annual cycle of biomass of a threatened population of the intertidal seagrass zostera japonica in...
TRANSCRIPT

S. Y. Lee
Annual cycle of biomass of a threatened populationof the intertidal seagrass Zostera japonica in Hong Kong
Received: 3 March 1997 /Accepted: 14 March 1997
Abstract The phenology and primary productivity of apopulation of Zostera japonica (Aschers. & Graebn.)threatened by the construction of Hong Kong's newinternational airport were studied over a 12-month pe-riod. The need to conserve the population, and the smallleaf size of Z. japonica rendered traditional destructiveor marking techniques inapplicable for percentage coverand biomass estimation. A nondestructive method basedon image analysis techniques was therefore devised forrepeated estimation of percentage cover, biomass andleaf area index. This technique, which involved randomquadrat sampling, photographic recording and imageanalysis, was able to provide data on the three param-eters with acceptable precision and was cost-e�ective inthe ®eld. Z. japonica demonstrated a strongly seasonalcycle of vegetative growth, with di�erent patterns forleaf density (peak in March) and overall bed area (peakin June). Total (above- and below-ground) net primaryproductivity was estimated at between 344 and 688 gAFDW m)2 yr)1. Percentage cover of Z. japonica wasnegatively correlated with total suspended solids (TSS)in the water column while total bed area was negativelycorrelated with water salinity. Increased sedimentationassociated with the new airport project was identi®ed asone important factor a�ecting the growth of the sea-grass, as TSS reached the high level of �1 g DW l)1
during the ®rst half of the study period. Sediment trapsset in the beds also recorded potential sedimentationrates at between 2.89 and 14.5 mg cm)2 d)1. This highturbidity resulted in a sharp decrease in the density of
Clithon spp., the dominant grazers of epiphytic algae onZ. japonica. E�ects of sedimentation and shading ongrowth of Z. japonica were investigated by ®eld manip-ulative experiments. Experimental increase of sedimen-tation rate and shade both resulted in larger decreases inpercentage cover and above-ground AFDW comparedwith the control.
Introduction
Four species of seagrasses, namely Zostera japonica(Aschers. & Graebn.), Ruppia maritima (L.S.L.), Halo-phila ovata Gaud. and H. beccarii Aschers., have beenrecorded from Hong Kong (Hodgkiss and Morton1978a, b). All four species are intertidal and small in size,e.g., the leaves of Z. japonica are usually <15 cm longwhile the other species have prostrate habits. Such habitsmake these seagrasses highly vulnerable to water columnstresses such as increased sedimentation, one commoncause of extinction of seagrass populations especially inrapidly developing areas such as the southeast Asianregion (Fortes 1988).
Zostera japonica and Halophila ovata are the mostabundant amongst the four Hong Kong seagrasses, butstill are restricted to only a few remote and less-dis-turbed localities in the territory (Lee 1994). Rapid de-velopment of the inshore environment and theassociated terrestrial catchments, together with the useof the o�shore waters for dredging of marine sand forconstruction or dumping of wastes all threaten the sur-vival of the Hong Kong seagrasses. In contrast to theimmense threats faced by these east Asian seagrasses,information on their ecology and biology is virtuallynonexistent. It is known that seagrasses, including thefour Hong Kong species, have fairly wide geographicranges along the Chinese coast (Huang et al. 1994), butvery little other information is available. The paucity ofinformation on the Chinese seagrasses is exempli®ed bythe fact that in the review by Mukai (1993) no infor-mation on the occurrence and distribution of the 17
Marine Biology (1997) 129: 183±193 Ó Springer-Verlag 1997
Communicated by T. Ikeda, Hakodate
S.Y. LeeDepartment of Ecology & Biodiversity andThe Swire Institute of Marine Science,The University of Hong Kong, Pokfulam Road,Hong Kong
Present address:(&) School of Applied Science, Gri�th University Gold Coast,Gold Coast Mail Centre PMB 50, Qld 4217, Australia

tropical seagrass species is available for the Chinesecoast. Basic information on aspects such as phenology,productivity and responses to common stresses, e.g.sedimentation, is therefore urgently needed for the ef-fective conservation of the seagrasses.
In Hong Kong, one of the sites where Zostera japo-nica occurs is under strong threat from constructionwork for a new international airport less than 1 kmaway. Increased sedimentation in the water column hasbeen identi®ed as a potential problem threatening thesurvival of the seagrass at the site. As part of a pro-gramme for the long-term rehabilitation and conserva-tion of seagrasses in Hong Kong, a 12-month study wasconducted on the phenology of growth to investigate thepotential impacts of sedimentation and other threats onZ. japonica near the airport site.
Materials and methods
Study site
All sampling was conducted at San Tau on the northern shores ofLantau Island, Hong Kong (113°55¢E; 22°17¢N). Lantau is thelargest island in Hong Kong and is located in the western sector ofthe territory, under the direct but seasonal in¯uence from the PearlRiver. The Zostera japonica beds occupy a narrow belt of the in-tertidal sand ¯at at �1.3 to 1.5 m above Chart Datum at San Tau.Dwarf mangroves of Kandelia candel, Aegiceras corniculatum,Bruguiera gymnorrhiza and Avicennia marina mark the upper in-tertidal limit of the seagrass belt whereas the lower limit is delin-eated by areas occupied by another seagrass, Halophila ovata. Thestudy site is directly opposite to the location of Hong Kong's newinternational airport currently under construction, separated by
only a narrow (<1 km) strip of water. The beds are thereforestrongly a�ected by disturbances and pollution arising from theconstruction work for the airport, one of the largest civil engi-neering projects currently being undertaken in the world (Fig. 1).
Hydrographic and sediment characteristics
Key hydrographic and sediment characteristics were monitoredregularly during the study period, which lasted from October 1993to September 1994 inclusive. Salinity (&), dissolved oxygen (mg l)1)and solar irradiation (W cm)2) were measured in situ using theappropriate meters. Subsurface water samples were collected forthe determination of total suspended solids. Subsamples of knownvolumes were ®ltered through Whatman GF/C papers, dried at80 °C to estimate total suspended solids, then combusted at 500 °Cfor 3 h to determine total suspended organic matter. Sedimentorganic matter was also measured using the loss-on-ignitionmethod (500 °C for 48 h) after acid pretreatment to remove car-bonate materials. Particle size distribution of the sediment wasdetermined with the dry sieving technique through a set of eightstandard sieves ranging from 8 mm to 63 lm mesh size.
