anatomy of disguise: camouflaging structures in nymphs of some
TRANSCRIPT

PUBLISHED BY THE AMERICAN MUSEUM OF NATURAL HISTORY
CENTRAL PARK WEST A T 79TH STREET, NEW YOR K, NY 10024
Number 3542, 18 pp., 9 figures, 3 tables December 7, 2006
Anatomy of Disguise: Camouflaging Structures inNymphs of Some Reduviidae (Heteroptera)
CHRISTIANE WEIRAUCH1
ABSTRACT
Immature stages of Reduvius personatus (Linnaeus) and some other Reduviidae are known tocamouflage themselves with a range of materials found in their environment. Even though thisbehavior has been observed in several species, camouflaging structures have never been studied ina comparative way. This study documents for the first time the structure that is involved in theapplication of camouflaging material, i.e., the hind tarsal fan, and reveals structures that assure thefastening of the camouflaging material, i.e., anchor setae and trichomes, in eight speciesrepresenting five subfamilies of Reduviidae. Whereas anchor setae assure the attachment ofcamouflaging material by their mechanical properties, short-projection trichomes, long-projectiontrichomes, and grouped trichomes are here proposed to secrete a sticky substance for this purpose.Primary homology hypotheses on the three types of trichomes are proposed. At least in somespecies, short-projection trichomes appear to be responsible for the fastening of the camouflaginglayer close to the integument, whereas long-projection trichomes may hold the outer layer ofcamouflaging material in place.
INTRODUCTION
Immature stages of the predatory assassinbug Reduvius personatus (Linnaeus, 1758), the‘‘masked hunter’’, have attracted the attentionof entomologists since their first descriptionalmost 250 years ago. The most remarkablefeature of these nymphs is a behavior that isknown in only few insects and is bestdocumented in the neuropteran Chrysopidae
family (Eisner et al., 1978; Stitz 1931): thenymphs camouflage themselves with a rangeof materials found in their environment.This behavior is accurately reflected in thename of Reduvius personatus: Fabricius’(1775) generic name Reduvius is derived fromthe Latin noun reduvia meaning ‘‘remainder orfragment’’, and Linnaeus’s (1758) epithet,the Latin adjective personatus, meaning‘‘masked’’.
Copyright E American Museum of Natural History 2006 ISSN 0003-0082
1 Division of Invertebrate Zoology, American Museum of Natural History ([email protected]).

The camouflage cover of R. personatusoften consists of fine soil or wood dust, whichtransforms the insect into a small walking pileof debris. The benefit for the nymph of R.personatus may be twofold, since the camou-flaged nymph might be less perceptible topotential predators, but the little predatormight also have an advantage in stalking itsown prey (Harz, 1952).
Camouflaging in nymphs of R. personatus—a behavior absent in the adult— was initiallythought to be a passive process, i.e., thenymphs were getting dirty merely because ofthe habitat they live in, as indicated by Fabre(1903) in his Souvenirs Entomologiques.However, Weber (1930) and Immel (1955)observed that nymphs throw substrate parti-cles on the dorsum using their hind legs andthat the cover is replaced after each moult.Dispons (1955), who referred to the camou-flaging behavior as ‘‘prosoponie’’, after theGreek word for ‘‘mask’’, gave a detailedaccount of the behavior for Reduvius persona-tus.
Structures that assist in throwing substrateparticles on the dorsum other than the hind-legs themselves—in the following referred toas ‘‘application structures’’—have never beendiscussed for Reduvius personatus. The struc-tures that fasten substrate particles on thenymphs’ integument, here called fasteningstructures, were likewise not studied in detail:Weber (1930) claimed that ‘‘erect setation’’ onthe integument accounted for the fastening.Madel (1951) and Immel (1955) argued thatsubstrate particles were held in place by shortthornlike and apically curved microtrichia,which are abundant on the dorsum of nymphsof R. personatus. Fabre (1903) and Miller(1956) believed that glandular secretions oreven glandular hairs were responsible for thefastening of camouflaging material, but theydid not document such secretions or hairs.
Nymphal camouflaging behavior is not re-stricted to R. personatus among Reduviinae,but is known for species of Acanthaspis Amyotand Serville (Brandt and Mahsberg, 2002;Ambrose, 1999; Louis, 1974; Miller, 1953;Odhiambo, 1958; Villiers, 1948; Butler, 1923),Alloeocranum Reuter (Ambrose, 1999), EdoclaStal (Ambrose, 1999), Empyrocoris Miller(Ambrose, 1999), Holotrichius Burmeister
(Kott, 2001; Dispons, 1955), Leogorrus (Lentand Wygodzinsky, 1979), Paredocla Jeannel(Brandt and Mahsberg, 2002; Villiers, 1948),Tetroxia Amyot and Serville (Louis, 1974), andspecies of Reduvius other than R. personatus(Dispons, 1955; Miller, 1953; Wygodzinskyand Usinger, 1946). In addition, camouflagingis not restricted to nymphs of Reduviinae,but also occurs in nymphs of Cetherinae(Louis, 1974), Salyavatinae (Ambrose, 1999;McMahan, 1982, 1983a, 1983b; Miller, 1953),Stenopodainae (Readio, 1927; Breddin, 1904),and Triatominae (Ambrose, 1999; Zeledon etal., 1973; Hase, 1940; Breddin, 1904). Cai et al.(2002) provided a literature summary oncamouflaging in several species of Reduviidae.Many of these descriptions state that camou-flaging is an active process during which thehind legs gather and apply camouflagingmaterial (e.g., Kott, 2001; McMahan, 1982;Louis, 1974; Dispons, 1955; Hase, 1940). As forR. personatus, all of the above studies lackdetailed examination of the hind legs that mayreveal application structures.
As to the camouflaging cover, it becomesclear from the above-mentioned referencesthat two types or layers of camouflagingmaterial are often distinguishable. The firstlayer is close to the integument, consists ofsmall soil and substrate particles, and isreferred to as ‘‘natural camouflaging’’ (Am-brose, 1999) or ‘‘dust coat’’ (Brand andMahsberg, 2002) by recent authors. Judgingfrom the literature, this dust-coat camouflag-ing seems to be present in all reduviid nymphsthat exhibit camouflaging. The second layer isfarther removed from the body and mayconsist of coarser particles, but also corpsesof insects the reduviid nymph has fed on.This outer layer is called ‘‘corpse camouflag-ing’’ by Ambrose (1999) or ‘‘backpack’’ byBrand and Mahsberg (2002). Corpse camou-flaging is known for several species ofAcanthaspis (Ambrose, 1999; Louis, 1974;Odhiambo, 1958), and for one species eachof Edocla (Ambrose 1999) and Paredocla(Brand and Mahsberg, 2002). Brandt andMahsberg (2002) demonstrated for a speciesof Paredocla that the corpse cover is impor-tant as protection from potential predators,whereas the dust cover may play a role inaggressive mimicry towards their ant prey.
2 AMERICAN MUSEUM NOVITATES NO. 3542

