 4 y 5, aun manteniendo 11 especies en común entre los dos.
Las especies comunes entre los sitios 7 y 10 fueron las siguientes: Chaetoceros decipiens,
Bacteriastrum furcatum, Coscinodiscus centralis, Fragilaria sp.2 (agujas largas),
Staurastrum sp., ME10 Diatomea anclada, ME13 Clorofícea (tren), Microcystis sp.,
Navicula sp. (dulces), Pediastrum duplex, Synechococcus sp.
Las especies en común entre los dos grupos 3-4-5 y 7-10 fueron: Bacteriastrum furcatum,
Chaetoceros decipiens, Coscinodiscus centralis, Synechococcus sp. El punto 10 resultó ser
el más distinto, lo cual se explica por su localización, la desembocadura del Río Dulce, lo
cual provoca una mezcla de agua dulce con agua salada, o sea una disminución en la
salinidad del agua y por lo tanto predomina en ese punto el desarrollo de algas de agua
dulce las cuales son muy diferentes a las de agua salada, Por esa razón es que este punto no
tiene coincidencia con las especies encontradas en otros puntos que son exclusivamente de
agua salada. En la muestra se encontraron especies marinas y de agua dulce. Es probable
que cerca del punto 7 desemboque algún riachuelo que provea localmente fitoplancton de
agua dulce.
Para algunas de las muestras los diferentes tratamientos si produjeron diferencias en cuanto
a las especies dominantes, aunque para otras no hubo mayor diferencia, como ocurrió con
la muestra 10 (Ver Tabla 10). Cada tratamiento de soluciones de enriquecimiento
presentaba una variante de la fórmula original del medio de cultivo f/2 de Guillard. Ver
Tabla 6 para la denominación de cada tratamiento.
La especie más común en los cultivos fue Navicula sp. 1 (arrocitos), la cual era muy raro
encontrarla en las muestras originales. Pocos de los tratamientos no presentaron esta

54
especie. Para la muestra 3 el tratamiento B (f/2 sin silicato) no presentó Navicula sp.,
dominando Scenedesmus sp. La ausencia del silicato debió haber limitado el crecimiento de
esta diatomea. Sin embargo, esta si se presentó en los otros tres tratamientos sin silicato
adicional [D (hierro), E (nitrato) y F (fosfato)]. Las condiciones del tratamiento 3B,
presencia de todos los nutrientes excepto el silicato, estimularon el crecimiento de
Scenedesmus sp., y esto evitó el desarrollo de diatomeas generalistas como Navicula sp.
Para la muestra 4 los tratamientos E y F fueron los más notables por presentar dos especies
(ME8 Dinoflagelado y Entomoneis sp.) en ausencia de Navicula sp. 1 (arrocitos). Los
tratamientos C y D si presentaron la mencionada especie, acompañados de dos especies de
diatomeas en abundancia (Melosira nummuloides, y Nitzschia longissima). El tratamiento C
(Plymouth-erdschreiber) es un medio de cultivo preparado con extracto de suelo, nitrato y
fosfato, que es utilizado para mantenimiento de cepas, por ser un medio no determinado
(Andersen, 2005). Su aplicación favoreció en el aislamiento de la especie Melosira
nummuloides.
La muestra 5 también presentó diferencia entre los tratamientos, dos de los cuales
presentaban dominancia de una especie (Melosira nummuloides en 5B y Scenedesmus sp.
en 5E) en ausencia de Navicula sp. 1 (arrocitos). Otros dos tratamientos (5D y 5F)
presentaron dominancia de esta especie. La abundancia de Scenedesmus sp. en la muestra
permitió que se encontrara en los tratamientos 5D y 5E.
La muestra 7 presentó una mayor diversidad de especies comunes en alta abundancia en
casi todos los tratamientos, pero siempre en presencia de Navicula sp. 1 (arrocitos). La
cianobacteria Synechococcus sp., la cual se considera un organismo contaminante, fue
común en tres de los tratamientos (7A, 7B y 7C).
En los cultivos de la muestra 10 hubo un bajo efecto de los tratamientos, habiendo
propiciado especialmente esta especie de Navicula. Esto se explica por la baja salinidad del
agua en que se colectó la muestra debido a la mezcla de agua dulce por la desembocadura
de Río Dulce. Aun habiendo microalgas marinas en la muestra estas no se reprodujeron en
cultivo. Las especies de diatomeas Chaetoceros decipiens, Coscinodiscus centralis y
Melosira nummuloides, y de dinoflagelados Ceratium spp. no se propagaron en mayor
cantidad en los cultivos. Los especímenes encontrados en algunos tratamientos podrían ser,
en parte, remanentes de las muestras originales, y no células que se reprodujeron bajo el
tratamiento de cultivo.
La mayoría de las especies de Navicula crecen adheridas a las rocas o sedimentadas en el
fondo (Tomas, 1997; Medlin et al, 1986). Esto explica que no sean muy abundantes en las
muestras originales, que fueron obtenidas por arrastre de red de plancton en aguas
superficiales con profundidades mayores a dos metros. Sin embargo, las escasas células
transportadas en las muestras son suficientes para que se establezcan colonias dominantes
en los cultivos.

55
III.2.2 Aislamiento por micromanipulación
Las muestras originales y los cultivos de enriquecimiento se utilizaron para realizar los
aislamientos por micromanipulación. De entre todas las especies provenientes de las
muestras y las propagadas en cultivo de enriquecimiento la mayoría fueron diatomeas
(Orden Bacillariophyta), no pudiéndose aislar dinoflagelados (Orden Pyrrhophyta). Es
posible que estos últimos sean más delicados a la manipulación y el traslado, y que el
medio del cultivo utilizado no les haya favorecido. Los cultivos de enriquecimiento
propiciaron la propagación de algunas especies que se encontraban en bajas densidades en
las muestras originales, aumentando las posibilidades de aislamiento por
micromanipulación. Entre las mencionadas anteriormente están las especies: Melosira
nummuloides, Odontella aurita, Coscinodiscus granii, Cyclotella sp., Entomoneis sp.,
Melosira nummuloides, Odontella longicruris, Pediastrum duplex, Scenedesmus y
Thalassionema nitzschioides.
De este modo se fueron adaptando algunas especies a las condiciones de cultivo artificial.
Sin embargo, algunas especies de las muestras originales, incluso de diatomeas, perecían en
estos cultivos. Tal fue el caso de Rhizosolenia sp., Chaetoceros decipiens y Coscinodiscus
centralis. Aunque fueran abundantes en las muestras, morían en las soluciones de
enriquecimiento. Estas especies tampoco se pudieron aislar a partir de las muestras
originales. El método de conservación en frío pudo haberles afectado, pero previno que las
muestras se deterioraran, comenzando a proliferar bacterias. Estas muestras pasaban hasta
48 horas bajo estas condiciones durante el transporte hasta el día que se llegaba al
laboratorio para poderlas procesar. El frío reduce la depredación por el zooplancton que
puede ir en las muestras y reduce la tasa metabólica, disminuyendo el consumo de las
reservas de las células algales, e inhibiendo el desarrollo de bacterias.
La primera especie que se aisló de este modo fue Odontella aurita. En un siguiente
muestreo se continuó con Melosira nummuloides, Coscinodiscus granii, Entomoneis sp.,
Thalassionema nitzschioides, Bacillaria paxillifera, Cyclotella sp., Odontella longicruris y
Pediastrum duplex (dulceacuícola). Odontella longicruris se aisló, pero no se ha
desarrollado con el mismo vigor que especies como O.aurita o Entomoneis sp.
III.2.3 Cultivos establecidos a partir de líneas de reproducción de soluciones
de enriquecimiento
Tetraselmis sp., Nannochloris sp. y Bacillaria paxillifera se obtuvieron por cultivos
consecutivos de algunas muestras originales. Estos cultivos fueron transferidos varias
ocasiones y los resultados fueron cultivos xénicos dominados por estas especies. El cultivo
de Tetraselmis sp. continúa viable desde su obtención hace un año, al igual que el de
Nannochloris sp. Bacillaria paxillifera se obtuvo mezclada con Navicula sp.1, de la cual se
aisló posteriormente por micromanipulación.
III.2.4 Purificación de especies aisladas
En pocos casos los cultivos establecidos por micromanipulación eran monoespecíficos. La
microalga Navicula sp.1 se encontraba frecuentemente propagándose en conjunción con la