A nondestructive method for the determinationof percentage cover and biomass
In order to monitor performance of the Zostera japonica popula-tion at San Tau, a nondestructive method was devised for the es-timation of performance indicators such as percentage cover andbiomass. Nondestructiveness of the method was essential, as thesmall beds (total area <160 m2) would not tolerate repeated ap-plication of the usual destructive methods, e.g., biomass harvesting(English et al. 1994). Other nondestructive methods such as themarking technique of Zieman (1974) are not easily applicable ± thesmall width (<2 mm) of the leaves of Z. japonica makes any formof marking di�cult. The method used in the present study, there-fore, involved the photographic recording of between 15 and 20randomly located 25 ´ 25 cm2 quadrats in the beds during low tide
Fig. 1 A map showing the environs of the site at San Tau (ST ),Lantau Island, Hong Kong. The Zostera japonica beds, where thestudy was conducted, were located <1 km from the construction siteof Hong Kong's new international airport on reclaimed land around
Chek Lap Kok Island (CLK, square enclosure on the map). See insetfor details of the local environment where Z. japonica occurs. Thelocation of the other major known population of Z. japonica at LaiChi Wo (LCW ) is also indicated
184

and subsequent application of image analysis techniques to esti-mate percentage cover of the seagrass.
Random quadrats (area = 0.0625 m2) were located in theseagrass beds during low tide. A colour photograph was taken foreach quadrat sample, and no other measurements were necessary inthe ®eld. Zostera japonica usually forms monospeci®c patches onthe open sediment and is thus amenable to photographic recording.Sometimes objects which may interfere with the image analysisprocess, e.g., pneumatophores of the mangrove Avicennia marina,had to be removed before taking the photograph. Leaf blades ofZ. japonica lie ¯at on the sediment surface during the low tide.
The photographs were then loaded into a calibrated LeicaQuantimet 500+ Image Analyser through a colour video camera,and the total area of the seagrass within the quadrats was estimatedusing colour detection and assisted by manual editing when nec-essary. Since Zostera japonica usually forms monotypic belts in themid-intertidal, the identi®cation of the seagrass-covered areas wasrelatively straightforward. Percentage cover of the seagrass wassubsequently calculated by the image analyser. Mean percentagecover was then determined from the replicate quadrats for eachsampling date. Sampling was made from biweekly to monthly in-tervals during the 12-month period.
Percentage cover values were converted to above-ground bio-mass readings in both dry (DW) and ash-free dry weight (AFDW)by a conversion factor obtained by drying and combusting knownareas of leaf blades. In order to estimate the actual percentagecover, the image analyser has to be calibrated such that a conver-sion factor relating actual and measured percentage cover could beobtained. This conversion was necessary as considerable overlap-ping of the leaf blades occurred when photographed during the lowtide. It was also expected that the relationship between actual andmeasured percentage cover might change for di�erent actual den-sities of leaves. In order to determine the relationship, ®ve quadratscovering a wide range of percentage coverage (9.5 to 44.2%) of theseagrass were photographed and then harvested. Actual above-ground biomass (both DW and AFDW) was measured from theharvested samples and percentage cover estimated following thetechnique described above. A calibration curve was constructed toestablish the relationship between biomass ®gures estimated usingthe image analysis technique and the actual values.
The precision of the method was also determined by taking ®vephotographs each of ®ve random quadrats of the seagrass bed. Theseagrass in the quadrats was manually reorientated before eachphotograph was taken. Percentage cover was then estimated fromthe ®ve photographs to evaluate the importance of orientation ofthe leaf blades, e.g., by tide and wave action, on the estimatedpercentage cover value.
The ratio of above- to below-ground biomass was determinedby collecting both components from ®ve 25 ´ 25 cm2 quadrats.The measurement was repeated for summer and winter periods soas to provide conversion factors which may vary according toseason. The calori®c value of the various components of the sea-grass was determined using a Parr 1200 bomb calorimeter.
Direct measurement of net above-ground primary productivity(NAPP, in g m)2 yr)1) of Zostera japonica was di�cult usingnondestructive techniques. Theoretically, NAPP can be estimatedfrom the following equation:
NAPP �X�DB� E � D� H� (Kemp et al. 1990),
where B is biomass in DW or AFDW (in g m)2, similarly below),DB is increase in biomass between successive measurements, E isbiomass loss due to excretion, D is biomass loss due to death andHis biomass loss due to herbivory. Since it is di�cult to apply themarking technique to the small leaf blades of Z. japonica, D couldnot be measured directly. H is assumed to be small for Z. japonicaas ®eld observations suggested few, if any, herbivores grazed on thespecies. Grazing gastropods such as Clithon spp. and Batillaria spp.utilized the epiphytic algae on Z. japonica but not the seagrassitself. E is assumed to be small compared with the overall biomasschanges. In order to estimate NAPP from changes in standingbiomass (DB), the turnover factor of the same species (4 to 8 times)measured by Harrison (1982) was adopted for estimation of NAPP.
NAPP of Z. japonica at San Tau was thus estimated to be between4 and 8 times SDB.
Since Zostera japonica normally grows through vegetativepropagation, the overall size of the beds can also serve as an in-dicator of the condition of the population. The total areal extent ofthe beds was measured from January 1994 to also make use of thisparameter as an indicator of performance. The areal extents of theZ. japonica patches (there are four major patches which occupyconsiderable areas at San Tau) were measured by marking out thecorners of the beds and taking measurements of the dimensions oftriangles, rectangles or other geometric shapes which would allowan overall approximation of the size of the beds. The total area ofthe beds was then worked out as the aggregate of the individualpatches.
In¯uence of physical conditions on growth
The relationship between physical conditions (turbidity measuredby total suspended solids, total suspended organic matter, dissolvedoxygen level and salinity) and two performance indices (percentagecover and total bed area) was explored by correlation analysis.Pearson correlation coe�cients were calculated using the monthlydata.