Even though some of the authors whodescribed camouflaging in reduviid nymphsexplained how camouflaging material wasfastened to the integument, illustrations offastening structures are limited to one linedrawing for Acanthaspis vitticollis by Louis(1974). No study so far has approached thefastening structures from a comparative per-spective, nor has any consideration been givento the fact that many species seem to applytwo layers of camouflaging material.
The present study aims to document appli-cation and fastening structures involved incamouflaging behavior in a sample of nymphsof Reduviidae using light and scanning elec-tron microscopy. Through its comparativeapproach, this study will assist in establishinga terminological framework for the descrip-tion of camouflaging structures and will helpto gain insights into their potential homolo-gies.
MATERIAL AND METHODS
Nymphs of eleven species of Reduviidaerepresenting eight subfamilies were examinedin this study using light microscopy andscanning electron microscopy. For six ofthese species in the subfamilies Reduviinae,Salyavatinae, and Triatominae, camouflagingbehavior was previously reported. For theremaining five species in the subfamiliesCetherinae, Hammacerinae, Harpactorinae,Peiratinae, Reduviinae, and Sphaeridopinae,evidence of camouflaging behavior or behav-ior of nymphs in general is absent in theliterature. Examination of these species mayreveal camouflaging structures, and may thuspredict if camouflaging behavior could occur.
Two aspects determined the choice of taxastudied. First, immatures are collected ratherinfrequently and—unless they are associatedwith adults—it is often impossible to identifythem even to generic level. The availability ofmaterial is therefore limited. Secondly, thespecies or genera were chosen to represent thewidest possible range of subfamilies.
LIGHT MICROSCOPY (LM): To examinedifferent layers of camouflaging material onthe integument, a living 4th instar nymph ofReduvius personatus was partly dismantledwith fine forceps and observed using a Wild
Heerbrugg 307800 stereomicroscope. Formore detailed light microscopic observation,the abdominal tergites and part of thepronotum were removed and macerated inwarm KOH (approximately 10%) for approx-imately four hours. After washing with 70%alcohol and staining with Chlorazol Black E,pieces of integument were set up for observa-tion on a slide in glycerin and examined usingeither a Leitz Diaplan, Zeiss Axioplan (FreieUniversitat Berlin) or a Nikon Eclipse 80i(American Museum of Natural History) com-pound microscope. Photographs were takenwith a Fujifilm FinePix S1 Pro digital cameraon the Zeiss Axioplan microscope.
SCANNING ELECTRON MICROSCOPY
(SEM): Specimens were dissected and mac-erated as described for light microscopicobservation. Pieces of integument were dehy-drated (alcohol 80%, 95%, absolute alcohol),coated on a Balzers Union SCD 014 (FreieUniversitat Berlin) or Denton Vacuum DeskII(American Museum of Natural History), andobserved on a Philips SEM 515 (FreieUniversitat Berlin) or a Hitachi S-4700(American Museum of Natural History).
TERMINOLOGY: The terminology of glan-dular structures follows Farine (1987) andNoirot and Quennedey (1991). Sclerotizedelements of a glandular unit are referred toas ductule or efferent duct, and comprisea receiving canal, a saccule, and a conductingcanal, all of which are observable aftermaceration of the glandular unit.
I use the term trichome to describe a com-posite structure comprising a hairlike portionon the outer surface of the cuticle, which isassociated with a glandular unit on the insideof the cuticle. The term is used in plantmorphology, but also in entomology, where itrefers to a glandular hair, most commonly onethat exudes a secretion attractive to ants(Nichols, 1989).
ABBREVIATIONS: aset, anchor setae; cc,conducting canal of a ductule; du, ductule(efferent duct, i.e., the sclerotized elements) ofa glandular unit; gtp, grouped trichome pro-jection; gtsac, grouped trichome saccule; htf,hind tarsal fan; ltp, long trichome projection;ltsac, long-projection trichome saccule; ma,muscle attachment; po, pore of trichome orglandular unit; rec, receiving canal of ductule;
2006 WEIRAUCH: CAMOUFLAGE IN REDUVIID NYMPHS 3