56
especie deseada. En estos casos se realizaba de nuevo el aislamiento hasta obtener cultivos
libres de Navicula sp.1. Para lograr esto no había que dejar pasar muchos días debido a que
Navicula sp.1 se reproducía con tal rapidez que dificultaba la purificación. Además, las
células de esta se adhieren a células más grandes de otras especies.
Los cultivos monoespecíficos se inocularon en caja de Petri utilizando el método del
estriado y por gota. De esta manera se pudieron obtener algunos cultivos libres de bacterias.
El estriado permite separar células de bacterias, hongos y microalgas, desarrollándose así
colonias puras de microalgas. Estas colonias luego se inocularon en otras cajas de Petri y
tubos de ensayo con medio de cultivo líquido. Las especies que se obtuvieron como
cultivos puros están listadas en la Tabla 9, entre estas Odontella aurita, Melosira
nummuloides, Entomoneis sp. y Scenedesmus sp.
III.2.5 Proporciones de inoculo para la producción de biomasa
Para todas las especies, un tubo de ensayo con 10 ml de cultivo concentrado fue suficiente
para inocular un frasco con 333ml de medio de cultivo. Para acelerar el crecimiento en la
fase inicial, un cultivo concentrado en frasco de 500 ml (con 333 ml de cultivo) se utilizaba
para inocular de 3 a 4 frascos de 1 Lt de medio de cultivo, lo cual iniciaba la propagación
en el nuevo envase.
III.2.6 Rendimiento en la producción de aceites por especie de microalga
Se logró la obtención de aceites de 12 especies de microalgas. Los resultados se pueden
consultar en la Tabla 11. Los porcentajes oscilaron entre el 0% (Pediastrum duplex) y 17%
en Nannochloris sp. Consideramos que es importante obtener mayor cantidad de biomasa
de cada especie (10 gr es poco), además de realizar varias extracciones de cada especie,
para obtener resultados más precisos. Sería importante también refinar el aceite,
separándolo de otras sustancias que son extraídas por el solvente y producen un error al
obtener el porcentaje de aceite.
El método de extracción mediante el aparato Sohxlet resultó ser eficiente, pero no es
necesariamente el más apropiado para extracciones a gran escala, ya que los costos sería
muy elevados debido al consumo de energía y recursos necesarios.

57
PARTE IV
IV.1 CONCLUSIONES
1. Los diferentes puntos de muestreo considerados en este estudio presentan una gran
variación con respecto a las diferentes especies de microalgas. Esto se debe a la geografía
heterogénea de cada punto y la presencia de drenajes, ríos, poblaciones, etc. Las variantes
geográficas, hídricas y humanas crean condiciones de variación de los parámetros químicos
(micronutrientes y macronutrientes, pH, compuestos tóxicos), físicos (turbidez, sedimentos,
temperatura y salinidad) y bióticos (bacterias y otros microorganismos) del agua. La
variación de estas condiciones incide sobre la diversidad y abundancia de microalgas en un
sitio.
2. Se identificaron 44 especies de microalgas, 32 en los cinco puntos de muestreo que
se compararon bajo tratamientos de cultivo, y 12 adicionales en los sitios no comparados.
Entre estos, 23 especies de diatomeas (Bacillariophyta), 7 especies de clorofíceas
(Chlorophyta) y 5 especies de dinoflagelados (Dinophyta), entre otras. (tabla 7 y 8).
3. Se utilizaron diferentes medios de cultivo, los cuales variaban en la presencia o
ausencia de macronutrientes y micronutrientes, con la intención de determinar si existía
predilección de algunas especies de microalgas por alguno de los medios de cultivo
empleados. No se observo una marcada diferencia, pero si obtuvo un efecto que facilitó el
aislamiento de algunas especies de microalgas. Los efectos de los diferentes medios de
cultivo sobre las muestras y especies de microalgas pueden ser evaluadas de manera más
precisa utilizando medios de cultivo definidos, en los cuales se utiliza como base agua de
mar artificial. El agua de mar natural contiene concentraciones variables de sustancias
iónicas y orgánicas, por lo cual es imposible preparar medios de cultivo definidos a partir
de esta. Se establecio el método de cultivo excitu para la reproducción.
4. El cultivo en frascos de 500 ml y 1L en medio de cultivo f/2 de Guillard probó ser
bastante efectivo, obteniéndose cultivos completamente desarrollados en 15 días para
especies como Nannochloris sp., Odontella aurita, Melosira nummuloides, Scenedesmus
sp., Thalassionema nitzschioides, Tetraselmis sp., Navicula sp., Entomoneis sp., y
Bacillaria paxilifera.
5. Se obtuvieron aceites por medio de extracción por solventes. Los mayores
porcentajes los presentaron las especies Nannochloris sp., Navicula sp.1, Scenedesmus sp.,
Melosira nummuloides y Odontella aurita, con 17%, 16%, 15%, 12% y 8%,
respectivamente. De estas, las que presentan mayor facilidad para ser cosechadas son
Melosira nummuloides y Odontella aurita, por su estructura filamentosa y formación de
conglomerados. Navicula sp.1 es difícil de cosechar, pues tiende a adherirse a la superficie
del recipiente de cultivo. Nannochloris sp. y Scenedesmus sp. producen cultivos en
suspensión homogénea, por lo que se requiere una técnica de filtración fina o la floculación
para obtener la biomasa. Debido a estos importantes aspectos prácticos, se sugiere el

58
cultivo de M. nummuloides y O. aurita, y evaluar de qué manera puede aumentarse su
producción de aceite en relación a la biomasa.
6. Se observo que varias de las especies encontradas al inicio en las muestras fijadas,
no se lograron aislar, posiblemente por ser altamente especialistas, siendo sensibles a
variaciones de temperatura, salinidad, fotoperiodo, presencia y concentración de nutrientes
específicos. Esto las hace más susceptibles como para ser cultivadas utilizando medios de
cultivo no específicos, y más delicadas para sobrevivir a las condiciones del transporte.
7. La especie de microalga más común fue Navicula sp. aunque los medios de cultivo
deficientes en silicato limitaron su crecimiento y desarrollo.
8. La técnica de micromanipulación resultó bastante eficaz para el aislamiento de
microalgas, aunque las especies más delicadas de microalgas como Rhizosolenia sp.,
Coscinodiscus centralis, Fragilaria spp., Nitzschia longissima, se dañaban al intentar
succionarlas y lavarlas. Esto impone un reto para la persona que realiza los aislamientos,
que debe poseer un alto control fino para atrapar células delicadas sin lastimarlas. La
técnica fue exitosa para especies unicelulares no alargadas (Entomoneis sp., Scenedesmus
sp., Navicula sp., Bacillaria paxilifera y Coscinodiscus granii) y colonias filamentosas
(Melosira nummuloides, Odontella aurita, Thalassionema nitzschioides). La dificultad
aumentaba en relación inversa al tamaño celular de algas unicelulares.
9. Si se encontraron especies de microalgas en la costa atlántica guatemalteca con
potencial para ser utilizadas en la obtención de biodiesel, especialmente dentro del grupo de
las diatomeas (Clase Bacillariophyta), y algunas Chlorophyta.

