analysis of mir-195 and mir-497 expression, regulation...
TRANSCRIPT

Human Cancer Biology
Analysis of MiR-195 and MiR-497 Expression, Regulation andRole in Breast Cancer
Dan Li1, Yulan Zhao1, Changxing Liu2, Xiaona Chen1, Yanting Qi8, Yue Jiang1,8, Chao Zou1, Xiaolong Zhang1,Shunying Liu1, Xuejing Wang5, Dan Zhao1, Qiang Sun5, Zhenbing Zeng3, Andreas Dress3, Marie C. Lin7,1,Hsiang-Fu Kung6,1, Hallgeir Rui8, Ling-Zhi Liu8, Feng Mao5, Bing-Hua Jiang4,8, and Lihui Lai1
AbstractPurpose: To investigate expression, regulation, potential role and targets of miR-195 and miR-497 in
breast cancer.
Experimental Design: The expression patterns of miR-195 and miR-497 were initially examined in
breast cancer tissues and cell lines by Northern blotting and quantitative real-time PCR. Combined bisulfite
restriction analysis and bisulfite sequencing were carried out to study the DNA methylation status of miR-
195 and miR-497 genes. Breast cancer cells stably expressing miR-195 and miR-497 were established to
study their role and targets. Finally, normal, fibroadenoma and breast cancer tissues were employed to
analyze the correlation between miR-195/497 levels and malignant stages of breast tumor tissues.
Results:MiR-195 andmiR-497 were significantly downregulated in breast cancer. The methylation state
of CpG islands upstream of the miR-195/497 gene was found to be responsible for the downregulation of
both miRNAs. Forced expression of miR-195 or miR-497 suppressed breast cancer cell proliferation and
invasion. Raf-1 and Ccnd1 were identified as novel direct targets of miR-195 and miR-497. miR-195/497
expression levels in clinical specimens were found to be correlated inversely with malignancy of breast
cancer.
Conclusions: Our data imply that both miR-195 and miR-497 play important inhibitory roles
in breast cancer malignancy and may be the potential therapeutic and diagnostic targets. Clin Cancer
Res; 17(7); 1722–30. �2011 AACR.
Introduction
MicroRNAs (miRNA) are a class of highly conserved,small noncoding RNAs, which regulate gene expression in avariety of eukaryotic organisms and execute pivotal roles in
physiological and pathological processes including devel-opment, differentiation, metabolism, immunity, cell pro-liferation and apoptosis (1–3). In general, these single-stranded miRNAs show a trend toward multiple targetingsites in the coding sequence (CDS) regions or the 30
untranslated (30UTR) regions of their target messengerRNAs (mRNAs) and cause translational repression ormRNA cleavage (4). There is growing evidence that differ-ential expressions of miRNAs are associated with tumortypes and cancer development (5).
Breast cancer is one of the leading causes of cancer death inwomen worldwide (6). miRNAs, such as miR-21, miR-27a,miR-17, miR-155, miR-10b, miR-125b, and miR-145, arefound to be dysregulated and serve as oncogenic agent ortumor suppressor in breast cancer (6–11). It has been knownthat these small molecules are involved in tumor cell pro-liferation, migration, invasiveness and metastasis. Recently,blood-based miRNA profiling studies demonstrate that sys-temic miR-195 levels are increased before surgery anddecreased after surgery in breast cancer patients (12). Inter-estingly, miR-497 has been found to be one of most pro-minently downregulatedmiRNAs inmale breast cancer (13).AsmiR-195/497 is a highly conservedmiRNA cluster locatedat Chromosome 17p13.1 (6), it would be interesting tosimultaneously study the roles of miR-195 and miR-497in breast cancer.
Authors' Affiliations: 1Institute of Molecular and Chemical Biology, EastChina Normal University; 2Department of General Surgery, Huashan Hos-pital, Shanghai Medical College, Fudan University; 3CAS-MPG PartnerInstitute and Key Lab for Computational Biology, SIBS, CAS, Shanghai,China; 4Department of Pathology, Jiangsu Key Lab of Cancer Biomarkers,Prevention and Treatment, Cancer Center, Nanjing Medical University,Nanjing, China; 5Department of Breast Surgery, Peking Union MedicalCollege Hospital, Chinese Academy of Medical Sciences and PekingUnion Medical College, Beijing, China; 6Faculty of Medicine, The ChineseUniversity of Hong Kong, HK, SAR, China; 7Brain Tumor Center, Neuro-surgery Division, Faculty of Medicine, PWH, The Chinese University ofHong Kong, Shatin, Hong Kong, China; and 8Department of Pathology,Anatomy and Cell Biology, Kimmel Cancer Center, Thomas JeffersonUniversity, Philadelphia, Pennsylvania
Note: Supplementary data for this article are available at Clinical CancerResearch Online (http://clinicalcancerres.aacrjournals.org/).
Note: D. Li, Y. Zhao, and C. Liu contributed equally to this work.
Corresponding Authors: Lihui Lai, Institute of Molecular and ChemicalBiology, East China Normal University, Shanghai 200062, China. Phone:86-21-62237226; Fax: 86-21-62237226; E-mail: [email protected] Bing-Hua Jiang, Department of Pathology, Nanjing Medical Univer-sity, Nanjing, China. Phone: 215-5036147; E-mail: [email protected]
doi: 10.1158/1078-0432.CCR-10-1800
�2011 American Association for Cancer Research.
ClinicalCancer
Research
Clin Cancer Res; 17(7) April 1, 20111722
on May 8, 2018. © 2011 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 24, 2011; DOI: 10.1158/1078-0432.CCR-10-1800

Accumulating evidence also demonstrates that geneticand epigenetic alterations cause the dysregulation of sub-sets of miRNAs (14, 15). Among them, promoter hyper-methylation is known for silencing the specific miRNAexpression to be involved in tumorigenesis (15). Mamma-lian DNA is predominantly methylated at the C-5-positionof complimentaryCpGbpbyDNAmethyltransferases (16).The aberrations of this epigenetic modification may leadto various diseases including cancer (15, 17). For miRNAexpression, DNA methylation of CpG islands was foundto be present in the promoter region of miRNAs withtumor suppressor features in human cancer such as miR-127, miR-124a, miR-1, as well as miR-148a and miR-34b(18–21). Thus, such DNA methylation might also play arole in tumorigenesis. The objective of this study was toreveal the roles of miR-195 and miR-497 in breast cancer.
