antibody-drug conjugates designed to eradicate tumors with ... · maytansinoid monoclonal antibody...
TRANSCRIPT

Antibody-Drug Conjugates Designed to Eradicate Tumors
with Homogeneous and Heterogeneous Expression
of the Target Antigen
Yelena V. Kovtun, Charlene A. Audette, Yumei Ye, Hongsheng Xie, Mary F. Ruberti,Sara J. Phinney, Barbara A. Leece, Thomas Chittenden, Walter A. Blattler,and Victor S. Goldmacher
ImmunoGen, Inc., Cambridge, Massachusetts
Abstract
Conjugates of the anti-CanAg humanized monoclonal anti-body huC242 with the microtubule-formation inhibitor DM1(a maytansinoid), or with the DNA alkylator DC1 (a CC1065analogue), have been evaluated for their ability to eradicatemixed cell populations formed from CanAg-positive andCanAg-negative cells in culture and in xenograft tumors inmice. We found that in culture, conjugates of either drugkilled not only the target antigen-positive cells but also theneighboring antigen-negative cells. Furthermore, we showedthat, in vivo , these conjugates were effective in eradicatingtumors containing both antigen-positive and antigen-negativecells. The presence of antigen-positive cells was required forthis killing of bystander cells. This target cell–activated killingof bystander cells was dependent on the nature of the linkerbetween the antibody and the drug. Conjugates linked via areducible disulfide bond were capable of exerting thebystander effect whereas equally potent conjugates linkedvia a nonreducible thioether bond were not. Our data offer arationale for developing optimally constructed antibody-drugconjugates for treating tumors that express the target antigeneither in a homogeneous or heterogeneous manner. (CancerRes 2006; 66(6): 3214-21)
Introduction
Antibody-drug conjugates, also called immunoconjugates, havebeen developed as anticancer agents with increased tumorselectivity through binding of their monoclonal antibody moietyto tumor-associated antigens (1). Antibodies that target differenttumor cell-surface antigens have been linked to various types ofcytotoxic compounds (2). Antibody conjugates with highlycytotoxic compounds, such as calicheamicins, maytansinoids,auristatins, CC1065 analogues (DC drugs), or novel taxoids, exhibitpotent and selective killing of target tumor cells in vitro andin animal models (3, 4). Several maytansinoid conjugates arecurrently in clinical development (5, 6) and a conjugate of an anti-CD33 antibody with calicheamicin, gemtuzumab ozogamicin(Mylotarg), has been approved for the treatment of relapsed acutemyeloid leukemia (7).Antibody-cytotoxic drug conjugates are designed to bind selec-
tively to, and then kill, antigen-positive cells. Consequently, the
antitumor efficacy of such conjugates might be impaired if the targetantigen is expressed in tumors in a heterogeneous fashion. It has beenreported that several types of antibody-targeted cytotoxic agents,such as radioimmunoconjugates, immunoliposomes, antibody-directed prodrug therapy (ADEPT), and gene-directed enzymeprodrug therapy (GDEPT), can kill not only antigen-positive tumorcells but also proximally located antigen-negative tumor cells (8–10).Such killing was termed ‘‘bystander effect.’’ Little, however, is knownabout the efficacy of antibody-cytotoxic drug conjugates againsttumors that express the target antigen in a heterogeneous manner.A conjugate of the anti-Ley antibody BR96 with doxorubicin (BR96-Dox) was tested in rats for its ability to eradicate human tumorxenografts with heterogeneous Ley antigen expression and found tobe ineffective against antigen-negative or low-expressing cells (9, 10).In contrast, a conjugate of the anti-CanAgmonoclonal antibody C242with the cytotoxic maytansinoid DM1 was effective in mice againsthuman tumor xenografts that expressed the antigen heterogeneously(11). It has been hypothesized (11, 12) that target cells efflux the drug,which then kills the neighboring cells independently of antigenexpression.Here we report studies elucidating the mechanism of this target
cell–activated killing of bystander cells. We observed that killing ofbystander antigen-negative cells through targeting of antigen-positive cells with a conjugate of the humanized C242 antibody(huC242), covalently linked either to DM1 or DC1 drugs, wasdependent on the nature of the linker used in the conjugate. Con-jugates with linkers that contained a disulfide bond, but not thoselinked through a thioether bond, showed extensive proximal cellkilling both in vitro and in vivo . We found that antigen-positive cellsprocess the disulfide-linked conjugate and release a maytansinoiddrug that is highly cytotoxic to antigen-negative cells. Meanwhile, aprocessing of the thioether-linked conjugate by antigen-positive cellsyields a poorly cytotoxic maytansinoid drug. Therefore, conjugateswith limited or prominent bystander cytotoxicity can be designedthrough manipulation of their linker composition.
Materials and Methods
Immunoconjugates. For the preparation of immunoconjugates, the may-tansinoid DM1 (N2V-deacetyl-N2V-(3-mercapto-1-oxopropyl)-maytansine)
was synthesized from the microbial fermentation product ansamitocin
P-3 as previously described (13). The synthesis of the analogue of CC-1065,
5-[(3-mercapto-1-oxopropyl)amino]-bis-indolyl-(seco)-1,2,9,9a-tetrahydrocy-clopropa[c]benz[e]indol-4-one compound, DC1, has been reported else-
where (14). The humanization of the C242 antibody (huC242) was done
by the resurfacing method that has previously been described (15).Antibody-drug conjugates were prepared using N -succinimidyl-4-
(2-pyridyldithio)pentanoate for disulfide linkage or N -succinimidyl-4-
(N -maleimidomethyl)cyclohexane-1-carboxylate (SMCC) for thioether
Requests for reprints: Yelena V. Kovtun, ImmunoGen, Inc., 128 Sidney Street,Cambridge, MA 02139. Phone: 617-995-4917; Fax: 617-995-2510; E-mail: [email protected].