rptp, rosettelike trichome projection; sac,saccule of a ductule; set, seta; sp1, abdominalspiracle 1; splat, sclerotized plate on abdom-inal tergites; stp, short trichome projection;stsac, short-projection trichome saccule; t1–t4,abdominal tergites 1–4; tar3, tarsomere three.
MATERIAL: Specimens studied belong tothe American Museum of Natural History(AMNH); were donated to the author byRalf Britz (Microtomus sp.), Peter Kott(Holotrichius tenebrosus) or Walter Sudhaus(Reduvius personatus); or were collected by theauthor (Peirates sp.; Rhynocoris erythropus). Adetermination to species level of nymphs notassociated with adult specimens can be pro-vided for only some of the taxa examined sinceimmatures are undescribed for most species ofReduviidae. Unless otherwise stated, the speci-mens were identified to species or genericlevel by the author. Cetherinae: Eupheno sp.,4th instar, Misiones, Argentina (AMNH).Hammacerinae: Microtomus sp., 3rd instar,USA, Florida, spring 2002, leg. R. Britz.Harpactorinae: Rhynocoris erythropus, 3rd in-star, France, Nimes, May 1999, leg. C.Weirauch. Peiratinae: Peirates sp., 3rd instar,Australia, Queensland, Bald Rock, June 1996,leg. C. Weirauch. Reduviinae: Acanthaspis sp.,3rd instar, India, South India: CoimbatoreDistrict, Marudamalar Hills, 1800 ft., Oct.1969, leg. P. S. Nathan (AMNH); 4th instar,Myanmar, February 1996, leg. C. Weirauch.Holotrichius tenebrosus, instars 1–5, labculture from P. Kott, det. P. Kott. Leogorrussp., 4th instar, Trinidad, W. I., ArimaValley, 800–1200 ft., Feb. 10–22, 1964, det.P. Wygodzinsky. Reduvius personatus, 4thinstar, USA, New Mexico, leg. W. Sudhaus.Salyavatinae: Salyavata sp., 5th instar,Guatemala, Tikal, Dec. 29, 1973, leg. D. H.,A. C., and K. M. C. Mistner, ex termite nest T820 (AMNH). Sphaeridopinae: Sphaeridopssp., 5th instar, [no locality label] (AMNH).Triatominae: Triatoma vitticeps, 3rd instar, [nolocality label], det. P. Wygodzinsky (AMNH).
RESULTS
The present study documents that immaturestages of certain Reduviidae (fig. 1) possessa range of specialized structures, which sup-posedly assist in applying and fastening the
camouflaging material used by these predatorsfor their disguise (figs. 2–9). According to theirfunctions, i.e., application versus fastening ofcamouflaging material, camouflaging struc-tures are here divided into two groups.
All species examined for which camouflagingbehavior was previously known were found topossess specialized camouflaging structures(table 1). This is true for Acanthaspis sp.,Holotrichius tenebrosus, Leogorrus sp., andReduvius personatus (Reduviinae); Salyavatasp. (Salyavatinae); and Triatoma vitticeps(Triatominae). Features that almost certainlyrepresent camouflaging structures were alsofound in two species for which no biologicaldata on immature stages exist, Eupheno sp.(Cetherinae) and Sphaeridops sp. (Sphaeri-dopinae). No camouflaging structures werefound in Microtomus sp. (Hammacerinae),Rhynocoris erythropus (Harpactorinae), andPeirates sp. (Peiratinae), for which camouflag-ing behavior has never been reported.
APPLICATION STRUCTURE: HIND TARSAL FAN
An application structure is located on thehind tarsus of the nymph of some species. It isformed by a fanlike arrangement of long, finesetae on the dorsoposterior margin of thedistal tarsomere (fig. 6E) and is here referredto as hind tarsal fan. Among the Reduviinaeobserved in this study, Acanthaspis sp.,Leogorrus sp., and Reduvius personatus(fig. 6E) possess a well-developed hind tarsalfan. This fan is also present in the species ofTriatominae examined here, Triatoma vitti-ceps. In addition, the hind tarsal fan haspreviously been described as ‘‘numerous verylong and delicate hairs’’, making it a characterof taxonomic and possibly group-definingimportance in nymphs of numerous speciesof Triatominae (Lent and Wygodzinsky,1979), but no reference was made to thepotential involvement of this structure incamouflaging behavior. Table 2 combinesresults from Zeledon et al. (1973) and Lentand Wygodzinsky (1979) and shows elevenspecies of Triatominae, for which bothcamouflaging habits and presence/absence ofthe hind tarsal fan are known. In these elevenspecies, presence of camouflaging behaviorcorrelates with the presence of the hind tarsal
4 AMERICAN MUSEUM NOVITATES NO. 3542

Fig. 1. Habitus and dorsal view of abdominal tergites of reduviid nymphs that possess camouflagingstructures. Only Holotrichius tenebrosus is shown in camouflaged condition, the other species are cleaned.
2006 WEIRAUCH: CAMOUFLAGE IN REDUVIID NYMPHS 5
fan and absence of camouflaging behaviorwith absence of the fan.
Among the species that possess fasteningstructures (discussed below), Eupheno sp.(Cetherinae), Holotrichius tenebrosus (Redu-
viinae), Salyavata sp. (Salyavatinae), andSphaeridops sp. (Sphaeridopinae) lack a hindtarsal fan. In H. tenebrosus, the fan is absentnot only in the 5th instar, as indicated intable 1, but in all five instars studied. The hindtarsal fan is also absent in the threes speciesthat lack fastening structures for camouflage(discussed below), Microtomus sp. (Ham-macerinae), Rhynocoris erythropus (Harpac-torinae), and Peirates sp. (Peiratinae).
The ventral surface of the distal tarsomereof Reduvius personatus is densely covered withsetae (fig. 6F). Some of these setae areconsiderably stouter than the rest and havea serrated ventral surface. Possibly, these setaeassist in loosening camouflaging particlesfrom the substrate.
FASTENING STRUCTURES: ANCHOR SETAE AND
TRICHOMES
Once the substrate particles have beenapplied to the abdominal and thoracic tergites,a second set of structures assures theirfastening. Two structural types of fastening
TABLE 1
Nymphal Instar, Presence and Absence of Hind Tarsal Fan, Trichomes, and Indication of CamouflagingLiterature for Taxa of Reduviidae Examined for Camouflaging Structures
Subfamily Genus/species Instar
Hind tarsal
fan
Trichomes
(gland + hair)
Behavior
previously
documented Citation
Cetherinae Eupheno sp. 4 absent present no Louis (1974)
documented for
closely related
Cethera spp.
Hammacerinae Microtomus sp. 3 absent absent no
Harpactorinae Rhynocoris erythropus 3 absent absent no
Peiratinae Peirates sp. 3 absent absent no
Reduviinae Acanthaspis sp. 5 present present yes Miller 1956, Odhiambo
1958, Louis 1974,
Brandt and
Mahsberg 2002
Holotrichius tenebrosus 5 absent present yes Dispons 1955, Kott
2001
Leogorrus sp. 4 present present yes Lent and Wygodzinsky
1979
Reduvius personatus 4 present present yes Weber 1930, Madel
1951, Immel 1955
Salyavatinae Salyavata sp. 5 absent present yes McMahan 1982, 1983
Sphaeridopinae Sphaeridops sp. 5 absent present no
Triatominae Triatoma vitticeps 3 present gland present,
hair absent
yes Zeledon et al. 1973
TABLE 2
Hind Tarsal Fan and Camouflaging Behavior inEleven Species of Triatominae According to Lent and
Wygodzinsky (1979) and Zeledon et al. (1973)
Hind tarsal
fan
Camouflaging
behavior
Dipetalogaster
maximus
present present
Panstrongylus herreri present present
P. megistus present present
Rhodnius prolixus absent absent
Triatoma barberi absent absent
T. infestans present present
T. lenti present present
T. maculata present present
T. protracta
navajoensis
absent absent
T. spinolai present present
T. vitticeps present present
6 AMERICAN MUSEUM NOVITATES NO. 3542