The amount of sediment accumulating on the leaf blades ofZostera japonica was estimated by carefully washing all surfaceparticles o� a large number (n = 84) of leaf blades and collectingthe particles on preweighed GF/C ®lters (1 lm pore size). The areaof leaf blade from which the particles was collected was measuredby image analysis technique using the same set up as above. Theamount of sediment on the leaves was then calculated in terms ofdry weight per unit leaf area. The density of Clithon spp., thedominant grazer of ephiphytes on Z. japonica, was measured bycounting all individuals of the gastropod in the random quadratsfor percentage cover estimation. In order to further assess the im-portance of sedimentation as a stress on the Z. japonica population,sediment traps were set up to measure the potential rate of sedi-mentation. Sediments deposited on the surface of leaf blades werealso estimated to give an indication of the scale of the problem.Four cylindrical sediment traps (dimensions: 60 mm diameter,80 mm height) were set up in the Z. japonica beds at San Tau in lateJune 1994. These sediment traps allowed the estimation of thepotential sedimentation rate, i.e. the maximum amount of sedimentwhich may be deposited on unit area of the substratum withoutsigni®cant resuspension. The traps have a cross-sectional area of28.3 cm2. Traps were set in situ to collect sediments from the watercolumn between successive visits, the interval of which was typi-cally between 7 and 10 d. The sediments collected in the traps werewashed out with a volume of distilled water, ®ltered on GF/C paper(pore size »1 lm) and dried to give sedimentation rate in grams dryweight per centimetre per day. Collection was conducted throughthe end of the study period.
The e�ect of increased sedimentation and solar radiation in-tensity on growth and performance of Zostera japonica was alsostudied using manipulative experiments. Eight patches of the sea-grass each of 0.0625 m2 were randomly selected from the beds atSan Tau. Four of the patches were assigned as treatment patcheswhile the other four were controls. Treatment took the form ofweekly addition of 1 g DW of resuspended silt particles to thepatches during low-tide periods. The silt particles were prepared bywet sieving of sediment collected from the same site through a63 lm mesh. The silt particles were dispersed in about 1 litre ofdistilled water and poured evenly onto the experimental patchesduring low tide. In order to minimize loss of the particles to areasoutside the patches, a rectangular frame of the same size as thepatch and 15 cm tall was positioned on the patch to preventspillage during the application. This addition of sediment particleswas equivalent to a mean sedimentation rate of 2.29 mg dry siltcm )2 d)1 in addition to the background sedimentation rate.
The performance indices of percentage cover and above-groundAFDW were used to compare the treatment and control patches.The two parameters were determined using the same image analysistechnique described above. Percentage change in the two perfor-
185

mance indices of the control and treatment patches between thestart and end of the experiment was compared by a Mann±Whitneytest.
The e�ect of heat stress on performance was investigated usinga similar experimental design. The intensity of solar irradiation onpatches of the seagrass was manipulated by erecting squares ofblack plastic meshes of two ``pore'' sizes [10 mm, ``half-shade''(HS); and 2 mm, ``full-shade'' (FS)] over seagrass patches of0.0625 m2 area. Four random patches each were assigned to thetwo shade treatments. In order to minimize the amount of distur-bance to the beds, the same four control patches were used for thesedimentation treatment. The meshes were maintained in positionat 10 cm above the beds by four thin sticks at the corners of thepatch. Preliminary measurements conducted over a wide range oflighting conditions in the ®eld suggested that on average irradiance(in W cm)2) was reduced by 55.7 and 64.7%, respectively, for theHS and FS treatments. This reduction in irradiation also resulted ina temperature decrease of between 1 and 2 °C on the surface of theexperimental patches compared with the control. Performance ofthe patches was compared at the end of the 3-month experimentalperiod by the same method outlined above.
Results
Physical conditions at the study site
Salinity at the site varied between 35 (winter) and 5&(summer), re¯ecting the strong in¯uence of the PearlRiver (Fig. 2a). The pattern of turbidity in the watercolumn did not, however, correspond directly with theexpected pattern of riverine in¯uence, i.e. high sedimentloads between June and September. Total suspendedsolids peaked at �1 g DW l)1 in October 1993 (Fig. 2b),when salinity was relatively high, suggesting that thesource of suspended material was something other thansediment of riverine origin. The values declined gradu-ally to much lower levels (<0.2 g l)1) thereafter, apartfrom a few sporadic peaks in May and July 1994. Theanthropogenic nature of the suspended material in the
Fig. 2 Variations in a salinity, b total suspended solids (TSS), cpercentage organic matter in suspended solids and d dissolved oxygenof the water column at San Tau during the study period. Note that the
peak of TSS did not correlate with the trough for salinity, suggestingthat the sediment was probably of anthropogenic rather than riverineorigin
186
initial half of the study period (October 1993 to April1994) is re¯ected by the low organic content, generally at<10% of the dry weight (Fig. 2c). Percentage organiccontent of the suspended solids demonstrated a signi®-cant increasing trend during the study period(r = 0.613, P < 0.001), probably mirroring a gradualshift in dominance from anthropogenic to natural (e.g.phytoplankton) sources of particles in the latter half ofthe 12-month period. The large amount of suspendedmatter in the water column resulted in heavy light at-tenuation. Between 60 and 95% of surface irradiancewas attenuated at 30 cm beneath the surface during theperiod May to July 1994. The fact that the dissolvedoxygen level of the water remained high (6 to 10 ppm)during the study period (Fig. 2d) further suggests therespective dominance by inorganic, anthropogenic sus-pended matter and phytoplankton in the two halves ofthe study period. The large amounts of sediment in late1993 probably arose from the reclamation and dredgingwork for the airport construction project.
The sediment at San Tau was dominated by coarse tomedium sand of sizes between 2 mm and 250 lm (/= )1 to 2). The silt-clay fraction (<63 lm, / ³ 4) onlycontributed 0.27% of the sediment in terms of dryweight at the start of the study but increased to a sig-ni®cantly higher ®gure of 1.05% by January 1994.