I2006 American Association for Cancer Research.doi:10.1158/0008-5472.CAN-05-3973
Cancer Res 2006; 66: (6). March 15, 2006 3214 www.aacrjournals.org
Research Article
Research. on May 2, 2020. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

linkage as described elsewhere (11, 13). The immunoconjugates used in thisstudy contained, on the average, 3.5 cytotoxic drug molecules, DM1 or DC1,
per antibody molecule (Fig. 1).
Cell lines. COLO 205 (human colon adenocarcinoma, ATCC CCL-222),
HL-60 (human acute promyelocytic leukemia, ATCC CCL-240), andNamalwa (human Burkitt’s lymphoma, ATCC CRL-1432) cultures were
maintained in RPMI 1640 supplemented with 10% heat-inactivated fetal
bovine serum (FBS) and 50 Ag/mL gentamicin sulfate. HT-29 (human colon
adenocarcinoma, ATCC HTB-38), A375 (human malignant melanoma,ATCC CRL-1619), and HepG2 (human hepatocellular carcinoma, ATCC HB-
8065) cultures were maintained in DMEM supplemented with 10% heat-
inactivated FBS and 50 Ag/mL gentamicin sulfate. SNU-16 (gastric
carcinoma, ATCC CRL-5974) culture was maintained in modified RPMI1640 (ATCC, 30-2003) supplemented with 10% heat-inactivated FBS and
50 Ag/mL gentamicin sulfate. All cell lines were cultured in a humidified
incubator at 37jC, 6% CO2.Binding of the huC242 antibody to CanAg. The binding of the huC242
antibody to CanAg-positive cells was evaluated by an indirect immunoflu-
orescence assay using flow cytometry. Cells (5 � 104 per well) were plated
in a round-bottomed 96-well plate and incubated at 4jC for 3 hours withserial dilutions of huC242 antibody in 0.2 mL of a-MEM supplemented with
2% (v/v) normal goat serum (Sigma, St. Louis, MO). Each sample was
assayed in triplicate. Control wells lacked huC242. The cells were then
washed with 0.2-mL cold (4jC) medium and stained with fluorescein-labeled goat anti-human immunoglobulin G (IgG) antibody for 1 hour at
4jC. The cells were again washed with medium, fixed in 1% formaldehyde/
PBS solution, and analyzed using a FACSCalibur flow cytometer (BDBiosciences, San Jose, CA).
Three-dimensional collagen cytotoxicity assay. COLO 205 cells (2 �105/mL) were treated with huC242-DM1 (1 � 10�8 mol/L of conjugated
antibody, 24 hours incubation at 37jC, 6% CO2), 1% formaldehyde(20 minutes incubation), or 5 Gy UV irradiation (UV Stratalinker 1800,
Stratagene, La Jolla, CA). Treated cells were washed thrice with 20 to 30 mL
of fresh culture medium and resuspended in culture medium at 1 � 108/mL.
Untreated COLO 205, Namalwa, A375, and HepG2 cells were washed and
resuspended in a similar manner. One microliter of each of the untreatedcell suspensions was then mixed with 0, 1, 2, 4, or 8 AL of the treated
COLO 205 cells. One hundred microliters of ice-cold collagen gel solution
(three-dimensional collagen cell culture system, Chemicon International,
Temecula, CA) were added to the cell mixtures and the cell-collagenmixtures were dispensed immediately (5 AL/well) in round-bottomed
96-well plates. The plates were incubated at 37jC for 1 hour to allow
polymerization of the collagen. Fresh culture medium (200 AL) was then
gently added to each well. Five to six days later, the number of viable cellswas determined using a colorimetric cell proliferation assay measuring the
reduction of 3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide
(MTT) to formazan by live cells. Fifty microliters of the MTT stock solution
(5 mg/mL in PBS) were added to each well. The plates were incubated at37jC for 3 hours and then centrifuged for 10 minutes at 860 � g . Culture
medium was aspirated from each well, the formazan crystals were
solubilized with DMSO (100 AL per well), and A540nm was measured. Therelative number of cells (cell survival) in each well was calculated by
correcting for the medium background reading and then dividing each
value by the average of the values in the control (untreated cells) wells.
Liquid culture cytotoxicity assay. Dilutions of immunoconjugates inthe appropriate medium were added to wells of 96-well round-bottomed
plates containing 2 � 103 COLO 205, 2 � 103 SNU-16, or 2 � 103 Namalwa
cells, or a mixture of 2 � 103 COLO 205 with 2 � 103 Namalwa cells, or a
mixture of 2 � 103 SNU-16 with 2 � 103 Namalwa cells. The plates wereincubated for 5 days at 37jC, 6% CO2. Pictures of the cells in each well were
then taken with a Nikon Diaphot 300 inverted microscope.
ELISA assays for the quantitation of huC242-DM1, huC242-SMCC-DM1, and maytansinoid drug released from the conjugate. The
methods have previously been described (16). Briefly, to measure the
concentrations of huC242-DM1, the conjugate from either the standard or
the test samples was captured on ELISA plates coated with a murine anti-maytansinoid monoclonal antibody (ImmunoGen, Cambridge, MA) and
detected with horseradish peroxidase–labeled donkey anti-human IgG
(Jackson ImmunoResearch, West Grove, PA). To determine the concentra-
tion of the maytansinoid drug released from the conjugate, protein wasprecipitated from samples by addition of 5 volumes of ice-cold acetone.
Precipitated protein was sedimented by centrifugation at 16,000 � g and
the concentrations of released drug in the supernatant were determinedusing a competition ELISA as follows. Plates were coated with DM1 that
had been conjugated to bovine serum albumin (BSA-DM1; ImmunoGen).
Dilution samples of a maytansine standard solution and test samples were
mixed with biotinylated murine anti-maytansinoid monoclonal antibody(ImmunoGen) and then incubated in the BSA-DM1–coated plate. Finally,
the bound biotinylated anti-maytansinoid antibody was detected with
streptavidin-horseradish peroxidase (Jackson ImmunoResearch).