Fig. 2. Long-projection trichomes, short-projection trichomes, and grouped trichomes in nymphs ofReduviidae in the subfamilies Cetherinae, Reduviinae, Salyavatinae, Sphaeridopinae, and Triatominae.
2006 WEIRAUCH: CAMOUFLAGE IN REDUVIID NYMPHS 7

structures were found in this study, oneformed by setae, the other by trichomes. Thesetal fastening mechanism appears to bepurely mechanical in that it holds the camou-flaging material with the anchor-shaped tip ofthe seta. This type is here referred to as anchorseta and was found only in specimens belong-ing to the genus Acanthaspis (Reduviinae)(figs. 4B, C, E). A second type of fasteningstructure is also present in Acanthaspis: itcomprises a glandular unit that is associatedwith a hairlike projection, which serves asexcretory structure of the glandular unit. Thiscomposite structure is here referred to astrichome (figs. 2, 4B–G). The ductules of theglandular units (figs. 4F, G) together with theexternal projections (figs. 4B–D) of thesetrichomes may be observed with light(fig. 4F) or electron microscopy (fig. 4G) evenin macerated specimens, in which the glandcells were destroyed. Presumably, the fluidsecreted by this trichome is sticky and thusholds the camouflaging particles in place.
Trichomes were found in all species thatwere previously reported to possess camou-flaging behavior (table 1) with the exceptionof Triatoma vitticeps. In this species, althoughglandular units are present, they do not openthrough a hairlike projection but directlyonto the exterior surface of the integument(fig. 2). Trichomes were also found inEupheno sp. (Cetherinae) and Sphaeridops sp.(Sphaeridopinae), two of the five speciesexamined, for which camouflaging behaviorwas previously undocumented. No fasteningstructures were found in the remaining threespecies, for which indication of camouflagingis lacking, Microtomus sp. (Hammacerinae),Rhynocoris erythropus (Harpactorinae), andPeirates sp. (Peiratinae) (table 1).
The trichomes found in this study showsubstantial variation between taxa (table 3).In addition, more than one type of trichomemay be present in a single species (figs. 2, 6A,7B, D, 8, 9A, D). The trichomes on theabdomen (or on abdomen and thorax in some
Fig. 3. Eupheno sp. (Cetherinae): Camouflaging structures on the abdominal tergites in the 4th instarnymph. A. Sclerotized plate surrounded by membrane, showing short-projection trichomes, LM. B.Sclerotized plate surrounded by membrane, showing short-projection trichomes, SEM. C. Close-up of partof B, showing margin of sclerotized plate with variously shaped short-projection trichomes. D. Tubercle-shaped short-projection trichome. E. Spinelike short-projection trichomes. F. Pore of a spinelike short-projection trichome.
8 AMERICAN MUSEUM NOVITATES NO. 3542

instances) are described briefly for eachspecies in the following (table 3). The tri-chomes are characterized according to thelength and shape of their external projections,
size and shape of the internal saccules, andarrangement of trichomes. Three types oftrichomes (short-projection trichome, long-projection trichome, and grouped trichome)
Fig. 4. Acanthaspis sp. (Reduviinae): Camouflaging structures on the abdominal tergites in the 5th instarnymph, A–E external views, F, G, internal views. A. Tergites with median sclerotized plates as seen in theSEM. B. Posterior margin of one of the median sclerotized plate with hairlike long-projection trichomes andanchor setae. C. Two lateral sclerotized plates with long-projection trichomes and anchor setae. D. Spinelikeshort-projection trichomes. E. Apex of anchor setae. F. Ductules of long-projection trichomes, lightmicroscope. G. Ductule of one long-projection trichome and one short-projection trichome, SEM.
2006 WEIRAUCH: CAMOUFLAGE IN REDUVIID NYMPHS 9
are distinguished, which are here treated aspotential primary homologous structures.
Eupheno sp. (figs. 1–3): Abdomen with onlyone type of trichome, trichome with shortprojection, no grouped trichomes. Projectioneither shaped into low tubercle (fig. 3D) orspine (fig. 3E, F). Low tubercles only in areasadjacent to the sclerotized plates, which serveas points for muscle insertions (fig. 1, whitearrows; fig. 3A, B). Spines distributed over thetergites (fig. 3A). Saccules elliptical (l 5approximately 10 mm).
Acanthaspis sp. (figs. 1, 2, 4): Trichomeswith short projections and with long projec-tions on abdomen, no grouped trichomes.Short-projection trichomes with slender spine-like projections (fig. 4D) and elliptical saccules(l 5 approximate 30 mm) (fig. FG). Short-projection trichomes evenly distributed on theabdominal tergites. Long-projection trichomeswith very long, slender, hairlike projectionsand elliptical saccules (l 5 approximately40 mm) (figs. 4B, C, F, G). Long-projectiontrichomes on abdominal tergites largely re-
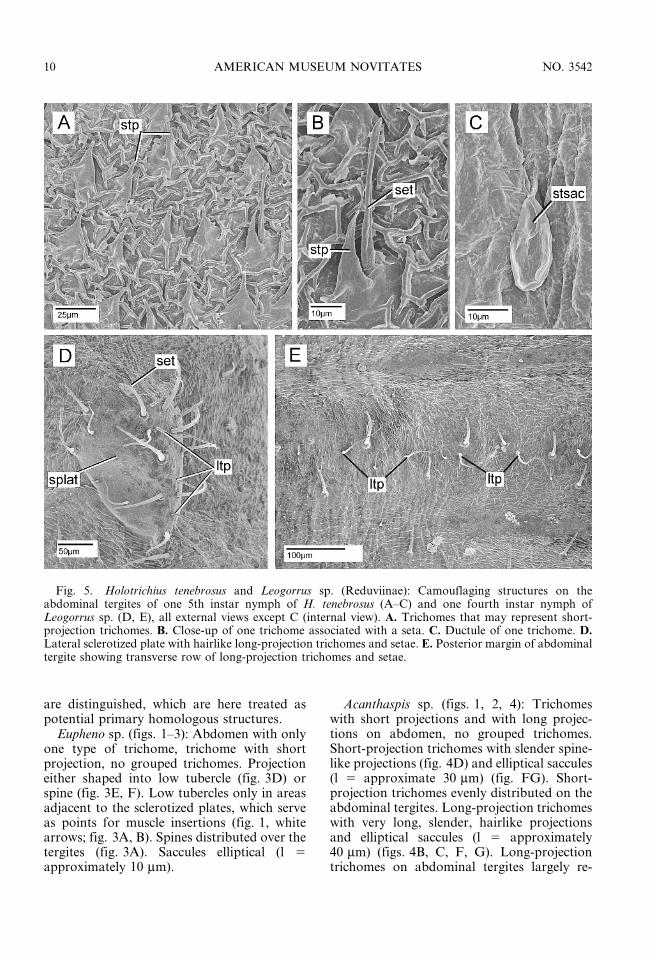
Fig. 5. Holotrichius tenebrosus and Leogorrus sp. (Reduviinae): Camouflaging structures on theabdominal tergites of one 5th instar nymph of H. tenebrosus (A–C) and one fourth instar nymph ofLeogorrus sp. (D, E), all external views except C (internal view). A. Trichomes that may represent short-projection trichomes. B. Close-up of one trichome associated with a seta. C. Ductule of one trichome. D.Lateral sclerotized plate with hairlike long-projection trichomes and setae. E. Posterior margin of abdominaltergite showing transverse row of long-projection trichomes and setae.
10 AMERICAN MUSEUM NOVITATES NO. 3542