Materials and Methods
Human tissue samplesHuman breast tumor samples and normal tissues were
obtained from Peking UnionMedical College Hospital andShanghai Huashan hospital. All participants provided writ-ten informed consent following institutional review boardapproval at the participating hospitals. Tissue samples werecollected and frozen in liquid nitrogen. These tissues wereprepared in the hospital biorepository, and clinical anno-tation is available through a database. Cases were classifiedand selected based on diagnosis using the CoPath Ana-tomic Pathology system, and no information regulated bythe Health Insurance Portability and Accountability Act(HIPPA) was included in the study, which qualifies for thestatus of NIH Exemption #4.
Cell culture and RNA preparationHuman breast cancer cell lines MDA-MB-231, MDA-MB-
435s, MDA-MB-453, ZR-75-30, SK-BR-3, T47D, andMCF 7were obtained from ATCC. Cells were maintained inDulbecco’s modified Eagle’s medium (DMEM; MDA-MB-231, MDA-MB-435s, MDA-MB-453, and T47D) or RPIM1640 medium (ZR-75-30, SK-BR-3) supplemented with10% FBS. MCF7 cells were maintained in MEM supple-mented with 10% FBS, and 0.01 mg/mL bovine insulin.Total RNAs were extracted from tissues and cells usingTrizol reagent (Invitrogen).
Combined bisulfite restriction analysis and bisulfitesequencing.
The University of California Santa Cruz (UCSC) data-base was used to identify CpG islands (CGI) spanningmiR-195 and miR-497 genes. Genomic DNA was isolated usingUniversal Genomic DNA Extraction Kit Ver.3.0 (Takara),and bisulfite conversion was performed using EZ DNAMethylation-Gold Kit� (Zymo Research). Bisulfite-con-verted genomic DNA that converts only unmethylatedcytosines to uracils, was amplified with specific primers.The purified PCR fragments were cloned into a pMD19-Tvector (Takara Code: D102A), and individual clones weresequenced. For combined bisulfite restriction analysis(COBRA), PCR fragments were digested with Taq I (NewEngland Biolabs Inc.) for 3 hours at 37�C. The restrictionproducts were electrophoresed on a 3% agarose gel, andvisualized by staining with Gold View (SBS Genetech). Theprimers used for amplification of CGI on miR-195 andmiR-497 genes are: miR-195 and miR-497-CG-BSF1:GTGTTTATTTGTAGTGATTT; miR-195 and miR-497-CG-BSR1: TAACTCCCTCAATCTCTTATTCTT.
5-Aza-dC treatment and precursor transfectionZR-75-30 cells were seeded 24 hours before treatment
with 5 mM 5-aza-20-deoxycytidine (5-Aza-dC; Sigma-Aldrich). After 5 days of treatment, the cells were harvestedand total RNAs were prepared and used for quantitativereal-time PCR analysis.
Pre-miR-195 and pre-miR-497 and pre-miR� miRNAprecursors were purchased from Ambion. Cells were trans-fected with the precursors at 100 nM using LipofectAMINE2000 reagent (Invitrogen).
Northern blot analysisTotal RNAs (25 mg each) were electrophoresed on 15%
acrylamide and 8 M urea denature gels, and transferredonto Hybond Nþmembrane (Amersham Biosciences). Themembranes were baked at 80�C for 2 hours before hybri-dization, then hybridized with oligo-nucleotide probescorresponding to the complementary sequences of thefollowing mature miRNAs: miR-195, 50-GCCAA-TATTTCTGTGCTGCTA-30 and miR-497, 50-ACAAACCA-CAGTGTGCTGCTG-30. Probes were 5-end labeled usingthe polynucleotide kinase in the presence of [g-32P] ATP.Hybridization was performed at 39�C in ULTRAhyb�-Oligo Hybridization Buffer (Ambion) for 16 hours. Mem-branes were washed at 42�C, 3 times with 2� SSC with0.1% SDS. The membranes were rehybridized after strip-ping the oligo nucleotides used as probes in 1% SDS for 30minutes at 65�C. The U6 RNA (50-GCTAATCTTCTCTG-TATCGTTCCAATTTT-30) was used as an internal control.
Real-time polymerase chain reaction assays formature miRNAs
Total RNAs were prepared from tissue samples and celllines for the analysis. The expression levels of miR-195and miR-497 were assayed using the primers designed byBiomics Company. Reverse transcription reaction was
Translational Relevance
In this study, we show that the promoter hypermethy-lation of miR-195/497 cluster causes their downregula-tion in breast cancer. The levels of miR-195/497expression are inversely correlated with tumor malig-nancy, and Raf-1 is identified as a direct target of bothmiRNAs. Our data imply that both miR-195 and miR-497 may be of potential interest as therapeutic anddiagnostic targets in breast cancer.
MiR-195 and MiR-497 in Breast Cancer
www.aacrjournals.org Clin Cancer Res; 17(7) April 1, 2011 1723
on May 8, 2018. © 2011 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 24, 2011; DOI: 10.1158/1078-0432.CCR-10-1800

performed using 30 ng of total RNAs and 1 mM loopedprimers. Real-time PCR was performed using the standardSYBR Green Assay protocol by the MX3000p Real-TimePCR Detection System (Stratagene). The 25 mL PCR reac-tion included 2 mL reverse transcription product, 1� PCRMaster Mix (Takara,) 1.5 mmol/L forward primer, and 0.7mmol/L reverse primer. The reactions were incubated in a96-well plate at 95�C for 10 seconds, followed by 40 cyclesof 95�C for 5 seconds, and 60�C for 30 seconds. The 2�Ct
method for relative quantification of gene expression wasused to determine miRNA expression levels. Each samplewas analyzed in triplicate, and U6 RNA was used tonormalize the miRNA levels.
