anatomy of shoots and tumors of in vitro habituated
TRANSCRIPT

616
American Journal of Botany 85(5): 616–628. 1998.
ANATOMY OF SHOOTS AND TUMORS OF IN VITRO
HABITUATED RHODODENDRON ‘MONTEGO’(ERICACEAE) CULTURES WITH TISSUE PROLIFERATION1
ERIC W. MERCURE,2 CYNTHIA S. JONES,3 MARK H. BRAND,2 AND
CAROL A. AUER2
2Department of Plant Science,1376 Storrs Road, U-67, The University of Connecticut, Storrs, Connecticut 06269-4067
Tissue proliferation (TP) is characterized primarily by the formation of galls or tumors at the crown of container-grownrhododendrons that were initially propagated in vitro. In the cultivar ‘Montego’, TP2like symptoms are first observed invitro as shoot clusters with small leaves and nodal tumors. In addition, unlike the normal in vitro non-TP (TP2) shoots, invitro TP (TP1) shoots proliferate rapidly without the presence of the plant growth regulator cytokinin in the tissue culturemedium. Comparisons of the anatomy of TP1 and TP2 shoot tips showed that TP1 shoots had a less developed vascularsystem, longer cells in the pith and cortex, and altered internodal elongation at the shoot apex. In addition, TP1 axillarybuds were abnormal in that they were displaced onto the stem above the leaf axil, and a small group of proliferating cellsreplaced the shell zone at the base of the bud. Initiation of tumor formation began with the expansion of this region of cellproliferation (RCP) and shoot growth from the abnormal axillary bud (tumor bud). Organization of the tumor bud andextension of the RCP characterized the further development of two types of tumors. In polar shoot tumors, shoot growthcontinued from the persistent tumor bud and the tumor at the base of the shoot remained small in size. The RCP extendsdownward to the vascular junction of the subtending leaf and the stem of the TP1 shoot. In nonpolar tumors, continuousde novo meristem formation led to the development of large tumors with or without shoots. The RCP is present throughoutthe tumor and is associated with de novo meristem formation. Comparisons to the anatomy of other tumor-like structuresshowed that TP tumors of Rhododendron ‘Montego’ are most similar to tobacco genetic tumors.
Key words: de novo meristem; Ericaceae; region of cell proliferation; Rhododendron; tissue proliferation; tumor bud.
A significant proportion of rhododendrons are current-ly propagated by tissue culture to provide a large numberof plants of different cultivars in a short amount of time.However, shoots with an altered morphology occasionallyoccur in clonal populations. An abnormal phenotype thathas been recently observed in container-grown rhododen-dron cultivars initially propagated by tissue culture iscalled tissue proliferation (TP). Tissue proliferation ischaracterized by the formation of callus-like or tumorousgrowths at the crown (LaMondia et al., 1992). On somecultivars, the crown tumors produce shoots that are small-er than normal, with short internodes and a whorled ar-rangement of small leaves (Brand, 1992; Linderman,1993; Brand and Kiyomoto, 1997). These disfiguredplants are not marketable. In many affected cultivars, TPis not observed until 1–3 yr after initial propagation, re-sulting in significant financial losses for nurserymen(Anonymous, 1992). However, TP can also occur earlierduring in vitro shoot culture, primarily with the cultivar‘Montego’. In ‘Montego’, shoots with TP-like symptoms
1 Manuscript received 5 May 1997; revision accepted 9 September1997.
The authors thank L. Khairallah and M. Cantino of the Electron Mi-croscope Laboratory of the Department of Physiology and Neurobiol-ogy for their technical help, and Natalie Kane, Jaren Madden, and Mat-thew Opel for their critical reading of the manuscript.
Storrs Agricultural Experiment Station scientific contribution 1744.This work was supported in part by funds made available through theHolden Arboretum, Mentor, OH, and the Connecticut Chapter of theAmerican Rhododendron Society.
3 Author for correspondence, current address: Department of Ecologyand Evolutionary Biology, 75 North Eagleville Road, U-43, The Uni-versity of Connecticut, Storrs, Connecticut 06269-3043.
(TP1 shoots) produce nodal tumors with many lateralbranches and the shoots have an altered leaf morphology.In addition, TP1 shoots are considered to be habituatedsince they grow and multiply rapidly in tissue culturewithout exogenous plant growth regulators in the medi-um, unlike in vitro non-TP (TP2) shoots, which requirecytokinin for shoot multiplication (Brand and Kiyomoto,1997).
Although TP appears similar in morphology to crowngalls, previous experimental evidence suggests that TP isnot normal crown gall development. No infective isolatesof the bacterium Agrobacterium tumafaciens, the causalagent of crown gall, have been found on rhododendronswith TP (Linderman, 1993). Known virulent strains ofAgrobacterium, inoculated by various techniques, havenot caused TP (Brand and Kiyomoto, unpublished data;LaMondia, Smith, and Rathier, 1997). Furthermore, theobservations that shoot formation is not common incrown gall and that TP is not contagious in a rhododen-dron production block suggest that TP is not crown gall(Linderman, 1993).
The morphology of TP also appears to be similar tolignotubers, and it has been theorized that TP results frominduced lignotuber formation. Lignotubers are perma-nent, woody extensions of the crown embedded with dor-mant axillary buds (James, 1984). Although lignotubersand many TP tumors have a woody core surrounded byfleshy cortex tissue (Linderman, 1993), several differ-ences exist between the two structures. TP tumors fromsome cultivars are poorly connected to the stem and areeasily removed, unlike lignotubers, which are integratedinto the stem (Brand and Kiyomoto, 1994). The dormant

May 1998] 617MERCURE ET AL.—IN VITRO TISSUE PROLIFERATION OF R. ‘MONTEGO’
buds in lignotubers regenerate a new canopy only afterthe main stem is removed, such as by fire (James, 1984),but TP tumors produce shoots while the main stem isintact. In addition, when the main stem of TP-affectedrhododendrons is removed, the TP tumors at the crownfail to regenerate permanent canopies (Brand and Kiyom-oto, 1994). Furthermore, regenerated shoots from someTP tumors are abnormal in appearance (LaMondia et al.,1992), and axillary buds are not present in TP tumors onmany cultivars (Linderman, 1993).
To further understand how tumors are formed, the nod-al anatomy of in vitro TP2 and TP1 shoots from Rho-dodendron ‘Montego’ was compared. Shoot tips were in-cluded in the study to determine what tissues/cells con-tributed to the initial formation of tumors, and whethertumor initiation was expressed only at the nodes. Wheth-er de novo meristem formation and/or unorganized pro-liferations of certain cell types contributed to tumorgrowth and development was determined. Finally, thesimilarities and differences in structure between TP tu-mors and crown gall in woody plants, lignotubers, andother tumor-like structures are discussed.