stricted to sclerotized patches not representingthe sclerotizations associated with muscleattachments (figs. 1, 4A–C), but some tri-chomes also on the membranous parts of theabdominal tergites. No intimate association oflong-projection trichome with single seta, butloose assemblage of trichomes and setae onsclerotized plates (fig. 4B, C).
Holotrichius tenebrosus (figs. 1, 2, 5A–C):Only one type of trichome on abdomen(fig. 5A), possibly representing short-projec-tion trichomes; no grouped trichomes.Projections in the form of moderately elongatespines with blunt apices and with correspond-ing pear-shaped saccules (l 5 40 mm) (fig. 2).
The situation in this species is not easilycompared with that in other species, in whichshort projections co-occur with small sacculesand long projections with large, elliptical
saccules, since the relatively short projectionin H. tenebrosus is associated with very largepear-shaped saccules. Primary homology as-sessment of the trichome in this species withthe short-projection trichome in other speciesis thus tentative. Sometimes the trichome isassociated with a single seta (fig. 5B).
Leogorrus sp. (figs. 1, 2, 5D, E): Short- andlong-projection trichomes present on abdo-men, no grouped trichomes (figs. 2, 5D, E).Short-projection trichome with slender anderect spines (l 5 approximately 40 mm) andelliptical saccules (l 5 approximately 12 mm).These projections abundant on the abdominaltergites. Long-projection trichomes very longand hairlike (l 5 approximately 160 mm) andwith elliptical saccules (l 5 approximately30 mm). These trichomes arranged in a trans-verse row in the posterior area of the tergite
Fig. 6. Reduvius personatus (Reduviinae): Camouflaging structures on the abdominal tergites in thefourth instar nymph, external views with exception of D (internal view). A. Hairlike long-projectiontrichome associated with one seta and several short-projection trichomes, LM. B. Hairlike long-projectiontrichome associated with one seta and several short-projection trichomes, SEM. C. Short-projectiontrichome. D. Ductules of long-projection trichomes and short-projection trichomes. E. Hind-tarsal fan. F.Setae with serrated ventral surface on ventral side of third tarsomere.
2006 WEIRAUCH: CAMOUFLAGE IN REDUVIID NYMPHS 11

(fig. 5E) and occurring in patches on thelateral sclerotization of the tergites (fig. 5D).No intimate association of long-projectiontrichome with individual seta, but looseassemblage of trichomes and setae on sclero-tized plates (fig. 5D).
Reduvius personatus (figs. 1, 2, 6): Short-and long-projection trichomes present onabdomen, no grouped trichomes. Short-pro-jection trichome thorn-shaped with pointedand curved tips (l 5 approximately 20 mm)and with elliptical saccules (l 5 approximately10 mm) (figs. 2, 6A–D). These projectionsabundant on the abdominal tergites (fig. 6A).Long-projection trichomes hairlike (l 5 ap-proximately 150 mm) and with elliptical sac-cules (l 5 approximately 50 mm) (figs. 6A, B,D). Long-projection trichomes sparsely dis-
tributed on abdominal tergites. Long-projec-tion trichomes usually closely associated witha single seta with few, fine branches (figs. 2,6A, B).
Careful dismantling of the wood-dust layeron a living nymph showed that two layers ofcamouflaging may be distinguished: a layer ofvery fine wood dust closely adhered to theintegument and a relatively coarser layer ofwood and bark particles was attached furtherfrom the body.
Salyavata sp. (figs. 1, 2, 7, 8): Long-pro-jection trichomes and short-projection tri-chomes on the abdomen, grouped trichomespredominantly on the thorax. Short-projec-tion trichome spinelike (l 5 approximately30 mm), with saccules elliptical (l 5 approxi-mately 15 mm), sparsely distributed on the
Fig. 7. Salyavata sp. (Salyavatinae): Camouflaging structures on pronotum and abdominal tergites inthe 5th instar nymph, SEM, all external view with exception of F (internal view). A. Rows of groupedtrichomes on the pronotum. B. Grouped trichome. C. Long-projection trichome with associated seta. D.Sort-projection trichomes. E. Apex of long-projection trichome with pore. F. Ductule of long-projection trichome.
12 AMERICAN MUSEUM NOVITATES NO. 3542

abdomen (fig. 7D). Long-projection trichomeswith hairlike projection (l 5 approximately100 mm) and elliptical saccules (l 5 approxi-mately 25 mm length) (fig. 7C, F, 8). Long-projection trichomes rather sparsely distribut-ed on the abdomen, mostly on the lateralmargins of the abdominal tergites.
Trichomes on the thorax agree with abovedescription of the long-projection trichomes,but occur in groups of two to seven trichomes(figs. 7A, B) and not as single trichomes.Long-projection trichomes and grouped tri-chomes are usually closely associated with oneseta (figs. 2, 7B, C).
Sphaeridops sp. (figs. 1, 2, 9): No singletrichomes, two types of grouped trichomes,the one on the thorax consisting of group oflong-projection trichomes (figs. 9A–C), theone on the abdomen externally rosette-shaped
(figs. 9D–F). Both grouped trichomes withsaccules of approximately 25 mm length.
Triatoma vitticeps (fig. 2): No trichomes onthe abdomen, but saccules of approximately10 mm length occurring in low numbers on themembranous parts of the abdominal tergites.
DISCUSSION
The present study is the first comparativeapproach to the study of camouflaging struc-tures in nymphs of Reduviidae and severalstructures associated with camouflaging aredescribed here for the first time. Camouflagingstructures in Reduviidae comprise a hindtarsal fan, which may assist in applyingcamouflaging material to the body, and twotypes of fastening structures, anchor setae andtrichomes. Whereas the anchor setae presum-ably hold the camouflaging material mechani-cally, the trichomes probably secrete a stickysubstance that attaches the particles to theinsect. Among the trichomes observed, short-projection trichomes are distinguishable fromlong-projection trichomes, and in two of theobserved species trichomes are not alwaysdistinctly separated from each other but mayform grouped trichomes.
Among the eleven species studied here,trichomes were not only found in the sixspecies for which camouflaging had previouslybeen described, but also in two species withunknown immature biology. The presence ofcamouflaging structures in these latter speciessuggests the presence of camouflaging behav-ior in nymphs of Eupheno sp. (Cetherinae) andSphaeridops sp. (Sphaeridopinae).
HIND TARSAL FAN: The presence of longand fine setae on the hind tarsus, i.e., the hindtarsal fan, was previously described fornymphs of numerous species of Triatominae(Lent and Wygodzinsky, 1979). Zeledon et al.(1973) documented the presence or absence ofcamouflaging behavior for several species ofTriatominae. However, presence or absence ofthe hind tarsal fan was never before discussedin combination with the presence or absenceof camouflaging behavior in Triatominae.Table 2 shows that the presence of the hindtarsal fan correlates with the presence ofcamouflaging behavior in the eleven speciesfor which both factors have been documented.
Fig. 8. Salyavata sp.: Long-projection trichomewith associated seta, 5th instar nymph, LM.
2006 WEIRAUCH: CAMOUFLAGE IN REDUVIID NYMPHS 13