Western blot analysisFor each sample, 25 mg of total protein extracts were
separated on SDS-PAGE gels, and transferred to PVDFmembrane. All the primary antibodies were incubatedovernight at 4�C, followed by the incubation with AP-conjugated goat anti-rabbit secondary antibody and detec-tion with BCIP/NBT (Amresco). Bands were quantifiedwith Labworks Instrument software (UVP LLC).
Soft agar colony-formation assaysMCF7 and ZR-75-30 cells were transfected withmiR-195,
miR-497, or negative control precursor miRNAs. After 24hours of transfection, cells were suspended in completemedium containing 0.35% agar, and overlaid on 0.6% agarin 6-well plates (2 � 103 cells/well). Each assay was per-formed in triplicates. The plates weremaintained for up to 2weeks. The number of colonies (>50 mm in diameter) wascounted under a microscopic field at 10� magnification.
Fluorescence-activated cell sorting (FACS) cell cycleanalysis
MCF7 and ZR-75-30 cells were seeded at 30% confluencein 12-well plate 24 hours before transfection. Cells weretreated with nocodazole for 16 to 20 hours after thetransfection of miR-195, miR-497, or negative controlprecursor at 100 nM; and fixed in 70% ethanol at 4�Cfor 24 hours. Fixed cells were then washed once with PBS,resuspended in 500 mL PBS containing10 mg/mL propi-dium iodide and 50 mg/mL RNase, and incubated for 30minutes at room temperature. The cells were then analyzedwith Fluorescent-Activated Cell Sorter (BD FACSaria cellsorter, BD Bioscences).
Lentivirus preparation, titration and infectionHEK293T cells were seeded in 10 cm plate at 6 � 106
cells, and transfected with 9 mg of pLemir-195 or pLemir-497 plasmid and 26 mL of the packaging mix stock usingArrest-In transfection reagent (Openbiosystems). The cul-ture supernatants were collected 48 and 72 hours aftertransfection, filted through 0.45-mm filters, and stored asvirus stocks. For virus titration, TLA-HEK293T cells wereseeded at 5 � 104 cells per well with 24-well tissue cultureplate in DMEM. The diluted virus were added to each well,and incubated at 37�C for 4 hours. Then, the transduction
mix was removed from cultures, cells were cultured for 48hours for the virus titration. For virus infection, cells wereincubated at 37�Cwith virus and polybrene (8 mg/mL) for 6hours, then for 2 days to select puromycin-resistant cells.
miRNA target prediction and luciferase activity assayMiRNA target prediction was performed by in-lab devel-
oped algorithm KeyTar. KeyTar miRNA target prediction isbased on miRNA: target sequence, structure, and function(unpublished). The human 30 UTR region of Ccnd1 genewas amplified by PCR using the following primers: Ccnd1–3UTR-F: 50-GGTACCGTTTGGCGTTTCCCAGAGT-30, andCcnd1–3UTR-R: 50-CGTCTAGATGGCTAAGTGAAGCAT-GAGG-30, and cloned into the KpnI and XbaI sites of thepGL3-control vector (Promega). Thehuman30 UTRof Raf-1genewas amplified byPCRusing the followingprimers: Raf-1–3UTR-F: 50-GAATTCGCAATGAAGAGGCTGGTA-30, andRaf-1–3UTR-R: 50- CTCGAGGCCCAAAGGGATAGAAA-30,and cloned into the EcoRI and XhoI sites of the pGL3-control vector. Nucleotide-substitutionmutations were car-ried out using PCR-based methods at the 30UTR regions ofCcnd1 and Raf-1. Primers were as below: Mut 30UTR ofCcnd1: 50-CGACGAACCGTTGACTTCCAGGCAC-30 and 50-GCAATAAGAAAATGGAGCTGCGGCCT-30; andMut 30UTRof Raf-1: 50-CGACGAGCTAAGGACCTTCTAGACT-30 and50-TTCTCTGAAAACATGTGTTCTGCCTC -30.
All constructswere verified by sequencing. The underlinednucleotides indicate the bases where mutations were made.For luciferase assay, the 293A cells were cultured in 24-wellplates, and transfected with 100 ng luciferase reporter plas-mid, 5 ng pRL-TK vector expressing the Renilla luciferase(Promega), and 50 pmoles ofmiR-195, miR-497, ormiRNAnegative control precursor. Transfection was performedusing Lipofectamine 2000 (Invitrogen). After transfectionfor 36 hours, firefly and renilla luciferase activities weremeasured using the Dual-Luciferase Reporter Assay (Pro-mega). Each transfection was repeated twice in triplicate.
Statistical analysisThe results are expressed as mean � SD. Data compar-
isons were performed using analysis of variance followedby Dunnett’s method using Microsoft Excel. The Mann–Whitney test was used to compare depot-specific differ-ences between cancer tissues and normal controls. Theresults were considered statistically significant at P < 0.05.
Results
MiR-195 and miR-497 are downregulated in humanbreast cancer tissues and cell lines
Initially, we examined the expression levels of miR-195and miR-497 in breast cancer tissues. The expression levelsof both miR-195 and miR-497 in breast cancer tissues weregreatly decreased when compared with those in the normaladjacent tissue (Fig. 1A and B). Similarly the levels of miR-195 and miR-497 were lower in different breast cancer celllines includingMCF7, ZR-75-30,MDA-453,MDA-435, andMDA-231 (Fig. 1B and C), indicating that miR-195 and
Li et al.
