antiproliferative effects of nerium oleander stem and ... · antiproliferative effects of nerium...
TRANSCRIPT

Send Orders for Reprints to [email protected]
Letters in Drug Design & Discovery, 2017, 14, 000-000 1
RESEARCH ARTICLE
1570-1808/17 $58.00+.00 ©2017 Bentham Science Publishers
Antiproliferative Effects of Nerium oleander Stem and Mitotic Arrest Induced by Cardenolide Odoroside B on NCI-H460 Cancer Cells
Kehkashan Arshad Qamara,*, Ahsana Dar Farooqa,b, Bina S. Siddiquia, Nurul Kabirb, Naseema Khatoona, Shakil Ahmedc, Shaista Erumc and Sabira Beguma
aH.E.J. Research Institute of Chemistry, bDr. Panjwani Center for Molecular Medicine and Drug Research, cIndustrial Analytical Center, International Center for Chemical and Biological Sciences, University of Karachi, Karachi 75270, Pakistan
Abstract: Background: Nerium oleander extract preparations have been used in the Arab folk-medicine for the treatment of solid tumors.
Objective: In the current investigation, bioassay-guided fractionation of N. oleander stem methano-lic extract was performed to identify the active compound(s) responsible for its antiproliferative activity and the mechanism of action of the active compounds was explored.
Methods: The methanolic extract, fractions and sub-fractions were screened against four human cancer cell lines: HT-144, MCF-7, NCI-H460 and SF-268 using sulforhodamine B assay. The ef-fects of the active compounds on the cytoskeleton and nuclei of NCI-H460 cells were studied using immunofluorescence microscopy.
Results: The more active petroleum ether insoluble sub-fraction led to the isolation of five pure compounds viz adynergenin, adynerin, hemidesmin-2, odoroside A and odoroside B. Odoroside A was the most potent compound with GI50: 0.04 and LC50: 0.74 µM against NCI-H460 cell line, while odoroside B demonstrated moderate growth inhibition and cytotoxicity (GI50: 6.7; LC50: 54 µM). After 24 hours’ treatment with odoroside B (50 µM) abnormal mitotic spindles were ob-served, while > 90% mitotic cells were arrested in the prophase stage.
Conclusion: N. oleander stem possesses significant antiproliferative effects against the aforemen-tioned cell lines and the cardenolide odoroside B induces mitotic arrest of NCI-H460 cells in the prophase stage.
Keywords: Nerium oleander, antiproliferative, odoroside B, cytoskeleton, immunofluorescence, mitotic arrest.
1. INTRODUCTION Nerium oleander (Apocynaceae family) is an evergreen tree distributed in Indo-Pakistan subcontinent, tropical and subtropical regions, and the Mediterranean. It has been used in the folklore medicine for the treatment of solid tumors and skin diseases like corns, eczema, herpes, psoriasis, ringworm infections, scabies, sores and warts [1-3]. Ingestion of parts of this plant causes toxicity due to the presence of cardiac glycosides such as oleandrin and oleandrigenin [4]. Many cardiac glycosides and pregnanes e.g. neridienone A and neridienone B have been isolated from the stem and twigs of N. oleander [5, 6]. Several of these compounds have demonstrated anticancer effects, e.g. neridienone A exhibited
*Address correspondence to this author at the H.E.J. Research Institute of Chemistry, University of Karachi, Karachi 75270, Pakistan; Tel: +92 21 34819017; Fax: +92 21 34819018; E-mail: [email protected]
IC50 of 0.68 and 2.5 µg/ml against human malignant fibro-blast and liver tumor cell lines, respectively [5]; the com-pound 3β-O-(β-D-diginosyl)-14-hydroxy-5β,14β-card-20(22)-enolide also demonstrated significant antiproliferative effects (IC50: 0.18 and 1.5 µM) against the aforementioned human cancer cell lines [7]. It has been recently demonstrated that the cardenolides from the Apocynaceae family including the genus Nerium exhibit the capability to regulate cancer cell survival and death through multiple signaling pathways [8].
The proteins involved in cell division such as microtu-bules are important targets for anticancer drugs [9]. Microtu-bules form the bipolar mitotic-spindle apparatus at the time of cell division, which is essential for the equal separation and distribution of chromosomes to the daughter cells [10]. This process requires a dynamic balance between polymeri-zation and depolymerization processes of microtubules. The treatment of cancer cells with anti-microtubule drugs dis-
A R T I C L E H I S T O R Y
Received: February 09, 2017 Revised: July 07, 2017 Accepted: July 11, 2017 DOI: 10.2174/1570180814666170727142145

2 Letters in Drug Design & Discovery, 2017, Vol. 14, No. 0 Qamar et al.
turbs this balance inducing interruption of mitosis, thus lead-ing to mitotic arrest and cell death [11].
Most anticancer drugs lack tumor specificity and kill rapidly proliferating tumor cells as well as normal cells of bone marrow, gastrointestinal mucosa and hair leading to severe side effects (myelosuppression, mouth ulcers, diar-rhea, alopecia etc.). Moreover, tumor cells quickly acquire resistance to these agents. Therefore, search for novel and safer anticancer agents is an on-going process. Currently, there is a revival of scientific interest in medicinal plants for lead compounds and/or the generation of their semi-synthetic derivatives. Despite reports on the anticancer potential of N. oleander there is no systematic bioassay-guided anticancer activity study on its stem. Keeping this in mind, in the pre-sent study N. oleander stem methanolic extract, fractions and sub-fractions were evaluated against four human cancer cell lines: HT-144 (melanoma, skin), MCF-7 (adenocarcinoma, breast), NCI-H460 (non-small cell lung cancer) and SF-268 (astrocytoma, CNS). Using immunofluorescence micros-copy, the effects of the active compounds derived from N. oleander stem were studied on the cytoskeleton and nuclei of NCI-H460 cells.