MATERIALS AND METHODS
Plant material—TP2 and TP1 rhododendron shoots of the cultivar‘Montego’ were grown in vitro on woody plant (WP) medium (Lloydand McCown, 1980), pH 5.2, with 3% sucrose, 0.3% agar and 0.1%Phytagel (Sigma Chemical Co., St. Louis, Missouri, USA). TP2 shootcultures were initiated from greenhouse-grown plants that were neveraffected by TP. In order for sustained shoot growth and multiplicationto occur on TP2 shoots, the medium was supplemented with 10 mmol/L of isopentenyladenine (IP). TP2 and TP1 shoots had been main-tained on their respective medium by subculturing at regular intervals(8–10 wk) for 18 mo prior to use in structural studies. During that time,no TP2 shoots had formed TP1 shoots. Variation in growth rates be-tween all shoot cultures was minimized by the placement of culturesunder 25–30 mmol photons·m22·s21 of cool white fluorescent light for16 h at 258C.
Preparation of nodes from in vitro TP2 and TP1 shoots for lightmicroscopy—Three stages of stem and tumor development were deter-mined for TP1 shoot cultures: shoot apices without visible tumors;small tumors (youngest tumor near the stem tip at nodes 7–9); medium-sized tumors with shoots (nodes 10–11); and large tumors with or with-out shoots (oldest tumors at base of stem, nodes 12–14). Nodal positionswere described in a basipetal series from the shoot apex, with the leafmost recently initiated from the shoot apical meristem designated LP1.In TP2 shoot cultures, axillary meristems from shoot apices and excisednodes were from the same nodal position as the small and large tumorsin TP1 shoots.
Four weeks after subculture, 5-mm nodal stem pieces from both cul-tures (with the leaf blade excised in TP2 shoots) were fixed in aldehydefixative (1.5% glutaraldehyde and 1.5% paraformaldehyde in 0.1 mol/L sodium cacodylate buffer, pH 7.3) for 24 h. The tissue pieces werethen washed with 3–4 changes of distilled water over a period of 2 h.The tissue was dehydrated using a standard ethanol series based ontissues fixed in formaldehyde fixatives (Ruzin, 1996). Tissue pieceswere embedded into JB-4 plastic (Polyscience Inc., Warrington, Penn-sylvania, USA) in molds per instructions of the manufacturer exceptthat, prior to plastic polymerization, the ratio of catalyzed JB-4 solutionA (Polyscience) to 100% ethanol was gradually increased from 1:4 to4:1 over 3 d. Tissue pieces were then placed in 100% catalyzed JB-4solution A for another 3 d, with fresh solution each day.
Plastic polymerized blocks were exposed to air for 1 d prior to sec-
tioning on an RMC MT-920 microtome (RMC, Tucson, Arizona, USA).Serial sections were 5–10 mm in thickness. Sections were stained by astandard periodic acid-schiff base series, and were counter-stained withtoluidine blue (0.05% in benzoate buffer, pH 4.4) (O’Brien and Mc-Cully, 1981).
Morphological observations of in vitro TP2 and TP1 shoots—Thir-ty in vitro TP2 and TP1 shoots from 4-wk-old cultures were measuredfor leaf area, internode length, and stem diameter. TP2 and TP1 shootswere chosen from ten randomly selected shoot cultures. In TP2 shoots,three of the shoots from each culture were randomly chosen for mea-surements. Because an individual TP1 shoot branches freely and growsas a cluster of shoots, one TP1 shoot was removed from each shootcluster for measurements. In both TP2 and TP1 shoots, the first fullyexpanded leaf below the shoot apex was chosen to measure the leafarea. Leaf area was estimated by multiplying the blade length from thebase to the apex with the maximum width of the blade. Below this leaf,the internode length and the stem diameter were measured.
RESULTS
Structure of TP2 shoots and axillary buds at differ-ent stages of development—In TP2 shoots, leaf primor-dia were initiated in an alternate phyllotaxy. Slight inter-nodal elongation had begun after the third leaf primor-dium (LP3) below the shoot apical meristem (SAM), butinternodal growth increased significantly after the fourthleaf below the SAM (Fig. 1).
The SAM had a tunica-corpus organization, with 2–3tunica cell layers distinguished by small, rectangular,densely stained cells (Fig. 2). These layers were confluentwith the adaxial surface of the youngest leaf primordium.The inversely hemispherical corpus consisted of irregu-larly shaped, more vacuolated meristematic cells (Fig. 2).Procambium differentiated acropetally from the olderregions of the stem into the youngest visible leaf pri-mordium (Fig. 2). The first differentiated tracheary ele-ment was observed in the internode below the fourthnode (Figs. 3, 4).
Early stages of axillary bud formation were character-ized by the presence of a mounded group of small, dense-ly cytoplasmic cells in the axil of the fourth node belowthe apex (Figs. 1, 3). The presence of meristematic cellsin the leaf axils in the first three nodes, while cell mat-uration continues in the petiole and stem proximal to theaxils, suggests that axillary buds are initiated as detachedmeristems. At the fifth node, the axillary bud was evidentas a dome displaced slightly toward the stem (Fig. 5).The apex of the dome consisted of small, densely stainedcells not yet characterized by a tunica-corpus organiza-tion, although an outer cell layer could be distinguishedby the presence of plastids in the cells. Below the apexof this bud, the mound of densely cytoplasmic cells wassubtended by small, thick-walled vacuolate cells thatformed an arc (Fig. 5). These cells formed the shell zonethat delimited the base of the axillary bud from the larger,vacuolate cells of the stem and petiole. In older axillarymeristems represented at nodes 12–14, additional leaf pri-mordia were not initiated; instead the prophylls increasedin size and overarched the meristem (Fig. 6). Cells in theshell zone also appeared more numerous and they hadthicker cell walls than in younger TP2 axillary buds (Fig.6). Procambium differentiation from the main shoot axisinto the axillary bud was not observed either in median
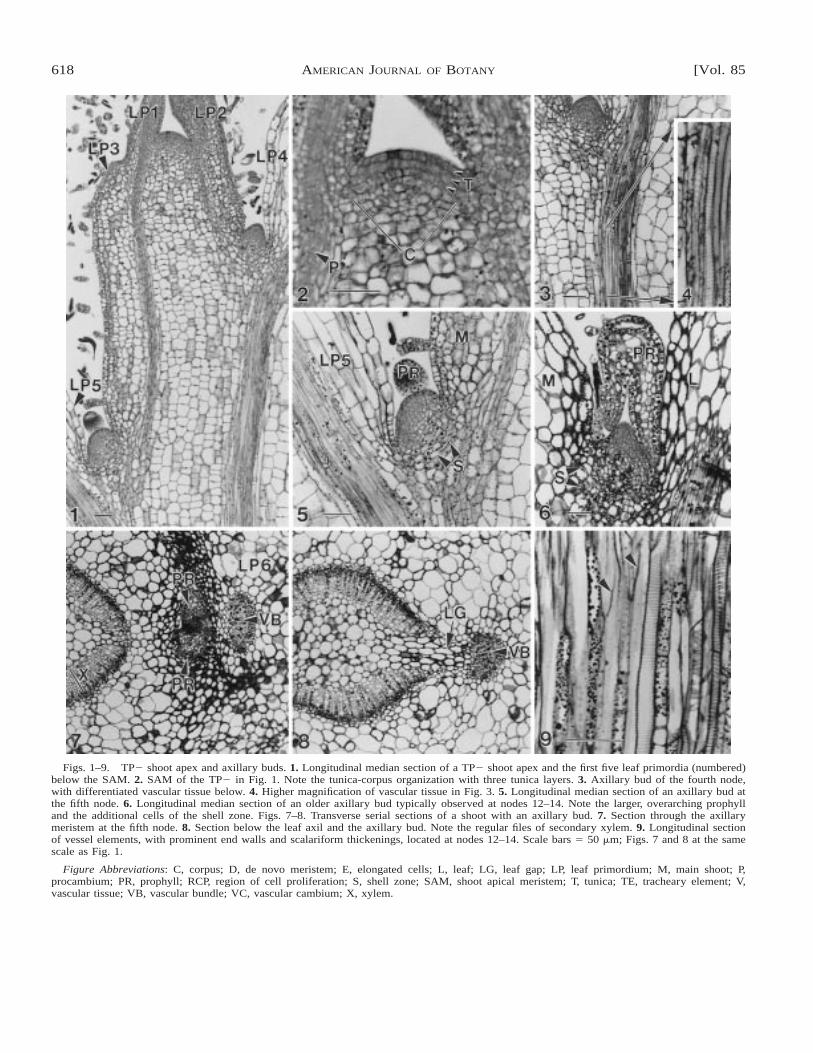
618 [Vol. 85AMERICAN JOURNAL OF BOTANY
Figs. 1–9. TP2 shoot apex and axillary buds. 1. Longitudinal median section of a TP2 shoot apex and the first five leaf primordia (numbered)below the SAM. 2. SAM of the TP2 in Fig. 1. Note the tunica-corpus organization with three tunica layers. 3. Axillary bud of the fourth node,with differentiated vascular tissue below. 4. Higher magnification of vascular tissue in Fig. 3. 5. Longitudinal median section of an axillary bud atthe fifth node. 6. Longitudinal median section of an older axillary bud typically observed at nodes 12–14. Note the larger, overarching prophylland the additional cells of the shell zone. Figs. 7–8. Transverse serial sections of a shoot with an axillary bud. 7. Section through the axillarymeristem at the fifth node. 8. Section below the leaf axil and the axillary bud. Note the regular files of secondary xylem. 9. Longitudinal sectionof vessel elements, with prominent end walls and scalariform thickenings, located at nodes 12–14. Scale bars 5 50 mm; Figs. 7 and 8 at the samescale as Fig. 1.
