annual report 2010 - college of agricultural and life sciences
TRANSCRIPT

Annual Report 2010 Biological control of hawkweeds, Pilosella spp.
G. Grosskopf-Lachat, C. Moffat, A. Tataru and A. Mosyakin
June 2011
www.cabi.org
KNOWLEDGE FOR LIFE


CABI Ref: VM00936
Issued June 2011
Biological control of hawkweeds,
Pilosella spp.
Annual Report 2010
G. Grosskopf-Lachat, C. Moffat, A. Tataru and A. Mosyakin
CABI Europe – Switzerland Rue des Grillons 1, CH-2800 Delémont, Switzerland
Tel: ++ 41 32 421 4870 Fax: ++ 41 32 421 4871 Email: [email protected]
Sponsored by: Ministry of Forests, Lands and Natural Resource Operations Montana Weed Trust Fund through Montana State University

This report is the Copyright of CAB International, on behalf of the sponsors of this work where appropriate. It presents unpublished research findings, which should not be used or quoted without written agreement from CAB International. Unless specifically agreed otherwise in writing, all information herein should be treated as confidential.

Table of Contents
Summary ........................................................................................................ 1
1. Introduction ............................................................................................. 2
2. Work programme for period under report ............................................ 3
3. Shipments of test plants ........................................................................ 4
4. Cheilosia psilophthalma (BECKER) (Diptera: Syrphidae) ...................... 4
4.1. Field collection of gravid females ................................................... 4
4.2. No-choice larval transfer tests ........................................................ 4
4.3. Conclusions and outlook ................................................................. 5
5. Aulacidea subterminalis NIBLETT (Hymenoptera: Cynipidae) .............. 5
6. Aulacidea hieracii (L.) (Hymenoptera: Cynipidae) ............................... 6
6.1. Emergence period ............................................................................ 6
6.2. Host-range investigations ................................................................ 7
6.2.1. Sequential no-choice gall development tests ............................... 7
6.2.2. Multiple-choice gall development tests in field cages ................... 8
6.3. Conclusions and outlook ................................................................. 8
7. MSc thesis research ............................................................................... 9
8. Proposed work programme for 2011 ................................................... 11
9. Acknowledgements .............................................................................. 11
10. References............................................................................................. 11

1
Summary
1. In 2010, we continued with no-choice larval transfer tests with the syrphid fly Cheilosia psilophthalma. Due to adverse weather conditions, only a few tests could be established, and none of the plants, including controls, were attacked. Since open-field tests with this potential agent have so far failed, we propose to discontinue host-range investigations for the time being. Without data obtained under open-field conditions, we are not able to adequately describe the ecological host range of this fly.
2. Aulacidea subterminalis, a univoltine gall wasp that galls the stolon tips of Pilosella officinarum and P. aurantiaca, was mass reared in 2010, and 253 A. subterminalis gall clusters were overwintered at CABI Europe – Switzerland (CABI E-CH). Two-thirds of the galls were sent to North America in March 2011 and we are using the remaining galls to maintain the rearing colony at CABI E-CH.
3. Two populations of the gall wasp A. hieracii were screened in 2010, i.e. A. hieracii reared from P. procera collected in Ukraine and a Russian population of A. hieracii. Sequential no-choice gall development tests were carried out with 12 different Pilosella and Hieracium species. Galls were retrieved from Pilosella sp. from southern Russia, P. procera and H. argutum, a native hawkweed species from California. Although plants in the right phenostage were available in 2010, no galls developed on P. caespitosa or other weedy hawkweeds in the genus Pilosella. Since gall production on P. caespitosa is an extremely rare event, we decided to discontinue host-range tests with A. hieracii.
4. Chandra Moffat started research for her MSc thesis in autumn 2009. She is being co-supervised by Dr Jason Pither and Dr Rob Lalonde at the University of British Columbia (UBC), Canada, and advised by Dr Rosemarie De Clerck-Floate of Agriculture and Agri-Food Canada (AAFC). Based at CABI E-CH in summer 2010, field work was conducted across the distribution range of Aulacidea pilosellae, a small cynipid galling the midrib of leaves, stolons and flower stems of several Pilosella species. In the past, we observed that the population from Poland and the Czech Republic has one generation per year whereas the Swiss and southern German population has two generations. We also observed differences in the fundamental host range of the two populations. Chandra is investigating the ecological host range, patterns of host selection and habitat preferences of the gall wasp. In 2010, Chandra’s work involved intensive field sampling of the hawkweeds, the gall wasps, the associated plant community and multiple abiotic features at more than 50 field sites across the study area, i.e. Switzerland, Germany, Poland and the Czech Republic.