Nondestructive estimation of percentage coverand biomass using image analysis techniques
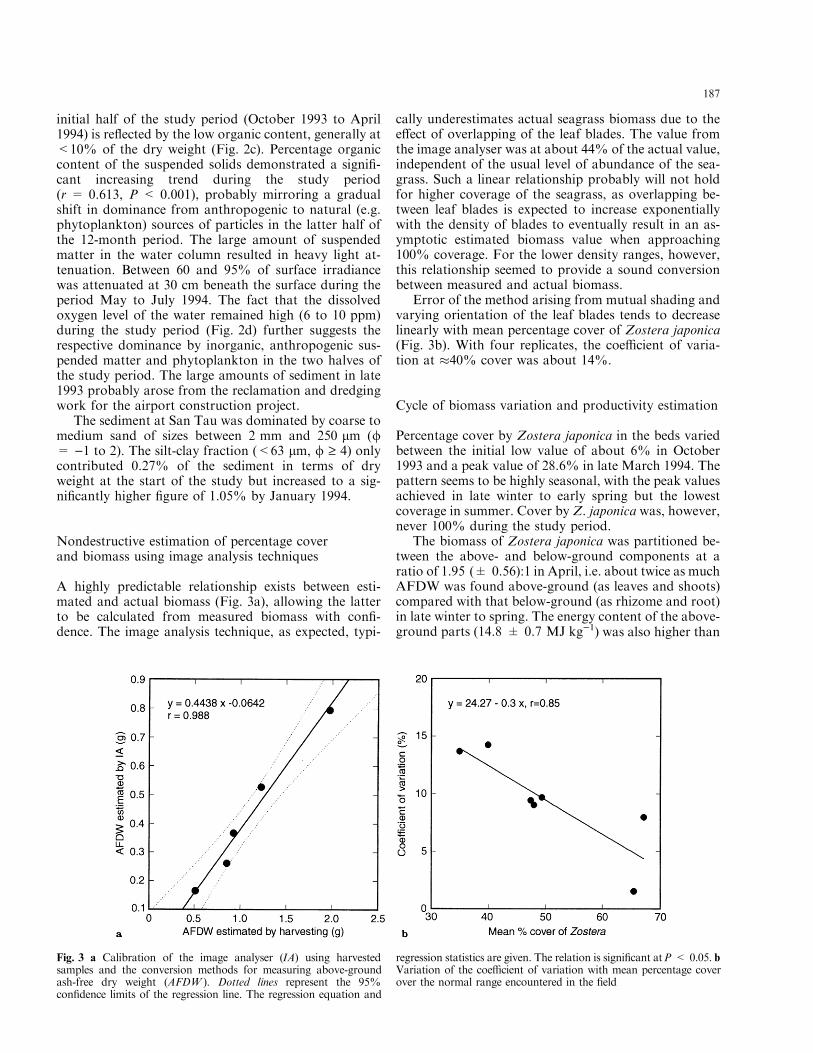
A highly predictable relationship exists between esti-mated and actual biomass (Fig. 3a), allowing the latterto be calculated from measured biomass with con®-dence. The image analysis technique, as expected, typi-
cally underestimates actual seagrass biomass due to thee�ect of overlapping of the leaf blades. The value fromthe image analyser was at about 44% of the actual value,independent of the usual level of abundance of the sea-grass. Such a linear relationship probably will not holdfor higher coverage of the seagrass, as overlapping be-tween leaf blades is expected to increase exponentiallywith the density of blades to eventually result in an as-ymptotic estimated biomass value when approaching100% coverage. For the lower density ranges, however,this relationship seemed to provide a sound conversionbetween measured and actual biomass.
Error of the method arising from mutual shading andvarying orientation of the leaf blades tends to decreaselinearly with mean percentage cover of Zostera japonica(Fig. 3b). With four replicates, the coe�cient of varia-tion at �40% cover was about 14%.
Cycle of biomass variation and productivity estimation
Percentage cover by Zostera japonica in the beds variedbetween the initial low value of about 6% in October1993 and a peak value of 28.6% in late March 1994. Thepattern seems to be highly seasonal, with the peak valuesachieved in late winter to early spring but the lowestcoverage in summer. Cover by Z. japonica was, however,never 100% during the study period.
The biomass of Zostera japonica was partitioned be-tween the above- and below-ground components at aratio of 1.95 (� 0.56):1 in April, i.e. about twice as muchAFDW was found above-ground (as leaves and shoots)compared with that below-ground (as rhizome and root)in late winter to spring. The energy content of the above-ground parts (14.8 � 0.7 MJ kg)1) was also higher than
Fig. 3 a Calibration of the image analyser (IA) using harvestedsamples and the conversion methods for measuring above-groundash-free dry weight (AFDW ). Dotted lines represent the 95%con®dence limits of the regression line. The regression equation and
regression statistics are given. The relation is signi®cant atP < 0.05. bVariation of the coe�cient of variation with mean percentage coverover the normal range encountered in the ®eld
187

that of the below-ground parts (14.1 � 0.3 MJ kg)1) inApril. These ®gures suggest that the overall ratio of theenergy content of the above-ground component to thatof the below-ground components was equal to 2.05:1 inApril.
In the summer (August), the picture was signi®cantlydi�erent. The energy content of the rhizomes and roots(15.2 � 0.2 MJ kg)1) was signi®cantly higher that of theabove-ground parts (14.3 � 0.3 MJ kg)1). The ratio ofabove-ground to below-ground AFDW in August alsochanged to 0.86:1, re¯ecting the change to a dominanceof the below-ground component. The overall energyvalue ratio was 0.81:1 (above-:below-ground compo-nents). There is therefore a dynamic shift of energy re-serves in Zostera japonica during di�erent seasons,which may be a result of either a natural phenology of
growth or the response towards stress imposed upon thepopulation.
Above-ground biomass (AFDW) and variation inpercentage cover of Zostera japonica followed a distinctseasonal pattern (Figs. 4, 5). Peak values of above-ground AFDW and percentage cover were achieved inApril whereas the highest areal extent of the bed wasrecorded in early June. The troughs for all three pa-rameters seem to be in winter, between October toJanuary. The large annual range of variation of theseparameters suggests a highly dynamic seasonal growthpattern.
Values of S(DB) obtained for the ®xed and randomquadrat samples were 53.24 and 47.28 g AFDW m)2,respectively. Assuming a turnover factor of between 4and 8 annually, these ®gures translate to NAPP valuesof 212.96 to 425.92 and 189.12 to 378.24 g AFDWm)2 yr)1. Taking the mean values for these two datasets, the range of 201.04 to 402.08 g AFDW m)2 yr)1 isobtained. Using the conversion ®gures obtained above,below-ground productivity is estimated to be 143.09 to286.18 g AFDW m)2 yr)1, giving a total productivity of344.13 to 688.26 g AFDW m)2 yr)1.