Human xenograft tumor models in mice. Five-week-old, female CB-17severe combined immunodeficient (SCID) mice were obtained from Taconic
(Germantown, NY). One week later, mice were inoculated s.c. with one
of the following cell lines: HT-29 (2 � 106 cells per mouse), COLO 205
(1 � 107), Namalwa (2 � 106), or a mixture of COLO 205 and Namalwa cells(1 � 107 and 2 � 106 cells per mouse, respectively). Nine to thirteen days
later, each mouse received daily i.v. bolus injections of PBS (control),
huC242-SMCC-DM1, or huC242-DM1 containing 25, 75, or 150 Ag/kg ofconjugated DM1 on 5 consecutive days (qd � 5). Tumor dimensions were
measured twice per week and the volume was calculated as 1/2 (L �W � H),
where L is the length, W the width, and H the height of the tumor.
Immunohistochemistry. Tumor tissues were fixed in 10% formalin andembedded in paraffin. Immunohistochemical examination of the paraffin
sections was done using murine C242 antibody to detect COLO 205 cells
and murine anti-CD38 antibody (RDI-CD38abm-290, Research Diagnostic,
Flanders, NJ) to detect Namalwa cells using the avidin-biotin-peroxidase(ABC) technique.
Results
HuC242-DM1 kills nontarget cells proximal to its target cellsin vitro . The immunoconjugate, cantuzumab mertansine orFigure 1. Structures of antibody-drug conjugates.
Bystander Effect of Antibody-Drug Conjugates
www.aacrjournals.org 3215 Cancer Res 2006; 66: (6). March 15, 2006
Research. on May 2, 2020. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

huC242-DM1, is effective in eradicating CanAg-expressing humantumor xenografts in mice, including those that express the antigenin a heterogeneous manner (11), but is not effective against CanAg-negative xenograft tumors.2 To determine if the huC242-DM1conjugate associated with the CanAg-positive cells has the abilityto kill neighboring nontargeted cells, we studied its behaviorin vitro using antigen-positive and antigen-negative mixed cellcultures in a three-dimensional collagen matrix. Cells can beembedded into the matrix at high densities simulating the in vivotumor environment.In a first series of experiments, treated and untreated antigen-
positive COLO 205 cells (Fig. 2) were used. One set of cells wasincubated in culture medium with huC242-DM1 at 37jC for 16hours. The unbound conjugate was then removed by extensivewashing and increasing numbers of treated cells were mixed withsamples of untreated cells to give ratios of treated to untreatedcells of 0:1 (control), 1:1, 4:1, and 8:1, respectively. The cell mixtureswere then embedded into collagen, incubated for 5 days, and thenumber of viable cells in each sample was determined in an MTTassay. In further control cultures, the immunoconjugate-treatedCOLO 205 cells were replaced by formaldehyde-treated or UVlight–treated COLO 205 cells. The results are shown in Fig. 3A . Thesurviving fraction of cells progressively decreased in the culturescontaining increasing numbers of conjugate-treated cells (Fig. 3A,black columns). Thus, untreated cells were killed in the mixedcultures. This killing was conjugate dependent and not due to anyinhibitory effect of dying COLO 205 cells on the untreated cellpopulation because COLO 205 cells killed by either formaldehydetreatment or UV irradiation did not significantly affect theproliferation of the untreated cells (Fig. 3A, gray and white columns ,respectively).In a second series of experiments, COLO 205 cells treated
with huC242-DM1, as described above, were mixed with antigen-
negative cell lines at a ratio of 1:1, embedded in collagen,incubated for 5 days, and then assayed for cell growth in anMTT assay. The antigen-negative cell lines were of diverseorigins: a Burkitt’s lymphoma (Namalwa), a melanoma (A375),and a hepatocarcinoma (HepG2). As shown in Fig. 3B , huC242-DM1-treated cells inhibited the growth of all cocultured, antigen-negative cell lines. Namalwa cells were the most sensitive tothe cytotoxic effect generated by conjugate pretreated COLO 205cells, as indicated by a decrease in the surviving fraction to <0.2;HepG2 cells were the least sensitive with a surviving fraction off0.7; and A375 and COLO 205 cells showed intermediatesensitivity.We next examined whether the target cell–dependent killing of
nontarget cells also occurs in liquid cell cultures. We used antigen-negative, nonadherent Namalwa cells as reporter cells in mixedcultures and semiadherent COLO 205 as target cells. The two celltypes were plated together in a 96-well round-bottomed plate.Within a few hours, all cells settled together in close proximity inthe middle of the well. The experiment included wells thatcontained 2 � 103 Namalwa cells alone, 2 � 103 COLO 205 cellsalone, and a mixture of 2 � 103 cells of each of the two cell lines, allin 0.2-mL liquid culture medium containing 1 nmol/L huC242-DM1. After incubation for 5 days at 37jC, we took photographsof the cell populations (Fig. 3C). In the absence of the conjugate(Fig. 3C, left column), the number of cells in all wells increasedsome 20-fold as determined by cell counting. The mixed cellpopulations were also analyzed by flow cytometry to confirm thatboth the Namalwa and COLO 205 cells had proliferated. Alexa-labeled huC242 served to stain CanAg-positive COLO 205 cellswhereas the B cell–specific Alexa-labeled anti-CD19 antibody wasused for detection of the Namalwa cells. This analysis revealed thatafter 5 days of proliferation, conjugate-free mixed culturesconsisted of 40% COLO 205 cells and 60% Namalwa cells,3 showing
Figure 2. Histograms of huC242 antibody bindingto colon adenocarcinoma cell lines COLO 205 andHT-29, the gastric carcinoma cell line SNU-16, themelanoma cell line A375, and the Burkitt’slymphoma cell line Namalwa. The cells wereincubated with huC242 and then stained withfluorescein-labeled antihuman IgG as describedin Materials and Methods. The cell-associatedfluorescence was measured on a fluorescence-activated cell sorter and the histograms are shownin solid lines. Dashed lines, histograms of cellsstained with fluorescein-labeled antihuman IgGwithout preincubation with huC242 (backgroundstaining). The percentages of antigen-positive andthe antigen-negative cells are given in numbers.