2
1. Introduction
Hawkweeds (Pilosella spp.) of Eurasian origin have been accidentally or deliberately introduced into other parts of the world, e.g. North and South America, and New Zealand. Several species in the genus Pilosella have become troublesome weeds outside their native range and are declared noxious weeds in North America (Scoggan, 1979; Gleason and Cronquist, 1991; USDA, NRCS, 2010). Meadow hawkweed, Pilosella caespitosa (syn. Hieracium caespitosum), and orange hawkweed, P. aurantiaca (syn. H. aurantiacum) (see Plate 1), are weeds of pastures and clear-cut areas (Wilson and Callihan, 1999). Pilosella officinarum, P. piloselloides, P. glomerata and P. floribunda are considered noxious weeds in the north-western part of the USA and in British Columbia, Canada. They reproduce by wind-dispersed seed, stolons (except for P. piloselloides) and, in the case of P. caespitosa, adventitious root buds (Wilson and Callihan, 1999). As observed for many hawkweed species in the genus Pilosella, vegetative reproduction results in a mat-forming growth.
In its native range, P. caespitosa is a plant of humid, nutrient-poor pastures and disturbed areas (e.g. roadsides) but it is not regarded as a weed. Pilosella aurantiaca is common in mountainous regions. Disturbance (e.g. overgrazing, burning and deforestation) as well as the lack of specialized natural enemies are thought to be the main reasons for hawkweed invasions outside their native range (Hunter, 1991; Syrett and Smith, 1998). Due to their low growth form and high regenerative capacity, hawkweeds escape mowing successfully and even profit from mechanical disturbance. Chemical control is possible with some broad-spectrum herbicides, e.g. 2,4-D (Meeklah, 1980). However, these are not selective and relatively expensive, and hawkweeds may recolonize pastures from forest margins, roadsides and conservation areas. In addition, herbicide applications are often not feasible in remote areas.
Surveys for phytophagous insects associated with hawkweeds carried out in New Zealand revealed the absence of specialized herbivorous insects (Syrett and Smith, 1998). A biological control programme was initiated in 1993 and five insect species have been approved for release in New Zealand. Since 2000, phytophagous insects have also been studied as potential biological control agents for North America. Oxyptilus pilosellae (Lepidoptera: Pterophoridae) proved to be difficult to rear and able to develop on native North American Hieracium species. Preliminary host-range investigations were undertaken with Aulacidea pilosellae (Hymenoptera: Cynipidae). This species causes small galls on the midrib of leaves and has a restricted host range, but it has not been possible so far to show that it has a negative impact on the growth of P. caespitosa. The multivoltine gall midge Macrolabis pilosellae (Diptera: Cecidomyiidae) has a preference for alien invasive hawkweeds. However, it accepted native North American hawkweed species under field conditions. Investigations on these three insect species have therefore been discontinued.
A petition for field release of the gall wasp Aulacidea subterminalis (Hymenoptera: Cynipidae) in the USA was submitted by Dr Jeff Littlefield (Montana State University, Bozeman). The Technical Advisory Group (TAG) recommended release of the insect and the environmental impact assessment was accepted.
A petition for field release is currently being prepared for the root-feeding hoverfly Cheilosia urbana (Diptera: Syrphidae). Work with C. psilophthalma will be

3
discontinued due to difficulties in obtaining field data regarding its specificity. In 2008, Aulacidea hieracii (Hymenoptera: Cynipidae) was added to the list of potential biological control agents. Galls were collected from Pilosella procera and Hieracium robustum in Ukraine and from an as-yet unidentified Pilosella sp. in southern Russia. In 2008, we obtained galls on P. caespitosa in no-choice gall development tests. This encouraged us, after unsuccessful attempts in 2003 and 2004, to continue host-range investigations.
In contrast to the situation in New Zealand, where all existing hawkweeds are naturalized species, native hawkweeds occur in North America. Indigenous North American hawkweed species belong to the genus Hieracium (subgenera Hieracium and Chionoracium) whereas most invasive species are in the genus Pilosella. While the majority of the invasive hawkweeds reproduce by stolons and by seeds, none of the native North American hawkweed species reproduces by stolons. Insects purely or mainly attacking stolons are therefore being given priority and this should reduce the risk of non-target effects.
Plate 1. Pilosella caespitosa (left) and P. aurantiaca (right) in flower.
2. Work programme for period under report
Cheilosia urbana (Diptera: Syrphidae)
Prepare and submit petition for field release;
Submit publication on host specificity.
Cheilosia psilophthalma (Diptera: Syrphidae)
Conduct additional no-choice larval transfer tests.
Aulacidea subterminalis (Hymenoptera: Cynipidae)
Conduct mass rearing for shipment to North America.
Aulacidea hieracii from Russia and Ukraine (Hymenoptera: Cynipidae)
Conduct no-choice gall development tests in screened rearing cages;
Conduct multiple-choice gall development tests in field cages and screened rearing cages;

4
Collect from the field in Ukraine (September/October 2009).
Project overview
Start compiling overview of the Hieracium project, including agents which failed, for later publication.
3. Shipments of test plants
During 2010, Marijka Haverhals provided us with seeds of Crepis bakeri collected in Idaho and living plants of Hieracium scouleri.
4. Cheilosia psilophthalma (BECKER) (Diptera: Syrphidae)
Cheilosia psilophthalma (Plate 2) is a univoltine hoverfly species, which oviposits onto the rosettes of several hawkweed species. Larvae feed on the above-ground plants parts, i.e. in the centre of the rosette, leaf axils and stolon tips. The larvae pupate in October and puparia overwinter on the ground.
Plate 2. Cheilosia psilophthalma adult (left); eggs (right).