Figure Abbreviations: C, corpus; D, de novo meristem; E, elongated cells; L, leaf; LG, leaf gap; LP, leaf primordium; M, main shoot; P,procambium; PR, prophyll; RCP, region of cell proliferation; S, shell zone; SAM, shoot apical meristem; T, tunica; TE, tracheary element; V,vascular tissue; VB, vascular bundle; VC, vascular cambium; X, xylem.

May 1998] 619MERCURE ET AL.—IN VITRO TISSUE PROLIFERATION OF R. ‘MONTEGO’
sections or in sections proximal or distal to the medianin the two stages of axillary bud development examined.
Transverse serial sections at the sixth node revealedthat the vascular cambium was continuous and had beguninitiation of secondary vascular tissue, with 7–9 vesselsper file of secondary xylem (Figs. 7, 8). The small leafgap facing the axillary bud and petiole reflects the di-vergence of the leaf primordium subtending the axillarybud (Fig. 7). The prophylls and the meristem of the ax-illary bud are distinguished from the cells of the sur-rounding leaf and stem by the appearance of small,densely stained cells (Fig. 8). The axillary bud is seatedat the junction of the leaf and stem, and the initial direc-tion of growth is parallel to the petiole axis (Fig. 7). Theleaf gap initially widens below the level of insertion ofthe axillary bud in serial sections toward the base of theshoot, and vacuolate cells similar to those of the pith, butmore elliptical in shape, are observed within the gap (Fig.8).
Few differences in structure of shoots were observedin younger nodes vs. older nodes. Tracheary elements inthe stem at nodes 12–14 were more mature with moreprominent end walls and helical thickenings in the lateralwalls of the vessel elements (Fig. 9). In addition, theolder stems had thicker epidermal and parenchyma cellwalls, but the amount of secondary tissue did not increasesignificantly.
Structure of TP1 shoot apices—Although TP2 shootsand TP1 shoots have alternate phyllotaxy, patterns ofinternodal elongation differ in TP1 shoots. Less inter-nodal elongation had occurred by the fourth leaf primor-dium (LP4)(Fig. 10). Internodes between LP4 and LP6were variable in length and in some shoots these inter-nodes were shorter than the same internodes in TP2shoots (compare Fig. 10 with Fig. 1). The average lengthof these internodes, however, was not significantly dif-ferent between TP2 and TP1 shoots (internode betweenLP5 and LP6 was 1.6 mm for both shoots).
The SAM in TP1 shoots appeared less organized thanthe TP2 SAM. Cells at the top of the meristem weredensely stained but had an irregular arrangement, leadingto an inability to distinguish discrete tunica layers (Fig.11). The area that the tunica occupied was smaller thanthe tunica of TP2 shoots (compare Fig. 11 with Fig. 2).The hemispheric corpus region was also smaller, withfewer meristematic cells present (Fig. 11). Differentiatedvacuolate cells occurred closer to the apex than in TP2shoots as disorganized files contiguous with those in old-er regions of the stem, and acropetally developing pro-cambium was first observed in the second primordium(Fig. 11). The maturation of vascular tissue began at thesame position as in TP2 shoots, in the internode betweenthe fourth and fifth leaf (Fig. 12). TP1 shoots had moreirregular-sized epidermal cells and longer pith and cortexcells than in TP2 shoots at the same nodal positions(compare Figs. 10 and 1).
In TP1 shoots, the formation of axillary buds appearedto originate at a slightly lower position than in TP2shoots. However, the distance of the axillary bud fromthe SAM was the same as in TP2 shoots due to theshorter internodes at the apex of TP1 shoots. In TP2shoots, axillary buds had initiated development by the
fourth node to form a mounded group of cells in the leafaxil, but axillary bud initiation in TP1 shoots was indi-cated by the presence of a few, small vacuolate cells thathad proliferated in the leaf primordium axil of the fifthnode (Fig. 13). At the next node below, the small vacu-olate cells had proliferated toward the cortex and theysubtended a small mounded group of densely cytoplasmiccells without a tunica-corpus organization (Fig. 14) as inTP2 axillary buds. The buds were more displaced towardthe stem axis than TP2 axillary buds; they extended fromthe stem above the axil, facing the petiole (Fig. 14). Ad-ditional differences from TP2 axillary buds were ob-served in transverse serial sections of TP1 axillary budsat the sixth node (Figs. 15, 16). The presence of prophyllswith the axillary bud at the sixth node was not consistent,unlike in TP2 axillary buds. When prophylls were pres-ent, usually one was observed, and it had the same small,densely cytoplasmic cells as the axillary dome (Fig. 15).Transverse sections confirmed that the axillary bud wasdisplaced above the leaf axil on the shoot, facing thepetiole (Fig. 15). Although axillary buds were nearly par-allel to the stem in TP2 shoots, axillary buds were per-pendicular to the stem in TP1 shoots (Fig. 15). Trans-verse sections also confirmed the presence of irregularlyshaped, meristematic cells that subtended the axillary budand had extended into the cortex (Fig. 15). In TP1shoots, sections through the axillary bud revealed that theleaf had already diverged from the stem, but in sectionsthrough the axillary buds of TP2 shoots, the leaf wasstill attached to the stem (compare Figs. 7 and 15). Thewider leaf gap facing the axillary bud and petiole in TP1shoots suggested that the divergence of the leaf subtend-ing the axillary bud was farther below the bud than inTP2 shoots (Fig 16). The leaf gap was also wider thanin TP2 shoots when leaves had begun to diverge fromthe stem (compare Figs. 8 and 16), but leaves were sig-nificantly smaller (2.3 mm2) than TP2 leaves (53.4 mm2).