3 K. Whiteman, unpublished.2 Unpublished results.
Cancer Research
Cancer Res 2006; 66: (6). March 15, 2006 3216 www.aacrjournals.org
Research. on May 2, 2020. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

that both cell types had proliferated at similar rates. As expected,huC242-DM1 killed most cells in wells containing CanAg-positiveCOLO 205 cells (Fig. 3C, top row) and did not significantly affect thegrowth in wells containing antigen-negative Namalwa cells (Fig. 3C,middle row). In wells with a mixed population of cells, theconjugate killed both COLO 205 and Namalwa cells (Fig. 3C, bottomrow). Thus, huC242-DM1-treated COLO 205 cells can eradicateproximal antigen-negative cells in liquid culture as well as in athree-dimensional collagen cell culture.To test if target cell lines other than COLO 205 can exert this
cytotoxic effect onto bystander cells, we did an analogousexperiment using another CanAg-positive cell line, the gastriccarcinoma SNU-16 (Fig. 2). As shown in Fig. 3D , huC242-DM1(1 nmol/L) killed SNU-16 cells, did not affect the proliferation ofNamalwa cells, and killed the mixed Namalwa/SNU-16 cellpopulation. Together, these experiments show that the in vitrobystander effect of huC242-DM1 is not limited either to a particulartype of CanAg-positive target cell or a particular type of CanAg-negative bystander cells.Bystander effect of immunoconjugates is dependent on
the nature of the antibody-drug linker. HuC242-DM1 consists
of DM1 molecules conjugated to the huC242 antibody through adisulfide-containing linker. This linker is readily reduced bythiols in vitro in a cell-free environment (data not shown), andtherefore is likely to be cleavable by abundant cell-associatedthiols. To test whether the presence of the disulfide bond in thelinker is required for generation of the bystander effect, wesynthesized huC242-SMCC-DM1, a conjugate in which DM1 islinked to the antibody via a nonreducible thioether bond(Fig. 1), and tested its in vitro cytotoxicity on COLO 205 cells,Namalwa cells, and mixed COLO 205/Namalwa cell populations.HuC242-SMCC-DM1 was as potent as huC242-DM1 in killingCOLO 205 target cells with an IC50 value of 4 � 10�11 mol/L(data not shown). This cytotoxicity was CanAg selective becauseneither of the conjugates was cytotoxic for the antigen-negativeNamalwa cells in the entire concentration range tested (up to1 � 10�9 mol/L). Unlike huC242-DM1, huC242-SMCC-DM1displayed only marginal, if any, cytotoxicity on bystander cellsin either the three-dimensional collagen matrix assay or liquidculture assay (Fig. 3B and C , respectively), suggesting that themechanism of the bystander effect for huC242-DM1 includes adisulfide bond cleavage step.
Figure 3. Immunoconjugates linked via a disulfide, but not thioether, bond kill nontarget cells located proximally to its target cells in vitro. A, effect of COLO 205cells treated with huC242-DM1, formaldehyde, or UV light, on the proliferation of untreated COLO 205 cells in three-dimensional collagen cell cultures. The untreatedcells were mixed with the treated cells at different ratios and the samples were incubated in collagen for 5 days; then viable cells were measured using an MTT assay.The fraction of surviving cells is shown for each cell mixture ratio. Black, gray, and white columns, results for cultures with huC242-DM1-treated cells, withformaldehyde-treated cells, and with UV light–treated cells, respectively. The experiment was repeated twice with similar results. B, effect of COLO 205 cells treatedwith huC242-DM1 or huC242-SMCC-DM1 on untreated COLO 205, Namalwa, A375, and HepG2 cells when grown in mixed three-dimensional collagen cell cultures.Samples of huC242-DM1 or huC242-SMCC-DM1 treated COLO 205 cells were mixed with equal numbers of Namalwa, A375, HepG2, or untreated COLO 205cells and the mixed cultures were incubated in a collagen matrix for 5 days; then the number of viable cells were determined using an MTT assay and compared withthose of control samples (untreated cells only). The fraction of surviving cells is shown for each cell line; black columns, COLO 205; gray columns, Namalwa;white columns, A375; and dashed columns, HepG2 cells. C, treatment of CanAg-positive COLO 205 cells, CanAg-negative Namalwa cells, or mixed COLO 205 andNamalwa cell populations with huC242-DM1, huC242-SMCC-DM1, huC242-DC1, or huC242-SMCC-DC1 in liquid cell culture. COLO 205 cells, Namalwa cells,and mixed populations of equal numbers of COLO 205 and Namalwa cells were grown in round-bottomed wells of tissue culture plates in the absence (left row ) orpresence (right row ) of 1 nmol/L of one of the antibody-drug conjugates. After 5 days of incubation, photographs of the wells were taken. D, results of an experimentanalogous to that in (C ) except that the antigen-positive cell line is SNU-16.