4.1. Field collection of gravid females
Between 21 and 30 April 2010, seven gravid C. psilophthalma females were collected in the garden at CABI E-CH (CABI Europe – Switzerland) to provide eggs for no-choice larval transfer tests (see below). The weather conditions in 2010 were unfavourable for field collection of C. psilophthalma. After a long and cold winter, we had a couple of weeks’ warm weather followed by three weeks of cool and rainy weather. During and after this long period of cold weather, no more flies were found in the field.
4.2. No-choice larval transfer tests
METHODS. Between 3 and 26 May 2010, we established 15 no-choice larval transfer tests. Batches of seven freshly hatched larvae were transferred onto potted plants of P. caespitosa, P. aurantiaca, and six native North American Hieracium species. The plants were each covered with a gauze bag, embedded in the garden

5
and watered as necessary. All pots were overwintered in a garden tunnel and checked for puparia in spring.
RESULTS. One H. albiflorum and one H. argutum plant died during the summer and were therefore not considered for evaluation. No puparia were retrieved from the remaining 13 plants.
4.3. Conclusions and outlook
The results obtained in 2010 are not representative due to the small number of replicates, i.e. we were able to expose only one P. caespitosa and one P. aurantiaca plant, and neither of these were attacked. Since open-field tests with this potential agent have failed so far, we propose to discontinue host-range investigations for the time being. Without data obtained under open-field conditions, we are not able to adequately describe the ecological host range of this fly.
5. Aulacidea subterminalis NIBLETT (Hymenoptera: Cynipidae)
Aulacidea subterminalis is a univoltine gall wasp that reproduces parthenogenetically. Females oviposit into the meristematic tissue of the stolon tips of P. aurantiaca and P. officinarum, which leads to galling of the daughter rosettes. In 2009, Dr Jeff Littlefield (Montana State University, Bozeman, USA), submitted a petition for field release. The Technical Advisory Group (TAG) recommended release, and in 2010, the environmental impact assessment (EIA) was approved.
In March 2010, overwintered A. subterminalis galls were shipped to the quarantine facility at Montana State University, Bozeman. Between 28 May and 10 July 2010, 241 A. subterminalis females emerged at CABI E-CH. We used all females for maintaining the rearing colony. Thirty-two large pots (diameter: 34 cm) containing P. officinarum plants were covered with gauze and ten A. subterminalis females were released onto each rearing pot. The pots were kept in the garden throughout the summer. The galls were harvested from 20 September onwards and 253 gall clusters were overwintered in an underground shelter. In March 2011, two-thirds of the galls were sent to the quarantine facility at Montana State University, Bozeman.
Plate 3. Aulacidea subterminalis female ovipositing into a stolon tip (left); pots used for mass rearing the gall wasp (right).

6
6. Aulacidea hieracii (L.) (Hymenoptera: Cynipidae)
Aulacidea hieracii is a univoltine gall wasp producing multi-chambered stem swellings on several Hieracium and Pilosella species. Females oviposit into the apical meristem of bolting plants (see Plate 4). In the past, the insect was reared from three different hawkweed species, i.e. P. procera collected in Ukraine, Pilosella sp. originating from southern Russia and H. robustum collected in Ukraine. Earlier host-range investigations suggested that the wasps reared from P. procera and H. robustum may be two different species.
Plate 4. Female of Aulacidea hieracii ovipositing into a stolon tip of Pilosella sp. from Russia.
6.1. Emergence period
METHODS. Galls of the Ukrainian and Russian population were obtained from host-range tests carried out at CABI E-CH in 2009. All galls were overwintered under semi-natural conditions in an underground shelter. In addition, Dr Viktor Fursov (National Ukrainian Academy of Sciences, Kiev) collected more than 1000 A. hieracii galls on P. procera in Ukraine between 20 and 27 September 2009. The galls were shipped to CABI E-CH and overwintered under semi-natural conditions. However, the majority of galls had emergence holes and were probably from 2008.
RESULTS. Between 3 and 23 June 2010, 160 male and 100 female A. hieracii of the Russian population emerged. Between 9 and 19 June, 49 A. hieracii females emerged from galls collected in Ukraine and 59 additional females emerged from galls obtained in host-range tests. All wasps were used for sequential no-choice gall development tests.
7
6.2. Host-range investigations
6.2.1. Sequential no-choice gall development tests
Sequential no-choice gall development tests with individual plants yielded very disappointing results in 2009. The attack rate was extremely low and none of the target weeds developed galls. In 2010, we therefore focused on sequential no-choice gall development tests in screened rearing cages with the Ukrainian and Russian populations of A. hieracii in order to test a maximum number of plant species and individual plants. All plants were in the right phenostage, i.e. the plants were in the process of bolting.
METHODS. Three potted plants of the same plant species were exposed to four A. hieracii females and where possible one or two A. hieracii males in screened rearing cages measuring 43 43 73 cm or 31 31 54 cm. After three days, all wasps were retrieved and transferred into a cage containing a different plant species, with dead insects being replaced. All tests were carried out in a polythene-covered garden tunnel. In cases of cold weather lasting several days, we prolonged the exposure period to up to one week. All plants exposed to gall wasps were afterwards embedded in a field cage. In August, we checked the plants for galls. In October 2010, we removed the galls from the plants and placed them into individual plastic cups in order to record adult emergence in spring 2011. Some 26 cages were set up with the Russian A. hieracii population and 46 cages with the Ukrainian gall wasp population.
