an mr 51,000 protein of mammalian spermatogenic cells that is common to the whole xy body and...
TRANSCRIPT

Chromosoma (1995) 103:591-596 C H R O M O S O M A �9 Springer-Verlag 1995
An Mr 51,000 protein of mammalian spermatogenic cells that is common to the whole XY body and centromeric heterochromatin of autosomes
Avril Smith, Ricardo Benavente
Department of Cell and Developmental Biology, Theodor-Boveri-Institute (Biocenter), University of Wtirzburg, Am Hubland, D-97074 Wtirzburg, Germany
Received: 24 December 1994; in revised form: 4 February 1995 / Accepted: 4 February 1995
Abstract. During mammalian male meiotic prophase the sex chromosomes form a structure called the XY body or sex vesicle. This structure is characterized by differ- ential condensation of chromatin and transcriptional in- activity. The reasons and mechanisms for the allocyclic behaviour of sex chromosomes with respect to autoso- rues are largely unknown. In order to gain insight into the process of XY-body formation we are involved in the characterization of proteins associated with meiotic sex chromosomes by immunological approaches. Here we report on the identification of an Mr 51,000 protein (p51) that is homogeneously distributed in the XY body of rodents as shown by immunocytochemistry with the novel monoclonal antibody 4EC. Interestingly, in germ line cells the antibody also labelled the centromeric het- erochromatin of autosomes. We speculate that p51 may be a component of the mechanisms that lead to wide chromosome regions becoming inaccessible for tran- scription and/or recombination events.
Introduction
During meiotic prophase of spermatogenesis the sex chromosomes X and Y of mammals form a conspicuous structure termed the XY body or sex vesicle, whereas the two X chromosomes of mammalian females behave during meiotic prophase in a manner indistinguishable from autosomes (Solari 1974). The XY body possesses several interesting features that are different from the sit- uation of autosomes: (a) the chromatin of sex chromo- somes shows clear differences in condensation, i.e. it shows allocyclic behaviour. (b) Recombination is re- stricted to the so-called pseudoautosomal region. (c) The XY body is largely transcriptionally inactive (Solari 1989). Thus, the XY body appears as an interesting model system for investigating the mechanisms that con-
Edited by: W. Hennig Correspondence to: R. Benavente
trol the structure and function of whole individual chro- mosomes.
The reasons for the allocyclic behaviour and tran- scriptional inactivity of sex chromosomes during mam- malian male meiotic prophase are poorly understood. In a recent hypothesis put forward by McKee and Han- del (1993) the conspicuous behaviour of sex chromo- somes during male meiosis was related to the recombi- national status of these chromosomes: differential con- densation of sex chromosomes during meiotic prophase would prevent deleterious recombination events in non- homologous regions of the X and Y chromosomes. Ac- cording to McKee and Handel (1993), the transcrip- tional inactivity of the XY body may be viewed as a consequence of chromatin condensation, rather than as a requirement for proper progression of spermatogene- sis as previously postulated (Lifschytz and Lindsley 1972).
The molecular mechanisms controlling XY-body for- mation are not known. Recent genetic investigations have suggested that XY-body formation is not an intrin- sic property of the sex chromosomes, but rather, may be controlled in trans by factors originating from autoso- rues or even by events occurring outside the meiotic cell (Handel et al. 1994). The mechanisms involved in XY- body formation also appear to be different from those controlling X-chromosome inactivation in somatic cells of female mammals. In contrast to the situation in fe- male tissues, the expression of Xist (X-inactive-specific transcript) in mouse testis is very low and does not fit temporally with XY-body formation (Kay etal. 1993; for alternative views see McCarrey and Dilworth 1992; Richler et al. 1992; Salido et al. 1992). Also in contrast to the situation in somatic cells, methylation events do not appear to be involved in sex chromosome inactiva- tion of male meiotic cells (Driscoll and Migeon 1990).