59
IV.2 RECOMENDACIONES
1. Se recomienda tomar en consideración, otros cuerpos de agua, como por ejemplo el
Océano Pacífico, lagos y ríos del país, para tener una base de datos completa de la
biodiversidad de especies con que se cuenta.
2. Para muestras tomadas a distancias muy grandes se recomendaría mejorar la cadena
de transporte, o sea utilizar un transporte más rápido, por ejemplo vía aérea para exponer el
menor tiempo a las microalgas a condiciones de estrés
3. Utilizar una red de plancton específica para fitoplancton ya que la que se empleo no
permitía separar al zooplancton de las muestras lo que afectaba el proceso de identificación,
aislamiento y en un momento dado se podría pensar que aún en frío el zooplancton podría
alimentarse de especies de microalgas especificas y evitar que se observarán por lo menos
tres días después de tomada la muestra.
4. Se sugiere la caracterización de las bacterias que se aislaron simultáneamente en los
cultivos, para que se pueda hacer uso de antibióticos específicos que permitan controlar su
crecimiento y se puedan obtener cultivos libres de contaminación por bacterias.
5. Se recomienda el estriado en placa, como técnica de aislamiento efectiva, ya que se
logra una buena separación a mediano plazo de algunas cepas de microalgas que se
encuentran contaminadas con bacterias y hongos incluso.
6. Se recomienda la refinación de los aceites obtenidos para conocer el grado de
pureza de estos.
7. Se sugiere buscar alternativas en las técnicas utilizadas para la extracción de los
aceites, con el objetivo de hacer más rentable y eficiente el proceso.

60
IV.3 REFERENCIAS BIBLIOGRÁFICAS
1. Andersen, R.A., J. A. Berges, P.J. Harrison y M.M. Watanabe. 2005. Recipes for
freshwater and seawater media. En: Andersen, R.A., editor. Algal Culturing
Techniques. Elsevier Academic Press, USA. pp. 429 – 538.
2. Andersen, R.A. Y M. Kawachi. 2005. Traditional Microalgae Isolation Techniques. En:
Andersen, R.A., editor. Algal Culturing Techniques. Elsevier Academic Press, USA.
pp. 83 – 100.
3. Arrivillaga, A. 2002. Evaluación de la presencia de Hydrilla verticillata en la región de
Río Dulce y el Lago de Izabal: diagnóstico general e identificación de medidas de
control. Oficina Técnica de Biodiversidad, Consejo Nacional de Áreas Protegidas, y
Fondo Nacional para la Conservación de la Naturaleza.
4. Arrivillaga A y M. García . 2004. Chapter 18. Status of the Coral Reefs of the
Mesoamerican Barier Reefs Systems Project Region and Reefs of El Salvador,
Nicaragua and the Pacific Coasts of Mesoamerica. Status of the Coral Reefs of the
World. 2004.
5. Becker, E.W. 1995. Microalgae: biotechnology and microbiology. Cambridge
University Press. Cambridge. 310 pp.
6. Bol, A. 2002. Análisis de la Contaminación presente en la Cuenca del Río Dulce y
Lago de Izabal, Guatemala, Tesis para optar al grado de Licenciatura en Química.
Facultad de Ciencias Químicas y Farmacia, Universidad de San Carlos de
Guatemala.
7. Carrillo L, Salaverría A, Pacas L. Martinez J, Otaolaurruchi R, Tay C. 2000. Evaluación
del Recurso Pesquero y Oceanográfico del Atlántico Guatemalteco, durante es año
2000. Unidad de Análisis de Información Geográfica del CEMA Universidad de San
Carlos de Guatemala Centro de Estudios del Mar y Acuicultura –CEMA– Dirección
General de Investigación –DIGI. 63 p.
8. Cazali, G. 1998. Informe Final de Tesis. Inventario de los pelecípodos de la Costa
Atlántica Guatemalteca con énfasis en especies comestibles. Escuela de Biología.

61
Facultad de Ciencias Químicas y Farmacia. Universidad de San Carlos de
Guatemala.
9. Chisti, Y. Modern systems of plant cleaning. En: Robinson, R., Batt, C., Patel, P.,
editores. Encyclopedia of food microbiology. Academic Press; 1999. p. 1806–15.
10. Chisti, Y. 2007. Biodiesel from microalgae. Biotechnology Advances. 25 (2007) 294–
306.
11. Cuevas, M.C. 1984. Contribución al Estudio de las Algas Marinas Macroscópicas del
Atlántico de la República de Guatemala. Tesis de Licenciatura. Escuela de Biología,
USAC.
12. Dawson, P. A. 1973. Observations on the structure of some forms of Gomphonema
parvulum Kütz. III. Frustule formation. J. Phycol. 9:353–64.
13. de Brouwer, J. F. C., y Stal, L. J. 2002. Daily fluctuation of exopolymers in cultures of
the benthic diatoms Cylindrotheca closterium amd Nitzschia sp. (Bacillariophyceae).
J. Phycol. 38:464–72.
14. Department of Commerce; NOAAM, USAID, PROARCA-COSTAS y MARN. 2001.
Atlas de los Recursos Costero-Marino e Índices de Sensibilidad Ambiental para la
zona del Puerto Quetzal en el Océano Pacífico y la Línea costera del Mar Caribe,
Guatemala. Department of Commerce /NOAAM/ USAID/ PROARCA-COSTAS/
MARN. EEUU-Guatemala
15. Díaz, L. 2004. Informe Final. Evaluación de la selectividad de cuatro artes de pesca
para los camarones blanco (Litopenaeus schmitti), café (Farfanpenaeus notalis) y
camaroncillo (Xiphopenaeus kroyeri). Ejercicio Profesional Supervisado. Escuela de
Biología. Facultad de Ciencias Químicas y Farmacia. Universidad de San Carlos de
Guatemala.
16. Dix, A., M. Maldonado, M. Dix, O. De Bocaletti, R. Girón, I. De la Roca, A.C. Bailey,
K. Herrera, J.F. Pérez, K. Pierola y G. Rivera. 1999. El impacto de la cuenca del río
Polochic sobre la integridad biológica del Lago de Izabal. Informe Final – Proyecto
No. 4. Universidad del Valle de Guatemala, Centro de Estudios Ambientales,
Consejo Nacional de Ciencia y Tecnología CONCYT, Fundación Defensores de la
Naturaleza. 148 p.

62
17. Fonseca, B.C, Arrivillaga A. 2003. Latin American Coral Reefs. Chapter: Coral Reefs
in Guatemala. Elsevier Science, B.V. All Rights Reserved.
18. Fuentes, A. 2007. Informe de Experiencias Docentes con la Comunidad-EPS- de la
carrera de Biología. Facultad de Ciencias Químicas y Farmacia. Universidad de San
Carlos de Guatemala.
19. Goodenough, U. W., y Heuser, J. E. 1985. The Chlamydomonas cell wall and its
constituent glycoproteins analyzed by the quick-freeze, deep-etch technique. J. Cell
Biol. 101:1550–68.
20. Granum, E., y Myklestad, S. M. 2001. Mobilization of �-1,3-glucan and biosynthesis
of amino acids induced by NH4+ addition to N-limited cells of the marine diatom
Skeletonema costatum (Bacillariophyceae). J. Phycol. 37:772–82.
21. Guillou, L., Chretiennot-Dinet, M.-J., Medlin, L. K., Claustre, H., Loiseaux-de Goer, S.,
y Vaulot, D. 1999. Bolidomonas: a new genus with two species belonging to a new
algal class, the Bolidophyceae (Heterokonta). J. Phycol. 35:368–81.
22. Goodwin, T. W. 1974. Carotenoids and biliproteins. In Algal Physiology and
Biochemistry, ed. W. D. P. Stewart, pp. 176–205. Berkeley: Univ. Calif. Press.
23. Grobbelaar JU. Algal nutrition. In: Richmond A, editor. Handbook of microalgal
culture: biotechnology and applied phycology.Blackwell; 2004. p. 97–115.
24. Herrera, K. 1999 Indicadores Biológicos de la calidad del agua en el Río Polochic y de
la integridad biológica del Lago de Izabal. Tesis de Grado. Universidad del Valle de
Guatemala.
25. Holdridge, L. R. 1967. «Life Zone Ecology». Tropical Science Center. San José, Costa
Rica. (Traducción del inglés por Humberto Jiménez Saa: «Ecología Basada en Zonas
de Vida», 1a. ed. San José, Costa Rica: IICA, 1982).
26. Huizing, H. J., Rietema, H., y Sietsma, J. H. 1979. Cell wall constituents of several
siphonaceous green algae in relation to morphology and taxonomy. Br. Phycol. J.
14:25–32.