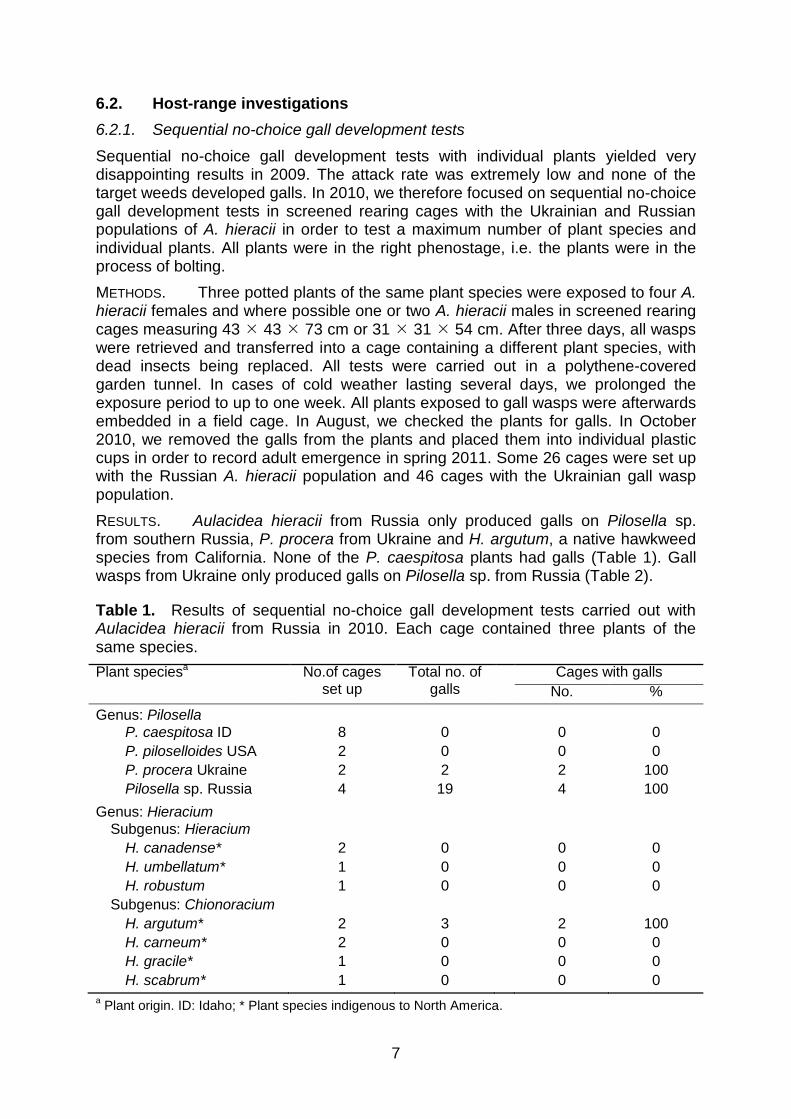
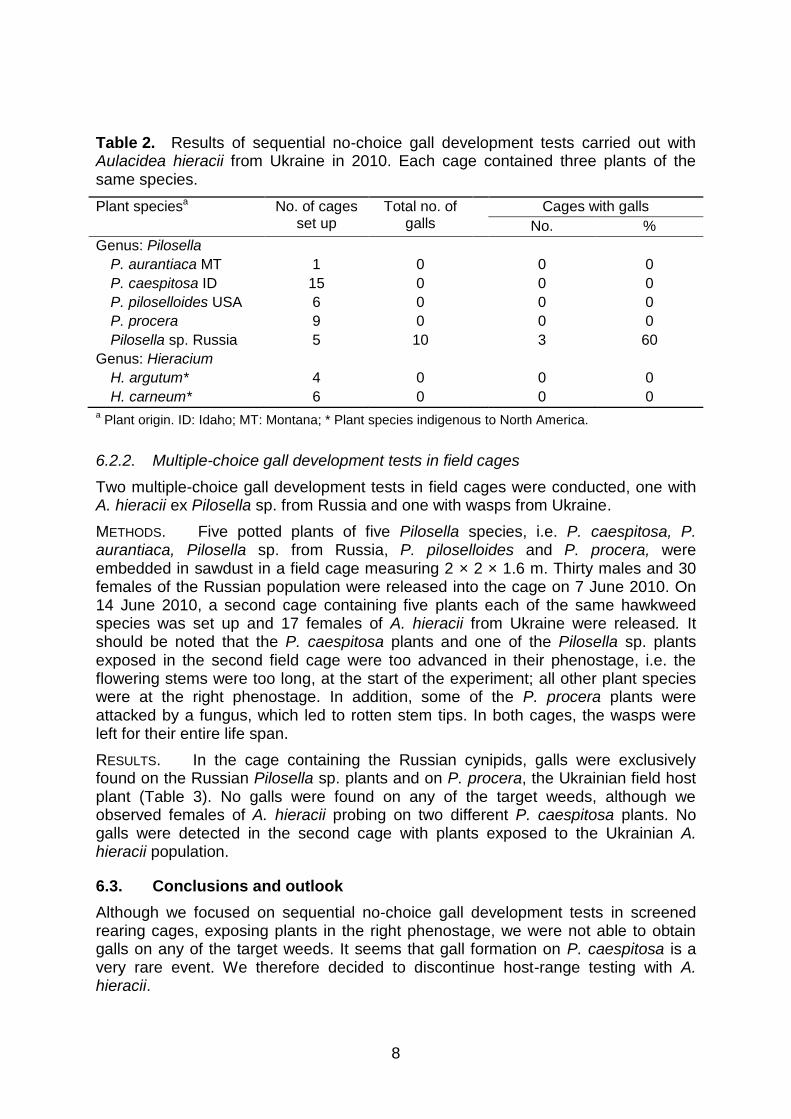
RESULTS. Aulacidea hieracii from Russia only produced galls on Pilosella sp. from southern Russia, P. procera from Ukraine and H. argutum, a native hawkweed species from California. None of the P. caespitosa plants had galls (Table 1). Gall wasps from Ukraine only produced galls on Pilosella sp. from Russia (Table 2).