2. MATERIALS AND METHODS
2.1. Materials and Stock Solutions Following cell culture grade chemicals were purchased from respective companies given in parentheses: Cy3-labeled β tubulin clone TUB 2.1 monoclonal antibody, 4, 6-diamidino-2-phenylindole (DAPI), dimethyl sulfoxide (DMSO), gentamycin sulphate, L-glutamine penicillin-streptomycin solution, poly-L-lysine, RPMI-1640, sulforho-damine B (SRB), trichloroacetic acid, trypan blue, trypsin-EDTA (Sigma, St. Louis, Mo, USA). Alexa 488 phalloidin, fetal bovine serum (heat inactivated) and zymed clearmount mounting medium (Invitrogen, Grand Island, NY 14072, USA). Amphotericin B, doxorubicin, paclitaxel and vin-blastine (MP Biochemicals, Solon, Ohio 44139, USA); tris base (Aldrich, Stienheim, Germany); triton-X-100 (Wako Pure Chemical Industries Ltd., Japan); acetic acid (Riedel-de-Haen, Sigma- Aldrich Gmbh D-30926, Germany); and for-maldehyde 4% (Carl Roth, Gmbh Co., Schoemperlenstraβe 76185, Karlsruhe, Germany).
The stock solutions of N. oleander stem methanolic ex-tract (40 mg /ml), fractions and sub-fractions (20 mg /ml), and pure compounds (20 mM) were prepared in DMSO while doxorubicin stock solution (20 mM) in sterile water. All the stock solutions were stored at -20°C and further dilu-tions were freshly prepared in RPMI-1640 containing gen-tamicin (50 µg/ml) to control bacterial contamination.
2.2. Extraction and Isolation Uncrushed stem of Nerium oleander (6 kg) was repeat-edly extracted with MeOH (3x) at room temperature. The concentrated syrupy residue obtained on removal of the sol-vent from the combined extract was shaken out with EtOAc and H2O. The EtOAc layer was extracted with 4% aqueous Na2CO3 solution to separate the acidic from the neutral frac-tion. The EtOAc layer containing the neutral fraction was
washed, dried over anhydrous Na2SO4, charcoaled and con-centrated to a thick syrup. It was divided into petroleum ether soluble and insoluble fractions. The petroleum ether insoluble fraction was subjected to column chromatography (petroleum ether, petroleum ether -EtOAc in order of in-creasing polarity). The fractions were combined on the basis of their thin layer chromatography (TLC) to yield 15 frac-tions. Fraction No. 2 (petroleum ether-EtOAc, 8.5:1.5 elu-ate), 5 (petroleum ether-EtOAc, 8:2 eluate), 6 (petroleum ether-EtOAc, 7.5:2.5 eluate), 8 (petroleum ether-EtOAc, 7:3 eluate) and 10 (petroleum ether-EtOAc, 7:3 eluate) were almost pure and were recrystallized from methanol to yield pure compounds identified as odoroside-A (15 mg) [12], adynerigenin (12 mg) [13], adynerin (30 mg) [13], odor-oside-B (10 mg ) [12] and hemidesmin-2 (20 mg) [14], re-spectively (Fig. 1).
2.3. Cell Lines HT-144 (melanoma, skin), MCF-7 (adenocarcinoma, breast), NCI-H460 (non-small cell carcinoma, lung) and SF-268 (anaplastic astrocytoma, CNS) human cancer cell lines were maintained in a humid atmosphere with 5% CO2 at 37°C. The RPMI-1640 medium was supplemented with heat inactivated fetal bovine serum (10%), glutamine (2 mM), penicillin (100 units/ml), streptomycin (100 µg/ml) and am-photericin-B (2.5 µg/ml). The cell concentration and viability were determined by using a hemocytometer.
2.4. Growth Inhibitory and Cytotoxicity Assay In vitro growth inhibition and cytotoxicity were deter-mined by employing sulforhodamine B (SRB) assay [15, 16]. About 1×104/well MCF-7 and NCI-H460 cells were added of 96-well plates while SF-268 and HT-144 cells were added at the density of 1.5×104 and 2×104 cells/well, respec-tively. After 24 hours’ incubation, different doses of N. ole-ander methanolic extract, fractions, sub-fractions, pure com-pounds and doxorubicin were added. After 48 hours, cells were fixed with ice-cold trichloroacetic acid (50 µl, 50%) and left for overnight air drying. The cells were stained with SRB dye (100 µl/well). Next day, the bound SRB dye was solubilized in Tris-base solution (10 mM) and optical density measurements were taken at 545 nm in a plate reader. By plotting the percentage of cell growth or cell killing versus the drug concentrations [16], drug concentrations causing growth inhibition and killing of 50% cells i.e. GI50 and LC50 values were calculated graphically.
2.5. Immunofluorescence Microscopy
Alexa Flour 488 phalloidin, Cy3-labeled β tubulin anti-body and DAPI were used to visualize F-actin filaments, microtubules and nuclei, respectively [17-19]. Lab-Tek II 8-chambered slides (Nalgene Nunc, USA) were coated with poly-L-lysine to allow firm attachment. About 1x 104 NCI-H460 cells/well were added and incubated overnight at 37°C. Next day, the culture medium containing pure com-pounds (1, 5, 10, 50 and 100 µM), or corresponding vehicle (0.1% DMSO) was added. After incubation for further 24 and/or 48 hours, the cells were fixed with formaldehyde (4%) and the slides washed with freshly prepared phosphate buffered saline (PBS). Permeabilization of the cells was car-

Antiproliferative Effects of Nerium oleander Stem and Mitotic Arrest Letters in Drug Design & Discovery, 2017, Vol. 14, No. 0 3
ried out with 1% Triton X-100 in PBS. The cells were incu-bated initially with Alexa Fluor 488 phalloidin to stain F-actin filaments, then with Cy3-labeled tubulin antibody for microtubules and lastly with DAPI. After extensive washing, the slides were mounted, cover-slipped and viewed under Nikon’s 90i epifluorescence microscope. Nikon DXM1200C cooled CCD camera was used to capture images with DAPI, FITC and TxRed filter cubes. Real-time deconvolution was performed on images by using NIS-elements 3.0 software (AR version) and images were processed with Adobe Photo-shop [20].