Clin Cancer Res; 17(7) April 1, 2011 Clinical Cancer Research1724
on May 8, 2018. © 2011 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 24, 2011; DOI: 10.1158/1078-0432.CCR-10-1800
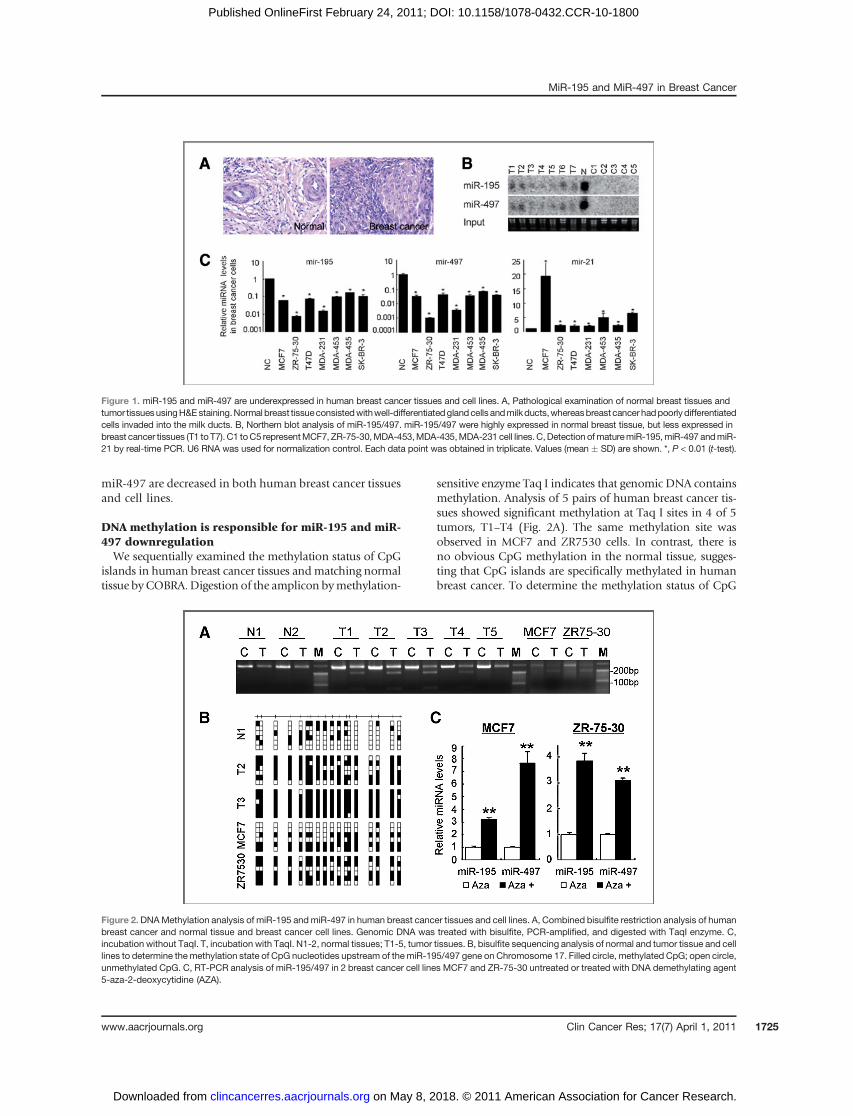
miR-497 are decreased in both human breast cancer tissuesand cell lines.
DNA methylation is responsible for miR-195 and miR-497 downregulationWe sequentially examined the methylation status of CpG
islands in human breast cancer tissues andmatching normaltissue byCOBRA.Digestion of the amplicon bymethylation-
sensitive enzyme Taq I indicates that genomic DNA containsmethylation. Analysis of 5 pairs of human breast cancer tis-sues showed significant methylation at Taq I sites in 4 of 5tumors, T1–T4 (Fig. 2A). The same methylation site wasobserved in MCF7 and ZR7530 cells. In contrast, there isno obvious CpG methylation in the normal tissue, sugges-ting that CpG islands are specifically methylated in humanbreast cancer. To determine the methylation status of CpG
Figure 1. miR-195 and miR-497 are underexpressed in human breast cancer tissues and cell lines. A, Pathological examination of normal breast tissues andtumor tissuesusingH&Estaining.Normal breast tissueconsistedwithwell-differentiatedglandcells andmilkducts,whereasbreastcancer hadpoorlydifferentiatedcells invaded into the milk ducts. B, Northern blot analysis of miR-195/497. miR-195/497 were highly expressed in normal breast tissue, but less expressed inbreast cancer tissues (T1 toT7). C1 toC5 representMCF7, ZR-75-30,MDA-453,MDA-435,MDA-231 cell lines.C,Detection ofmaturemiR-195,miR-497 andmiR-21 by real-time PCR. U6 RNA was used for normalization control. Each data point was obtained in triplicate. Values (mean � SD) are shown. *, P < 0.01 (t-test).
Figure 2. DNAMethylation analysis of miR-195 andmiR-497 in human breast cancer tissues and cell lines. A, Combined bisulfite restriction analysis of humanbreast cancer and normal tissue and breast cancer cell lines. Genomic DNA was treated with bisulfite, PCR-amplified, and digested with TaqI enzyme. C,incubation without TaqI. T, incubation with TaqI. N1-2, normal tissues; T1-5, tumor tissues. B, bisulfite sequencing analysis of normal and tumor tissue and celllines to determine themethylation state of CpG nucleotides upstream of themiR-195/497 gene on Chromosome 17. Filled circle, methylated CpG; open circle,unmethylated CpG. C, RT-PCR analysis of miR-195/497 in 2 breast cancer cell lines MCF7 and ZR-75-30 untreated or treated with DNA demethylating agent5-aza-2-deoxycytidine (AZA).
MiR-195 and MiR-497 in Breast Cancer
www.aacrjournals.org Clin Cancer Res; 17(7) April 1, 2011 1725
on May 8, 2018. © 2011 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 24, 2011; DOI: 10.1158/1078-0432.CCR-10-1800

islands in the nucleotide sequence upstream ofmiR-195 andmiR-497 promoters on Chromosome 17, we carried outbisulfite sequencing analysis of genomic DNA isolated fromnormal and tumor tissues, and breast cancer cell lines. Theresults showed densemethylation of certain CpGs located inthe region upstream of miR-195 and miR-497 promoters inthe tumor tissuesandcancer cellsMCF7andZR7530,whereasonly a few scattered CpGs were methylated in the regionupstream of miR-195 and miR-497 promoters from normalbreast tissue (Fig. 2B). To study whether CpG methylationaffectsmiR-195andmiR-497expression, thecancer cellsweretreatedwithDNAdemethylatingagent5-aza-2-deoxycytidine(5-Aza-dC). The cells treated with 5-Aza-dC increased theexpression of both miR-195 and miR-497 (Fig. 2C). Thus,DNA methylation is responsible for the low expression ofmiR-195 and miR-497 in breast cancer cells.