Expansion of the Zostera japonica beds was, again,seasonal. The peak areal extent of the beds occurred inlate May to early June, closely following that for per-centage cover and above-ground biomass (April). Totalcoverage of the beds varied between a low of 79 m2 inFebruary 1994 and the usual values of >120 m2 formost other times of the study period (Fig. 6).
E�ect of shade and sedimentation on growth
The potential sedimentation rate varied between 14.5and 2.9 mg cm)2 d)1 during June to September 1994(Table 1). The sedimentation rate so recorded seemed
Fig. 4 Zostera japonica. Variations in above-ground ash-free dryweight (AFDW ) of Z. japonica at San Tau during the study period
Fig. 5 Zostera japonica. Variations in percentage cover of theZ. japonica population at San Tau during the study period
Fig. 6 Zostera japonica. Variations in total coverage of Z. japonicaon the intertidal sand¯at at San Tau during the study period
188

to correlate positively with the total suspended matterin the water column recorded for the same period(Fig. 2b). The average amount of sediment particles onthe leaves was estimated to be 4.52 mg DW cm)2 leafarea (only one surface used in the calculation). Theleaves therefore had signi®cant sediments deposited ontheir surfaces.
The importance of this sedimentation on the seagrassbeds can be seen also through its e�ect on grazers tap-ping the epiphytes on Zostera japonica leaf blades. Themost common grazers are the gastropods Clithon spp.,which occur at high densities in healthy Z. japonica beds.The density of Clithon spp. actually exhibited a distinctdecreasing trend from March 1993 (Fig. 7), falling fromthe high values of between 25 and 40 to <3 individualsper 0.0625 m2 by July 1994. Since the trough valuesactually fell far below those recorded for the same timein the previous year, there are reasons to believe thatthese grazers had been seriously impacted by, probably,sedimentation on the Z. japonica leaf blades.
Results of Pearson correlations between percentagecover, total bed area and above-ground biomass and thefour major environmental parameters measured suggesta negative relationship between total suspended solids inthe water column and Zostera japonica performance,and a similar relationship between total bed area andsalinity (Table 2).
Seagrass percentage cover and above-ground AFDWboth decreased during the 3-month experimental periodin all (treatment and control) experimental patches.Although increased sedimentation did result in generallylower percentage cover and above-ground AFDW in the
treatment patches (Table 3), the di�erences were notstatistically signi®cant. The large variance among theplots probably reduced the power of the test with respectto the magnitude of the treatment e�ect.
Similar to the sedimentation experiment, results ofthe shading experiment suggest larger mean decrease inpercentage cover and above-ground AFDW in theshaded compared with the control plots (Table 3), butthe di�erence was again found to be statisticallynonsigni®cant.
Table 1 Potential sedimentation rate (in mg cm)2 d)1 �1 SD) asmeasured by sediment traps in the Z. japonica beds at San Tau forthe period June to September 1994
Date Sedimentation rate
23±27 Jun 14.52 � 9.6927 Jun±8 Jul 6.00 � 2.498±19 Jul 3.51 � 1.061±8 Aug 2.89 � 1.1023±30 Aug 7.09 � 3.7930 Aug±15 Sep 6.45 � 1.19
Fig. 7 Changes in the density of the gastropod Clithon spp. whichgrazed on the epiphytic algae on leaves ofZ. japonica during the studyperiod
Table 2 Pearson correlation coe�cients relating the two perfor-mance indices (percentage cover and above-ground AFDW) andthe hydrographic parameters. Only total suspended solids (TSS)has a signi®cant negative correlation with percentage cover ofZ. japonica (DO dissolved oxygen; TSOM total suspended organicmatter; * signi®cant correlation at P < 0.01)
Parameter % cover Total bed area AFDW
DO 0.5314 0.1129 0.3110Salinity )0.0041 )0.6986* 0.3227TSS )0.7096* 0.1610 )0.5224TSOM )0.5041 0.1033 )0.2919
Table 3 Zostera japonica. Re-sults of the manipulative ex-periment on the e�ects ofincreased sedimentation andreduced irradiation on the per-formance of Z. japonica. Datashown are means � SD per-centage change between the in-itial and ®nal values for theplots. Despite the more pro-nounced mean changes (de-crease) in both indices in theexperimental plots, no sig-ni®cant di�erence can be de-tected between the responsesgiven by the treated and controlplots (NS nonsigni®cant)
Performance Control patches Treatment patches Mann± Pindex (%) (%) Whitney U,W
Increased sedimentation% cover )25.1�34.7 )42.7�35.9 7, 17 0.77, NSAbove-groundAFDW (g m)2 )
)46.5�31.9 )67.0�19.4 5, 15 0.39, NS
Full-shade Half-shade Kruskal± Ppatches (%) patches (%) Wallis v2
Reduced irradiation% cover )25.1�34.7 )49.8�29.7 )48.9�15.5 2.00 0.37, NSAbove-groundAFDW (g m)2 )
)46.5�31.9 )56.3�31.8 )69.4�11.4 1.19 0.55, NS
189

Discussion
Image analysis techniques have been applied to the es-timation of macrophyte distribution and extent (Leh-mann et al. 1994; Long et al. 1994) and seagrass leafvolume (Gerber et al. 1994). Most of the previous ap-plications to biomass estimation involved extensive bedsand the use of aerial photography. Whereas these tech-niques can provide a rapid estimation of biomass inthese large-scale studies, the method reported uponherein aims at measuring gradual, within-bed variationsin biomass for populations with limited areal extent.This image analysis technique allows nondestructive,replicate measurements of percentage cover and biomassof threatened seagrasses. The method has the advantageof being particularly applicable to small-size species suchas members of the genera Zostera and Syringodium,which are not amenable to traditional marking tech-niques. Since this method does not rely on personaljudgement, e.g., of abundance as in the method ofMellors (1991) and similar methods summarised in En-glish et al. (1994), it is less prone to errors due to dif-ference in experience of the observer and also requiresless cross-calibration. It is also comparatively moretime-e�cient than the visual technique of Mellors(1991).
Calibration is a crucial step in the validation of suchindirect methods. It seems from the example of the HongKong Zostera japonica that linearity is maintained be-tween actual and estimated biomass for the normallevels of seagrass abundance. Caution has to be exer-cised when working on dense beds as the method mayincreasingly underestimate biomass. As a matter of fact,most if not all nondestructive methods, e.g. the visualmethod of Mellors (1991), will encounter the sameproblem while estimating percentage cover and biomassof extremely dense beds. This is not usually a practicalproblem as such luxurious beds are probably capable oftolerating some destructive sampling.