63
27. Ikan, R. 1969. Natural Products. Laboratory Guide. Department of Organic Chemistry.
Hebrew University, Jerusalem. Academic Press. New Cork.
28. Lavens, P. y P. Sorgeloos. 1996. Manual on the Production and Use of Live Food for
Aquaculture. FAO Fisheries Technical Paper. No. 361. FAO (Food and Agriculture
Organization of the United Nations), Rome. 295 pp.
29. Lee, R.E. 2008. Phycology. 4ta ed. Cambridge University Press. New York. 547 pp.
30. Leiva, X. 2006. Informe de Tesis de Biología. Pastos marinos: composición
comunitaria, biomasa de pastos marinos y morfometría de Thalassia testudinum, en
dos sitios de Bahía La Graciosa, Izabal, Guatemala. Facultad de Ciencias y
Humanidades. Universidad del Valle de Guatemala.
31. Mague, T. H., y Holm-Hansen, O. 1975. Nitrogen fixation on a coral reef. Phycologia
14:87–92.
32. Mansour, M. P., Volkman, J. K., Jackson, A. E., y Blackburn, S.I. 1999. The fatty acid
and sterol composition of five marine dinoflagellates. J. Phycol. 35:710–29.
33. Meeks, J. C., Campbell, E. L., Summers, M. L., y Wong, F.C. 2002. Cellular
differentiation in the cyanobacterium Nostoc punctiforme. Arch. Microbiol.
178:395–403.
34. Medlin, L. K., Crawford, R. M., y Andersen, R. A. 1986. Histochemical and
ultrastructural evidence for the function of the labiate process in the movement of
centric diatoms. Br. Phycol. J. 21:297–301.
35. Metting FB. Biodiversity and application of microalgae. J Ind Microbiol 1996;17:477–
89.
36. Molina Grima E, Acién Fernández FG, García Camacho F, Chisti Y. Photobioreactors:
light regime, mass transfer, and scaleup. J Biotechnol 1999;70:231–47.
37. Naciones Unidas. 1992. Convención Marco de las Naciones Unidas sobre el Cambio
Climático. FCCC/INFORMAL/84.GE 05-62301 (S) 220705 220705.

64
38. Naciones Unidas. 1998. Protocolo de Kyoto de la Convención Marco de las Naciones
Unidas sobre el Cambio Climático. FCCC/INFORMAL 83*/ge 05-61702 (s)130605
130605.
39. Orellana, I. 2004. Informe Final de Tesis. Relación de la Estructura de tallas y biometría
del camarón café (Farfanpenaeus notalis) de la Bahía de Amatique, Izabal,
Guatemala. Centro de Estudios del Mar y Acuicultura. Universidad de San Carlos de
Guatemala.
40. Otero, A., y Vincenzini, M. 2004. Nostoc (Cyanophyceae) goes nude: extracellular
polysaccharides serve as a sink for reducing power under unbalanced C/N
metabolism. J. Phycol. 40:74–81.
41. Padan, E. 1979. Facultative anoxygenic photo syn thesis in cyanobacteria. Annu. Rev.
Plant Physiol. 30:27–40.
42. Phillips, R.C. y E. G. Meñez. 1988. Seagrasses. Smithsonian Contributions to The
Marine Sciences. No 34. Smithsonian Institution Press. Washington D.C. 104 pp.
43. Prado, L. 1990. Informe Final de Tesis. Colecta, clasificación y distribución de las
especies de gasterópodos en la Costa Atlántica de Guatemala. Escuela de Biología.
Facultad de Ciencias Químicas y Farmacia. Unviersidad de San Carlos de
Guatemala.
44. Pulz O. Photobioreactors: production systems for phototrophic microorganisms. Appl
Microbiol Biotechnol 2001;57:287–93.
45. Ratledge C,Wynn JP. The biochemistry and molecular biology of lipid accumulation in
oleaginous microorganisms. Adv Appl Microbiol 2002;51:1–51.
46. Rodolfi, L. et al. 2008. Microalgae for Oil: Strain Selection, Induction of Lipid
Synthesis and Outdoor Mass Cultivation in an Low-Cost Photobioreactor.
Biotechnology and Bioengineering. Vol. 102, No. 1, January 1, 2009
47. Romero, R. 2005. Informe de Tesis. Elaboración, Análisis y Comparación de Biodiesel
a partir del aceite de Palma usado (Elaeis guineensis) mediante dos procesos, a nivel

65
de Planta Piloto. Escuela de Ingeniería Química. Facultad de Ingeniería. Universidad
de San Carlos de Guatemala.
48. Romero-Oliva, C. 2007. Informe Final de Tesis. Caracterización del hábitat del manatí
(Trichechus manatus manatus) Linnaeus 1758, en el Golfo de Honduras. Escuela de
Biología, Facultad de Ciencias Químicas y Farmacia. Universidad de San Carlos de
Guatemala. 98pp.
49. Sawayama S, Inoue S, Dote Y, Yokoyama S-Y. CO2 fixation and oil production
through microalgae. Energy Convers Manag 1995;36: 729–31.
50. Salaverria, A., y F. Rosales, 1993. Ecología Pesquera de la Costa Atlántica de
Guatemala. Evaluación Inicial. Bahía de Amatique, Izabal. Informe de Avance,
Septiembre 1993. Centro de Estudios del Mar y Acuicultura. USAC. 105 p.
51. Schmaljohann, R., y Röttger, R. 1978. The ultrastructure and taxonomic identity of the
symbiotic algae of Heterostegina depressa (Foraminifera: Nummulitidae). J. Mar.
Biol. Assoc. UK 58:227–37.
52. Sheehan, J., T. Dunahay, J. Benemann, P. Roessler, 1998. A look back at the U.S.
Department of Energy´s Aquatic Species Program-Biodiesel from Algae. National
Renewable Energy Laboratory. U.S. Department of Energy`s. Office of Fuel
Development. Golden, Colorado. 328 pp.
53. Sitio Web de Culture Collection of Algae and Protozoa (CCAP). 2008. Disponible en
http://www.ccap.ac.uk/index.htm
54. Smith, G. M. 1955. Cryptogamic Botany, Vol. 1, 2da ed. New York: McGraw-Hill.
55. Stanier, R. Y. 1973. Autogrophy and hetero trophy in unicellular blue-green algae. In
The Biology of the Blue-Green Algae, ed. N. G. Carr and B. A. Whitton, pp. 501–
18. Berkeley: Univ. Calif. Press.
56. Stewart, W. D. P., y Pearson, H. W. 1970. Effects of aerobic and anaerobic conditions
on growth and metabolism of blue-green algae. Proc. R. Soc. Lond. [B] 175:293–
311.