MiR-195 and miR-497 inhibit breast cancer cell colonyformation and invasion in vitro
The breast cancer cells were transiently transfected withmiR-195, miR-497 or negative control precursors to study
their effects in colony formation. Overexpression of miR-195 and miR-497 in both ZR-75-30 and MCF7 cells exhib-ited a significant reduction of colony number when com-pared with the cells transfected with the negative control(Fig. 3A). To analyze the role of miR-195 in cell invasion,ZR-75-30 andMCF7 cells were transfected with miR-195 ormiR-497, and analyzed by Transwell assay. MiR-195 ormiR-497 transfection significantly inhibited cell invasionwhen compared with the control (Fig. 3B). To understandthe potential mechanism of miR-195, miR-497 in inhibit-ing these biological effects, we found that miR-195 andmiR-497 overexpression induced cell cycle G1 arrest48 hours after transfection (Fig. 3C), and 24 and 36 hoursafter transfection (Supplementary Fig. S1).
MiR-195 and miR-497 directly target Raf-1To further understandmolecular mechanism ofmiR-195
and miR-497 in inhibiting breast cancer cells, we searchedfor potential targets of miR-195 andmiR-497 by the KeyTarmiRNA target prediction algorithm that is based on se-quences and real physiological situation. This algorithm
Figure 3. miR-195/497 inhibitbreast cancer cell growth andinvasion in vitro. A, Colonyformation in soft agar ofMCF7 andZR-75-30 cells transfected withmiR-195, miR-497 or the negativecontrol oligo (a mismatchedmiRNA oligo, mir-mm). Colonies >50 mm in diameter were countedunder a microscopic field at 10xmagnifications. Values (mean �SD, n ¼ 3) are shown. *, P < 0.05;**, P < 0.01 (t-test). B, Matrigelinvasion assay of MCF7 and ZR-75-30 cells that were transfectedwith control (upper) or miR-195/497 oligo (lower; �200).A significant decrease wasobserved in miR-195- or miR-497-transfected breast cells.*, P < 0.05. C, Cell cycle analysisby FACS in MCF7 and ZR-75-30cells transfected with miR-195 ormiR-497. Mean � SD wereshown. *, P < 0.01 (t-test).
Li et al.
Clin Cancer Res; 17(7) April 1, 2011 Clinical Cancer Research1726
on May 8, 2018. © 2011 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 24, 2011; DOI: 10.1158/1078-0432.CCR-10-1800

suggested that Raf-1 is a novel direct target of both miR195and miR-497 (Fig. 4B). The potential sites of miR195 andmiR-497 in Raf-1 3’UTR were even more conserved thanthose in Ccnd1 3’UTR, a known target of miR-195 andmiR-497. Luciferase assays showed that miR-195 and miR-497 significantly reduced the activity of the Raf-1 genereporter, but not of the mutated gene reporter (Fig. 4C).Western blot analysis of Raf-1 and Ccnd1 in ZR-75-30 andMCF7 cell lines showed that miR-195 or miR-497 transfec-tion inhibited Raf-1 and Ccnd1 expression and ERK1/2phosphorylation (Fig. 4D and E). To confirm the datafurther, we subsequently established stable-expression ofmiR-195 and miR-497 breast cancer cells by lentivirustransfection (Fig. 4F). Similarly, we found that miR-195and miR-497 inhibited cell growth and survival (data notshown), and led to significant downregulation of both Raf-
1 and Ccnd1 in vitro (Fig. 4G). The results further demon-strate that Raf-1 is a novel target of miR-195 and miR-497in different experimental systems.
MiR-195 and miR-497 levels correlate inversely withmalignancy of human breast tumors
We also investigated the relationship between the expres-sion levels of miR-195 and miR-497 and the malignantstages of human breast tumors. As shown in Figure 5A,miR-195 and miR-497 levels were significantly downregu-lated in malignant tumors (n ¼ 71, Stage I-III) whencompared with those in the normal tissues (P < 0.0001),and to those in the benign tumors, known as fibroadnoma(P < 0.05). No significant difference existed between nor-mal and benign tumor. Such data suggested that levelsof miR-195 and miR-497 are inversely associated with
Figure 4. Raf1 and Ccnd1 are direct targets of miR-195/497. A, Structure of human miR-195/497 locus. B, Schematic diagram of putative miR-195/497binding sites in the 30UTR of Raf-1 and Ccnd1. The possible binding sites of miR-195/497 in the 30UTR or CDS of the target genes were searchedusing MiRanda v1.0b (36). The conservation of the corresponding sites in the genomes of 28 species was analyzed using UCSC Genome Browser (NCBI36/hg18, http://genome.ucsc.edu/). The mutated nucleotides are indicated in red box. The sites for Raf1 were even more conserved thanthose for Ccnd1, a previously reported target of miR-195/497. C, Luciferase assays of Raf-1 and Ccnd1 in miR-195/497-overexpressed breast cells.Compared with negative control, the overexpression of both miR-195 and miR-497 could inhibit luciferase reporter activities of Raf-1 and Ccnd1. n ¼ 3; *,**,P < 0.05. D, Western blot analysis of Raf-1 and Ccnd1 levels in the cells transfected with miR-195/497 precursors. Significant decrease levels of Raf-1 andCcnd1were observed after the forced expression of miR-195/497. The expression pattern of Raf1 was similar to that of Ccnd1. E, Western blot analysis of Raf-1 and its downstream genes in miR-195/497 overexpressed MCF-7 cells. MiR-195/497 repressed the expression levels of Raf-1 and ERK1/2, and inhibitedthe phosphorylation of Erk1/2. F, Expression ofmiR-195/497 in lentivirus-transfectedMDA231 cells. Higher miR-195 andmiR-497 expression inmiR-195/497-transfected cells was detected by semi-quantitative RT-PCR. G, Western blot analysis of Raf-1 and Ccnd1 levels in MDA231 cells. Raf-1 andCcnd1 levels were greatly suppressed in miR-195- and mir-497-expressingMDA231 cells when compared with those in the cells expressing precursor control(scr-mda231). Beta-actin was used as an internal control.