Although the vascular cambium in TP1 shoots wascontinuous, vascular tissue could not be clearly distin-guished from the cambium due to the presence of fewerand more irregular files of cells (Figs. 15, 16). Less sec-ondary growth corresponded to the smaller diameter ofTP1 shoots (0.4 vs. 0.8 mm of TP2 shoots). In addition,the vasculature of the main shoot, leaf, and the axillarybud was radially aligned in TP2 shoots, but not in TP1shoots because of divergence of the leaf to one side ofthe bud (compare Figs. 7 and 15). This displacement waspossibly due to the fact that the petioles were smaller thanthe petioles of TP2 shoots.
Tumor initiation—The initial stages of tumor forma-tion were characterized at node 7 by a mass of dividingcells, here designated as a region of cell proliferation(RCP), that extended from the cortex (Fig. 17) acrope-tally into the axillary bud (Fig. 18). Procambium haddeveloped toward the apex of the axillary bud (Fig. 18),but it could not be clearly distinguished near the RCP.Serial sections revealed the presence of cells that weremore elongated in length within the RCP (Fig. 17), butwhether these cells were procambial connections from theaxillary bud to the main TP1 shoot could not be deter-mined. Due to cell division in the RCP, the resulting ax-illary bud was enlarged at the base to form a bulbous

620 [Vol. 85AMERICAN JOURNAL OF BOTANY
Figs. 10–16. TP1 shoot apex and axillary buds. 10. Longitudinal median section of a TP1 shoot apex and the first six nodes (numbered)below the SAM. 11. SAM of the TP1 shoot in Fig. 10. Note the altered tunica-corpus organization as compared to TP2 SAM. 12. Trachearyelements first observed in the internode below the fourth node. 13. Axillary bud initiation at the fifth node of the shoot in Fig. 10. 14. Axillarybud at the sixth node of the shoot in Fig. 1. Note the displacement of the bud above the axil and the additional proliferation of cells at the baseof the bud. Figs. 15–16. Transverse serial sections of a shoot at node 6. 15. Section through the axillary meristem. Note that the bud is parallel,not transverse, to the plane of the section. 16. Section below the leaf axil and the axillary meristem. Scale bars 5 25 mm in Figs. 12–14 and 50mm in Figs. 10–11, 15–16.
structure above the leaf axil (Fig. 18). This bulbous struc-ture marked the initial development of the tumor, and theabnormal axillary bud is now designated a tumor bud.The meristem of the tumor bud was similar in structureto the TP1 SAM, in which no discrete tunica layers couldbe distinguished and files of large vacuolated cells werepresent close to the meristem (Fig. 18). Shoot develop-
ment was indicated by the presence of regularly initiatedleaves at the tumor bud apex (Fig. 18).
Tumor development pattern A: polar tumors—Aftertumor initiation, one of two different types of tumors wasobserved to develop: polar shoot tumors or nonpolar tu-mors. In polar shoot tumors, tumor growth was domi-

May 1998] 621MERCURE ET AL.—IN VITRO TISSUE PROLIFERATION OF R. ‘MONTEGO’
Figs. 17–18. Serial longitudinal median sections through a TP1 shoot with a tumor initiating development at node 7. Scale bars 5 50 mm.
nated by a single shoot (tumor shoot) that developed fromthe original tumor bud, as leaves and leaf primordia wereinitiated in a regular sequence. Two types of polar tumorscan develop after tumor initiation.
In polar tumor type 1 (PT1), the RCP, after initiationat the base of the tumor, extended basipetally toward thejunction of the main shoot and subtending leaf vascula-ture of nodes 8–9 (Figs. 19–20). The RCP contributed tothe formation of the widened base of the tumor. Elon-gated cells in the RCP, and in linear files that extendedfrom the vascular junction (Fig. 19), were present at thetumor base, but the cells could not be readily identifiedas procambium. Serial sections revealed that the elon-gated cells became twisted around cells of the RCP, withno differentiation of vascular tissue. However, procam-bium development into the most recently initiated leafprimordia of the tumor bud was clearly observed (Fig.20). These observations suggest that tumor shoots wereconnected to the main TP1 shoot by the RCP, not bystrong vascular connections. Because of the displacementof the abnormal axillary bud onto the main TP1 stem,further growth of the tumor shoot caused the subtendingleaf to become more perpendicular to the TP1 shoot (Fig.19). Epidermal, cortex, and pith tissues could be readilydistinguished in all stages of tumor shoot development(Fig. 20). The apical meristem of the tumor became moreorganized and was similar in structure to the TP2 SAM,with 2–3 tunica layers and an inversely hemisphericalcorpus (Fig. 20).
Further development of PT1 tumors was examined atnodes 10–11. These tumors exhibited continued growthof the single tumor shoot without additional proliferationof the RCP beyond the base of the tumor shoot. Thetumor shoots had some similarities to TP2 shoots, in thatthe cells of the cortex and pith were not elongated in
length and normal internodal elongation had begun (Fig.21). In addition, the tumor apical meristem remained sim-ilar in structure to that of TP2 shoots and younger PT1tumors (Fig. 22). However, as in TP1 shoots, more dif-ferentiated, vacuolate cells were present close to the SAM(Fig. 22) and the axillary buds on the tumor shoot weredisplaced onto the main tumor axis (Fig. 23). A newlydeveloped RCP subtended the axillary buds and had be-gun to proliferate into the cortex of the tumor shoot (Fig.23). Therefore, the PT1 tumor shoots had an intermediatestructure with characteristics similar to both TP2 andTP1 shoots.
In type 2 (PT2) polar tumors, as in PT1 tumors, theRCP extended basipetally toward the junction of the mainshoot and subtending leaf vasculature, and no procam-bium or mature vascular strands could be readily distin-guished within the RCP. However, the PT2 tumors thatwere examined at nodes 8–9 had a larger base than PT1tumors due to the acropetal extension of the RCP intothe tumor, a larger RCP, and the presence of larger, moreirregularly shaped epidermal and cortical cells (Fig. 24).The area of the RCP and the area of the cortical andepidermal regions of the tumor that extended beyond thecircumference of the main shoot were proportional in size(Fig. 24).
The interior of the RCP consisted of round, pith-likecells surrounded by a band of smaller, proliferating cells(Fig. 24) with individually differentiated tracheary ele-ments that were perpendicular and parallel to the tumoraxis in the same section (Fig. 25). These proliferatingcells were continuous with the weak vascular cambiumof the main shoot axis (Fig. 24). Therefore, the greaterincrease in width at the base of the tumor was due tobidirectional production of parenchyma cells (with iso-lated tracheary elements toward the interior) by the RCP.