Table 1. Results of sequential no-choice gall development tests carried out with Aulacidea hieracii from Russia in 2010. Each cage contained three plants of the same species.
Plant speciesa No.of cages
set up
Total no. of
galls
Cages with galls
No. %
Genus: Pilosella
P. caespitosa ID 8 0 0 0
P. piloselloides USA 2 0 0 0
P. procera Ukraine 2 2 2 100
Pilosella sp. Russia 4 19 4 100
Genus: Hieracium
Subgenus: Hieracium
H. canadense* 2 0 0 0
H. umbellatum* 1 0 0 0
H. robustum 1 0 0 0
Subgenus: Chionoracium
H. argutum* 2 3 2 100
H. carneum* 2 0 0 0
H. gracile* 1 0 0 0
H. scabrum* 1 0 0 0
a Plant origin. ID: Idaho; * Plant species indigenous to North America.
8
Table 2. Results of sequential no-choice gall development tests carried out with Aulacidea hieracii from Ukraine in 2010. Each cage contained three plants of the same species.
Plant speciesa No. of cages set up
Total no. of galls
Cages with galls
No. %
Genus: Pilosella
P. aurantiaca MT 1 0 0 0
P. caespitosa ID 15 0 0 0
P. piloselloides USA 6 0 0 0
P. procera 9 0 0 0
Pilosella sp. Russia 5 10 3 60
Genus: Hieracium
H. argutum* 4 0 0 0
H. carneum* 6 0 0 0 a Plant origin. ID: Idaho; MT: Montana; * Plant species indigenous to North America.
6.2.2. Multiple-choice gall development tests in field cages
Two multiple-choice gall development tests in field cages were conducted, one with A. hieracii ex Pilosella sp. from Russia and one with wasps from Ukraine.
METHODS. Five potted plants of five Pilosella species, i.e. P. caespitosa, P. aurantiaca, Pilosella sp. from Russia, P. piloselloides and P. procera, were embedded in sawdust in a field cage measuring 2 × 2 × 1.6 m. Thirty males and 30 females of the Russian population were released into the cage on 7 June 2010. On 14 June 2010, a second cage containing five plants each of the same hawkweed species was set up and 17 females of A. hieracii from Ukraine were released. It should be noted that the P. caespitosa plants and one of the Pilosella sp. plants exposed in the second field cage were too advanced in their phenostage, i.e. the flowering stems were too long, at the start of the experiment; all other plant species were at the right phenostage. In addition, some of the P. procera plants were attacked by a fungus, which led to rotten stem tips. In both cages, the wasps were left for their entire life span.