Bystander Effect of Antibody-Drug Conjugates
www.aacrjournals.org 3217 Cancer Res 2006; 66: (6). March 15, 2006
Research. on May 2, 2020. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from
Bystander effect is not limited to antibody-maytansinoidconjugates. To determine if the bystander effect is a uniqueproperty of antibody-maytansinoid conjugates, we examinedconjugates of huC242 with DC1. DC1 is an analogue of theminor groove–binding DNA alkylator CC-1065 and differs fromDM1 in both its structure (Fig. 1) and mechanism of action.Previously, we had reported potent antigen-selective conjugatesof DC1 with an anti-CD19 antibody and an anti-CD56 antibody(14). We constructed analogous conjugates of DC1 with huC242,in which the drug and the antibody were conjugated via eithera disulfide-containing linker (huC242-DC1) or a thioether-containing linker (huC242-SMCC-DC1) as shown in Fig. 1, andtested their ability to kill antigen–positive and bystander cellsin vitro . At concentrations between 2 � 10�10 and 2 � 10�9
mol/L, either conjugate killed most of the cells in antigen-positive COLO 205 cultures (Fig. 3C, top row , results for the 1 �10�9 mol/L concentration) but not the cells in antigen-negativeNamalwa cultures (Fig. 3C, middle row), confirming that theircytotoxicity is antigen dependent. When tested on mixed target/nontarget cell populations, huC242-DC1 was able to kill mostcells of both kinds whereas huC242-SMCC-DC1 was not (Fig. 3C,bottom row). These experiments show that the ability to killbystander cells is not an exclusive property of DM1 conjugatesbut applies to other antibody-drug conjugates linked bydisulfide-containing bonds.Bystander effect of huC242-DM1 is generated through
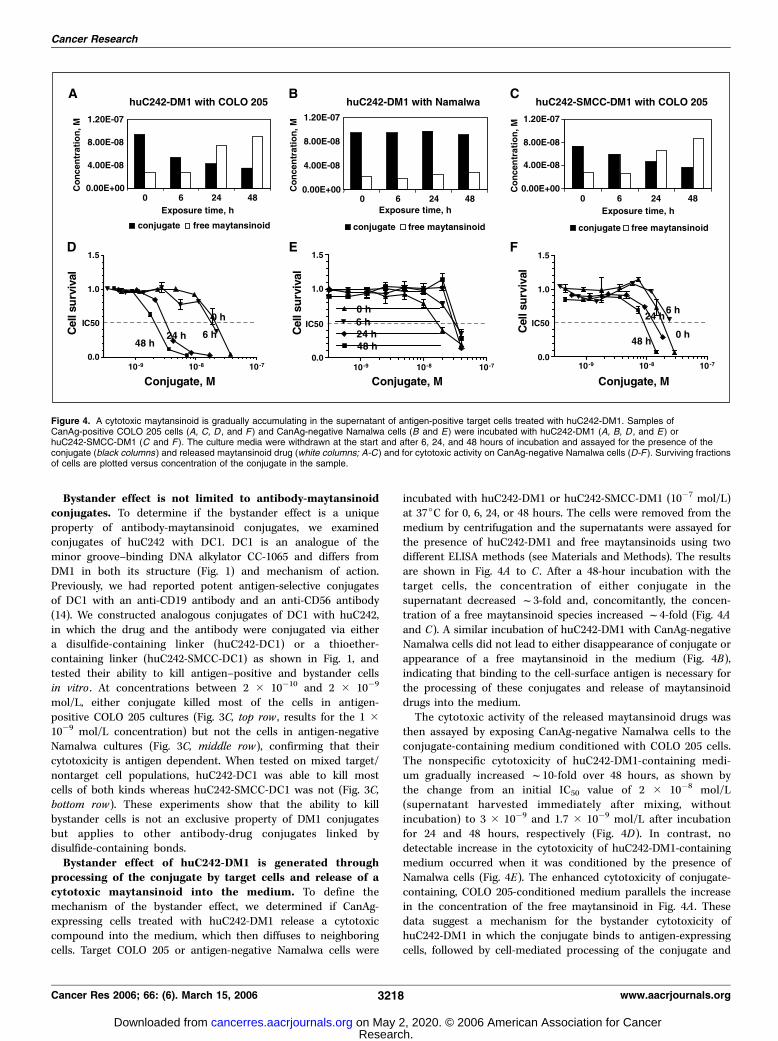
processing of the conjugate by target cells and release of acytotoxic maytansinoid into the medium. To define themechanism of the bystander effect, we determined if CanAg-expressing cells treated with huC242-DM1 release a cytotoxiccompound into the medium, which then diffuses to neighboringcells. Target COLO 205 or antigen-negative Namalwa cells were
incubated with huC242-DM1 or huC242-SMCC-DM1 (10�7 mol/L)at 37jC for 0, 6, 24, or 48 hours. The cells were removed from themedium by centrifugation and the supernatants were assayed forthe presence of huC242-DM1 and free maytansinoids using twodifferent ELISA methods (see Materials and Methods). The resultsare shown in Fig. 4A to C . After a 48-hour incubation with thetarget cells, the concentration of either conjugate in thesupernatant decreased f3-fold and, concomitantly, the concen-tration of a free maytansinoid species increased f4-fold (Fig. 4Aand C). A similar incubation of huC242-DM1 with CanAg-negativeNamalwa cells did not lead to either disappearance of conjugate orappearance of a free maytansinoid in the medium (Fig. 4B),indicating that binding to the cell-surface antigen is necessary forthe processing of these conjugates and release of maytansinoiddrugs into the medium.The cytotoxic activity of the released maytansinoid drugs was
then assayed by exposing CanAg-negative Namalwa cells to theconjugate-containing medium conditioned with COLO 205 cells.The nonspecific cytotoxicity of huC242-DM1-containing medi-um gradually increased f10-fold over 48 hours, as shown bythe change from an initial IC50 value of 2 � 10�8 mol/L(supernatant harvested immediately after mixing, withoutincubation) to 3 � 10�9 and 1.7 � 10�9 mol/L after incubationfor 24 and 48 hours, respectively (Fig. 4D). In contrast, nodetectable increase in the cytotoxicity of huC242-DM1-containingmedium occurred when it was conditioned by the presence ofNamalwa cells (Fig. 4E). The enhanced cytotoxicity of conjugate-containing, COLO 205-conditioned medium parallels the increasein the concentration of the free maytansinoid in Fig. 4A . Thesedata suggest a mechanism for the bystander cytotoxicity ofhuC242-DM1 in which the conjugate binds to antigen-expressingcells, followed by cell-mediated processing of the conjugate and
Figure 4. A cytotoxic maytansinoid is gradually accumulating in the supernatant of antigen-positive target cells treated with huC242-DM1. Samples ofCanAg-positive COLO 205 cells (A, C, D , and F ) and CanAg-negative Namalwa cells (B and E) were incubated with huC242-DM1 (A, B, D , and E ) orhuC242-SMCC-DM1 (C and F ). The culture media were withdrawn at the start and after 6, 24, and 48 hours of incubation and assayed for the presence of theconjugate (black columns ) and released maytansinoid drug (white columns; A-C ) and for cytotoxic activity on CanAg-negative Namalwa cells (D-F ). Surviving fractionsof cells are plotted versus concentration of the conjugate in the sample.