622 [Vol. 85AMERICAN JOURNAL OF BOTANY

May 1998] 623MERCURE ET AL.—IN VITRO TISSUE PROLIFERATION OF R. ‘MONTEGO’
←
Figs. 19–27. Polar shoot tumor development from nodes 8 to 12. Figs. 19–20. Serial longitudinal sections of a young PT1 tumor at nodes 8–9. 19. PT1 tumor showing the extension of the RCP down at the vasculature junction of the main TP1 shoot and of the leaf that subtends thetumor. 20. Median longitudinal section of the tumor. 21. PT1 tumor at the nodes 10–11. 22. The SAM of the PT1 tumor shown in 21. 23. Axillarybud of the PT1 tumor shoot of 21. Note the displacement onto the stem. 24. Median section transverse to the main TP1 shoot of a PT2 tumor atnodes 10–11, with a larger RCP and a more abnormal tumor shoot. The main TP1 shoot to which the tumor is attached is bracketed. 25. Isolated,distorted tracheary elements within the RCP of the PT2 tumor of 24. 26. Apex of the PT2 tumor shoot of 24. 27. Axillary bud of the PT2 tumorshoot of 24. Scale bars 5 25 mm in Fig. 22, 50 mm in Figs. 19–20, 23, and 25–27, and 100 mm in Figs. 21 and 24.
The bidirectional growth indicated that the RCP had acambium-like function. Because no vascular strands werepresent within the RCP at the base of the tumor, thesePT2 tumors were also connected to the main TP1 shootby the RCP.
Displacement of the axillary bud caused tumor devel-opment to be perpendicular to the main TP1 shoot (Fig.24). PT2 tumor shoots developed an epidermis and cortexwith more uniform cell sizes than the tumor base, butthese shoots had less internodal elongation than in TP1shoots (Fig. 24). In addition, the structure of the tumorbud remained unorganized after tumor initiation, with thestructure of the SAM similar to that of the TP1 shoot(Fig. 26). Axillary buds of the tumor shoots were char-acterized by a mounded group of small densely cyto-plasmic cells located in the leaf axils in a similar positionto that of TP2 shoots (Fig. 27). However, no shell zonesubtended the axillary bud (Fig. 27). Therefore, the PT2tumor shoots also had an intermediate structure withcharacteristics of both TP2 and TP1 shoots, but theywere more similar to TP1 shoots.
Tumor development pattern B: nonpolar tumors—Inthe second tumor type that developed, tumor initiationwas similar to that of polar shoot tumors (Figs. 17, 18)with the base of the tumor bud swollen due to prolifer-ation of the RCP. However, subsequent development dif-fered from polar shoot tumors in that tumor growth wasnot dominated by the formation of a single shoot meri-stem. Instead, tumor development was characterized atnodes 8–9 by the formation of apparent de novo meri-stems in addition to the original tumor bud (Figs. 28, 29).Because tumor development resulted from growth inmore than one direction, these tumors are called nonpolartumors. The meristems of these de novo lateral buds con-sisted of only very small, densely cytoplasmic cells thatwere not subtended by shell zones (Fig. 28). Files ofelongated cells, presumably procambium, were observedto extend from the vasculature of the main shoot andfrom the leaf subtending the tumor toward the lateralbuds (Fig. 28). Instead of a shell zone, clusters of small,densely cytoplasmic cells subtended de novo meristems;larger, more vacuolated cells of the RCP were presentthroughout the tumor (Figs. 28, 29). Unlike polar shoottumors, extensive proliferation of the RCP toward thejunction of the main TP1 shoot and subtending leaf vas-culature did not occur (Figs. 28, 29).
Growth of nonpolar tumors as represented by tumordevelopment at nodes 10–11 was due to proliferation ofthe RCP. The RCP had extended more into the cortex ofthe main TP1 shoot by this stage of development (Fig.30). Isolated regions of vascular tissue had differentiatedwithin the RCP at the base of the tumor (Fig. 31). As in
PT2 tumors, individually distorted tracheary elementswere also observed to be in parallel and perpendicular tothe plane of section. Tumors were composed of a centralcore of RCP tissue, with epidermal cells and cells outsidethe RCP that were irregular in size and shape and cellsinside the RCP that were all similar in size, smaller, andthicker walled (Figs. 32, 33). In median sections, the RCPappeared as globular extensions of proliferating cells ad-jacent to each other that were partly surrounded by long,narrow parenchyma cells (Figs. 32, 33). Farther withinthe large region of parenchyma tissue of the tumor, iso-lated vessel elements cells continued to differentiate fromcells of the RCP (Fig. 34).
Lateral tumor shoots did develop from nonpolar tu-mors (Figs. 33, 38). Vascular tissue had differentiated atthe base of the shoot, but vascular strands of the devel-oping tumor shoot did not extend through the RCP (Fig.33) to form a direct vascular connection from the tumorshoot to the main TP1 shoot. Most tumor shoots on non-polar tumors shared many features that characterizedTP1 shoots, including many leaves initiated close to theSAM, a SAM that lacked distinguishable tunica layers,and axillary meristems that were displaced onto the stemof the tumor shoot (Fig. 35). In addition, the tumor shootswere short and wide at the base as in PT2 tumor shoots(Fig. 35). In other nonpolar tumor shoots, the SAM wasmore organized with distinguishable tunica layers (Fig.36).
The formation of lateral tumor shoots on the originaltumor was associated with the continued formation of denovo buds on the external surface of the tumor. As inyounger nonpolar tumors, the meristems consisted ofsmall, densely cytoplasmic cells. These meristems ap-peared to be external proliferations of the RCP that werepresent close to the tumor surface below the epidermis(Fig. 37). Tumor shoot formation from the buds was in-dicated by the regular initiation of leaves and procam-bium development in association with leaf primordia,with procambium extending basipetally into the RCP(Fig. 37).
Additional lobed tumor growths on the surface of theoriginal tumor also involved de novo bud formation andextension of the RCP. These tumor growths, unlike thetumor shoots, consisted of RCP surrounded by parenchy-ma tissue and they did not develop leaves with axillarybuds (Fig. 37). The RCP of these tumors was locatedproximally to the tumor bud (Fig. 38). The simultaneousdevelopment of tumor and tumor shoots with nondirec-tional growth from buds that did not form in leaf axils(Figs. 37, 38) suggested that the subsequent developmentof nonpolar tumors occurred from adventitious meri-stems.