2.6. Mitotic Index Assay The cells were identified to be in either interphase or mitotic stage or undergoing cell death. Control interphase
nuclei were similar in size, smooth, round or oval shaped. The dead cells showed condensed and/or fragmented nuclei. The mitotic cells exhibited condensed chromosomes without nuclear membrane. Prophase stage was characterized by nu-clear envelope breakdown and chromosome condensation. In the metaphase stage of the cell cycle, replicated chromo-somes were aligned at the spindle equator. The anaphase
OO
O O
OH
H
HO
CH3
OCH3
H
Odoroside-A OO
O O
OH
H
HO
CH3
OCH3
H
Odoroside-B
O
HHO
O O
OO
O O
O
H
HO
CH3
OCH3
H
OO
O
O
H3CO
HO OCH3
HOH2C
Adynerigenin
Adynerin
Hemidesmin-2
Fig. (1). Structures of compounds.
4 Letters in Drug Design & Discovery, 2017, Vol. 14, No. 0 Qamar et al.
stage began when the chromosomes segregated towards the spindle poles to form two new daughter cells [21]. The telo-phase stage was evident when the cleavage furrow appeared till the completion of cytokinesis [22]. The mitotic cells in prophase, metaphase, anaphase, or telophase were identified, and their percentages determined for the control cells and after treatment with odoroside B (50 µM) for 24 and 48 hours. The percent mitotic index was calculated by dividing the number of cells in mitosis by the total number of cells and multiplying the result with 100.
2.7. Statistical Analysis Statistical software SPSS 16 was used to analyze the data which was expressed as mean ± SEM. ANOVA (Analysis of variance) followed by LSD (least significance difference) and Duncan’s multiple comparison tests were used for statis-tical comparisons between the control and the treated groups and between the various treatments. p values of less than 0.05 were considered statistically significant (*p < 0.05; **p < 0.01; ***p < 0.001, versus control).
3. RESULTS 3.1. Cytotoxic Effects of Extract, Fractions and Sub-fractions
The methanolic extract, fractions and sub-fractions ex-hibited significantly maximum cytotoxic effects against NCI-H460 lung cancer cell line as reflected in their lowest LC50 values (Table 1). The methanolic extract and its ethyl acetate soluble fraction showed similar cytotoxicity against MCF-7 cell line with LC50 of ~ 6 µg /ml. The petroleum ether insoluble sub-fraction demonstrated maximum cyto-toxic activity against all the cell lines. The aqueous fraction exhibited minimum cytotoxicity against MCF-7 cell line (LC50 > 100 µg/ml).
3.2. Growth Inhibition and Cytotoxicity of Extract, Frac-tion, Sub-fraction and Pure Compounds
The methanolic extract and the ethyl acetate soluble frac-tion exhibited similar GI50 of ~ 0.3 µg/ml to the pure com-pound adynerin, while the petroleum ether insoluble sub-fraction demonstrated comparable growth inhibition to odor-oside A and the reference compound doxorubicin (Table 2). The GI50 potency order was odoroside A = doxorubicin = PEIS > EA = adynerin = ME > hemidesmin-2 > odoroside B. The petroleum ether insoluble sub-fraction demonstrated ~2-fold greater cytotoxic effects than odoroside A and doxoru-bicin (LC50: 0.16 µg/ml). The potency order for LC50 was PEIS > odoroside A = doxorubicin > EA = ME > adynerin = hemidesmin-2 > odoroside B.
3.3. Immunofluorescence Microscopy
Immunocytochemistry was employed to explore the mechanism of antiproliferative effects of the pure com-pounds isolated from N. oleander stem against NCI-H460 cells. F-actin filaments, microtubules and the nuclei were visualized using Alexa 488 phalloidin, Cy3-labeled tubulin antibody and DAPI, respectively. The effects of these com-pounds were compared with control, and those induced by doxorubicin, paclitaxel and vinblastine. Odoroside A in-duced cell death at the dose of 1 µM, while odoroside B (50 µM) exhibited remarkable changes in the morphology and percent mitotic stages, hence further studies were conducted on the latter compound.