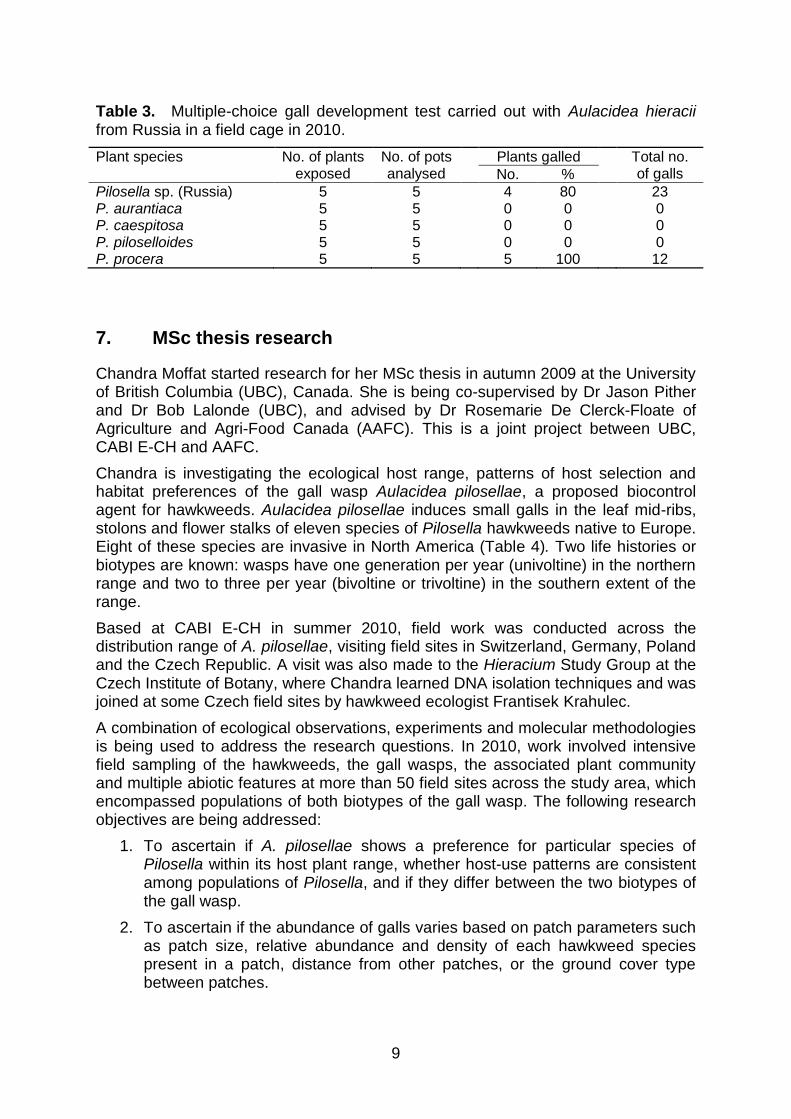
RESULTS. In the cage containing the Russian cynipids, galls were exclusively found on the Russian Pilosella sp. plants and on P. procera, the Ukrainian field host plant (Table 3). No galls were found on any of the target weeds, although we observed females of A. hieracii probing on two different P. caespitosa plants. No galls were detected in the second cage with plants exposed to the Ukrainian A. hieracii population.
6.3. Conclusions and outlook
Although we focused on sequential no-choice gall development tests in screened rearing cages, exposing plants in the right phenostage, we were not able to obtain galls on any of the target weeds. It seems that gall formation on P. caespitosa is a very rare event. We therefore decided to discontinue host-range testing with A. hieracii.
9
Table 3. Multiple-choice gall development test carried out with Aulacidea hieracii from Russia in a field cage in 2010.
Plant species No. of plants exposed
No. of pots analysed
Plants galled Total no. of galls No. %
Pilosella sp. (Russia) 5 5 4 80 23 P. aurantiaca 5 5 0 0 0 P. caespitosa 5 5 0 0 0 P. piloselloides 5 5 0 0 0 P. procera 5 5 5 100 12
7. MSc thesis research
Chandra Moffat started research for her MSc thesis in autumn 2009 at the University of British Columbia (UBC), Canada. She is being co-supervised by Dr Jason Pither and Dr Bob Lalonde (UBC), and advised by Dr Rosemarie De Clerck-Floate of Agriculture and Agri-Food Canada (AAFC). This is a joint project between UBC, CABI E-CH and AAFC.
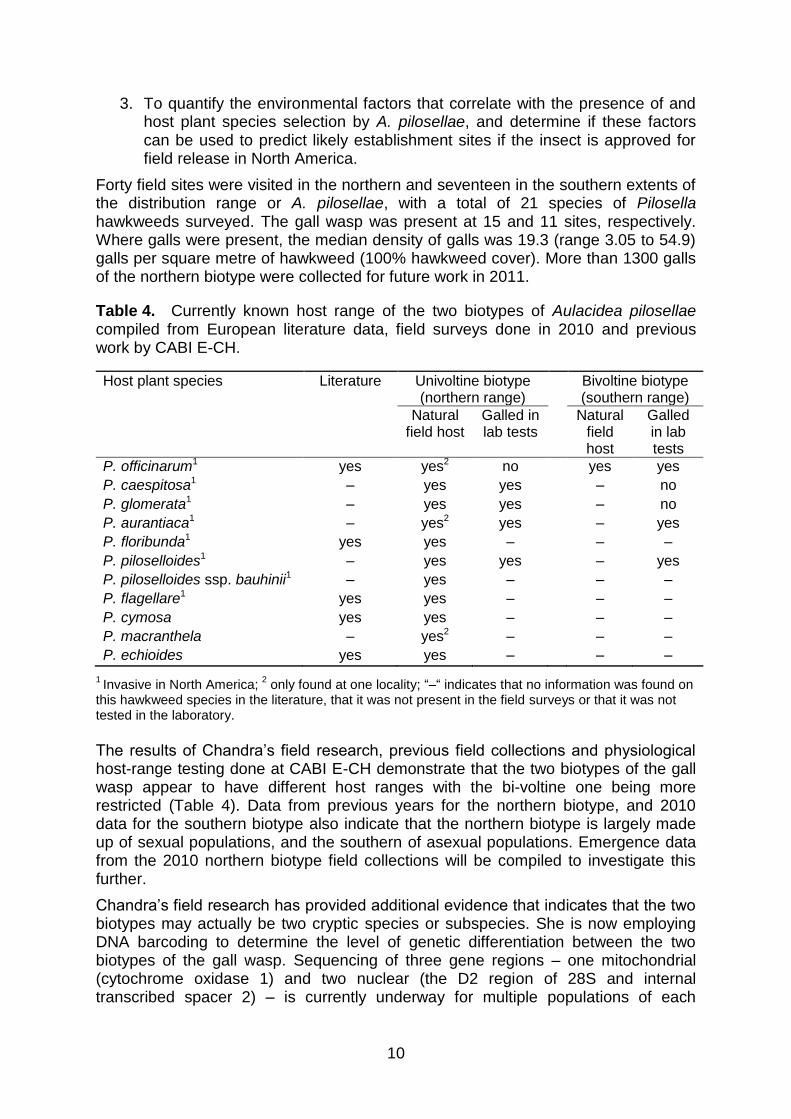
Chandra is investigating the ecological host range, patterns of host selection and habitat preferences of the gall wasp Aulacidea pilosellae, a proposed biocontrol agent for hawkweeds. Aulacidea pilosellae induces small galls in the leaf mid-ribs, stolons and flower stalks of eleven species of Pilosella hawkweeds native to Europe. Eight of these species are invasive in North America (Table 4). Two life histories or biotypes are known: wasps have one generation per year (univoltine) in the northern range and two to three per year (bivoltine or trivoltine) in the southern extent of the range.