MiR-195 and MiR-497 in Breast Cancer
www.aacrjournals.org Clin Cancer Res; 17(7) April 1, 2011 1727
on May 8, 2018. © 2011 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 24, 2011; DOI: 10.1158/1078-0432.CCR-10-1800

malignancy of human breast cancer. But there were nosignificant differences between each stage of breast cancer(P > 0.05), indicating that miR-195 andmiR-497 could notbe used in staging of malignant breast tumors. In addition,malignant breast tumors showed much higher expressionlevels of Raf-1 and Erk1/2 when compared with the normaltissues (Fig. 5B). Thus, the expression levels of Raf-1 areinversely correlated with expression levels of miR-195and miR-497 in human cancer tissues. In addition, weanalyzed the association between miR-195/497 and hor-mone receptors. No significant difference of miR-195/497expression was found between estrogen receptor (ER)positive (n ¼ 45) versus ER negative (n ¼ 26) breastcancers. There was no significant difference of the 2 miR-NAs between progesterone receptor (PR) positive and PRnegative tumors (n ¼ 40 vs. 31, P > 0.05).
Discussion
In this study, miR-195 and miR-497 were found to besignificantly downregulated in breast cancer tissues and celllines. MiR-195 was initially reported to be upregulated in
cardiac hypertrophy, and its overexpression led to patho-logical heart growth and heart failure in transgenic mice(22). Recently, miR-195 was found to be downregulated ina variety of cancers, including gastric cancer, liver cancer,bladder cancer, and adrenocortical cancer (23–26). Intro-duction of miR-195 markedly suppressed colony forma-tion in vitro and tumor development in nude mice (24).MiR-497 expression was found to be downregulated indifferent cancers, such as primary peritoneal carcinoma,gastric cancer and breast cancer (23, 27). However, the roleof miR-497 in human cancer is not clear yet. Our datasuggested that both miR-195 and miR-497 were signifi-cantly downregulated in breast cancer, and that they wereregulated by a common mechanism of CpG methylationupstream of miR-195 and miR-497 promoters. Recentstudies showed that certain miRNA genes such as miR-1,miR-127, miR-148a, and miR-34b/c contain CpGs that aresusceptible toDNAmethylation for decreasing their expres-sion (18, 20, 21). To understand whether CpGmethylationwas responsible for downregulation of miR-195 and miR-497, we found that DNA demethylating agent treatmentrestored the expression of miR-195 and miR-497 in breastcancer cells, suggesting that DNA methylation in CpGinucleotides upstream of miR-195 and miR-497 promotershas a functional role to decrease their expression. Otherthan DNA methylation, p53 inactivation or genetic dele-tion could lead to miRNA deregulation (28, 29). In chroniclymphocytic leukemia patients, chromosome 17p deletionmay influence a set of miRNAs, such as miR-21 and miR-34a (30, 31).
In addition, forced expression of miR-195 and miR-497had similar effect to suppress breast cancer cell proliferation,blocked cell cycle G1 progression, and induced apoptosis.Such results suggest thatmiR-195 andmiR-497have similareffects to play a tumor-suppressor role in breast cancer bythe same cluster of gene regulation. Furthermore, we pre-dicted and confirmed that Raf-1 is a novel target of bothmiR-195 and miR-497. Raf-1 has been found to be over-expressed or overactivated in a variety of cancers, includingrenal cell carcinoma, hepatocellular carcinoma, non–smallcell lung cancer, melanoma, and papillary thyroid carci-noma (32). Recently, Raf-1was found tobe a target ofmiR-7and miR-125b (33, 34). In breast cancer, miR-125b couldtarget the same regionof c-Raf-1 (34). In addition, Raf-1wasidentified as a direct target of miR-7 in several cancer celllines, including breast cancer (33). Here, we present the firstreport that Raf-1 is a direct target of bothmiR-195 andmiR-497 in breast cancer, extending the potential target networkofmiRNAs. It would be interesting to testwhether this novelregulation mechanism of miR-195 and miR-497 in breastcancer, and their common target of Raf-1 may also beextended to other human cancers.
Finally, our data showed that expression levels of miR-195 and miR-497 correlated inversely with malignancy ofhuman breast tumors. Such expression pattern couldpotentially identify malignant tumors from normal orbenign tumors. Recent finding in miRNA study suggestssome miRNAs to be used for diagnosis or prognosis of
Figure 5.MiR-195/497 levels correlate inversely withmalignancy in humanbreast tumors. A, miR-195/497 levels in primary breast tumor tissues.Normal: tissues from nondiseased individuals (n ¼ 25); breastfibroadenoma (n ¼ 9) and invasive ductal carcinoma (total: 71, includingStage I: 22; Stage II: 23; and Stage III: 26) were employed. U6 rRNA wasused as a loading control. No significant difference existed betweennormal and benign tumor tissues. Mir-195/497 levels were significantlydownregulated in each stage of malignant tumors (Stages I–III) ascompared with normal (P < 0.0001) or benign tumor tissues (P < 0.05).B, Western blot analysis for Raf-1 and its downstream proteins in breastcancer tissues. Horizontal bars denote median. Malignant breast tumorshad higher expression of Raf-1 (6/6), Erk1/2 (6/6) and phospho-Erk1/2(3/6), as compared with those of normal tissue.
Li et al.