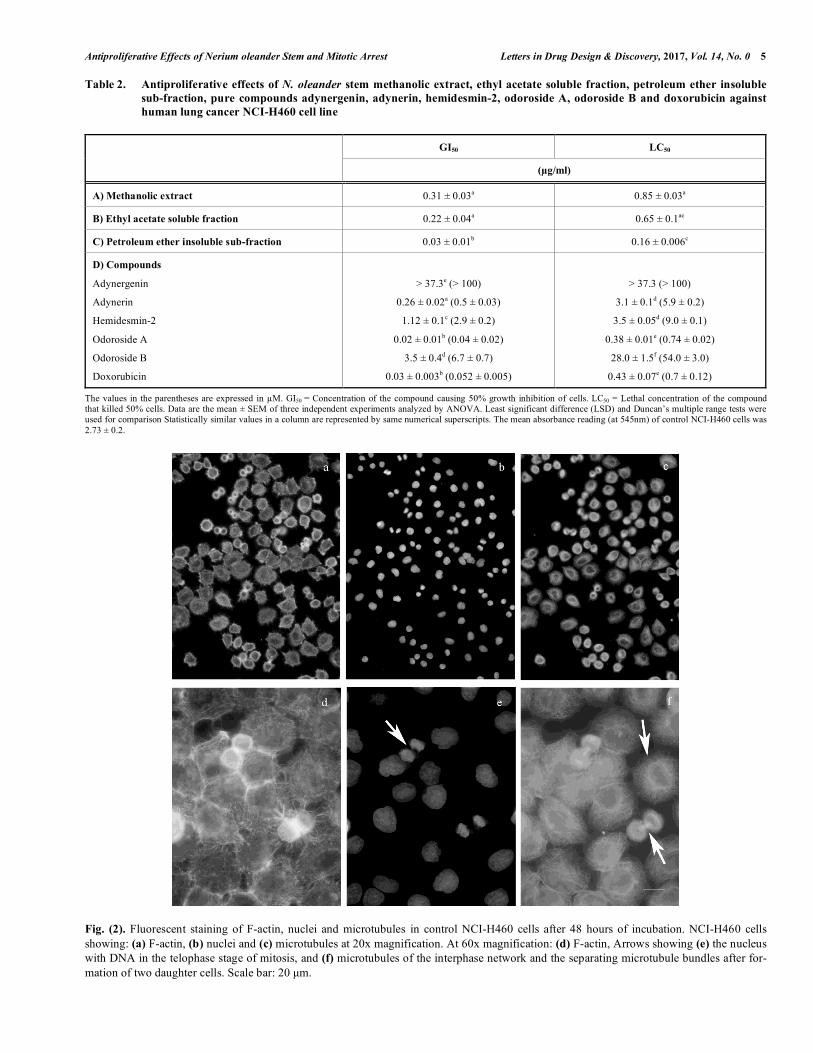
3.3.1. Cytoskeleton and Nuclear Morphology of Control NCI-H460 Cells The control NCI-H460 cells were polygonal in shape but rounded at the time of cell division with fine, pointed and thread-like F-actin processes (Fig. 2a, d). The interphase nuclei were smooth, round or oval with 2-3 invaginations
Table 1. Cytotoxic effects of N. oleander stem methanolic extract, fractions and sub-fractions against HT-144, MCF-7, NCI-H460 and SF-268 human cancer cell lines.
Cell Lines
HT-144 MCF-7 NCI-H460 SF-268
(LC50: µg/ml)
A) Extract
Methanolic
1.8 ± 0.14 a,1
6.0 ± 0.6 a,2
0.85 ± 0.03a,1
7.5 ± 1.3 a,2
B) Fractions
Aqueous
Ethyl acetate soluble
81.3 ± 5.7b,1
0.94 ± 0.2c,1
> 100
6.2 ± 1b,2
17.1 ± 0.9b,2
0.65 ± 0.1a,1
44.3 ± 0.7 b,3
5.1 ± 0.8a,2
C) Sub-fractions
Petroleum ether soluble
Petroleum ether insoluble
94.7 ± 3.6 d,1
0.72 ± 0.04c,1,2
31.5 ± 0.5b,2
1.5 ± 0.4c,1
23.0 ± 0.5c,2
0.16 ± 0.006d,2
61.0 ± 1.4c,3
0.37 ± 0.12d,2
Data are the mean ± SEM of three independent experiments analyzed by ANOVA. For comparison of different test agents, least significant difference (LSD) and Duncan’s multiple range tests were used. LC50 = Lethal concentration of the extract/fraction/sub-fraction that killed 50% cells. The mean absorbance reading (at 545nm) of control cells of the afore-mentioned cell lines was 2.8 ± 0.12. In columns statistically different values are represented by dissimilar alphabetical superscripts. In rows statistically different values are repre-sented by dissimilar numerical superscripts.
Antiproliferative Effects of Nerium oleander Stem and Mitotic Arrest Letters in Drug Design & Discovery, 2017, Vol. 14, No. 0 5
Table 2. Antiproliferative effects of N. oleander stem methanolic extract, ethyl acetate soluble fraction, petroleum ether insoluble sub-fraction, pure compounds adynergenin, adynerin, hemidesmin-2, odoroside A, odoroside B and doxorubicin against human lung cancer NCI-H460 cell line
GI50 LC50
(µg/ml)
A) Methanolic extract 0.31 ± 0.03a 0.85 ± 0.03a
B) Ethyl acetate soluble fraction 0.22 ± 0.04a 0.65 ± 0.1ae
C) Petroleum ether insoluble sub-fraction 0.03 ± 0.01b 0.16 ± 0.006c
D) Compounds Adynergenin
Adynerin
Hemidesmin-2
Odoroside A
Odoroside B
Doxorubicin
> 37.3e (> 100)
0.26 ± 0.02a (0.5 ± 0.03)
1.12 ± 0.1c (2.9 ± 0.2)
0.02 ± 0.01b (0.04 ± 0.02)
3.5 ± 0.4d (6.7 ± 0.7)
0.03 ± 0.003b (0.052 ± 0.005)
> 37.3 (> 100)
3.1 ± 0.1d (5.9 ± 0.2)
3.5 ± 0.05d (9.0 ± 0.1)
0.38 ± 0.01e (0.74 ± 0.02)
28.0 ± 1.5f (54.0 ± 3.0)
0.43 ± 0.07e (0.7 ± 0.12)
The values in the parentheses are expressed in µM. GI50 = Concentration of the compound causing 50% growth inhibition of cells. LC50 = Lethal concentration of the compound that killed 50% cells. Data are the mean ± SEM of three independent experiments analyzed by ANOVA. Least significant difference (LSD) and Duncan’s multiple range tests were used for comparison Statistically similar values in a column are represented by same numerical superscripts. The mean absorbance reading (at 545nm) of control NCI-H460 cells was 2.73 ± 0.2.
Fig. (2). Fluorescent staining of F-actin, nuclei and microtubules in control NCI-H460 cells after 48 hours of incubation. NCI-H460 cells showing: (a) F-actin, (b) nuclei and (c) microtubules at 20x magnification. At 60x magnification: (d) F-actin, Arrows showing (e) the nucleus with DNA in the telophase stage of mitosis, and (f) microtubules of the interphase network and the separating microtubule bundles after for-mation of two daughter cells. Scale bar: 20 µm.