Aside from this intriguing correlation, thehind legs were reported to play the central rolein the process of gathering and loadingcamouflaging material onto the abdominaltergites in several species of Triatominae andother Reduviidae (McMahan 1982, Hase1940, Louis 1974, Kott 2001, Weber 1930).The fan, which is formed by long, fine setae,most likely assists in this process. The hindtarsal fan is present also in several Reduviinae,Acanthaspis sp., Leogorrus, and Reduviuspersonatus, and is here proposed to have thesame function as the hind tarsal fan inTriatominae. However, the application ofcamouflaging material must be possible with-out a hind tarsal fan, because the hind tarsalfan is absent in several species that are known
to camouflage or show camouflaging struc-tures, e.g., Eupheno sp. (Cetherinae), Holo-trichius tenebrosus (Reduviinae), Salyavatasp. (Salyavatinae), and Sphaeridops sp.(Sphaeridopinae). Future observation ofcamouflaging behavior in these species andexamination of substrate used for camouflagemay help to explain why some species possessan application structure and other species lackit. Furthermore, the hind tarsal fans inTriatominae and some species of Reduviinaeare not necessarily homologous structures, butmay have independent origins. This questionwill be answered only through a cladisticanalysis of Reduviidae.
Future research into systematic distributionand biological function of the serrated setae
Fig. 9. Sphaeridops sp. (Sphaeridopinae): Camouflaging structures on pronotum and abdominal tergitesin the 5th instar nymph, SEM, all external view with exception of F (internal view). A–C. Grouped trichomeson the pronotum. D, E. Rosettelike projection trichomes on the abdominal tergites. F. Saccules of therosettelike projection trichomes seen in the light microscope.
14 AMERICAN MUSEUM NOVITATES NO. 3542

TA
BL
E3
Ch
ara
cter
isti
cso
fS
ho
rt-P
roje
ctio
nT
rich
om
es,
Lo
ng
-Pro
ject
ion
Tri
cho
mes
,a
nd
Gro
up
edT
rich
om
esin
Eig
ht
Sp
ecie
so
fR
edu
viid
ae
Sh
ort
-tri
cho
me
pro
ject
ion
Sh
ort
-tri
cho
me
sacc
ule
Dis
trib
uti
on
Lo
ng-t
rich
om
e
pro
ject
ion
Lo
ng-t
rich
om
e
sacc
ule
Dis
trib
uti
on
Gro
up
ed
tric
ho
me
pro
jec-
tio
ns
Gro
up
ed
tric
ho
me
sacc
ule
Dis
trib
uti
on
Euphen
osp
.
(Cet
her
inae)
tub
ercl
eo
r
spin
e
elli
pti
cal,
,10
mmab
du
nd
an
t,
tub
ercl
escl
ose
tom
usc
le
att
ach
men
ts
ab
sen
tab
sen
tab
sen
tab
sen
tab
sen
tab
sen
t
Aca
nth
asp
issp
.
(Red
uvii
nae)
slen
der
erec
t
spin
e
elli
pti
cal,
,30
mmab
du
nd
an
tver
ylo
ng,
hair
lik
e
elli
pti
cal,
,40
mmd
ense
on
scle
rot.
pla
tes,
few
on
mem
br.
part
s
ab
sen
tab
sen
tab
sen
t
H.
teneb
rosu
s
(Red
uvii
nae)
spin
esw
ith
blu
nt
ap
ex
pea
r-sh
ap
ed,
,40
mmver
yab
un
dan
tab
sen
tab
sen
tab
sen
tab
sen
tab
sen
tab
sen
t
Leo
gorr
us
sp.
(Red
uvii
nae)
slen
der
erec
t
spin
e
elli
pti
cal,
,12
mmab
un
dan
tver
ylo
ng,
hair
lik
e
elli
pti
cal,
,30
mmin
tran
sver
se
row
so
nte
rgit
es,
an
do
nla
t.
scle
rot
patc
hes
,
no
tco
mm
on
ab
sen
tab
sen
tab
sen
t
R.
per
sonatu
s
(Red
uvii
nae)
tho
rnw
ith
po
inte
dan
d
curv
edti
p
elli
pti
cal,
,10
mmab
un
dan
tver
ylo
ng,
hair
lik
e
elli
pti
cal,
,50
mmir
regu
lar,
no
t
com
mo
n
ab
sen
tab
sen
tab
sen
t
Saly
ava
tasp
.
(Saly
avati
nae)
curv
edsp
ines
wit
hsu
bap
ical
po
re
elli
pti
cal,
,15
mmir
regu
lar
on
ab
do
men
lon
gan
d
slen
der
elli
pti
cal,
25
mmab
sen
tlo
ng
slen
der
hair
sin
gro
up
s
elli
pti
cal,
25
mmab
un
dan
t,
incl
ud
ing
late
ral
terg
al
spin
es
Sphaer
idops
sp.
(Sp
haer
ido
-
pin
ae)
ab
sen
tab
sen
tab
sen
tab
sen
tab
sen
tab
sen
tro
sett
eel
lip
tica
l,
20
mmab
un
dan
to
n
ab
do
men
,
tho
rax
wit
h
gro
up
ed
hair
-typ
e
tric
ho
mes
T.
vitt
icep
s
(Tri
ato
min
ae)
ab
sen
tel
lip
tica
l,
,10
mmn
ot
ver
y
com
mo
n
ab
sen
tab
sen
tab
sen
tab
sen
tab
sen
tab
sen
t
2006 WEIRAUCH: CAMOUFLAGE IN REDUVIID NYMPHS 15