Cancer Research
Cancer Res 2006; 66: (6). March 15, 2006 3218 www.aacrjournals.org
Research. on May 2, 2020. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

progressive release of a cytotoxic maytansinoid species capable ofkilling neighboring cells.Incubation of huC242-SMCC-DM1 with COLO 205 cells led to
only a modest 2-fold increase in nonspecific cytotoxicity of theculture medium (Fig. 4F). Because both the disulfide-linkedconjugate and the thioether linked conjugate are processed byCOLO 205 cells and release a comparable amount of freemaytansinoid species into the medium (Fig. 4A and C), distinctmetabolites are likely formed from huC242-DM1 and huC242-SMCC-DM1. We have recently identified the chemical structures ofthe metabolites and confirmed that some of the products of thehuC242-DM1 processing are 100- to 10,000-fold more potent thanthe metabolite of huC242-SMCC-SM1 (17).Bystander effect of huC242-DM1 in human xenograft tumor
models. By analogy with the in vitro mixed cell culture systems,we developed mixed xenograft tumor models consisting ofCanAg-expressing COLO 205 cells and CanAg-negative Namalwacells. A mixture of COLO 205 and Namalwa cells was injected s.c.into SCID mice (a total of 1.2 � 107 cells per mouse). Everyanimal (a total of 23 mice) developed a measurable tumor and, 9days after the cell implanting, the mean tumor volume reachedf100 mm3. Two 9-day-old tumors were removed from sacrificedanimals and analyzed for the presence of COLO 205 cells(CanAg+/CD38�) and Namalwa cells (CD38+/CanAg�) byimmunohistochemical staining of the tissues with the murineC242 antibody and a murine anti-CD38 antibody, respectively. Themarker for COLO 205 cells (Fig. 5A-a) and that for Namalwa cells(Fig. 5A-b) stained areas of approximately equal size, confirmingthe mixed nature of the tumors. The vast majority of both COLO205 cells and Namalwa cells appeared viable, with little necrosisapparent. Groups of mice bearing such mixed tumors of f100mm3 in size were then treated with five i.v. injections, given on5 consecutive days (qd � 5), of PBS (control group), huC242-SMCC-DM1, or huC242-DM1, at daily doses of the conjugatescontaining 150 Ag/kg of linked DM1. One day after the lastinjection, two mice from each group were sacrificed and theirtumors were analyzed by immunohistochemistry as describedabove. In the PBS-treated control group, the tumor sectionslooked similar to those obtained from untreated tumors (Fig. 5A ,compare c with a , and d with b), with similarly-sized areas ofCOLO 205 and Namalwa tissues, both displaying only a minimaldegree of necrosis. Sections from tumors treated with huC242-SMCC-DM1 showed strongly necrotic COLO 205 tissue (50-60%necrotic, Fig. 5A-e) and healthy Namalwa tissue (Fig. 5A-f ). Finally,sections from tumors treated with huC242-DM1 showed a highdegree of necrosis (70-80%) in both COLO 205 and Namalwatissues (Fig. 5A, g and h). These in vivo data mirror our in vitroresults using mixed cell populations. The activity of the conjugatewith the nondisulfide linker was restricted to the cells thatexpress the target antigen whereas the conjugate with thedisulfide-containing linker killed both the target and thenontarget tumor cells in the same tumor.These immunohistochemistry results were in agreement with
the effects of huC242-DM1 and huC242-SMCC-DM1 on the tumorgrowth. Treatment with either conjugate was equally effectiveagainst CanAg-positive COLO 205 tumors (Fig. 5B-a): tumors infour of five mice completely regressed and did not relapse untilday 90, when the experiment was terminated, and in each group,one mouse relapsed leading to a 28-day delay in the tumorgrowth. Neither conjugate was active on nontarget Namalwatumors: huC242-SMCC-DM1 treatment did not delay tumor
growth whereas huC242-DM1 effected only a modest 7-day delayin tumor growth (Fig. 5B-b), confirming that the antitumoractivities are CanAg selective. The two conjugates differedmarkedly in their antitumor activity against mixed COLO 205-Namalwa tumors. HuC242-SMCC-DM1 did not delay the progres-sion of mixed tumors whereas huC242-DM1 caused completeregressions of tumors within 2 weeks (Fig. 5B-c) with four of fivemice remaining tumor-free until the end of the study (150 days,i.e., f34 tumor doubling times). Immunohistochemical analysisof the only relapsed tumor in this group revealed that thetumor consisted entirely of Namalwa cells (data not shown).In conclusion, only the conjugate with the disulfide linker showedan ability to kill bystander cells in vivo .We further assessed the in vivo bystander effect of huC242-
DM1 and huC242-SMCC-DM1 in the HT-29 human colon cancerxenograft model in SCID mice. HT-29 tumors are examples ofspecial cases of ‘‘mixed’’ tumors that are generated spontaneouslyfrom a single cell line yet are heterogeneous with respect toantigen expression. HT-29 cells express the huC242 antibodytarget, CanAg, only on a minority of cells (20-40%) in vitro (Fig. 2)or in xenograft tumors (11). Mice bearing established (f130mm3) HT-29 tumors were treated with either huC242-DM1 orhuC242-SMCC-DM1 at various doses (25, 75, or 100 Ag/kg oflinked DM1, qd � 5), and tumor growth was monitored. Whereasthe disulfide-linked conjugate huC242-DM1 induced markedtumor growth delays at each dose (5-25 days depending on thedose; Fig. 6A), the huC242-SMCC-DM1 conjugate produced onlymarginal antitumor effects (2.5-3.5 days of growth delay; Fig. 6B).Taken together, these data provide evidence that huC242-SMCC-DM1 conjugate containing a nondisulfide linker is efficacious onlyagainst tumors in which all proliferating cells express the targetantigen. In contrast, the bystander effect associated with thedisulfide-containing conjugate renders this conjugate also effec-tive against tumors in which only a fraction of cells expresses thetarget antigen.