66
57. Terrones, J. 2007. Biocombustibles. Congreso ADIAT 2007. Clusters de Innovación
para el Desarrollo Regional Sustentable. Promotora de Palma de Soconusco, S.A. de
C.V.
58. Terry KL, Raymond LP. System design for the autotrophic production of microalgae.
Enzyme Microb Technol 1985; 7:474–87.
59. Tomas, C.R. 1997. Identifying Marine Phytoplankton. Academic Press. San Diego,
California. 858 pp.
60. Tomkins, J., M. DeVille, J.G. Day y M.F. Turner. 1995. Culture Collection of Algae
and Protozoa. Catalog of Strains. Ambleside, UL. 204 pp.
61. Torrentera, L., A. Blanco y G.J. Tacon. 1989. Proyecto GCP/RLA/075/ITA. Apoyo a
las actividades regionales de Acuicultura para América Latina y El Caribe.
Organización de las Naciones Unidas para la Agricultura y El Caribe -FAO-,
Brasilia.
62. Tredici, M.R. Bioreactors, photo. En: Flickinger, M.C., Drew, S.W., editores.
Encyclopedia of bioprocess technology: fermentation, biocatalysis and
bioseparationWiley; 1999. p. 395–419.
63. Vides-Llarena, G. 2003. Caracterización y Comparación de los Arrecifes Coralinos de
Bajo de Languaja, King Fish, Faro Rojo, Arrecife de Chatarra y Barco Hundido, de
la Bahía de Amatique, Izabal, Guatemala. FUNDAECO.
64. Weiss, T. 1983. Food Oils and their Uses. 2a edición. EEUU: AVI Publishing
Company.
65. Yoshii, Y., Takaichi, S., Maoka, T., e Inouye, I. 2003. Photosynthetic pigment
composition in the primitive green alga Mesostigma viride (Prasinophyceae):
phylogenetic and evolutionary implications. J. Phycol. 39:570–6.

67
IV.4 ANEXOS

68
ANEXO 1 IV.4.1 Micrografías de las especies de microalgas
Fuente: Todas las microfotografías fueron tomadas y editadas por el Br. Rodrigo Blanco
Estrada, Auxiliar de Investigación en el presente proyecto de investigación.
IV.4.1.1 Orden Bacillariophyta
Fotografía 1
Phylum: Heterokontophyta
Clase: Bacillariophyceae
Orden:Biddulphiales
Familia: Chaetocerotaceae
Especie: Bacteriastrum furcatum
Shadbolt
Aumento: 400X
Microscopio invertido con contraste de
fases.
Abundancia: Rara en pocas muestras.
No aislada.
Marina Fuente: Proyecto Fodecyt 43-2007
Fotografía 2
Phylum: Heterokontophyta
Clase: Bacillariophyceae
Orden:Biddulphiales
Familia: Chaetocerotaceae
Especie: Chaetoceros decipiens
Cleve
Aumento: 400X
Microscopio invertido con contraste de fases.
Abundancia: Muy abundante en varias muestras.
No aislada.
Polihalofila (Marina y agua dulce). Fuente: Proyecto Fodecyt 43-2007

69
Fotografía 3
Phylum: Heterokontophyta
Clase: Bacillariophyceae
Orden: Biddulphiales
Familia: Coscinodiscaceae
Especie: Coscinodiscus centralis
Ehrenberg
Aumento: 1000X
Microscopio óptico
convencional
Abundancia: Común en varias
muestras.
No aislada.
Dulceacuícola Fuente: Proyecto Fodecyt 43-2007
Fotografía 4
Phylum: Heterokontophyta
Clase: Bacillariophyceae
Orden: Biddulphiales
Familia: Coscinodiscaceae
Especie: Coscinodiscus granii
Gough
Aumento: 400 X
Microscopio óptico con
contraste de fases.
Abundancia: Común en
algunas muestras.
Aislada en medio de cultivo
líquido y sólido.
Marina Fuente: Proyecto Fodecyt 43-
2007

70
Fotografía 5
Phylum: Heterokontophyta
Clase: Bacillariophyceae
Orden: Thalassiosirales
Familia:
Stephanodiscaceae
Especie: Cyclotella sp.
Aumento: 400X
Microscopio invertido con
contraste de fases.
Abundancia: Abundante
en pocas muestras.
Aislada en medio de
cultivo líquido.
Marina Fuente: Proyecto Fodecyt
43-2007
Fotografía 6
Phylum: Heterokontophyta
Clase: Bacillariophyceae
Orden: Bacillariales
Familia: Bacillariaceae
Especie: “diatomea verde”
Aumento: 400X
Microscopio invertido con
contraste de fases.
Aislada en medio de cultivo
líquido. Fuente: Proyecto Fodecyt 43-
2007