on the ventral surface of the distal tarsomere,which were here documented only for Re-duvius personatus (fig. 6F) and hypothesizedto assist in loosening camouflaging particlesfrom the substrate, could provide furtherinsights into the process of camouflaging.
ANCHOR SETA: Among the eight examinedspecies that have fastening structures, only thetwo specimens of Acanthaspis from SouthIndia and Myanmar were found to possessanchor setae. Possibly, therefore, anchor setaerepresent a unique feature of species of thegenus Acanthaspis. Examination of additionalspecies of Acanthaspis and other, possiblyclosely related taxa such as Paredocla andEdocla, will be necessary to test this hypoth-esis.
It may be worth noting that the very longand slender anchor setae occur in the onlytaxon examined for which corpse camouflagehas been reported (Ambrose, 1999; Brand andMahsberg, 2002; Louis, 1974; Odhiambo,1958). Possibly the comparatively heavy andbulky corpse cover requires mechanical fas-tening in addition to the adhesion offered bythe trichome secretions. Examination of theother two taxa for which corpse camouflageis known, Edocla (Ambrose, 1999) andParedocla (Brand and Mahsberg, 2002), mayprovide additional evidence for this hypothe-sis.
PRIMARY HOMOLOGY OF SHORT- AND LONG-PROJECTION TRICHOMES: Given the compara-tive approach of this study, but the lackof understanding of relationships withinReduviidae, only tentative primary homologystatements may be formulated for the tri-chomes. A distinction between short- andlong-projection trichomes seems evident inthose cases where both types of trichomesappear in one species, because the two typesare clearly differentiated by the shape andlength of the projections and the size of thesaccules. Such clear distinctions are seen inthree taxa of Reduviinae, Acanthaspis,Leogorrus, and Reduvius (table 3). In thesethree species, short-projection trichomes are ofroughly similar size (projection and saccule) asare the long-projection trichomes. A primaryhomology statement for the short-projectiontrichomes as well as the long-projectiontrichomes among these taxa seems tenable.
Holotrichius tenebrosus is difficult to interpretin this context: only one type of trichome ispresent, which comprises relatively short,blunt, spinelike projections of approximately20 mm length which are sometimes associatedwith a seta, but which have very large saccules(approximately 30 mm). Considering the equaland rather dense distribution of the trichomeson the abdominal tergites, it appears reason-able to interpret them as short-projectiontrichomes. Examination of additional speciesof this genus may provide further insights.
With respect to size and shape of projectionand saccule, the short-projection trichomein Eupheno (Cetherinae) and Salyavata(Salyavatinae) may be considered primaryhomologous to the short-projection trichomesin Reduviinae.
POTENTIAL FUNCTION OF SHORT- AND LONG-PROJECTION TRICHOMES: Short- and long-projection trichomes and anchor setae mayserve different functions: Brandt and Mahs-berg (2002) found that two layers of camou-flaging material are applied by Acanthaspis,consisting of a dust cover close to the in-tegument, and a cover of corpses and othercoarse material that is further removed fromthe body. Short-projection trichomes andlong-projection trichomes plus anchor setae,respectively, may be responsible for these twolayers. Brandt and Mahsberg (2002) couldfurther demonstrate that the dust cover playsa role in aggressive mimicry, whereas thecorpse cover seems to be important as pro-tection from potential predators.
A two-layer camouflage is also present inReduvius personatus, although in a less spec-tacular way. Examination of a living nymphduring this project showed that one layer ofcamouflaging material consists of small parti-cles that adhere very closely to the integument,whereas coarser substrate particles form a sec-ond layer that is further removed from theintegument. Whether the two layers of thiscover serve the same function as the layers inAcanthaspis remains open to speculation andfurther experimentation.
SETA ASSOCIATED WITH LONG-PROJECTION
TRICHOME: At least in some instances, thelong-projection trichome is associated witha single, long, slender seta, which may bemechanosensory. This association is seen in
16 AMERICAN MUSEUM NOVITATES NO. 3542

Reduvius personatus, in which the seta isslightly plumose, in Salyavata sp., and alsoin the single type of trichome present inHolotrichius tenebrosus. In Acanthaspis andLeogorrus, long-projection trichomes are moreloosely associated with setae on the sclerotizedplates; setae are rather evenly distributed onthe thorax and abdomen in Sphaeridops.Eupheno sp. lacks long-projection trichomesas well as setae in association with trichomes.Assuming that the setae close to the trichomesare in fact mechanoreceptive, they mightprovide information to the animal on its‘‘loading status’’.
GROUPED TRICHOMES IN SALYAVATINAE
AND SPHAERIDOPINAE: Among the examinedspecies, only Salyavata sp. (Salyavatinae) andSphaeridops sp. (Sphaeridopinae) possessgrouped trichomes. Given the length and shapeof their projections and size of their saccules,these trichomes might be hypothesized as de-rived from long-projection trichomes thatgrouped together. Salyavata sp. possesses a com-bination of single long-projection trichomes andgrouped trichomes with long projections.Sphaeridops, in contrast, possesses only groupedtrichomes, which comprise trichomes with longprojections on the thorax and rosette-shapedtrichomes, i.e., trichomes whose exterior projec-tions are extremely short, on the abdomen. Thesimilar aspect of the grouped long-projectiontrichomes on the thorax of Salyavata sp. withthose on the thorax in Sphaeridops sp. makesa hypothesis on primary homology of thesestructures reasonable. When it is also taken intoaccount that Salyavatinae and Sphaeridopinaeshare several other morphological characteris-tics, such as highly modified mouthparts, unlikethose seen in any species among Reduviidae,and similar structure of the antenna and femalegenitalia (personal obs., unpubl.), a closer re-lationship of these two taxa does appear likely.The rosette-shaped trichomes on the abdomenof Sphaeridops might then be derived fromgrouped long-projection trichomes of the typeseen on the thorax in Sphaeridops and inSalyavata by a drastic shortening in thoseprojections.
OUTLOOK: The present paper providesa first and albeit preliminary approach to thecomparative study of camouflaging structuresin Reduviidae. As more nymphs of
Reduviidae become available, the picturesketched in this paper will be improvedconsiderably. Including camouflaging struc-tures in a cladistic analysis of Reduviidae,currently in preparation, will test the homol-ogy of certain structures described here, suchas the hind tarsal fan, the short- and long-projection trichomes, and the grouped tri-chomes in Salyavata and Sphaeridops. Finally,improved taxon sampling together with a cla-distic analysis will shed light on where andhow often camouflaging structures and there-fore camouflaging behavior evolved withinReduviidae.
ACKNOWLEDGEMENTS
Randall Toby Schuh (AMNH) allowed meto study several immature specimens of PedroWygodzinsky’s alcohol collection, and RalfBritz, Peter Kott, and Walter Sudhaus kindlydonated additional specimens studied duringthis project. To all these individuals a warm‘‘thank you’’.
The results presented here are largely de-rived from my PhD dissertation, and wereobtained while working at the Freie Uni-versitat Berlin and during a three-month visitto the AMNH. I thank Walter Sudhaus forsupervising my dissertation and the AGEvolutionsbiologie for discussions. For finan-cial support I would like to acknowledgethe Nachwuchsforderungsgesetz des LandesBerlin (NaFoG), Anette Kade GraduateStudent Fellowship Program at the AMNH,and the Berliner Programm zur Forderung derChancengleichheit fur Frauen in Forschungund Lehre.
Thanks also to Randall Toby Schuh andDimitri Forero for kindly commenting on themanuscript.
REFERENCES
Ambrose, D.P. 1999. Assassin bugs. Enfield:Science Publishers, 337 pp.
Brandt, M., and D. Mahsberg. 2002. Bugs witha backpack: the function of nymphal camou-flage in the West African assassin bugsParedocla and Acanthmaspis spp. AnimalBehaviour 63: 277–284.
2006 WEIRAUCH: CAMOUFLAGE IN REDUVIID NYMPHS 17