De novo bud formation on the surface of tumors in

624 [Vol. 85AMERICAN JOURNAL OF BOTANY

May 1998] 625MERCURE ET AL.—IN VITRO TISSUE PROLIFERATION OF R. ‘MONTEGO’
←
Figs. 28–38. Nonpolar tumor development from nodes 8 to 11. 28–29. Serial longitudinal sections of a tumor at nodes 8–9 in which a de novomeristem has formed with the tumor bud. Figs. 30, 32–33, 37–38. Longitudinal serial sections through a tumor at nodes 9–11. 30. Tumor developmentfrom the main TP1 shoot. The RCP has extended into the cortex of the shoot. 31. Isolated, distorted tracheary elements within the RCP of thetumor in Fig. 30. 32. Median section of the tumor showing the large, whorled RCP. 33. Tumor with one of many tumor shoots. 34. Continueddifferentiation of isolated tracheary elements (arrowheads) within the RCP of the tumor in Fig. 33. 35. Longitudinal section of the tumor shootapex of Fig. 33. 36. Longitudinal section of a more organized SAM of another tumor shoot. 37. Tumor with de novo meristem formation, some ofwhich have initiated shoot development. 38. Tumor shoot with the development of tumors on each side of the shoot. Scale bars 5 25 mm in Figs.34 and 36, 50 mm in Figs. 28, 31, and 35, and 200 mm in Figs. 30, 32–33, and 37–38. Fig. 29 is in the same scale as in Fig. 28.
association with a massive, proliferating core of RCP, andthe formation of tumor and tumor shoots from these buds,continued in the development of the largest nonpolar tu-mors as observed at nodes 12–14 (Figs. 39, 40). In sometumors, lateral growth was continuous and became direc-tional, resulting in the development of elongated tumorsthat could be compared to a ‘‘swollen stem’’ (Fig. 39).Epidermal cells and cortical parenchyma cells outside theRCP continued to be irregular in cell size and shape,while cells inside the RCP remained small, round, andthick-walled (Figs. 39, 40). Some of the lateral tumorshoots were extremely compressed with little internodalelongation (Fig. 40).
The oldest nonpolar tumors had patterns of develop-ment that were different from the younger nonpolar tu-mors. Most of the cells of the RCP differentiated into avast region of parenchyma tissue interspersed with somevessel elements. However, more vascular tissue differ-entiation had occurred in the oldest tumors, but even soregions of vascular tissue appeared isolated from eachother (Fig. 39). The presence of the RCP adjacent to thevascular tissue of the tumor (Figs. 39, 40) suggested thatthe RCP was involved in vascular tissue differentiation.Differentiation of vascular tissue was not always parallelto the axis of tumor growth (Fig. 39). Serial sectionsrevealed that the vascular tissue close to the base of thetumor was connected within the tumor, and that the vas-culature of the tumor was weakly connected to the TP1shoot vasculature (shown in longitudinal section in Fig.39; transverse section in Fig. 40). In tumors that formeda large globular mass of lateral tumors, de novo meri-stems not only formed on lateral tumors, but were alsoobserved in the axils that formed between lateral tumors(Fig. 40). Finally, de novo meristem formation did notoccur on the surface of some mature lateral tumors.
DISCUSSION
During growth and development of in vitro R. ‘Mon-tego’ cultures, differences in the morphology of shootswith and without TP are readily apparent. TP2 shootswere characterized by a single shoot with apical domi-nance, but TP1 shoots branched readily from nodal tu-mors causing shoots to be shorter and thinner with smallerleaves. The differences in shoot morphologies can be ex-plained by the observation of four significant differencesin the structure of TP1 shoots as compared to TP2 shoots:a smaller, less organized SAM, shorter internodes towardthe apex, a less active cambium, and axillary buds that aredisplaced from the axil onto the shoot. For example, thesmaller SAM and the less active cambium lead to the for-mation of smaller leaves on the more narrow TP1 shoots.
The timing of shell zone formation is the major dif-
ference in the anatomy of axillary buds between R. ‘Mon-tego’ and other woody plants. The shell zone initiatesaxillary bud development as early as the first node belowthe SAM in Fraxinus shoots (Remphrey, 1989), but usu-ally in the second to fourth node below the SAM in Bet-ula, Syringa, Euptelea (Garrison, 1949a, b) and Arcto-staphylos (Remphrey and Steeves, 1984). In TP2 shoots,the arcuate group of vacuolated cells that form the shellzone subtends the axillary bud at the fifth node belowthe SAM. However, the axillary bud one node above con-sists of a smaller dome of meristematic cells without adistinguishable shell zone present. Therefore, unlike inother woody plants, it is unclear whether the shell zoneinitiates axillary bud formation in TP2 shoots. TP1shoots are more remarkable in that a shell zone is notpresent at all. Instead, axillary buds are initiated aftercells proliferate in the cortex adjacent to the fifth leaf axilbelow the SAM. Both the shell zone in TP2 shoots andthe proliferated cells in TP1 shoots subtend axillarybuds. This indicates that the proliferated cells in the TP1leaf axils, which correspond to the same location as theshell zone in TP2 shoots, precede axillary bud devel-opment in TP1 shoots.
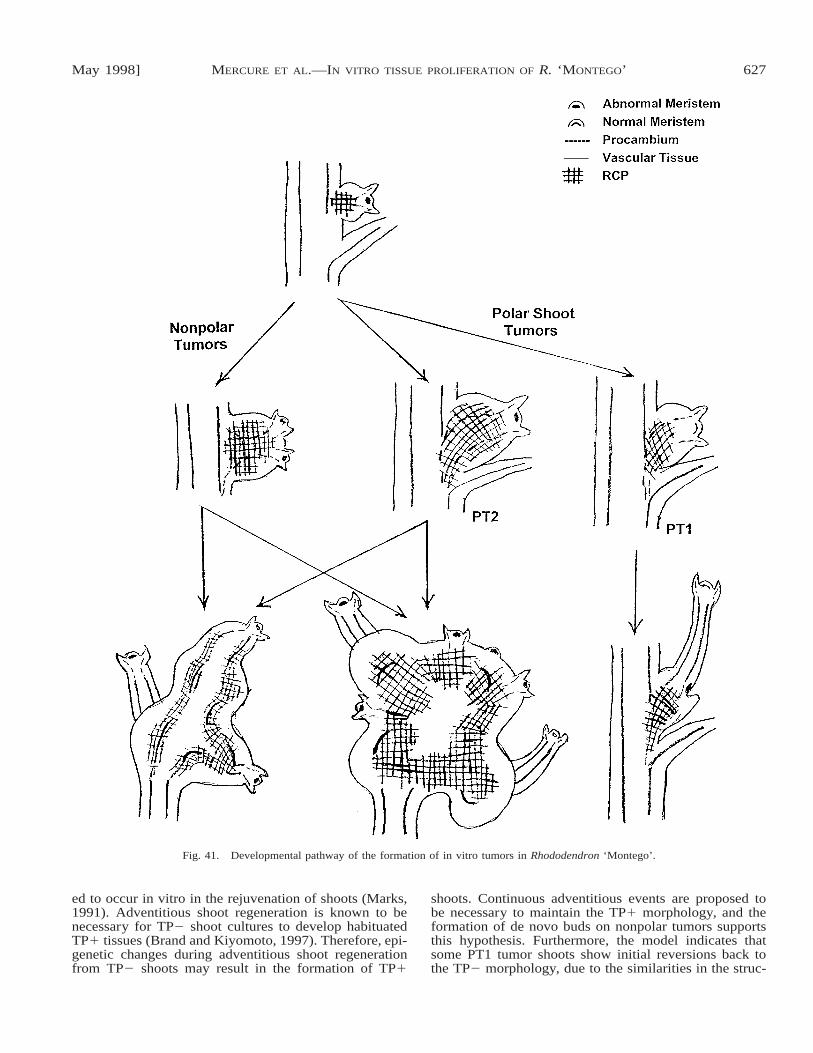
Cell proliferation in the axils of TP1 shoots leads tothe multiple branching pattern as axillary buds are initi-ated, but were not inhibited in growth. Tumors resultfrom such uninhibited growth when cell proliferationcontinues, forming an RCP, while the axillary buds ini-tiate shoot development. A model for tumor developmentin Fig. 41 shows that subsequent organization of the ax-illary meristem of the tumor, with or without de novomeristem formation, and extension of the RCP charac-terizes the formation of either polar shoot tumors or non-polar tumors. As shown in the model, some PT2 tumorsmay develop into nonpolar tumors, since the major dif-ference between the two tumor types is the presence ofa single shoot in PT2 tumors.