The values obtained for Zostera japonica from thismethod are within the broad range obtained for sea-grasses in the literature, e.g., for Z. marina (Aioi et al.1981; Roman and Able 1988); Posidonia australis (Westand Larkum 1979) and Z. noltii and Cymodocea nodosa(van Lent et al. 1991) but are signi®cantly lower thanthose recorded by Mukai et al. (1979) for Z. marina inJapan and by Gallegos et al. (1993) for Thalassiatestudinum in Mexico. The values from these studieswere, however, obtained from a variety of methods andfrom species with distinct growth forms or sizes. Thesecan therefore only serve as rough reference ®gures.
Using the average energy value obtained from theApril and August samples (14.6 MJ kg)1), the NAPPvalue is equivalent to 2.9±5.9 MJ m)2 yr)1. Again, usingthe above- to below-ground biomass energy ratio of1.43:1, below-ground energy production is calculated tobe 2.1 to 4.1 MJ m)2 yr)1, giving a total productivity of5.0 to 10.0 MJ m)2 yr)1. As the beds almost grew to
double (a 90.8% increase between February and May1994) the original size, a generally acceptable level ofperformance is indicated.
Performance and biomass cycle of Zostera japonica
Strong seasonality of growth has widely been docu-mented for various seagrasses especially for populationsat higher latitudes, e.g., for Zostera marina (Aioi 1980),Z. japonica and Ruppia maritima (Harrison 1982), andSyringodium ®liforme (Short et al.1993). Similar to mostother seagrasses, e.g., Z. muelleri (Kerr and Strother1989, 1990), Z. noltii (Vermaat et al. 1987), Thalassiatestudinum (MarbaÁ et al. 1994), and Thalassodendronpachyrhizum (Kirkman and Cook 1987), Z. japonicademonstrates a strong seasonality in vegetative growthand reproduction. Thus large seasonal variations havebeen observed in performance indicators such as per-centage cover and the total areal extent of the beds.Sexual reproduction is also limited to a brief period inthe year, between March and May. Although the pop-ulation at Lai Chi Wo (an undisturbed location withZ. japonica distribution in Hong Kong) had not beenfollowed in detail, it was observed that this populationhas the same ¯owering time as the San Tau population.
Although most seagrasses tend to exhibit strongseasonality in growth and reproduction, the apparentphenology of the Zostera japonica population at SanTau could be a result of the superimposition of checks(and release) on growth imposed by external, anthro-pogenic, stress factors on the natural seasonal cycle.
An evaluation of the level of performance of theZostera japonica population at San Tau is di�cult dueto the general lack of comparative data. This study is the®rst comprehensive endeavour to investigate the ecologyof any seagrass in Hong Kong. While seagrasses, in-cluding Z. japonica, are known to occur on the coast ofmainland China, there is apparently no accessible liter-ature on their ecology. The case of Z. japonica furthersu�ers from the wide latitudinal distribution of thespecies, any comparative data available are, of necessity,from very di�erent latitudes. The interpretation of theperformance data is therefore largely speculative.
The general level of performance of Zostera japonicaat San Tau is compared with published data on per-centage cover, above-ground biomass and NAPP forother seagrasses belonging to the same genus or those inthe same geographic region (Table 4). It can be seenfrom the table that levels of the four performance indi-cators for the Z. japonica population at San Tau areinferior to those of the other seagrasses reported in theliterature. Whereas this is probably partly explicable bythe morphology of the local species, i.e. a narrow, short,leaf blade, the readings are still low compared to otherspecies of the same genus, e.g. Z. noltii, and species ofSyringodium, which have equally short and narrow leafblades (Phillips and MenÄ ez 1988). Despite the relativelylarge volume of data on seagrass production ecology
190

and growth dynamics, there is yet no established rela-tionship between production and other indices of per-formance and latitude. Duarte (1989) explored possiblelatitudinal e�ects on the seasonality of seagrass biomassand the production to peak biomass ratio. Z. japonica atSan Tau demonstrates two apparent deviations from thegeneral patterns established by this author: (a) the peakof biomass occurs in spring (March) rather than insummer (July to August) and (b) the annual productionvalue (201 to 402 g AFDW m)2 yr)1) is much higherthan that predicted from peak biomass by the regressionequation (123 g DW m)2 yr)1) generated based on11 species of seagrasses occurring from 9 to 52° latitude.Direct application of Duarte's expectation of the pro-duction/biomass (P/B) ratio to Z. japonica at San Tau is,however, questionable, as (a) 12 out of the 14 data setsused for the calculation of the annual P/B ratio werefrom higher (>22°) latitudes and (b) all data points hadhigher (>50 g DW) biomass than the local Z. japonica.Nevertheless, Duarte (1989) pointed out the importanceof environmental conditions on the particular patternsof biomass seasonality and annual P/B ratio. Localdi�erences in environmental conditions could oftenmask any latitudinal trend in biomass seasonality, and ahigh P/B ratio may be indicative of the need to coun-teract epiphytic accretion by leaf sloughing (Wahbeh1984). The lack of a comprehensive model for growthand production of seagrasses along latitudinal gradients,and, especially, one incorporating the data from thepoorly studied West Paci®c tropics, makes interpopula-tion comparison of performance di�cult. This is par-ticularly true in the case of species with wide geographicdistributions such as Z. japonica. Notwithstanding such
confounding factors, the wide di�erences in the fourperformance indices between the local Z. japonica andits congeners and other species suggest that the condi-tion of the San Tau population is not indicative of ahealthy and thriving population.
The phenology of growth and reproduction of mostintertidal organisms in Hong Kong is strongly in¯u-enced by the seasonal exposure to solar irradiation(Williams 1994). Thus intertidal macroalgae only pro-liferate in the intertidal region during the summermonths whereas little growth is discernable in the sum-mer. This seasonality is attributed to one characteristicfeature of the Hong Kong tidal cycle: the spring lowtides usually occur in the late afternoon in the summermonths, at the time when solar irradiation is strongeston the shore. This intense heat can lead to massivemortality of animals on the mid to high shore, andprevent the colonization of these areas by most macro-algae. The same stress can be expected for the intertidalsoft-bottom organisms such as Zostera japonica.