Clin Cancer Res; 17(7) April 1, 2011 Clinical Cancer Research1728
on May 8, 2018. © 2011 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 24, 2011; DOI: 10.1158/1078-0432.CCR-10-1800

breast cancer in the future. MiR-21 was significantly corre-lated with advanced clinical stage, lymph node metastasis,and poor survival of the patients (35). As miRNAs haveunique expression profile in cancer tissues, and theirexpression is more stable than mRNA. miRNAs are promis-ing for application as biomarkers. Future work shouldfocus on the evaluation of the diagnostic or prognosticvalue of miR-195/497. In summary, the results suggest thatboth miR-195 and miR-497 are downregulated in breastcancer, and their silence is caused by DNA methylation.Our data imply that miR-195/497 may be potential ther-apeutic targets in breast cancer therapy.
Disclosure of Potential Conflicts of Interest
No potential conflicts of interest were disclosed.
Acknowledgments
We thank Chuan-Xiu Bian, Xiaonan Fu, Daoyuan Dong, Zhongshi Liang,Yun Liu, and Ying Wu for some technical assistance.
Grant Support
This work was supported in part by the National Basic Research Programof China (2007CB947001, 2007CB947002), by State High TechnologyDeveloping Project (2008AA02Z115), by the Key Programs of NationalScience Foundation of China (30430240), by Shanghai Metropolitan Fundfor Research and Development (04DZ14005 and 04JC14096), and byNational Cancer Institute, NIH Grant R01CA109460.
The costs of publication of this article were defrayed in part by thepayment of page charges. This article must therefore be hereby markedadvertisement in accordance with 18 U.S.C. Section 1734 solely to indicatethis fact.
Received July 7, 2010; revised January 31, 2011; accepted February 6,2011; published OnlineFirst February 24, 2011.
References1. He L, Hannon GJ. MicroRNAs: Small RNAs with a big role in gene
regulation (vol 5, pg 522 2004). Nat Rev Genet 2004;5: 522-þ.2. Vasudevan S, Tong YC, Steitz JA. Switching from repression to
activation: MicroRNAs can up-regulate translation. Science 2007;318:1931–4.
3. Vasudevan S, Tong Y, Steitz JA. Switching from repression to activa-tion: microRNAs can up-regulate translation. Science 2007;318:1931–4.
4. Filipowicz W, Bhattacharyya SN, Sonenberg N. Mechanisms of post-transcriptional regulation by microRNAs: are the answers in sight?NatRev Genet 2008;9:102–14.
5. Calin GA, Croce CM.MicroRNA signatures in human cancers. Nat RevCancer 2006;6:857–66.
6. Iorio MV, Casalini P, Tagliabue E, Menard S, Croce CM. MicroRNAprofiling as a tool to understand prognosis, therapy responseand resistance in breast cancer. Eur J Cancer 2008;44:2753–9.
7. Lu J, Getz G, Miska EA, Alvarez-Saavedra E, Lamb J, Peck D, et al.MicroRNA expression profiles classify human cancers. Nature2005;435:834–8.
8. Iorio MV, Ferracin M, Liu CG, Veronese A, Spizzo R, Sabbioni S, et al.MicroRNA gene expression deregulation in human breast cancer.Cancer Res 2005;65:7065–70.
9. Hossain A, Kuo MT, Saunders GF. Mir-17–5p regulates breast cancercell proliferation by inhibiting translation of AIB1 mRNA. Mol CellularBiol 2006;26:8191–201.
10. Mertens-Talcott SU, Chintharlapalli S, Li X, Safe S. The oncogenicmicroRNA-27a targets genes that regulate specificity protein tran-scription factors and the G2-M checkpoint in MDA-MB-231 breastcancer cells. Cancer Res 2007;67:11001–11.
11. Lehmann U, Hasemeier B, Christgen M, M€uller M, R€omermann D,L€anger F, et al. Epigenetic inactivation of microRNA gene hsa-mir-9–1in human breast cancer. J Pathol 2008;214:17–24.
12. Heneghan HM, Miller N, Lowery AJ, Sweeney KJ, Newell J, Kerin MJ.Circulating microRNAs as novel minimally invasive biomarkers forbreast cancer. Ann Surg 2010;251:499–505.
13. Lehmann U, Streichert T, Otto B, Albat C, Hasemeier B, Christgen H,et al. Identification of differentially expressed microRNAs in humanmale breast cancer. BMC Cancer 2010;10:109.
14. Zhang L, Huang J, Yang N, Greshock J, Megraw MS, Giannakakis A,et al. microRNAs exhibit high frequency genomic alterations in humancancer. Proc Natl Acad Sci U S A 2006;103:9136–41.
15. Baylin SB. DNA methylation and gene silencing in cancer. Nat ClinPract Oncol 2005;2 Suppl 1:S4–11.
16. Goll MG, Bestor TH. Eukaryotic cytosine methyltransferases. Ann RevBiochem 2005;74:481–514.
17. Esteller M. Cancer epigenomics: DNA methylomes and histone-mod-ification maps. Nat Rev Genet 2007;8:286–98.
18. Saito Y, Liang G, Egger G, Friedman JM, Chuang JC, Coetzee GA,et al. Specific activation of microRNA-127 with downregulation of theproto-oncogene BCL6 by chromatin-modifying drugs in human can-cer cells. Cancer Cell 2006;9:435–43.
19. Lujambio A, Ropero S, Ballestar E, FragaMF, Cerrato C, Seti�en F, et al.Genetic unmasking of an epigenetically silenced microRNA in humancancer cells. Cancer Res 2007;67:1424–9.
20. Datta J, Kutay H, Nasser MW, Nuovo GJ, Wang B, Majumder S, et al.Methylation mediated silencing of MicroRNA-1 gene and its role inhepatocellular carcinogenesis. Cancer Res 2008;68:5049–58.
21. Lujambio A, Calin GA, Villanueva A, Ropero S, S�anchez-C�espedes M,Blanco D, et al. A microRNA DNA methylation signature for humancancer metastasis. Proc Natl Acad Sci U S A 2008;105:13556–61.