Comparisons of the structure of TP tumors to crowngalls and lignotubers could provide evidence that TP iscaused by crown gall disease or abnormal lignotuber for-mation. In the Ericaceae, crown gall of Vaccinium (Row-land, 1990) and lignotubers of Kalmia (Del Tridici, 1992)and Rhododendron (James, 1984) are known to occur.Although the anatomy of these structures in the Ericaceaehas not been studied, they have been described for otherwoody plants. Our comparisons are based on the structureof crown gall in apple (Riker and Keitt, 1926; Sylwesterand Countryman, 1933) and on the extensive morpholog-ical and anatomical descriptions of lignotubers in Euca-lyptus (Carr, Jahnke, and Carr, 1984), but they are limiteddue to differences in the growth of whole vs. tissue-cul-tured plants.

626 [Vol. 85AMERICAN JOURNAL OF BOTANY
Figs. 39–40. Largest nonpolar TP tumors at nodes 12–14. 39. Longitudinal section through the main TP1 shoot (bottom) to which an elongatedtumor is attached. 40. Transverse section through the main TP1 shoot (bracketed) to which a globular tumor is attached. Scale bar 5 200 mm.
In the crown gall of apple, the gall is composed mostlyof parenchyma tissue early in development that later isreplaced by xylem (Riker and Keitt, 1926; Slywester andCountryman, 1933). Although both the meristematic areaand the RCP initiate development of crown galls and TPtumors, respectively, several differences exist betweenthe meristematic area and the RCP. Unlike the meriste-matic area in crown gall, the RCP persists throughoutdevelopment and remains connected to additional RCPsthat develop. Although vessels that differentiate from in-itials of the meristematic area are distorted in width, theTP tumor RCP produces initials that differentiate intodistorted parenchyma and unevenly distributed, randomlyoriented vascular tissue. In addition, an axillary meristemis not involved in crown gall initiation. These differencesin anatomy suggest that TP is not crown gall.
Lignotubers in Eucalyptus are located at the crown,usually at the cotyledonary nodes, and consist of swollenwoody tissue located above and/or below the axillary bud(Carr, Jahnke, and Carr, 1984). If present, additional (ac-cessory) buds are located with the axillary bud. Ligno-tubers in some Eucalyptus genera are remarkably similarto small TP tumors, in which the lignotuber is part of aswollen axillary bud with accessory buds on either side(Carr, Jahnke, and Carr, 1984). The lignotuber cambium,like the TP tumor RCP, produces many more initials thanthe main stem cambium. However, unlike TP tumors, lig-notubers only grow from continued division of its cam-bium, eventually surrounding all the buds with parenchy-ma, and later, vascular tissue. Once the accessory budshave formed, no new ones are formed in lignotubers,while de novo meristems are continually produced on thesurface of existing TP tumors. Another interesting dif-ference between lignotubers and in vitro TP tumor de-velopment is that the lignotuber cambium produces muchmore vascular tissue than parenchyma, while the oppositeoccurs in the RCP of TP tumors. In addition, the swirling
of vascular tissue observed in lignotubers appears to oc-cur around the accessory buds during cambial growth,unlike the random orientation of vascular tissue in TPtumors.
A final comparison between crown galls, lignotubers,and TP tumors is that the structure of the main shoot towhich the crown gall or lignotuber is attached is not al-tered prior or during crown gall or lignotuber formation.However, the structure of the main TP1 shoot is alteredprior to tumor initiation and development. This providesfurther evidence that TP is not crown gall or lignotuberdevelopment.
Another tumor-like growth similar to TP is the tobaccogenetic tumor. These tumors with short shoots sponta-neously occur in the leaf axils in hybrid plants of Nico-tiana glauca and N. langsdorffii (Levine, 1937; Smith,1972). TP tumors and the genetic tumors are similar inanatomy and in in vitro behavior. Tobacco genetic tu-mors, like TP tumors, consist of proliferations of paren-chyma and vascular tissue with many adventitious budslocated on the surface of the tumors (Levine, 1937). Asin the oldest TP tumors, internal meristematic areas areassociated with the growth of new tumors from de novobuds (Levine, 1937).
One other similarity between tobacco genetic tumorsand TP1 tissues is that they appear to be habituated whencultured under in vitro conditions (Smith, 1972). Habit-uation, rejuvenation, and dwarfism are examples of mor-phological shifts that result from epigenetic changes dur-ing tissue culture (Meins, 1989; Moore, Robbins, andSjulin, 1991). Epigenetic change involves persistentchanges in gene expression that leads to changes in plantdevelopment (Meins, 1989; Skirvin, McPheeters, andNorton, 1994). Although habituation was first observedin callus cultures, shoot tip cultures have been known tobecome habituated during tissue culture (Gaspar et al.,1991). In rhododendron, epigenetic changes are suggest-
May 1998] 627MERCURE ET AL.—IN VITRO TISSUE PROLIFERATION OF R. ‘MONTEGO’
Fig. 41. Developmental pathway of the formation of in vitro tumors in Rhododendron ‘Montego’.
ed to occur in vitro in the rejuvenation of shoots (Marks,1991). Adventitious shoot regeneration is known to benecessary for TP2 shoot cultures to develop habituatedTP1 tissues (Brand and Kiyomoto, 1997). Therefore, epi-genetic changes during adventitious shoot regenerationfrom TP2 shoots may result in the formation of TP1
shoots. Continuous adventitious events are proposed tobe necessary to maintain the TP1 morphology, and theformation of de novo buds on nonpolar tumors supportsthis hypothesis. Furthermore, the model indicates thatsome PT1 tumor shoots show initial reversions back tothe TP2 morphology, due to the similarities in the struc-

628 [Vol. 85AMERICAN JOURNAL OF BOTANY
ture of TP2 shoots and PT1 tumor shoots. In R. ‘Mon-tego’, TP1 shoots are observed to occasionally revertback at a low frequency to a morphology resembling thatof TP2 shoots (Ruan, 1995). Epigenetic changes areknown to be unstable, as reversions back to the originalmorphology can occur once the inducing agent is re-moved (Moore, Robbins, and Sjulin, 1991; Skirvin,McPheeters, and Norton, 1994). In tobacco callus cul-tures that are habituated to cytokinin, plants can be re-generated that produce lines of habituated and cytokinin-requiring callus, indicating that reversions in habituationreadily occur withn a clonal population of cells found inthe callus (Meins and Hansen, 1986). However, in TP1shoots, we anticipate that reversions occur at a lower fre-quency in TP1 shoots since these reversions would in-volve changes in meristematic regions (the RCP and theSAM) within the differentiated TP1 shoot, which maynot occur simultaneously.