6 Letters in Drug Design & Discovery, 2017, Vol. 14, No. 0 Qamar et al.
and 2-4 nucleoli. The condensed chromatin visible in ana-phase mitotic stage is marked by an arrow (Fig. 2e). Thread-like, fine microtubules were arranged in a circular ring around the nucleus forming a regular network in the periphery (Fig. 2f). Highly fluorescent bundles of microtubules forming the mitotic spindles and connecting the two daughter cells at the end of cell division were clearly visible (Fig. 2f).
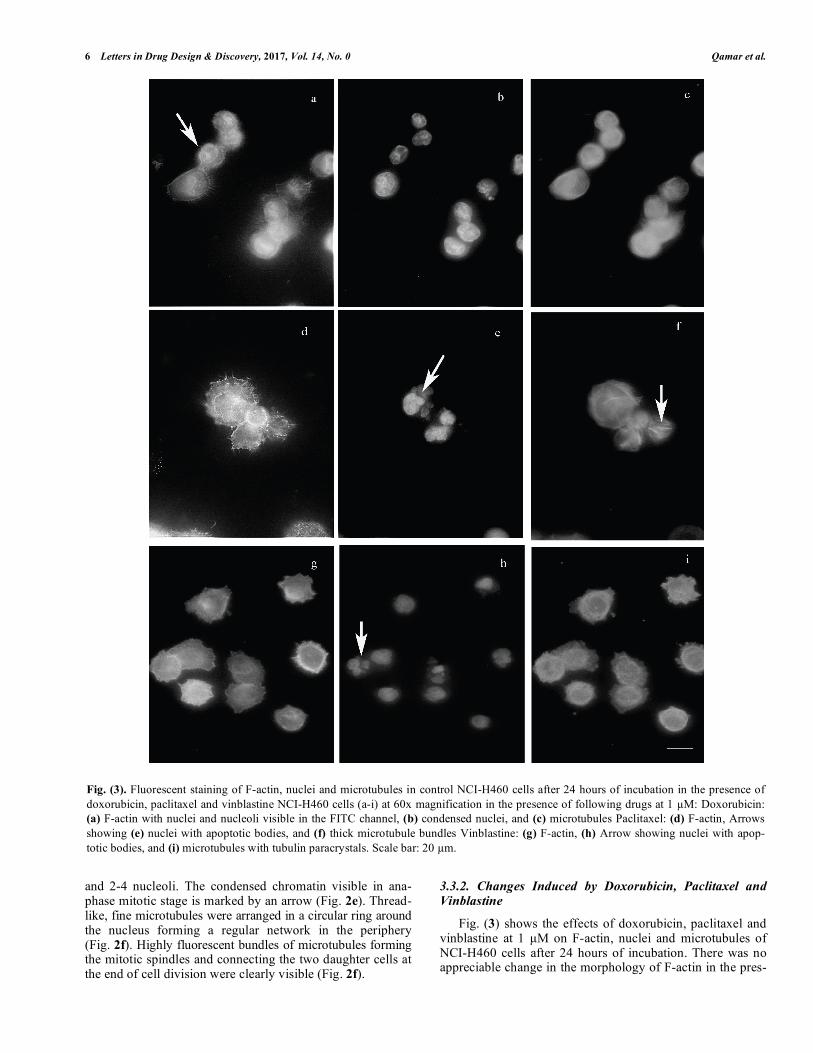
3.3.2. Changes Induced by Doxorubicin, Paclitaxel and Vinblastine Fig. (3) shows the effects of doxorubicin, paclitaxel and vinblastine at 1 µM on F-actin, nuclei and microtubules of NCI-H460 cells after 24 hours of incubation. There was no appreciable change in the morphology of F-actin in the pres-
Fig. (3). Fluorescent staining of F-actin, nuclei and microtubules in control NCI-H460 cells after 24 hours of incubation in the presence of doxorubicin, paclitaxel and vinblastine NCI-H460 cells (a-i) at 60x magnification in the presence of following drugs at 1 µM: Doxorubicin: (a) F-actin with nuclei and nucleoli visible in the FITC channel, (b) condensed nuclei, and (c) microtubules Paclitaxel: (d) F-actin, Arrows showing (e) nuclei with apoptotic bodies, and (f) thick microtubule bundles Vinblastine: (g) F-actin, (h) Arrow showing nuclei with apop-totic bodies, and (i) microtubules with tubulin paracrystals. Scale bar: 20 µm.

Antiproliferative Effects of Nerium oleander Stem and Mitotic Arrest Letters in Drug Design & Discovery, 2017, Vol. 14, No. 0 7
ence of doxorubicin (Fig. 3a). However, since doxorubicin is a fluorescent drug and binds to DNA, hence nuclei were visible in the FITC channel also (Fig. 3a). Few nuclei ap-peared irregular, condensed and fragmented (Fig. 3b). Moreover, the fine network of microtubules, as seen in con-trol cells, was not visible (Fig. 3c), Tubulin stabilizing agent paclitaxel induced long, thick microtubule bundle formation
(Fig. 3f), whereas vinblastine caused tubulin paracrystals formation (Fig. 3i). After treatment with both aforemen-tioned drugs, no detectable change in F-actin was observed (Fig. 3d, g). Most of the nuclei treated by either paclitaxel or vinblastine appeared fragmented and apoptotic bodies were clearly visible as marked by arrows (Fig. 3e, h).
Fig. (4). Fluorescent staining of F-actin, nuclei and microtubules in NCI-H460 cells after 24 hours of incubation with odoroside B NCI-H460 cells (a-c) at 20x, (d-i) at 60x magnification in the presence of odoroside B: At 50 µM: (a) F-actin, Arrows showing (b) many nuclei in pro-phase stage of mitosis, and (c) microtubules (d) F-actin with rounding of cell at the time of cell division, Arrows showing (e) two nuclei in the prophase stage of mitosis, and (f) mitotic spindle. At 100 µM: (g) F-actin, (h) nuclei, and (i) Arrow showing irregular microtubule bun-dles. Scale bar: 20 µm.