One feature of the growth cycle of Zostera japonica atSan Tau is the asynchrony of maximal percentage coverand bed size (Figs. 4, 5, 6). A sharp decline in percentagecover (and thus above-ground biomass) was evident inlate April, accompanied by a rapid increase in overallbed area. This probably re¯ects a shift in biomass allo-cation from the above-ground (leaves and shoots) to thebelow-ground (rhizomes and roots) components duringsummer.This change in allocation pattern is suggestedby a signi®cant di�erence in the above- to below-groundbiomass ratio revealed by the destructive samples col-lected in April (1.96:1) and August (0.86:1), respectively.Such a rapid inversion of biomass allocation has rarely
Table 4 Published data on the performance of seagrasses com-pared with that recorded for the Z. japonica population at San Tau,Hong Kong. All values apart from the above-ground primary
productivity (NAPP) refer to peak values; all biomas and NAPPvalues are AFDW except otherwise stated (ND no data)
Species % cover Biomass NAPP Source(g m)2) (g m)2 yr)1)
Zostera japonica 28.6±32.4 28.7±34.2 201±402 Present studyThalassodendron pachyrhizum ND 500 ND Kirkman and
(dry weight) Cook (1987)Thalassia testudinum ND >1000 1350±4050 Gallegos et al.
(dry weight) (dry weight) (1993)Syringodium ®liforme ND ND 664 Short et al.
(dry weight) (1993)Syringodium isoetifolium ND 519 ND Aioi and
(dry weight)a Pollard (1993)Zostera noltii 100 118.7 212 van Lent et al.
(1991)Zostera marina ND 26.8 548 Aioi et al.
(dry weight) (dry weight)b (1981)Z. marina ND 221.6 2628 Mukai et al.
(dry weight) (dry weight)c (1979)Z. marina ND 305 987 Roman and
(dry weight) (dry weight) Able (1988)Z. marina ND 192.8 ND Aioi (1980)
(dry weight)
a Includes above- and below-ground componentsb No indication of whether source refers to above-ground or total productionc For the spring/summer peroid only
191

been reported for seagrasses. This inversion may be re-lated to the desiccation and heat stresses experienced bythe intertidal Z. japonica, imposed by the occurrence ofthe lowest tides in the hottest part of the day in the HongKong summer (between May and September). McMillan(1984) compared the thermal tolerances of seven generaof tropical seagrasses and concluded that a narrow andsmall leaf morphology is strongly correlated with successin shallow tropical waters. Z. japonica probably dem-onstrates this relationship well by being able to surviveintertidally in the summer heat of Hong Kong.
Sedimentation has been suggested to be one of themain causes of the global decline of seagrasses. Tur-bidity arising from eutrophication and high sedimentloadings (from dredging and reclamation works) hasbeen identi®ed as the major cause of decline and failureof recovery of Zostera marina in the Dutch Wadden Sea(Giesen et al. 1990). Fortes (1988) also pointed to theimportance of increased sedimentation rates on thesurvival of seagrasses in the Indo-Paci®c region. Tur-bidity reduces photosynthesis (Libes 1986) and may in-crease vulnerability of the seagrass to infection such as``wasting disease'' (Vergeer and den Hartog 1991). Di-rect deposition of sediments on the leaf blades will alsoreduce epiphytic productivity, which is often an impor-tant component of community production in seagrassbeds. Many consumers which depend on the epiphytesfor food will then be adversely a�ected. The recordeddecline in density of the epiphytic grazer Clithon mayhave been a result of increased turbidity at San Tau. Thehigh sedimentation rate recorded by the traps and thesigni®cant amount of sediment laid on the leaf surfacefurther support this e�ect. The sediment accumulation islikely a result of the airport construction work, as theparticles had a lower organic content and occurrenceduring the dry than the wet season. Nevertheless, theincreasing organic content of the suspended particlessuggests that phytoplankton was probably the cause ofturbidity in the latter half of the study period, also re-ducing the performance of Z. japonica.
Although manipulation of the sedimentation rate andirradiance level did not yield statistically signi®cant dif-ferences between control and treatment patches ofZostera japonica, the lack of signi®cance was probablyattributable to the small sample size rather than a lack ofe�ect. Increased sedimentation as well as reduced irra-diation both caused a decrease in percentage cover andabove-ground AFDW in the treated plots compared tothe control. Vermaat et al. (1993) conducted a similarshading experiment on Z. noltii in Mauritania and adetected signi®cant decrease in leaf growth when shad-ing exceeded 94%. This level of light reduction isthought to have gone below the light compensationpoint for the species. Sample size of the present experi-ment was somehow restricted by the small overall bedsize as well as the need to avoid excess disturbance to thestudy population, a situation probably common tostudies on dwindling populations in nature. The smallsample size also prevented the application of the re-
gression approach to the analysis of the data, whichcould have avoided the problem of unequal treatmentlevels in ANOVA (Vermaat et al. 1993). Similar exper-iments can be repeated with more replication in largerand less-threatened natural beds or with controlledmesocosms in the laboratory to further explore the im-pact of sedimentation and shading on Z. japonica.
Acknowledgements This study was supported by a grant from theAirport Authority Hong Kong. The Director of Agriculture andFisheries is thanked for permission to publish the data collected inthe project. T. Fong o�ered capable ®eld and laboratory assistancethroughout the course of the study. I am also grateful for the as-sistance and enthusiasm of Messrs. C. C. Lay, Y. K. Chan andD. Cheung of the Agriculture and Fisheries Department, HongKong Government, during the study. Comments from two anon-ymous referees also improved the manuscript.