In other plants that have an increased level of the en-dogenous plant hormone cytokinin due to overexpressionof an introduced cytokinin biosynthesis gene, the plantsare similar to TP1 shoots in that they lack apical domi-nance, have smaller leaves, form new shoots from denovo meristems, and also appear habituated (Smigockiand Owens, 1988; Medford et al., 1989; Smigocki, 1991;Li, Hagen, and Guilfoyle, 1992). During the release ofaxillary buds from apical dominance, cytokinin, in thepresence of auxin, promotes xylem differentiation duringthe connection of vascular strands of buds to the mainshoot (Sorokin and Thimann, 1965). Tumor initiation canbe considered as abnormal buds being released from api-cal dominance, and elongated cells adjacent to the RCPat the base of the tumor may be distorted procambialconnections between the developing tumor and the mainshoot. These observations suggest that changes in endog-enous plant hormones may be involved in the TP phe-notype. Metabolism of exogenous cytokinins and endog-enous cytokinin levels is currently being comparedamong TP2 and TP1 shoots and TP tumors to determinewhether altered hormone biochemistry is involved in thein vitro TP of R. ‘Montego’.
LITERATURE CITED
ANONYMOUS. 1992. News watch, phenomenon continues to defy an-swers. American Nurseryman 176: 17–20.
BRAND, M. H. 1992. Tissue culture variations: problems. AmericanNurseryman 175: 60–65.
, AND R. KYOMOTO. 1994. Tissue proliferation apparently notlignotubers. Yankee Nursery Quarterly 3: 5–6.
, AND . 1997. In vitro behavior of Rhododendron ‘Mon-tego’ shoots with and without tissue proliferation and the inductionof habituation via adventitious events. HortScience 32: 989–994.
CARR, D. J., R. JAHNKE, AND S. G. M. CARR. 1984. Initiation, devel-opment, and anatomy of lignotubers in some species of Eucalyptus.Australian Journal of Botany 32: 415–437.
DEL TREDICI, P. 1992. Seedling versus tissue-cultured Kalmia latifolia:the case of the missing burl. Combined Proceedings of the Inter-national Plant Propagators’ Society 42: 476–482.
GARRISON, R. 1949a. Origin and development of axillary buds: Syringavulgaris L. American Journal of Botany 36: 205–213.
. 1949b. Origin and development of axillary buds: Betula pa-pyrifera Marsh. and Euptelea polyandra Sieb. and Zucc. AmericanJournal of Botany 36: 379–389.
GASPAR, T., D. HAGEGE, C. KEVERS, C. PENEL, M. CREVECOEUR, I. EN-GELMANN, H. GREPPIN, AND J. M. FOIDART. 1991. When plant ter-
atomas turn into cancers in the absence of pathogens. PhysiologicaPlantarum 83: 696–791.
JAMES, S. 1984. Lignotubers and burls—their structure, function, andecological significance in Mediterranean ecosystems. Botanical Re-view 50: 225–266.
LAMONDIA, J. L., T. M. RATHIER, V. L. SMITH, T. M. LIKENS, AND M. H.BRAND. 1992. Tissue proliferation/crown gall in rhododendron.Yankee Nursery Quarterly 2: 1–3.
, , AND . 1997. Tissue proliferation in Rhododen-dron: association with disease and effect on plants in the landscape.HortScience 32: 1001–1003.
LEVINE, M. 1937. Tumors of tobacco hybrids. American Journal of Bot-any 24: 250–256.
LI, Y., G. HAGEN, AND T. J. GUILFOYLE. 1992. Altered morphology intransgenic tobacco plants that overproduce cytokinins in specifictissues and organs. Developmental Biology 153: 386–395.
LINDERMAN, R. G. 1993. Tissue proliferation. American Nurseryman178: 56–67.
LLOYD, G. B., AND B. H. MCCOWN. 1980. Commercially-feasible mi-cropropagation of mountain laurel, Kalmia latifolia, by use of shoottip culture. Combined Proceedings of the International Plant Prop-agation Society 30: 421–426.
MARKS, T. R. 1991. Rhododendron cuttings. I. Improved rooting follow-ing ‘‘rejuvenation’’ in vitro. Journal of Horticultural Science 66:102–111.
MEDFORD, J. I., R. HORGAN, Z. EL-SAWI, AND H. J. KLEE. 1989. Alter-ations of endogenous cytokinins in transgenic plants using a chi-meric isopentenyl transferase gene. Plant Cell 1: 403–413.
MEINS, F. Jr. 1989. Habituation: heritiable variation in the requirementof cultured plant cells for hormones. Annual Review of Genetics23: 395–408.
, AND C. E. HANSEN. 1986. Epigenetic and genetic factors reg-ulating the cytokinin requirement of cultured cells. In M. Bopp[ed.], Plant growth substances 1985, 333–340. Springer-Verlag,Berlin.
MOORE, P. P., J. A. ROBBINS, AND T. M. SJULIN. 1991. Field performanceof ‘Olympus’ strawberry subclones. HortScience 26: 192–194.
O’BRIEN, T. P., AND M. E. MCCULLY. 1981. The study of plant structure:principles and selected methods. Termarcarphi Pty. Ltd., Mel-bourne.
REMPHREY, W. R. 1989. Shoot ontogeny in Fraxinus pennsylvanica(green ash). I. Seasonal cycle of terminal meristem activity. Ca-nadian Journal of Botany 67: 1624–1632.
, AND T. A. STEEVES. 1984. Shoot ontogeny in Arctostapylos uva-ursi (bearberry): origin and early development of lateral vegetativeand floral buds. Canadian Journal of Botany 62: 1933–1939.
RIKER, A. J., AND G. W. KEITT. 1926. Studies of crown gall and woundovergrowth on apple nursery stock. Phytopathology 16: 765–807.
ROWLAND, L. J. 1990. Susceptibility of blueberry to infection by Agro-bacterium tumefaciens. HortScience 25: 1659.
RUAN, Y. 1995. Callus induction and shoot organogenesis in Rhododen-dron, and in vitro responses of tissues from rhododendron plantswith or without tissue proliferation. Master’s thesis, University ofConnecticut, Storrs, CT.
RUZIN, S. E. 1996. Plant microtechnique. University of California,Berkeley, CA.
SKIRVIN, R. M., K. D. MCPHEETERS, AND M. NORTON. 1994. Sourcesand frequency of somaclonal variation. HortScience 29: 1232–1237.
SMIGOCKI, A. C. 1991. Cytokinin content and tissue distribution inplants transformed by a reconstructed isopentenyl transferase gene.Plant Molecular Biology 16: 105–115.
, AND L. D. OWENS. 1988. Cytokinin gene fused with a strongpromoter enhances shoot organogenesis and zeatin levels in trans-formed plant cells. Proceedings of the National Academy of Sci-ences USA 85: 5131–5135.
SMITH, H. H. 1972. Plant genetic tumors. In A. C. Braun and F. Hom-burger [eds.], Progress in experimental tumor research-plant tumorresearch, vol. 15, 138–164. S. Karger, Basel.
SOROKIN, H. P., AND K. W. THIMANN. 1965. The histological basis forinhibition of axillary buds in Pisum sativum and the effects ofauxins and kinetin on xylem development. Protoplasma 59: 326–350.
SYLWESTER, E. P., AND M. C. COUNTRYMAN. 1933. A comparative his-tological study of crown gall and wound callus on apple. AmericanJournal of Botany 20: 328–340.