Discussion
Antibody-drug conjugates show targeted antigen-dependentkilling of cells in vitro and antigen-selective antitumor activity incancer models in vivo (2). In this study, we report that twoantibody-drug conjugates containing different drugs, linked via adisulfide bond, can kill antigen–negative cells in mixed antigen-positive and antigen-negative cell populations in vitro . Further-more, studies in vivo with mixed s.c. xenograft tumors in SCIDmice produced similar results. These results support a mechanismof cytotoxicity for these antibody-drug conjugates, which includesbinding of the conjugate to target cells, cleavage of the conjugatedisulfide bond (presumably through a disulfide exchange reac-tion), and release of a drug capable of killing nearby nontargetcells. Similar cytotoxicity in a vicinity of target cells was pre-viously described for radioimmunoconjugates, immunoliposomes,ADEPT, and GDEPT, and referred as bystander effect (8). Thisreport is a demonstration that antibody-drug conjugates can beengineered either to produce a bystander effect (disulfide-linkedconjugates) or exert a precise killing of antigen-presenting cellswithout damaging proximal antigen-negative tissues (thioether-linked conjugates).Immunohistochemistry studies with antibodies binding to
tumor-associated antigens revealed that many solid tumors expressthe target antigen in a heterogeneous fashion and are populated
Bystander Effect of Antibody-Drug Conjugates
www.aacrjournals.org 3219 Cancer Res 2006; 66: (6). March 15, 2006
Research. on May 2, 2020. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

with both antigen-positive and antigen negative cells (18, 19). Forexample, tumors in about half of patients in the cantuzumabmertansine phase I study expressed CanAg either heterogeneouslyor focally (6). This heterogeneity of antigen expression wasconsidered a barrier to the effective treatment of such tumorswith antibody-drug conjugates because of the presumed target-restricted cell killing by immunoconjugates (20). The in vitro andin vivo bystander cytotoxicity associated with disulfide linker–containing conjugates establishes a rationale for immunoconjugate
treatment of tumors even if they exhibit heterogeneous antigenexpression.The bystander effect adds a degree of nonselective killing activity
to the target cell–restricted cytotoxicity of antibody-drug con-jugates. Potentially, this could be a drawback if normal cells intissues surrounding the cancer tissue are affected. However, thiscollateral toxicity might be well tolerated if it is limited to a smallnumber of cells in intimate proximity to the tumor tissues.Furthermore, normal tissue barriers might prevent the cytotoxic
Figure 5. Bystander cytotoxicity of immunoconjugates in xenograft tumor models. A, immunohistochemical analysis of mixed COLO 205/Namalwa xenografttumors grown in SCID mice and treated either with huC242-DM1 or huC242-SMCC-DM1. Slides were stained with murine C242 antibody to detect COLO 205cells (left column ) or an anti-CD38 antibody to detect Namalwa cells (right column ). a and b, consecutive sections of an untreated tumor; c and d, from a tumorof the control group of animals treated with PBS; e and f, from a tumor in an animal treated with huC242-SMCC-DM1; and g and h, from a tumor in an animal treatedwith huC242-DM1. Arrows, necrotic areas. B, activity of huC242-DM1 and huC242-SMCC-DM1 conjugates against xenograft tumors of CanAg-positive targetCOLO 205 cells (a), antigen-negative Namalwa cells (b), and mixed populations of COLO 205 and Namalwa cells (c ). Animals with established tumors off100 mm3 size were treated on 5 consecutive days with PBS (n, control group), huC242-SMCC-DM1 (o, .), or huC242-DM1 (4, E) at daily doses of theconjugates that contained 150 Ag/kg of linked DM1. Tumor volumes in mm3 were plotted versus time (days after cell inoculation).
Cancer Research
Cancer Res 2006; 66: (6). March 15, 2006 3220 www.aacrjournals.org
Research. on May 2, 2020. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

effect and the potential toxicities contributed by the bystander effectto normal tissues might be mitigated by the inherent resistance ofnondividing cells to the antimitotic action of maytansinoids (21, 22).As a potential advantage, the bystander cytotoxicity may damagetissues intricately involved in supporting the tumor growth, such asendothelial cells and pericytes of tumor neovasculature, or tumorstromal cells, resulting in enhanced antitumor activity of theconjugate against tumors expressing the antigen either homoge-neously or heterogeneously.
Acknowledgments
Received 11/3/2005; revised 12/23/2005; accepted 1/13/2006.The costs of publication of this article were defrayed in part by the payment of page
charges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.
We thank Dr. Dorfman, Brigham and Women’s Hospital (Boston, MA), for hisexpert evaluation of immunohistochemistry slides; Sharon Wilhelm, Kate Lai, RobertZhao, and Hans Erickson for preparation of the antibody-drug conjugates; KathleenWhiteman for her help with tissue culture experiments; and Hans Erickson, JohnLambert, Robert Lutz, Rita Steeves, and Wayne Widdison for numerous enlighteningdiscussions during the project.
Figure 6. Activity of huC242-DM1 and huC242-SMCC-DM1 conjugates against HT-29 xenografttumors, which express the target antigen, CanAg, in aheterogeneous fashion. Groups of five mice bearing13-day-old s.c. tumors of a mean volume of f170mm3 were treated on 5 consecutive days with PBS(n, control group), huC242-DM1 (A ), or with huC242-SMCC-DM1 (B) at daily doses of the conjugates thatcontained 25 Ag/kg (w ), 75 Ag/kg (y), or 150 Ag/kg (o)of linked DM1. Tumor volumes in mm3 were plottedversus time (days after cell inoculation).