Breddin, G. 1904. Rhynchoten aus Ameisen- undTermitenbauten. Annales de la SocieteEntomologique Belge 48: 407–416.
Butler, E.A. 1923. A biology of the BritishHemiptera-Heteroptera. London: H. F. & G.Witherby, 682 pp.
Cai, W., P. Zhao, and Q. Mi. 2002. Camouflagingin assassin bugs. Entomological Knowledge 39:317–319. [In Chinese, English summary]
Dispons, P. 1955. Les Reduviides de l’Afrique nord-occidentale. Memoires du Museum Nationald’Histoire Naturelle, Nouvelle Serie Serie AZoologie 10: 93–240.
Eisner, T., K. Hicks, and D.S. Robson. 1978. Wolf-in-sheep’s-clothing strategy of a predaciousinsect larva. Science 199(4330): 790–794.
Fabre, J.-H. 1903. Souvenirs entomologiques. 8eserie. VI. Le Reduve a masque. Paris, 380 pp.
Fabricius, J.C. 1775. Systema entomologiae, sistensinsectorum classes, ordines, genera, species,adjectis synonymis, locis, descriptionibus, etobservationibus. Kortii: Flensburgi & Lipsiae,i–xxx, 832 pp.
Farine, J.-P. 1987. The exocrine glands ofDysdercus cingulatus (Heteroptera, Pyrrho-coridae): morphology and function of thenymphal glands. Journal of Morphology 194:195–207.
Harz, K. 1952. Ein Beitrag zur Biologie vonReduvius personatus L. (Rynchita [sic!]/Heteroptera). Nachrichtenblatt der Baye-rischen Entomologen 1(10): 73–75.
Hase, A. 1940. Uber Triatoma dimidiata(Hemiptera, Triatomidae). I. Teil. Zeitschriftfuer Parasitenkunde 11: 419–429.
Immel, R. 1955. Zur Biologie und Physiologie vonReduvius personatus L. Zeitschrift fuerMorphologie und Oekologie der Tiere 44:163–195.
Kott, P. 2001. Holotrichius tenbrosus Burmeister,1835 (Hemiptera): Beobachtungen zurMaskierung der Larven. Verhandlungen desWestdeutschen Entomologen Tags 2000:229–232.
Lent, H., and P. Wygodzinsky. 1979. Revision ofthe Triatominae (Hemiptera, Reduviidae), andtheir significance as vectors of Chagas’ disease.Bulletin of the American Museum of NaturalHistory 163: 123–520.
Linnaeus, C. 1758. Systema naturae per regna trianaturae, secundum classes, ordines, genera,species, cum characteribus, differentiis, syno-nymis, locis. Editio decima, reformata. Salvii:Holmiae, i–v, 824 pp.
Louis, D. 1974. Biology of Reduviidae of cocoafarms in Ghana. American Midland Naturalist91: 68–89.
Madel, W. 1951. Beobachtungen an der Staub-wanze Reduvius personatus L. Zeitschrift fuerParasitenkunde 15: 102–108.
McMahan, E.A. 1982. Bait-and-capture strategy ofa termite-eating assassin bug. Insectes Sociaux29: 346–351.
McMahan, E.A. 1983a. Bugs angle for termites.Natural History 83: 40–46.
McMahan, E.A. 1983b. Adaptations, feeding pref-erences, and biometrics of a termite-baitingassassin bug (Hemiptera: Reduviidae). Annalsof the Entomological Society of America 76:483–486.
Miller, N.C.E. 1953. Notes on the biology of theReduviidae of Southern Rhodesia. Tran-sactions of the Zoological Society of London27: 541–672.
Miller, N.C.E. 1956. Biology of the Heteroptera.London: Leonard Hill Books, 162 pp.
Nichols, S.W. 1989. The Torre-Bueno glossary ofentomology. New York: Entomological So-ciety, 840 pp.
Noirot, C., and A. Quennedey. 1991. Glands, glandcells, glandular units: some comments onterminology and classification. Annales de laSociete Entomologique de France 27: 123–128.
Odhiambo, T.R. 1958. The camouflaging habits ofAcanthaspis petax Stal (Hem., Reduviidae) inUganda. Entomologist’s Monthly Magazine94: 47.
Readio, P.A. 1927. Studies on the biology of theReduviidae of America north of Mexico.University of Kansas Science Bulletin 17:5–291.
Stitz, H. 1931. Planipennia. In P. Schulze (editor),Biologie der Tiere Deutschlands. Teil, 35:68–304. Berlin: Borntraeger.
Villiers, A. 1948. Faune de l’empire francais. IX.Hemipteres Reduviides de l’Afrique noire.Paris: Edition du Museum, 488 pp.
Weber, H. 1930. Biologie der Hemipteren. Berlin:Julius Springer, 543 pp.
Wygodzinsky, P., and R.L. Usinger. 1946. Thegenus Reduvius Fabricius in western NorthAmerica (Reduviidae, Hemiptera, Insecta).American Museum Novitates 2175: 1–15.
Zeledon, R., C.E. Valerio, and J.E. Valerio. 1973.The camouflage phenomenon in several speciesof Triatominae (Hemiptera, Reduviidae).Journal of Medical Entomology 10: 209–211.
18 AMERICAN MUSEUM NOVITATES NO. 3542