22. van Rooij E, Sutherland LB, Liu N, Williams AH, McAnally J, GerardRD, et al. A signature pattern of stress-responsivemicroRNAs that canevoke cardiac hypertrophy and heart failure. Proc Natl Acad Sci U S A2006;103:18255–60.
23. Guo JM, Miao Y, Xiao BX, Huan R, Jiang Z, Meng D, et al. Differentialexpression of microRNA species in human gastric cancer versus non-tumorous tissues. J Gastroenterol Hepatol 2009;24:652–7.
24. Xu T, Zhu Y, Xiong Y, Ge YY, Yun JP, Zhuang SM. MicroRNA-195suppresses tumorigenicity and regulates G1/S transition of humanhepatocellular carcinoma cells. Hepatology 2009;50:113–21.
25. Ichimi T, Enokida H, Okuno Y, Kunimoto R, Chiyomaru T, KawamotoK, et al. Identification of novel microRNA targets based on microRNAsignatures in bladder cancer. Int J Cancer 2009;125:345–52.
26. Soon PS, Tacon LJ, Gill AJ, Bambach CP, Sywak MS, Campbell PR,et al. miR-195 and miR-483–5p Identified as Predictors of PoorPrognosis in Adrenocortical Cancer. Clin Cancer Res 2009;15:7684–92.
27. Flavin RJ, Smyth PC, Laios A, O'Toole SA, Barrett C, Finn SP, et al.Potentially important microRNA cluster on chromosome 17p13.1 inprimary peritoneal carcinoma. Mod Pathol 2009;22:197–205.
28. Raver-Shapira N, Marciano E, Meiri E, Spector Y, Rosenfeld N,Moskovits N, et al. Transcriptional activation of miR-34a contributesto p53-mediated apoptosis. Mol Cell 2007;26:731–43.
29. Spizzo R, Nicoloso MS, Lupini L, Lu Y, Fogarty J, Rossi S, et al. miR-145 participates with TP53 in a death-promoting regulatory loop andtargets estrogen receptor-alpha in human breast cancer cells. CellDeath Differentiation;17:246–54.
30. Rossi S, Shimizu M, Barbarotto E, Nicoloso MS, Dimitri F, Sampath D,et al. microRNA fingerprinting of CLL patients with chromosome 17pdeletion identify a miR-21 score that stratifies early survival.Blood;116:945–52.
31. Zenz T, H~abe S, Denzel T, Mohr J, Winkler D, B€uhler A, et al. Detailedanalysis of p53 pathway defects in fludarabine-refractory chroniclymphocytic leukemia (CLL): dissecting the contribution of 17p dele-
MiR-195 and MiR-497 in Breast Cancer
www.aacrjournals.org Clin Cancer Res; 17(7) April 1, 2011 1729
on May 8, 2018. © 2011 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 24, 2011; DOI: 10.1158/1078-0432.CCR-10-1800

tion, TP53 mutation, p53-p21 dysfunction, and miR34a in a prospec-tive clinical trial. Blood 2009;114:2589–97.
32. Gollob JA,Wilhelm S, Carter C, Kelley SL. Role of Raf kinase in cancer:Therapeutic potential of targeting the Raf/MEK/ERK signal transduc-tion pathway. Semin Oncol 2006;33:392–406.
33. Webster RJ, Giles KM, Price KJ, Zhang PM, Mattick JS, LeedmanPJ. Regulation of Epidermal Growth Factor Receptor Signalingin Human Cancer Cells by MicroRNA-7. J Biol Chem 2009;284:5731–41.
34. HofmannMH, Heinrich J, Radziwil G, Moelling K. A Short Hairpin DNAAnalogous to miR-125b Inhibits C-Raf Expression, Proliferation, andSurvival of Breast Cancer Cells. Mol Cancer Res 2009;7:1635–44.
35. Yan LX, Huang XF, Shao Q, Huang MY, Deng L, Wu QL, et al.MicroRNA miR-21 overexpression in human breast cancer is asso-ciated with advanced clinical stage, lymph node metastasis andpatient poor prognosis. RNA 2008;14:2348–60.
36. Enright AJ, John B, Gaul U, Tuschl T, Sander C, Marks DS. MicroRNAtargets in Drosophila. Genome Biol 2003;5:R1.
Li et al.
Clin Cancer Res; 17(7) April 1, 2011 Clinical Cancer Research1730
on May 8, 2018. © 2011 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 24, 2011; DOI: 10.1158/1078-0432.CCR-10-1800

2011;17:1722-1730. Published OnlineFirst February 24, 2011.Clin Cancer Res Dan Li, Yulan Zhao, Changxing Liu, et al. in Breast CancerAnalysis of MiR-195 and MiR-497 Expression, Regulation and Role
Updated version
10.1158/1078-0432.CCR-10-1800doi:
Access the most recent version of this article at:
Material
Supplementary
http://clincancerres.aacrjournals.org/content/suppl/2011/03/30/1078-0432.CCR-10-1800.DC1
Access the most recent supplemental material at:
Cited articles
http://clincancerres.aacrjournals.org/content/17/7/1722.full#ref-list-1
This article cites 33 articles, 15 of which you can access for free at:
Citing articles
http://clincancerres.aacrjournals.org/content/17/7/1722.full#related-urls
This article has been cited by 7 HighWire-hosted articles. Access the articles at:
E-mail alerts related to this article or journal.Sign up to receive free email-alerts
Subscriptions
Reprints and
To order reprints of this article or to subscribe to the journal, contact the AACR Publications Department at
Permissions
Rightslink site. Click on "Request Permissions" which will take you to the Copyright Clearance Center's (CCC)
.http://clincancerres.aacrjournals.org/content/17/7/1722To request permission to re-use all or part of this article, use this link
on May 8, 2018. © 2011 American Association for Cancer Research. clincancerres.aacrjournals.org Downloaded from
Published OnlineFirst February 24, 2011; DOI: 10.1158/1078-0432.CCR-10-1800