Based at CABI E-CH in summer 2010, field work was conducted across the distribution range of A. pilosellae, visiting field sites in Switzerland, Germany, Poland and the Czech Republic. A visit was also made to the Hieracium Study Group at the Czech Institute of Botany, where Chandra learned DNA isolation techniques and was joined at some Czech field sites by hawkweed ecologist Frantisek Krahulec.
A combination of ecological observations, experiments and molecular methodologies is being used to address the research questions. In 2010, work involved intensive field sampling of the hawkweeds, the gall wasps, the associated plant community and multiple abiotic features at more than 50 field sites across the study area, which encompassed populations of both biotypes of the gall wasp. The following research objectives are being addressed:
1. To ascertain if A. pilosellae shows a preference for particular species of Pilosella within its host plant range, whether host-use patterns are consistent among populations of Pilosella, and if they differ between the two biotypes of the gall wasp.
2. To ascertain if the abundance of galls varies based on patch parameters such as patch size, relative abundance and density of each hawkweed species present in a patch, distance from other patches, or the ground cover type between patches.
10
3. To quantify the environmental factors that correlate with the presence of and host plant species selection by A. pilosellae, and determine if these factors can be used to predict likely establishment sites if the insect is approved for field release in North America.
Forty field sites were visited in the northern and seventeen in the southern extents of the distribution range or A. pilosellae, with a total of 21 species of Pilosella hawkweeds surveyed. The gall wasp was present at 15 and 11 sites, respectively. Where galls were present, the median density of galls was 19.3 (range 3.05 to 54.9) galls per square metre of hawkweed (100% hawkweed cover). More than 1300 galls of the northern biotype were collected for future work in 2011.
Table 4. Currently known host range of the two biotypes of Aulacidea pilosellae compiled from European literature data, field surveys done in 2010 and previous work by CABI E-CH.
Host plant species Literature Univoltine biotype (northern range)
Bivoltine biotype (southern range)
Natural field host
Galled in lab tests
Natural field host
Galled in lab tests
P. officinarum1 yes yes2 no yes yes
P. caespitosa1 – yes yes – no
P. glomerata1 – yes yes – no
P. aurantiaca1 – yes2 yes – yes
P. floribunda1 yes yes – – –
P. piloselloides1 – yes yes – yes
P. piloselloides ssp. bauhinii1 – yes – – –
P. flagellare1 yes yes – – –
P. cymosa yes yes – – –
P. macranthela – yes2 – – –
P. echioides yes yes – – –
1 Invasive in North America;
2 only found at one locality; “–“ indicates that no information was found on
this hawkweed species in the literature, that it was not present in the field surveys or that it was not tested in the laboratory.
The results of Chandra’s field research, previous field collections and physiological host-range testing done at CABI E-CH demonstrate that the two biotypes of the gall wasp appear to have different host ranges with the bi-voltine one being more restricted (Table 4). Data from previous years for the northern biotype, and 2010 data for the southern biotype also indicate that the northern biotype is largely made up of sexual populations, and the southern of asexual populations. Emergence data from the 2010 northern biotype field collections will be compiled to investigate this further.
Chandra’s field research has provided additional evidence that indicates that the two biotypes may actually be two cryptic species or subspecies. She is now employing DNA barcoding to determine the level of genetic differentiation between the two biotypes of the gall wasp. Sequencing of three gene regions – one mitochondrial (cytochrome oxidase 1) and two nuclear (the D2 region of 28S and internal transcribed spacer 2) – is currently underway for multiple populations of each

11
biotype. Both biotypes are also being screened for the presence of the bacterial endo-symbiont Wolbachia which, if present, may be responsible for the differences in sex ratios observed in the two biotypes. Chandra will continue the molecular work and data analysis from the 2010 field study during the summer of 2011.