71
Fotografía 7
Phylum: Heterokontophyta
Clase: Bacillariophyceae
Orden: Bacillariales
Familia: Bacillariaceae
Especie: “Diatomea anclada”
Aumento: 400X
Microscopio invertido con contraste de
fases.
Abundancia: Rara
No aislada.
Dulceacuícola Fuente: Proyecto Fodecyt 43-2007
Fotografía 8
Phylum: Heterokontophyta
Clase: Bacillariophyceae
Orden: Surirellales
Familia: Entomoneidaceae
Especie: Entomoneis sp.
Aumento: 400X
Microscopio invertido con contraste de
fases.
Aislada en medio de cultivo líquido y
sólido.
Marina. Fuente: Proyecto Fodecyt 43-2007
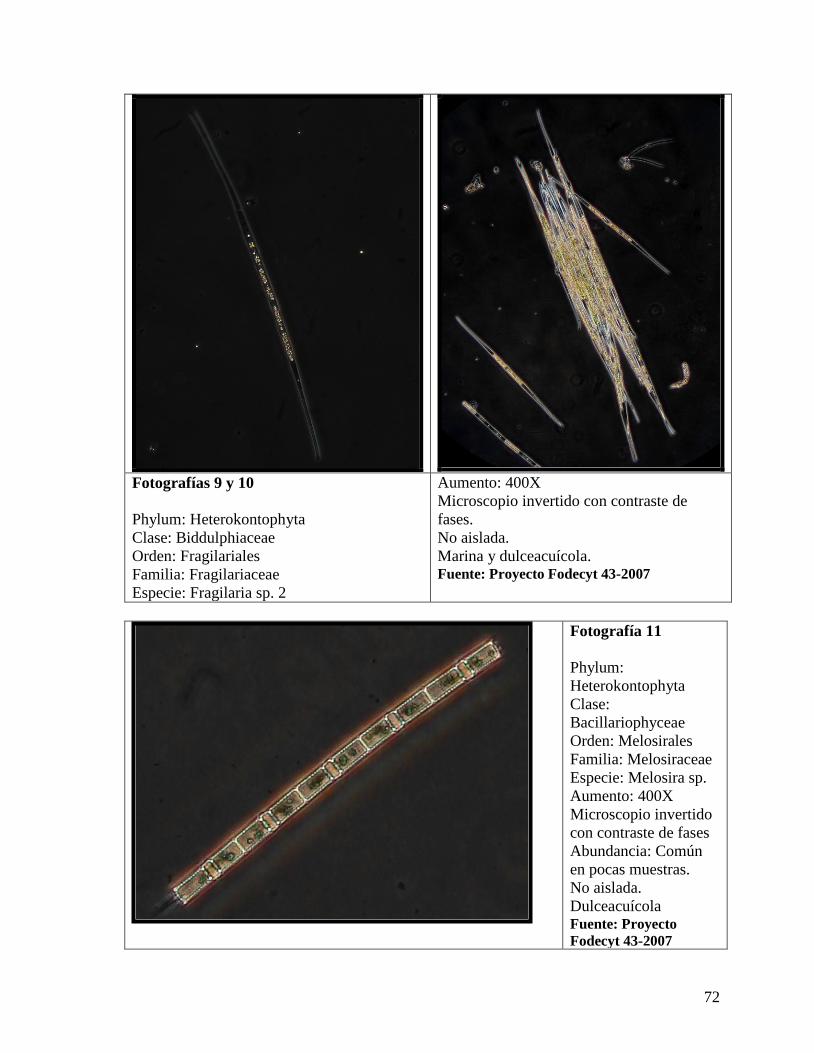
72
Fotografías 9 y 10
Phylum: Heterokontophyta
Clase: Biddulphiaceae
Orden: Fragilariales
Familia: Fragilariaceae
Especie: Fragilaria sp. 2
Aumento: 400X
Microscopio invertido con contraste de
fases.
No aislada.
Marina y dulceacuícola. Fuente: Proyecto Fodecyt 43-2007
Fotografía 11
Phylum:
Heterokontophyta
Clase:
Bacillariophyceae
Orden: Melosirales
Familia: Melosiraceae
Especie: Melosira sp.
Aumento: 400X
Microscopio invertido
con contraste de fases
Abundancia: Común
en pocas muestras.
No aislada.
Dulceacuícola Fuente: Proyecto
Fodecyt 43-2007
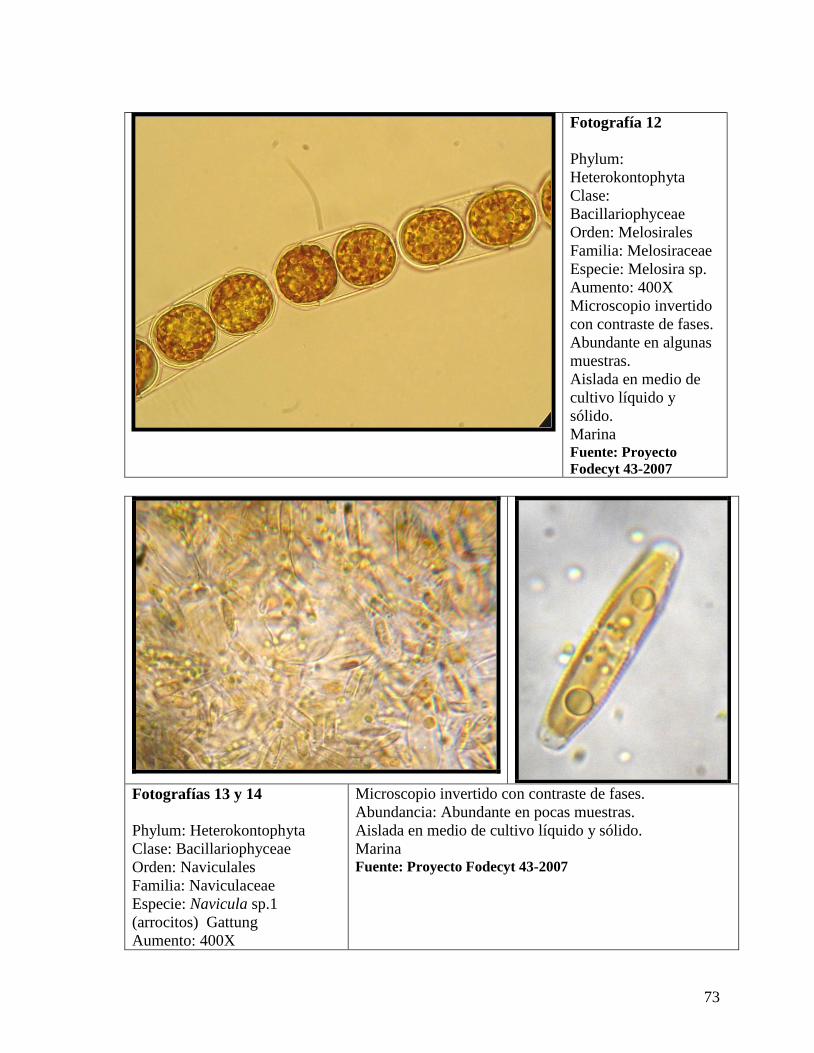
73
Fotografía 12
Phylum:
Heterokontophyta
Clase:
Bacillariophyceae
Orden: Melosirales
Familia: Melosiraceae
Especie: Melosira sp.
Aumento: 400X
Microscopio invertido
con contraste de fases.
Abundante en algunas
muestras.
Aislada en medio de
cultivo líquido y
sólido.
Marina Fuente: Proyecto
Fodecyt 43-2007
Fotografías 13 y 14
Phylum: Heterokontophyta
Clase: Bacillariophyceae
Orden: Naviculales
Familia: Naviculaceae
Especie: Navicula sp.1
(arrocitos) Gattung
Aumento: 400X
Microscopio invertido con contraste de fases.
Abundancia: Abundante en pocas muestras.
Aislada en medio de cultivo líquido y sólido.
Marina Fuente: Proyecto Fodecyt 43-2007

74
Fotografía 15
Phylum: Heterokontophyta
Clase: Bacillariophyceae
Orden: Naviculales
Familia: Naviculaceae
Especie: Navicula sp.
Aumento: 400X
Microscopio invertido con
contraste de fases.
Abundancia: Abundante en
muestras cultivadas.
Aislada en medio de
cultivo líquido.
Marina Fuente: Proyecto Fodecyt
43-2007
Fotografía 16
Phylum: Heterokontophyta
Clase: Bacillariophyceae
Orden: Bacillariales
Familia: Bacillariaceae
Especie: Nitzschia sp.
Aumento: 400X
Microscopio invertido con
contraste de fases.
Abundancia: Común en pocas
muestras.
Aislada en medio de cultivo
sólido.
Marina Fuente: Proyecto Fodecyt 43-2007

75
Fotografía 17
Phylum: Heterokontophyta
Clase: Bacillariophyceae
Orden: Bacillariales
Familia: Bacillariaceae
Especie: Pseudinitzschia longissima
(Brébisson in Kützing)
Ralfs
Aumento: 1000X
Microscopio óptico convencional.
Abundancia: Rara en pocas
muestras.
No aislada. Marina Fuente: Proyecto Fodecyt 43-2007
Fotografías 18, 19 y 20
Phylum: Heterokontophyta
Clase: Bacillariophyceae
Orden: Biddulphiales
Familia: Triceratiaceae
Especie: Odontella aurita (Lyngbye) C.
A. Agardh
Aumento: 400X
Microscopio invertido con contraste de
fases.
Abundancia: Común en pocas muestras.
Aislada en medio ce cultivo líquido y
sólido.
Marina Fuente: Proyecto Fodecyt 43-2007

76
Fotografías 21 y 22
Phylum: Heterokontophyta
Clase: Bacillariophyceae
Orden: Biddulphiales
Familia: Triceratiaceae
Especie: Odontella longicruris (Greville) Hoban
Aumento: 400X
Microscopio invertido con contraste de
fases.
Abundancia: Común en pocas muestras.
Aislada en medio de cultivo líquido.
Marina Fuente: Proyecto Fodecyt 43-2007
Fotografía 23
Phylum: Heterokontophyta
Clase: Bacillariophyceae
Orden: Naviculales
Familia: Pleurosigmataceae
Especie: Pleurosigma sp.
Aumento: 400X
Microscopio invertido con contraste de
fases.
Abundancia: Rara, en pocas muestras.
Aislada en medio de cultivo líquido.
Marina Fuente: Proyecto Fodecyt 43-2007