8 Letters in Drug Design & Discovery, 2017, Vol. 14, No. 0 Qamar et al.
3.3.2. Changes in Cytoskeletal and Nuclear Morphology Induced by Odoroside B
The fluorescence microscopy showed no appreciable change in the morphology of F-actin, interphase nuclei and microtubules after 24 and 48 hours’ treatment with odoroside B (50 µM) as compared to the aforementioned control (Fig. 4d, e, f; 5a, b, c). After 24 hours’ incubation with odoroside B (50 µM), an increase in the mitotic cells in the prophase stage could be clearly seen as marked by an arrow at 20x magnification (Fig. 4b). When the cells were incubated with higher concentration of odoroside B (100 µM) for 24 hours, the microtubules appeared as thickened, irregular, highly disorganized and entangled fibers (Fig. 4i). After 48 hours, these aforementioned changes were more pronounced (Fig. 5f). Additionally, the treated cells lost their processes and became rounded (Fig. 5d). Nuclear fragmentation was visible and apoptotic bodies were seen as marked by an arrow (Fig. 5e).
3.3.3. Effects of Odoroside B on Cell Number and Percent Mitotic Index
The average number of control cells counted in single field at 20x magnification after 24 hours (240 ± 15) was sig-nificantly reduced (67%) after treatment with odoroside B (50 µM) (Fig. 6a). A significant increase in the mean percent mitotic index of control (5.4) vs. treated cells (9.7) was observed (Fig. 6b). The corresponding mean percentages of control mitotic cells after 24 hours’ incubation were about 29, 18 and 53 in prophase, metaphase and anaphase/ telophase stages. However, 24 hours of odoroside B (50 µM) treatment induced a significant increase in prophase stage (96.8%) accompanied by a significant decline in metaphase (1.6%) and anaphase/telophase stages (1.6%) (Fig. 6c). A significant decline was observed in the total number of the cells after incubation for 48 hours (p < 0.001) (Fig. 6a), however, the percent mitotic index was slightly reduced from 4.83 in the control to 3.1 in the treated cells (Fig. 6b).
Fig. (5). Fluorescent staining of F-actin, nuclei and microtubules in NCI-H460 cells after 48 hours of incubation with odoroside B. Arrows showing NCI-H460 cells (a-f) at 60x magnification in the presence of odoroside B: At 50 µM: (a) F-actin with the blebs, (b) nuclei in prophase stage of mitosis, and (c) microtubules. At 100 µM: (d) F-actin, (e) fragmented nuclei, and (f) irregular microtubule bundles. Scale bar: 20 µm.

Antiproliferative Effects of Nerium oleander Stem and Mitotic Arrest Letters in Drug Design & Discovery, 2017, Vol. 14, No. 0 9
4. DISCUSSION AND CONCLUSIONS The search for substances of plant origin with anticancer properties have prompted studies on all parts of N. oleander including roots [20]. To our knowledge, the present study provides the first evidence of the antiproliferative activity of N. oleander stem methanolic extract, fractions and sub-fractions against the aforementioned cell lines. Moreover, this is the first report of the anticancer activity of the pure compounds: adynerin, hemidesmin-2, odoroside A and odor-oside B derived from N. oleander stem against NCI-H460 cell line. The current investigation reports that N. oleander stem methanolic extract, fractions and sub-fractions exhib-ited potent cytotoxic effects against all the four human can-cer cell lines tested, however, they were more cytotoxic against NCI-H460 cells. In the present study, odoroside A (5β) (GI50: 0.04; LC50: 0.74 µM) was the most potent com-pound exhibiting comparable activity to the reference com-pound doxorubicin against NCI-H460 cell line, while its stereoisomer odoroside B (5α) was ~70-fold less cytotoxic (LC50: 54 µM) emphasizing the importance of stereochemis-try for biological activities.
The petroleum ether insoluble sub-fraction was most cy-totoxic (LC50: 0.16 µg/ml) demonstrating ~2-fold better cell killing activity than the standard anticancer drug doxorubicin against NCI-H460 cell line. The additive or synergistic ef-fects of compounds residing in the sub-fraction could have been responsible for its greater cytotoxicity as seen in tradi-tional Chinese medicine (TCM), where composite and com-plex remedies are used which possibly act in a synergistic fashion to increase therapeutic effects and reduce adverse side effects on healthy tissues [24].
It is well known that certain drugs like paclitaxel and vinblastine exert their anti-tumor actions by disrupting the dynamics of microtubule assembly, thereby disturbing the formation and function of the mitotic spindle apparatus in dividing cells, arresting cells in mitosis and induction of cell death [25]. It is essential that all the chromosomes are at-tached to the mitotic spindle before the onset of anaphase; this process is strictly controlled by the mitotic or the spindle assembly checkpoint [26]. The microtubule-disrupting agents possibly arrest cells in mitosis by triggering the mi-totic checkpoint that ensures proper attachment of chromo-
Fig. (6). Effects of odoroside B on the number of cells, percent mitotic index and mitotic stages of NCI-H460 cells. Effects of odoroside B (50 µM, ) after 24 and 48 hours of incubation in NCI-H460 cells on: (a) Number of cells, (b) Percent mitotic index, and (c) Percent mitotic stages after 24 hours of incubation. Control (□), n = 3. * = p < 0.05, ** = p < 0.01, *** = p < 0.001.

10 Letters in Drug Design & Discovery, 2017, Vol. 14, No. 0 Qamar et al.
somes to the mitotic spindle before the onset of anaphase [27]. Paclitaxel induces mitotic arrest by promoting stabiliza-tion of microtubules [28], whereas vinblastine acts by de-polymerizing the microtubules [29]. In the present investiga-tion, odoroside B induced mitotic arrest with ~90% of NCI-H460 mitotic cells in the prophase stage along with a sub-stantial decline (90%) in the metaphase and anaphase/ telophase stages indicating that it probably interferes with either the function or the structure of the microtubules or both. Dose and time dependent changes were seen at higher concentration (100 µM); the microtubules appeared as thick, irregular and disorganized fibers. Moreover, after 48 hours of treatment, cell rounding and nuclear fragmentation indicated cell death.