References
Aioi K (1980) Seasonal changes in the standing crop of eelgrass(Zostera marina L.) in Odawa Bay, central Japan. Aquat Bot 8:343±354
Aioi K, Mukai H, Koike I, Ohtsu M, Hattori A (1981) Growth andorganic production of eelgrass (Zostera marina L.) in temperatewaters of the Paci®c coast of Japan. II. Growth analysis inwinter. Aquat Bot 10: 175±182
Aioi K, Pollard PC (1993) Biomass, leaf growth and loss rate of theseagrass Syingodium isoetifolium on Dravuni Island, Fiji. AquatBot 46: 283±292
Duarte C (1989) Temporal biomass variability and production/biomass relationships of seagrass communities. Mar Ecol ProgSer 51: 269±276
English S, Wilkinson C , Baker V (1994) Survey manual for trop-ical marine resources. Australian Institute of Marine Science,Townsville, Australia
Fortes MD (1988) Mangroves and seagrass beds of east Asia:habitat under stress. Ambio 17: 207±213
Gallegos ME, Merino M, MarbaÁ N , Duarte CM (1993) Biomassand dynamics of Thalassia testudinum in the Mexican Caribbe-an: elucidating rhizome growth. Mar Ecol Prog Ser 95: 185±192
Gerber DT, Ehlinger TJ, Les DH (1994) An image analysis tech-nique to determine the surface area and volume for dissectedleaves of aquatic macrophytes. Aquat Bot 48: 175±182
Giesen WBJT, van Katwijk MM, den Hartog C (1990) Eelgrasscondition and turbidity in the Dutch Wadden Sea. Aquat Bot37: 71±85
Harrison PJ (1982) Comparative growth of Zostera japonica Asc-hers. & Graebn. and Z. marina L. under simulated intertidaland subtidal conditions. Aquat Bot 14: 373±379
Hodgkiss IJ, Morton BS (1978a) Zostera nana Roth. (Pot-amogetonaceae) ± a new record for Hong Kong. Mem HongKong nat Hist Soc 13: 23±27
Hodgkiss IJ, Morton BS (1978b) Halophila beccarii Ascherson(Hydrocharitaceae) ± a new record for Hong Kong with noteson other Halophila species. Mem Hong Kong nat Hist Soc 13:28±32
Huang ZG et al. (eds) (1994) Marine species and their distributionin China's Seas. China Ocean Press, Beijing, China
Kemp M, Murray L, McRoy CP (1990) Primary productivity. In:Phillips RC, McRoy CP (eds) Seagrass research methods.UNESCO, Paris, pp 153±159
Kerr EA, Strother S (1989) Seasonal changes in leaf growth rate ofZostera muelleri Irmisch ex Aschers. in south-eastern Australia.Aquat Bot 33: 131±140
Kerr EA, Strother S (1990) Seasonal changes in standing crop ofZostera muelleri in south-eastern Australia. Aquat Bot 38: 369±376
192

Kirkman H, Cook IH (1987) Distribution and leaf growth ofThalassodendron pachyrhizum den Hartog in southern WesternAustralia. Aquat Bot 27: 257±266
Lee SY (1994) Seagrass rehabilitation ®nal report. Report to theHong Kong Government. The Swire Institute of Marine Sci-ence, University of Hong Kong, Hong Kong
Lehmann A, Jaquet J-M, Lachavanne J-B (1994) Contribution ofGIS to submerged macrophyte biomass estimation and com-munity structure modeling, Lake Geneva, Switzerland. AquatBot 47: 99±117
Libes M (1986) Productivity±irradiance relationship of Posidoniaoceanica and its epiphytes. Aquat Bot 26: 285±306
Long BG, Skewes TD, Poiner IR (1994) An e�cient method forestimating seagrass biomass. Aquat Bot 47: 277±291
MarbaÁ N, Gallegos ME, Merino M, Duarte CM (1994) Verticalgrowth of Thalassia testudinum: seasonal and interannualvariability. Aquat Bot 47: 1±11
McMillan C (1984) The distribution of tropical seagrasses withrelation to their tolerance of high temperatures. Aquat Bot 19:369±379
Mellors JE (1991) An evaluation of a rapid visual technique forestimating seagrass biomass. Aquat Bot 42: 67±73
Mukai H (1993) Biogeography of the tropical seagrasses in thewestern Paci®c. In: Pollard PC, Koike I, Mukai H, RobertsonAI (eds) Tropical seagrass ecosystems: structure and dynamicsin the Indo-West Paci®c. CSIRO, Melbourne, Australia, pp 1±17
Mukai H, Aioi K, Koike I, Ohtsu M, Hattori A (1979) Growth andorganic production of eelgrass (Zostera marina L.) in temperatewaters of the Paci®c coast of Japan. I. Growth analysis inspring±summer. Aquat Bot 7: 47±56
Phillips RC, MenÄ ez EG (1988) Seagrasses. Smithson Contr mar Sci34: 1±104
Roman CT, Able KW (1988) Production ecology of eelgrass(Zostera marina L.) in a Cape Cod salt marsh±estuarine system,Masschusetts. Aquat Bot 32: 353±363
Short FT, Montgomery J, Zimmermann CF, Short CA (1993)Production and nutrient dynamics of a Syringodium ®liformeKuÈ tz. seagrass bed in Indian River Lagoon, Florida. Estuaries16: 323±334
van Lent F, Nienhuis PH, Verschuure JM (1991) Production andbiomass of the seagrass Zostera noltii Hornem. and Cymodoceanodosa (Ucria) Aschers. at the Banc d'Arguin (Mauritania, NWAfrica): a preliminary approach. Aquat Bot 41: 353±367
Vergeer LHT, den Hartog C (1991) Occurrence of wasting diseasein Zostera noltii. Aquat Bot 40: 155±163
Vermaat JE, Beiher JAJ, Gijlstra R, Hootsmans MJM, PhilippartCJM, van der Brink NW, van Vierssen W (1993) Leaf dynamicsand standing stocks of intertidal Zostera noltii Hornem. andCymodocea nodosa (Ucria) Ascherson on the Banc d'Arguin(Mauritania). Hydrobiologia 258: 59±72
Vermaat JE, Hootsmans MJM, Nienhuis PH (1987) Seasonal dy-namics and leaf growth of Zostera noltii Hornem., a perennialintertidal seagrass. Aquat Bot 28: 287±299
Wahbeh MI (1984) The growth and production of the leaves of theseagrass Halodule stipulacea (Forsk.) Aschers. from Aqaba,Jordan. Aquat Bot 20: 33±41
West RJ, Larkum AWD (1979) Leaf productivity of the seagrass,Posidonia australis, in eastern Australian waters. Aquat Bot 7:57±65
Williams GA (1994) The relationship between shade and molluscangrazing in structuring communities on a moderately-exposedtropical rocky shore. J exp mar Biol Ecol 178: 79±95
Zieman JC (1974) Methods for the study of the growth and pro-duction of turtle grass Thalassia tetsudinum KoÈ nig. Aquacul-ture, Amsterdam 4: 139±142
193