References
1. Wu AM, Senter PD. Arming antibodies: prospects andchallenges for immunoconjugates. Nat Biotechnol2005;23:1137–46.
2. Payne G. Progress in immunoconjugate cancer ther-apeutics. Cancer Cell 2003;3:207–12.
3. Goldmacher VS, Blatter WA, Lambert JM, Chari RVJ.Immunotoxins and antibody-drug conjugates for cancertreatment. In: Muzykantov VTV, editor. Biomedicalaspects of drug targeting. Boston: Kluwer AcademicPublishers; 2002. p. 291–309.
4. Lambert JM. Drug-conjugated monoclonal antibodiesfor the treatment of cancer. Curr Opin Pharmacol2005;5:543–9.
5. Helft PR, Schilsky RL, Hoke FJ, et al. A phase I study ofcantuzumab mertansine administered as a singleintravenous infusion once weekly in patients withadvanced solid tumors. Clin Cancer Res 2004;10:4363–8.
6. Tolcher AW, Ochoa L, Hammond LA, et al. Cantuzu-mab mertansine, a maytansinoid immunoconjugatedirected to the CanAg antigen: a phase I, pharmacoki-netic, and biologic correlative study. J Clin Oncol 2003;21:211–22.
7. Bross PF, Beitz J, Chen G, et al. Approval summary:gemtuzumab ozogamicin in relapsed acute myeloidleukemia. Clin Cancer Res 2001;7:1490–6.
8. Allen TM. Ligand-targeted therapeutics in anticancertherapy. Nat Rev Cancer 2002;2:750–63.
9. Muldoon LL. Effect of antigenic heterogeneity on theefficacy of enhanced delivery of antibody-targetedchemotherapy in a human lung cancer intracerebralxenograft model in rats. Neurosurgery 2003;53:1406–12.
10. Muldoon LL, Neuwelt EA. BR96-DOX immunoconju-gate targeting of chemotherapy in brain tumor models.J Neurooncol 2003;65:49–62.
11. Liu C, Tadayoni BM, Bourret LA, et al. Eradication oflarge colon tumor xenografts by targeted delivery ofmaytansinoids. Proc Natl Acad Sci U S A 1996;93:8618–23.
12. Johnson RK, Wong J, Sucheki P, et al. Role of abystander effect in the efficacy of SB-408075, anantibody-maytansinoid complex, in colon carcinomaxenografts. Proc Am Assoc Cancer Res 2000;41:1926.
13. Chari RV, Martell BA, Gross JL, et al. Immunoconju-gates containing novel maytansinoids: promising anti-cancer drugs. Cancer Res 1992;52:127–31.
14. Chari RV, Jackel KA, Bourret LA, et al. Enhancementof the selectivity and antitumor efficacy of a CC-1065analogue through immunoconjugate formation. CancerRes 1995;55:4079–84.
15. Roguska MA, Pedersen JT, Keddy CA, et al. Human-ization of murine monoclonal antibodies throughvariable domain resurfacing. Proc Natl Acad Sci U S A1994;91:969–73.
16. Xie H, Audette C, Hoffee M, Lambert JM, Blattler WA.Pharmacokinetics and biodistribution of the antitumorimmunoconjugate, cantuzumab mertansine (huC242–1),and its two components in mice. J Pharmacol Exp Ther2004;308:1073–82.
17. Erickson HK, Park PU, Widdison WC, et al. Antibody-maytansinoid conjugates are activated in targetedcancer cells by lysosomal degradation and linker-dependent intracellular processing. Cancer Res. In press2006.
18. Greiner JW. Modulation of antigen expression inhuman tumor cell populations. Cancer Invest 1986;4:239–56.
19. Christiansen J, Rajasekaran AK. Biological impedi-ments to monoclonal antibody-based cancer immuno-therapy. Mol Cancer Ther 2004;3:1493–501.
20. Cheng JD, Rieger PT, von Mehren M, Adams GP,Weiner LM. Recent advances in immunotherapy andmonoclonal antibody treatment of cancer. Semin OncolNurs 2000;16:2–12.
21. Drewinko B, Patchen M, Yang LY, Barlogie B.Differential killing efficacy of twenty antitumor drugson proliferating and nonproliferating human tumorcells. Cancer Res 1981;41:2328–33.
22. Rao PN, Freireich EJ, Smith ML, Loo TL. Cell cyclephase-specific cytotoxicity of the antitumor agentmaytansine. Cancer Res 1979;39:3152–5.
Bystander Effect of Antibody-Drug Conjugates
www.aacrjournals.org 3221 Cancer Res 2006; 66: (6). March 15, 2006
Research. on May 2, 2020. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from

2006;66:3214-3221. Cancer Res Yelena V. Kovtun, Charlene A. Audette, Yumei Ye, et al. Target Antigenwith Homogeneous and Heterogeneous Expression of the Antibody-Drug Conjugates Designed to Eradicate Tumors
Updated version
http://cancerres.aacrjournals.org/content/66/6/3214
Access the most recent version of this article at:
Cited articles
http://cancerres.aacrjournals.org/content/66/6/3214.full#ref-list-1
This article cites 20 articles, 11 of which you can access for free at:
Citing articles
http://cancerres.aacrjournals.org/content/66/6/3214.full#related-urls
This article has been cited by 44 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications
Permissions
Rightslink site. (CCC)Click on "Request Permissions" which will take you to the Copyright Clearance Center's
.http://cancerres.aacrjournals.org/content/66/6/3214To request permission to re-use all or part of this article, use this link
Research. on May 2, 2020. © 2006 American Association for Cancercancerres.aacrjournals.org Downloaded from