It is established that the addition of paclitaxel stabilizes the microtubules and increases the polymerization of tubulin, while the reverse is seen after the addition of vinblastine [30]. Western blotting can be employed to determine whether odoroside B induces polymerization or depolymeri-zation of tubulin in NCI-H460 cells, as demonstrated for 1H-[1, 2, 4] oxadiazolo[4,3-a]quinoxalin-1-one that prevents microtubule polymerization in HeLa cells [31].
It is being suggested that the aforementioned effects of odoroside B on cell cycle progression be substantiated by DNA flow cytometry which can also evaluate the distribu-tion of actively dividing cells in the cell cycle and the per-centage of apoptotic and non- apoptotic cells in each phase of the cell cycle [32]. Moreover, to confirm the mode of cell death induced by odoroside B, measurement of mitochon-drial membrane potential and release of cytochrome c pro-viding an early indication of the initiation of cellular apopto-sis should be carried out [33]. When it is confirmed that the cell death induced by odoroside B is due to the apoptotic process, then Western blot analysis could also be done to examine the involvement of specific caspases and/or the cleavage of poly (ADP-ribose) polymerase [34].
These results indicate that N. oleander stem methanolic extract, fractions and sub-fractions possess significant anti-proliferative effects against HT-144, MCF-7, NCI-H460 and SF-268 human cancer cell lines justifying the traditional use of this plant. The pure compounds (adynerin, hemidesmin-2, odoroside A and odoroside B) were cytotoxic against NCI-H460 cell line. The importance of stereochemistry for bio-logical activities was evident by 70-fold greater cytotoxicity exhibited by odoroside A (5β) against NCI-H460 cell line than its isomer odoroside B (5α). It is also being suggested that the antiproliferative effects of odoroside B were mainly due to its interaction with microtubules and induction of cell death. Therefore, it is concluded that N. oleander is a poten-tially valuable candidate for cancer chemotherapy.
CONSENT FOR PUBLICATION
Not applicable.
CONFLICT OF INTEREST There is no actual or potential conflict of interest includ-ing any financial, personal or other relationships with other people or organizations that could have inappropriately in-fluenced, or be perceived to influence our work.
ACKNOWLEDGEMENTS We are thankful to National Cancer Institute, Frederick, Md. USA, for kindly providing us the cell lines under Mate-rial Transfer Agreement.
REFERENCES [1] Duke, J.A. Handbook of Medicinal Herbs, CRC Press LLC, 2000,
N.W. Corporate Blvd., Boca Raton, Florida 33431, USA. 1985. [2] Baytar, I.A. T. M. A. An Encyclopaedia of Medicine and Nutrition.
Dar Al-Kutub, Lebanon, 1992. [3] Yesilada, E.; Honda, G.; Sezik, E.; Tabata, M.; Fujita, T.; Tanaka,
T.; Takeda, Y.; Takaishi, Y. Traditional medicine in Turkey. V. Folk medicine in the inner Taurus Mountains. J. Ethnopharmacol., 1995, 46, 133-152.
[4] Langford, S.D.; Boor, P.J. Oleander toxicity: an examination of human and animal toxic exposures. Toxicology, 1996, 109, 1-13.
[5] Bai, L.; Wang, L.; Zhao, M.; Toki, A.; Hasegawa, T.; Ogura, H.; Kataoka, T.; Hirose, K.; Sakai, J. Bai, J.; Ando, M. Bioactive Preg-nanes from Nerium oleander. J. Nat. Prod., 2007, 70, 14-18.
[6] Zhao, M.; Bai, L.; Wang, L.; Toki, A.; Hasegawa, T.; Kikuchi, M.; Abe, M.; Sakai, J.; Hasegawa, R.; Bai, Y.; Mitsui, T.; Ogura, H.; Kataoka, T.; Oka, S.; Tsushima, H.; Kiuchi, M.; Hirose, K.; Tomida, A.; Tsuruo, T.; Ando, M. Bioactive cardenolides from the stems and twigs of Nerium oleander. J. Nat. Prod., 2007, 70, 1098-1103.
[7] Zhao, M.; Bai, L.; Toki, A.; Hasegawa, R.; Sakai, J.; Hasegawa, T.; Ogura, H.; Kataoka, T.; Bai, Y.; Ando, M.; Hirose, K., Ando, M. The structure of a new cardenolide diglycoside and the biological activities of eleven cardenolide diglycosides from Nerium olean-der. Chem. And Pharm. Bull., 2011, 59, 371-7.
[8] Wen, S.; Chen, Y.; Lu, Y.; Wang, Y.; Ding, L.; Jiang, M. Carde-nolides from the Apocynaceae family and their anticancer activity. Fitoterapia, 2016, 112, 74–84.
[9] Jordan, M.A.; Thrower, D.; Wilson, L. Effects of vinblastine, podophyllotoxin and nocodazole on mitotic spindles: Implications for the role of microtubule dynamics in mitosis. Journal of Cell Science, 1992, 102, 401-416.
[10] Rusan, N.M.; Fagerstrom, C.J.; Yvon, A.M.; Wadsworth, P. Cell cycle-dependent changes in microtubule dynamics in living cells expressing green fluorescent protein-alpha tubulin. Mol. Biol. Cell, 2001, 12, 971-80.
[11] Pasquier, E.; Honor´e, S.; Braguer, D. Microtubule-targeting agents in angiogenesis: where do we stand? Drug Resist Updates, 2006, 9, 74-86.
[12] Ahmad, V.U.; Basha, A. Spectroscopic Data of Steroid Glycosides, Volume 4, 2047-2049. Springer-Verlag New York Inc. 2006.
[13] Ahmad, V.U.; Basha, A. Spectroscopic Data of Steroid Glycosides, Volume 4, 2038-2039. Springer-Verlag New York Inc. 2006.