77
Fotografía 24
Phylum: Heterokontophyta
Clase: Bacillariophyceae
Orden: Rhizosoleniales
Familia: Rhizosoleniaceae
Especie: Rhizosolenia sp.
Aumento: 1000X
Microscopio óptico convencional.
Abundancia: Abundante, en pocas
muestras.
No aislada.
Polihalófila (marina y dulceacuícola) Fuente: Proyecto Fodecyt 43-2007
Fotografía 25
Phylum:
Heterokontophyta
Clase:
Bacillariophyceae
Orden: No asignado
Familia: Diatomaceae
Especie: Tabellaria
sp.
Aumento: 400X
Microscopio
invertido con
contraste de fases.
Abundancia: Rara, en
pocas muestras.
No aislada.
Marina Fuente: Proyecto
Fodecyt 43-2007

78
Fotografía 26
Phylum:
Heterokontophyta
Clase:
Bacillariophyceae
Orden:
Thalassionematales
Familia:
Thalassionemataceae
Especie: Thalassionema
frauenfeldii
(Grunow)
Hallegraeff
Aumento: 400X
Microscopio invertido con
contraste de fases.
Abundancia: Rara, en pocas
muestras.
Aislada en medio de cultivo
líquido. Marina Fuente: Proyecto Fodecyt 43-
2007
Fotografía 27
Phylum:
Heterokontophyta
Clase:
Bacillariophyceae
Orden:
Thalassionematales
Familia:
Thalassionemataceae
Especie:
Thalassionema
nitzschioides
(Grunow)
Mereschkowsky
Aumento: 400X
Microscopio invertido con
contraste de fases.
Abundancia: Rara, en pocas
muestras.
Aislada en medio de cultivo
líquido. Marina Fuente: Proyecto Fodecyt 43-2007

79
Fotografía 28
Phylum: Heterokontophyta
Clase: Bacillariophyceae
Orden: Biddulphiales
Familia: Thalassiosiraceae
Especie: Thalassisira sp.
Aumento: 400X
Microscopio óptico convencional.
Abundancia: Rara, en pocas muestras.
No aislada.
Marina Fuente: Proyecto Fodecyt 43-2007

80
IV.4.1.2 Phylum Chlorophyta
Fotografía 29
Phylum: Chlorophyta
Clase: Chlorophyceae
Orden: Chlorococcales
Familia: Dictyosphaeriaceae
Especie: Botryococcus braunii
Kützing
Aumento: 400X
Microscopio invertido con
contraste de fases.
Abundancia: Abundante.
Aislada en medio de cultivo
líquido.
Dulceacuícola Fuente: Proyecto Fodecyt 43-
2007
Fotografía 30
Phylum: Chlorophyta
Clase: Chlorophyceae
Orden: No determinado
Familia: No determinado
Especie: “Clorofícea tren”
Aumento: 1000X
Microscopio óptico
convencional.
Abundancia: Rara, en pocas
muestras.
No aislada.
Dulceacuícola Fuente: Proyecto Fodecyt 43-
2007
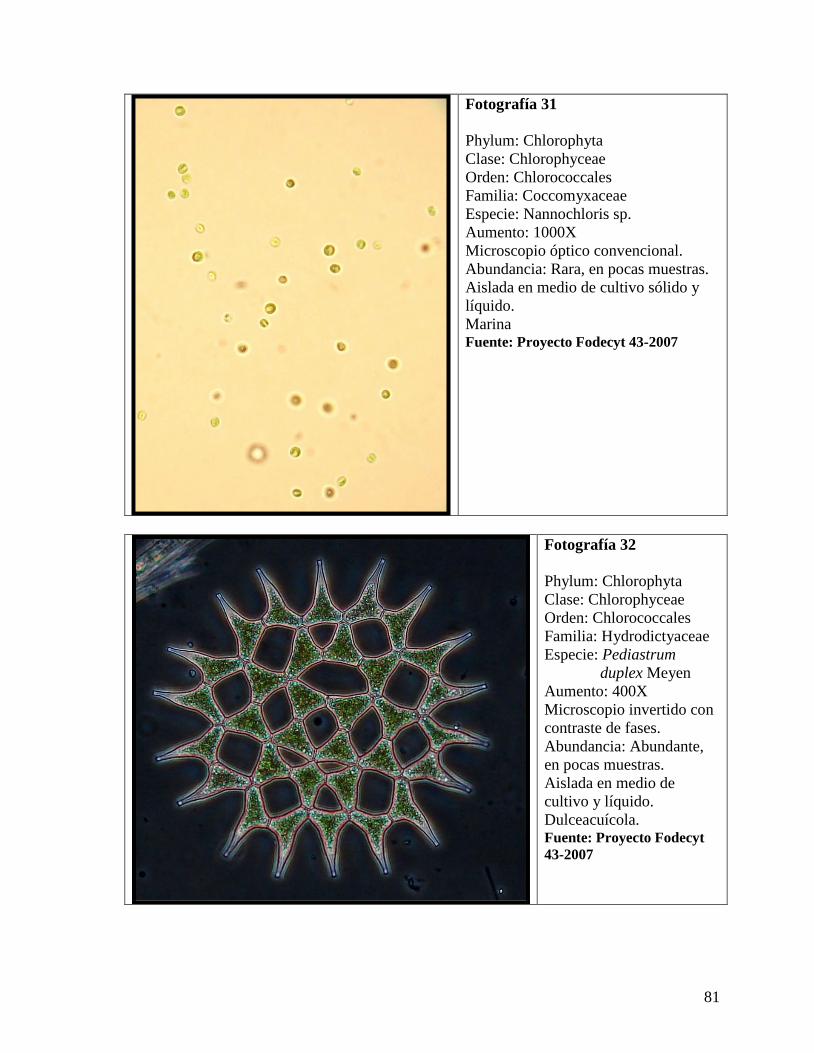
81
Fotografía 31
Phylum: Chlorophyta
Clase: Chlorophyceae
Orden: Chlorococcales
Familia: Coccomyxaceae
Especie: Nannochloris sp.
Aumento: 1000X
Microscopio óptico convencional.
Abundancia: Rara, en pocas muestras.
Aislada en medio de cultivo sólido y
líquido.
Marina Fuente: Proyecto Fodecyt 43-2007
Fotografía 32
Phylum: Chlorophyta
Clase: Chlorophyceae
Orden: Chlorococcales
Familia: Hydrodictyaceae
Especie: Pediastrum
duplex Meyen
Aumento: 400X
Microscopio invertido con
contraste de fases.
Abundancia: Abundante,
en pocas muestras.
Aislada en medio de
cultivo y líquido.
Dulceacuícola. Fuente: Proyecto Fodecyt
43-2007
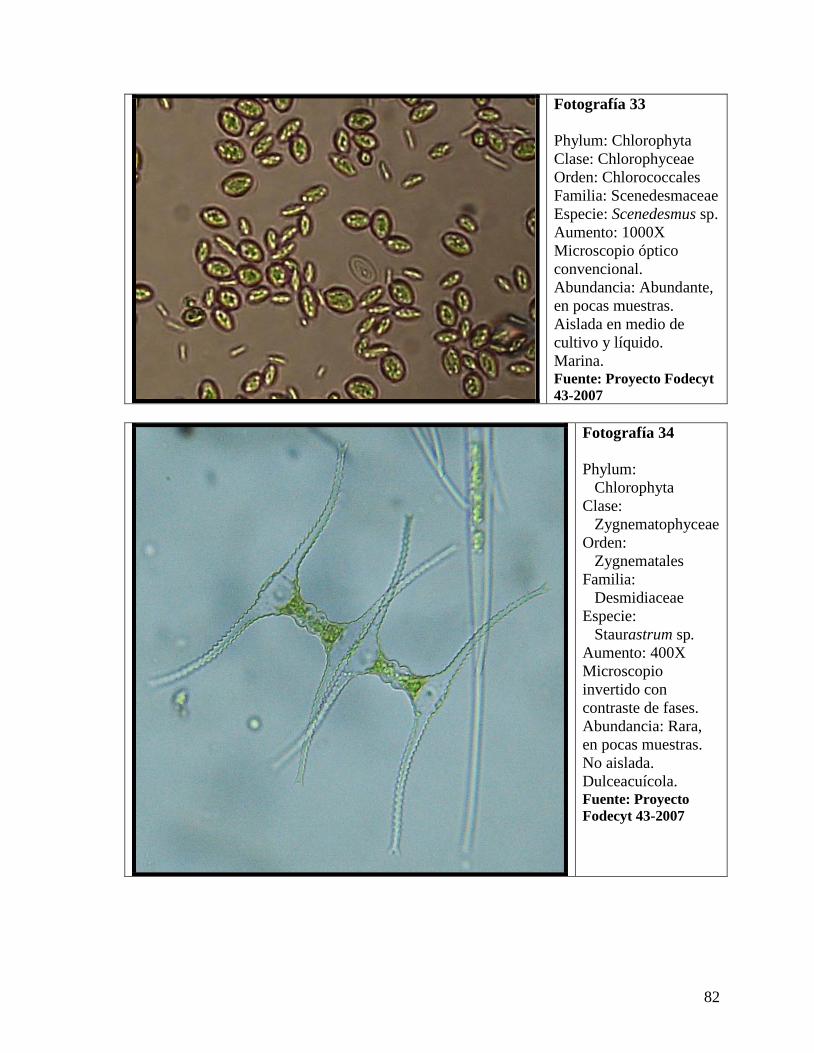
82
Fotografía 33
Phylum: Chlorophyta
Clase: Chlorophyceae
Orden: Chlorococcales
Familia: Scenedesmaceae
Especie: Scenedesmus sp.
Aumento: 1000X
Microscopio óptico
convencional.
Abundancia: Abundante,
en pocas muestras.
Aislada en medio de
cultivo y líquido.
Marina. Fuente: Proyecto Fodecyt
43-2007
Fotografía 34
Phylum:
Chlorophyta
Clase:
Zygnematophyceae
Orden:
Zygnematales
Familia:
Desmidiaceae
Especie:
Staurastrum sp.
Aumento: 400X
Microscopio
invertido con
contraste de fases.
Abundancia: Rara,
en pocas muestras.
No aislada.
Dulceacuícola. Fuente: Proyecto
Fodecyt 43-2007