8. Proposed work programme for 2011
Aulacidea subterminalis (Hymenoptera: Cynipidae)
Conduct mass rearing for shipment to North America;
Collect additional galls in the Black Forest, Germany.
Aulacidea pilosellae (Hymenoptera: Cynipidae)
Conduct no-choice and choice gall development tests concentrating on native North American hawkweed species not tested yet;
Field collect galls of the univoltine and bivoltine biotype.
Cheilosia urbana (Diptera: Syrphidae)
Prepare and submit petition for field release.
9. Acknowledgements
We would like to thank Dr Mark Schwarzländer and Marijka Haverhals (University of Idaho, USA) for sending us native North American test plants, and Christian Léchenne and Florence Willemin (both CABI E-CH) for growing plants. Many thanks are also due to Dr Victor Fursov (Institute of Zoology, National Ukrainian Academy of Sciences, Kiev) for field collection of A. hieracii galls in Ukraine. We greatly appreciated the help by Frantisek Krahulec and Jindřich Chrtek, Jun. from the Czech Institute of Botany, who facilitated Chandra’s field trip to the Czeck Republic. Dr Hariet Hinz (CABI E-CH) kindly reviewed this report. In 2010, work by CABI E-CH was funded by the Ministry of Forests, Lands and Natural Resource Operations and the Montana Weed Trust Fund through Montana State University, while Chandra Moffat received support from the Natural Sciences and Engineering Research Council of Canada and the Irving K. Barber School of Arts and Sciences International Education Travel Subsidy.
10. References
Gleason, H.A. and Cronquist, A. (1991) Manual of vascular plants of northeastern United States and adjacent Canada. Second edition. The New York Botanical Garden, USA.
Hunter, G.G. (1991) The distribution of hawkweeds (Hieracium spp.) in the South Island, indicating problem status. Journal of the New Zealand Mountain Lands Institute Review 48, 21–31.

12
Meeklah, F.A. (1980) Chemical weed control. New Zealand Ministry of Agriculture and Fisheries, Agricultural Research Division, Annual Report 1978/79, 266 pp.
Scoggan, H.J. (1979) The flora of Canada. Part 4 – Dicotyledoneae (Loasaceae to Compositae). In: National Museum of Natural Sciences Publications in Botany, No. 7. National Museums of Canada, Ottawa, pp. 1563–1570.
Syrett, P. and Smith, L. (1998) The insect fauna of four weedy Hieracium (Asteraceae) species in New Zealand. New Zealand Journal of Zoology 25, 73–83.
USDA, NRCS (2010) The PLANTS Database. National Plant Data Center, Baton Rouge, Louisiana, USA. http://plants.usda.gov [accessed April 2010].
Wilson, L.M. and Callihan, R.H. (1999) Meadow and orange hawkweed. In: Sheley, R.L. and Petroff, J.K. (eds) Biology and management of noxious rangeland weeds. Oregon State University Press, Corvallis, USA, pp. 238–248.

13
Distribution list
Siegfried Bräutigam
Rob Bourchier
Rosemarie DeClerck-Floate
Simon Fowler
Viktor Fursov
Alison Halpern
Richard Hansen
Frantisek Krahulec
Boris Korotyaev
Jeff Littlefield
George Markin
Brian Marschman
Peter Mason
Brett Richardson
Mark Schwarzländer
Alexander Sennikov
Lincoln Smith
Susan Turner
Linda Wilson
CABI library (2)


www.cabi.org
KNOWLEDGE FOR LIFE
contact CABI europe CABI Head Office
Nosworthy Way, Wallingford, Oxfordshire, OX10 8DE, UK T: +44 (0)1491 832111 CABI Europe – UK
Bakeham Lane, Egham, Surrey, TW20 9TY, UK T: +44 (0)1491 829080 CABI Europe – Switzerland
Rue des Grillons 1, CH-2800 Delémont, Switzerland T: +41 (0)32 4214870 E: [email protected]
asia CABI South Asia
Opposite 1-A, Data Gunj Baksh Road, Satellite Town, Rawalpindi-Pakistan T: +92 (0)51 9290132 CABI Southeast and East Asia
PO Box 210, 43400 UPM Serdang, Selangor, Malaysia T: +60 (0)3 89432921 CABI South Asia – India
2nd Floor, CG Block, NASC Complex, DP Shastri Marg, Opp. Todapur Village, PUSA, New Delhi – 110012, India T: +91 (0)11 25841906 CABI Southeast and East Asia – China, Beijing Representative Office
Internal Post Box 56, Chinese Academy of Agricultural Sciences, 12 Zhongguancun Nandajie, Beijing 100081, China T: +86 (0)10 82105692
africa CABI Africa
ICRAF Complex, United Nations Avenue, Gigiri, PO Box 633-00621, Nairobi, Kenya T: +254 (0)20 7224450/62
americas CABI Caribbean & Latin America
Gordon Street, Curepe, Trinidad and Tobago T: +1 868 6457628 CABI North America
875 Massachusetts Avenue, 7th Floor, Cambridge, MA 02139, USA T: +1 617 3954051