[14] Das, P.C.; Joshi, P.C.; Mandal, S.; Das, A.; Chatterjee, A. New Coumarinolignoids from Hemidesmus indicus. Indian Journal of Chemistry, 1992, 31B, 342-345.
[15] Skehan, P.; Streng, R.; Scudiero, D.; Monks, A.; McMahon, J.; Vistica, D.; Warren, J.T.; Bokesch, H.; Kenney, S.; Boyd, M.R. New colorimetric cytotoxicity assay for anti-cancer drug screening. Journal of National Cancer Institute, 1990, 82, 1107-1112.
[16] Monks, A.; Scudiero, D.; Skehan, P.; Shoemaker, R. Feasibility of a high-flux anticancer drug screen using a diverse panel of cultured human tumor cell lines. Journal of National Cancer Institute, 1991, 11, 757-766.
[17] Schaefer, A.W.; Kabir, N.; Forscher, P. Filopodia and actin arcs guide the assembly and transport of two populations of microtu-bules with unique dynamic parameters in neuronal growth cones. Journal of Cell Biology, 2002, 158, 139-152.
[18] Kabir, N.; Schaefer, A.W.; Nakhost, A.; Sossin, W.S.; Forscher, P. Protein kinase C activation promotes microtubule advance in neu-ronal growth cones by increasing average microtubule growth life-times. Journal of Cell Biology, 2001, 152, 1033-1044.
[19] Forscher, P.; Smith, S.J. Actions of cytochalasins on the organiza-tion of actin filaments and microtubules in a neuronal growth cone. Journal of Cell Biology 1988, 07, 1505-1516.
[20] Qamar, K.A.; Dar, A.; Siddiqui, B.S.; Kabir, N.; Aslam, H.; Ah-med, S.; Erum, S.; ursolic acid on the cytoskeleton of MCF-7 hu-

Antiproliferative Effects of Nerium oleander Stem and Mitotic Arrest Letters in Drug Design & Discovery, 2017, Vol. 14, No. 0 11
man breast cancer cells. Letters in Drug Design & Discovery, 2010, 7, 726-736.
[21] McIntosh, J.; Grishchunk, E.; West, R. Chromosome-microtubule interactions during mitosis. Annu. Rev. Cell Dev. Biol., 2002, 18, 193-219.
[22] Cha, H., Dangi, S., Machmer, C.E., Shapiro, P., Inhibition of mixed-lineage kinase (MLK) activity during G2-phase disrupts mi-crotubule formation and mitotic progression in HeLa cells. Cellular signaling, 2006, 18, 93-104.
[23] Gunes, A.; Guler, E.; Un, R.N.; Demir, B.; Barlas, F.B.; Yavuz, M.; Coskunol, H.; Timur, S. Niosomes of Nerium oleander ex-tracts: In vitro assessment of bioactive nanovesicular structures. Journal of Drug Delivery, 2017, 37, 158-165.
[24] Efferth, T.; Li, P.C.H.; Konkimalla, V.S.B.; Kaina, B. From tradi-tional Chinese medicine to rational cancer therapy. Trends in Mo-lecular Medicine, 2007, 13, 353-361.
[25] Zhou, J.; Giannakakou, P. Targeting microtubules for cancer che-motherapy. Curr. Med. Chem. Anticancer Agents, 2005, 5, 65-71.
[26] Ricke, R.M.; van Deursen, J.M. Correction of microtubule–kinetochore attachment errors: Mechanisms and role in tumor sup-pression. Seminars in cell & developmental biology, 2011, 22, 559-65.
[27] Blajeski, A.L.; Phan, V.A.; Kottke, T.J.; Kaufmann, S.H. G1 and G2 cell-cycle arrest following microtubule depolymerization in human breast cancer cells. J. Clin. Investig., 2002, 110, 91-99.
[28] Milross, C.G.; Mason, K.A.; Hunter, N.R.; Chung, W.K.; Peters, L.J.; Milas, L. Relationship of mitotic arrest and apoptosis to anti-tumor effect of paclitaxel. J. Natl. Cancer Inst., 1996, 88, 1308-1314.
[29] Jordan, M.A.; Wilson, L. Microtubules as a target for anticancer drugs. Nat. Rev. Cancer, 2004, 4, 253-265.
[30] Wilson, L.; Jordan, M.A. Pharmacological probes of microtubule function. In: J. S. Hyams and C. W. Lloyd (eds.), Microtubules, 59–83. New York: Wiley-Liss, 1994.
[31] Kim, S.Y.; An, J.M.; Lee, H.G.; Du, S.K.; Cheong, C.U.; Seo, J.T. 1H-[1,2,4] oxadiazolo[4,3-a]quinoxalin-1-one induces cell cycle ar-rest and apoptosis in HeLa cells by preventing microtubule polym-erization. Biochemical and Biophysical Research Communications, 2011, 408, 287-292.
[32] Xiao, D.; Pinto, J.T.; Soh, J.-W.; Deguchi, A.; Gundersen, G.G.; Palazzo, A.F.; Yoon, J.-T.; Shirin, H.; Weinstein, I.B. Induction of apoptosis by the garlic-derived compound S-allylmercaptocysteine (SAMC) is associated with microtubule depolymerization and JNK1 activation. Cancer Res, 2003, 63, 6825-37.
[33] Gamen, S.; Anel, A.; Pérez-Galán, P.; Lasierra, P.; Johnson, D.; Piñeiro, A.; Naval, J. Doxorubicin treatment activates a Z-VAD-sensitive caspase, which causes deltapsim loss, caspase-9 activity, and apoptosis in Jurkat cells. Exp. Cell Res. 2000, 258, 223-35.
[34] Vogel, C.; Hager, C.; Bastians, H. Mechanisms of mitotic cell death induced by chemotherapy-mediated G2 checkpoint abroga-tion. Cancer Res. 2007, 67, 339-345.