It is conceivable that the mechanisms controlling XY- body formation and maintenance involve nuclear pro- teins that one would expect are spermatogenesis specific and specifically associated with sex chromosomes. In our laboratory we are undertaking efforts to identify
592
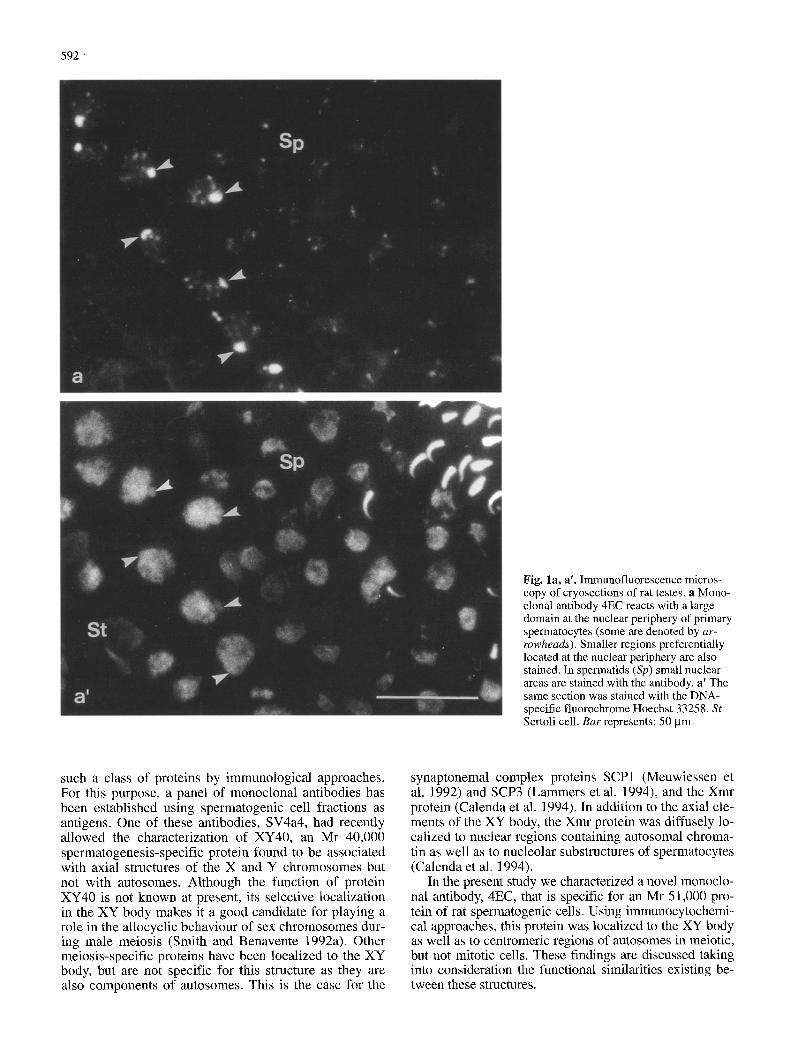
Fig. la, a'. Immunofluorescence micros- copy of cryosections of rat testes, a Mono- clonal antibody 4EC reacts with a large domain at the nuclear periphery of primary spermatocytes (some are denoted by ar- rowheads). Smaller regions preferentially located at the nuclear periphery are also stained. In spermatids (Sp) small nuclear areas are stained with the antibody, a' The same section was stained with the DNA- specific tluorochrome Hoechst 33258. St Sertoli cell. Bar represents: 50 gm
such a class of proteins by immunological approaches. For this purpose, a panel of monoclonal antibodies has been established using spermatogenic cell fractions as antigens. One of these antibodies, SV4a4, had recently allowed the characterization of XY40, an Mr 40,000 spermatogenesis-specific protein found to be associated with axial structures of the X and Y chromosomes but not with autosomes. Although the function of protein XY40 is not known at present, its selective localization in the XY body makes it a good candidate for playing a role in the allocyclic behaviour of sex chromosomes dur- ing male meiosis (Smith and Benavente 1992a). Other meiosis-specific proteins have been localized to the XY body, but are not specific for this structure as they are also components of autosomes. This is the case for the
synaptonemal complex proteins SCP1 (Meuwiessen et al. 1992) and SCP3 (Lammers et al. 1994), and the Xmr protein (Calenda et al. 1994). In addition to the axial ele- ments of the XY body, the Xmr protein was diffusely lo- calized to nuclear regions containing autosomal chroma- tin as well as to nucleolar substructures of spermatocytes (Calenda et al. 1994).
In the present study we characterized a novel monoclo- nal antibody, 4EC, that is specific for an Mr 51,000 pro- tein of rat spermatogenic cells. Using immunocytochemi- cal approaches, this protein was localized to the XY body as well as to centromeric regions of autosomes in meiotic, but not mitotic cells. These findings are discussed taking into consideration the functional similarities existing be- tween these structures.