83
IV.4.1.3 Clase Cyanophyceae
Fotografía 35
Phylum: Bacteria
Clase: Cyanobacteria
Orden: No asignado
Familia: Microcystaceae
Especie: Microcystis wesenbergii
(Komárek) Komárek
Aumento: 400X
Microscopio invertido con
contraste de fases.
Abundancia: Abundante, en
pocas muestras.
No aislada.
Dulceacuícola Fuente: Proyecto Fodecyt 43-2007
IV.4.1.4 Orden Dynophyta
Fotografía 36
Phylum: Dinophyta
Clase: Dinophyceae
Orden: Peridiniales
Familia: Ceratiaceae
Especie: Ceratium furca
(Ehrenberg) Claparéde &
Lachmann
Aumento: 400X
Microscopio invertido con
contraste de fases.
Abundancia: Abundante, en
algunas muestras.
No aislada.
Marina Fuente: Proyecto Fodecyt 43-
2007

84
Fotografía 37
Phylum: Dinophyta
Clase: Dinophyceae
Orden: Peridiniales
Familia: Ceratiaceae
Especie: Ceratium fusus
(Ehrenberg) Dujardin
Aumento: 400X
Microscopio invertido con
contraste de fases.
Abundancia: Abundante, en
algunas muestras.
No aislada.
Marina Fuente: Proyecto Fodecyt 43-2007
Fotografía 38
Phylum: Dinophyta
Clase: Dinophyceae
Orden: Peridiniales
Familia: Ceratiaceae
Especie: Ceratium
trichoceros
(Ehrenberg)
Kofoid
Aumento: 400X
Microscopio invertido con
contraste de fases.
Abundancia: Abundante,
en algunas muestras.
No aislada.
Marina Fuente: Proyecto Fodecyt
43-2007

85
Fotografía 39
Phylum: Dinophyta
Clase: Dinophyceae
Orden: Peridiniales
Familia: Ceratiaceae
Especie: Ceratium sp.
Aumento: 400X
Microscopio invertido
con contraste de fases.
Abundancia:
Abundante, en
algunas muestras.
No aislada.
Marina Fuente: Proyecto
Fodecyt 43-2007
Fotografía 40
Phylum: Dinophyta
Clase: Dinophyceae
Orden: Peridiniales
Familia: Ceratiaceae
Especie: Ceratium tripos (O.
F. Müller)
Nitzsch
Aumento: 400X
Microscopio invertido con
contraste de fases.
Abundancia: Abundante, en
algunas muestras.
No aislada.
Marina Fuente: Proyecto Fodecyt 43-
2007

86
Fotografía 41
Phylum: Dinophyta
Clase: Dinophyceae
Orden: Peridiniales
Familia:
Protoperidiniaceae
Especie:
Protoperidinium sp.
Aumento: 400X
Microscopio invertido
con contraste de fases.
Abundancia: Rara, en
pocas muestras.
No aislada.
Marina Fuente: Proyecto
Fodecyt 43-2007
IV.4.1.5 Otros
Fotografía 42
Phylum: Ochrophyta
Clase: Eustigmatophyceae
Orden: No asignado
Familia: Monodopsidaceae
Especie: Nannochloropsis sp.
Aumento: 1000X
Microscopio óptico
convencional.
Abundancia: Rara, en pocas
muestras.
No aislada.
Marina

87
PARTE V
V.1 Informe Financiero

88
V.3 Cronograma de Actividades
OBJETIVOS ACTIVIDADES RESULTADOS
En
ero
Feb
rero
Ma
rzo
Ab
ril
Ma
yo
Ju
nio
Ju
lio
Ag
ost
o
Sep
tiem
bre
Oct
ub
re
No
vie
mb
re
Dic
iem
bre
GENERAL:
Identificar las algas marinas de la costa atlántica guatemalteca y
determinar la presencia y concentración de aceites esenciales de
estas.
ESPECIFICO:
Identificar
taxonómicamente
las algas
(microscópicas y
macroscópicas)
predominantes.
Colecta muestras de
macroalgas y
microalgas en 5
visitas.
Obtención de
muestras de Algas
microscópicas y
macroscópicas.
Identificación
taxonómica de algas
marinas del Caribe
guatemalteco
Primera
caracterización de las
algas más abundantes
en el Caribe
guatemalteco
Determinación de
las algas más
comunes de los dos
grupos (microalgas
y macroalgas)
ESPECIFICO:
Establecer el
método de cultivo
exsitu para la
reproducción de
las algas marinas
predominantes.
Establecimiento y
monitoreo de
acuarios para la
reproducción de las
algas más comunes.
Establecimiento de
método de cultivo
para microalgas y
macroalgas del Caribe
guatemalteco
ESPECIFICO:
Comparar la
presencia y
concentración de
aceites en los
diferentes grupos
de algas
macroscópicas y
microscópicas del
Caribe
guatemalteco
Extracción,
saponificación y
determinación de
aceites de cada una
de las especies
reproducidas.
Establecer método
para la determinación
del porcentaje de
aceites para
microalgas y
macroalgas de la costa
Caribe guatemalteca.
Proponer
recomendaciones
para el cultivo de la
especie más
abundante.
Determinación de la
especie de alga
marina apta para la
reproducción y
potencial materia para
la producción de
Biodiesel
Divulgación

89