593
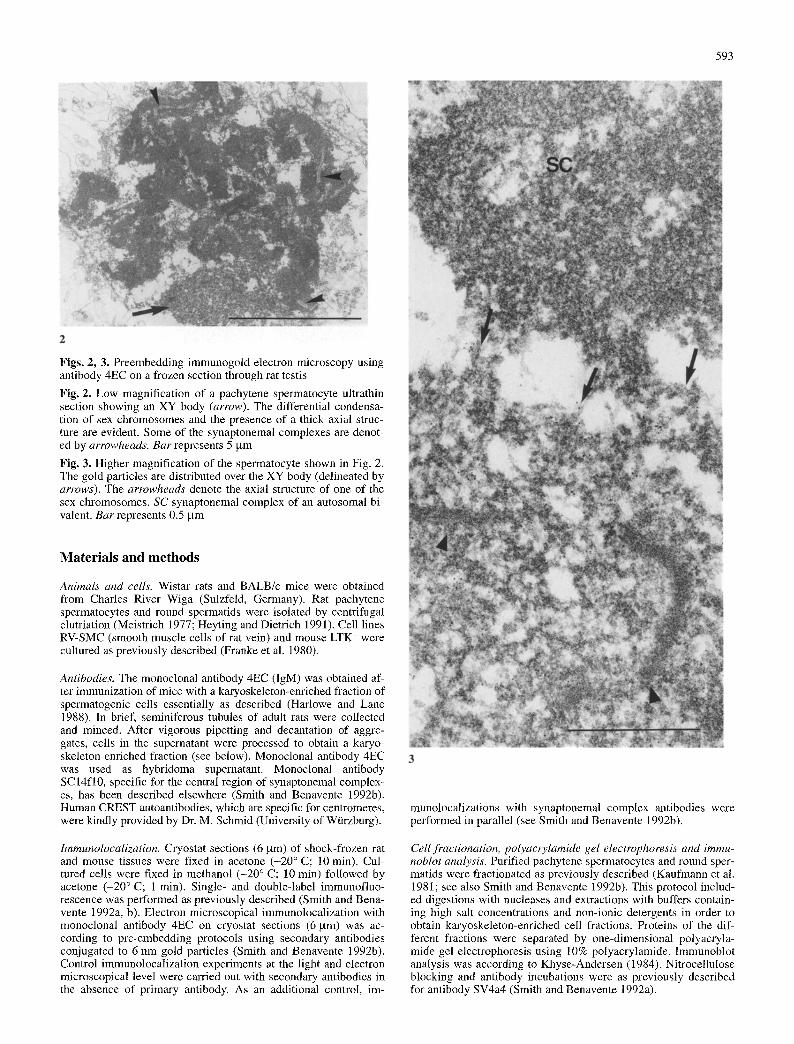
Figs. 2, 3. Preembedding immunogold electron microscopy using antibody 4EC on a frozen section through rat testis
Fig. 2. Low magnification of a pachytene spermatocyte ultrathin section showing an XY body (arrow). The differential condensa- tion of sex chromosomes and the presence of a thick axial struc- ture are evident. Some of the synaptonemal complexes are denot- ed by arrowheads. Bar represents 5 ~tm
Fig. 3. Higher magnification of the spermatocyte shown in Fig. 2. The gold particles are distributed over the XY body (delineated by arrows). The arrowheads denote the axial structure of one of the sex chromosomes. SC synaptonemal complex of an autosomal bi- valent. Bar represents 0.5 gm
Materials and methods
Animals and cells. Wistar rats and BALB/c mice were obtained from Charles River Wiga (Sulzfeld, Germany). Rat pachytene spermatocytes and round spermatids were isolated by centrifugal elutriation (Meistrich 1977; Heyting and Dietrich 1991). Cell lines RV-SMC (smooth muscle cells of rat vein) and mouse LTK- were cultured as previously described (Franke et al. 1980).
Antibodies. The monoclonal antibody 4EC (IgM) was obtained af- ter immunization of mice with a karyoskeleton-enriched fraction of spennatogenic cells essentially as described (Harlowe and Lane 1988). In brief, serniniferous tubules of adult rats were collected and minced. After vigorous pipetting and decantation of aggre- gates, cells in the supernatant were processed to obtain a karyo- skeleton-enriched fraction (see below). Monoclonal antibody 4EC was used as hybridoma supernatant. Monoclonal antibody SC14fl0, specific for the central region of synaptonemal complex- es, has been described elsewhere (Smith and Benavente 1992b). Human CREST autoantibodies, which are specific for centromeres, were kindly provided by Dr. M. Schmid (University of Wtirzburg).
Immunolocalization. Cryostat sections (6 gm) of shock-frozen rat and mouse tissues were fixed in acetone (-20 ~ C; 10 min). Cul- tured cells were fixed in methanol (-20 ~ C; 10 min) followed by acetone (-20 ~ C; 1 min). Single- and double-label immunofluo- rescence was performed as previously described (Smith and Bena- vente 1992a, b). Electron microscopical immunolocalization with monoclonal antibody 4EC on cryostat sections (6 gin) was ac- cording to pre-embedding protocols using secondary antibodies conjugated to 6 nm gold particles (Smith and Benavente 1992b). Control immunolocalization experiments at the light and electron microscopical level were carried out with secondary antibodies in the absence of primary antibody. As an additional control, ira-
munolocalizations with synaptonemal complex antibodies were performed in parallel (see Smith and Benavente 1992b).
Cell fractionation, polyacrylamide gel electrophoresis and immu- noblot analysis. Purified pachytene spermatocytes and round sper- matids were fractionated as previously described (Kaufmann et al. 1981; see also Smith and Benavente 1992b). This protocol includ- ed digestions with nucleases and extractions with buffers contain- ing high salt concentrations and non-ionic detergents in order to obtain karyoskeleton-enriched cell fractions. Proteins of the dif- ferent fractions were separated by one-dimensional polyacryla- mide gel electrophoresis using 10% polyacrylamide. Immunoblot analysis was according to Khyse-Andersen (1984). Nitrocellulose blocking and antibody incubations were as previously described for antibody SV4a4 (Smith and Benavente 1992a).
594
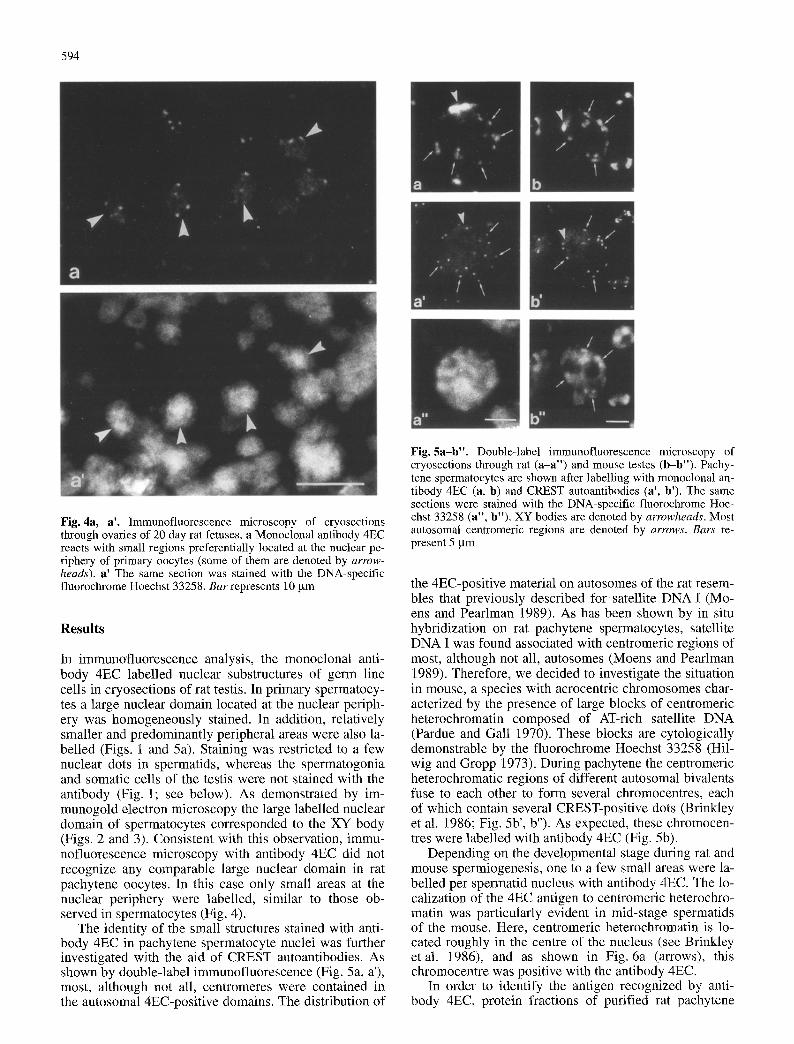
Fig. 4a, a'. Immunofluorescence microscopy of cryosections through ovaries of 20 day rat fetuses, a Monoclonal antibody 4EC reacts with small regions preferentially located at the nuclear pe- riphery of primary oocytes (some of them are denoted by arrow- heads) , a' The same section was stained with the DNA-specific fluorochrome Hoechst 33258. B ar represents 10 gm
Results
In immunofluorescence analysis, the monoclonal anti- body 4EC labelled nuclear substructures of germ line cells in cryosections of rat testis. In primary spermatocy- tes a large nuclear domain located at the nuclear periph- ery was homogeneously stained. In addition, relatively smaller and predominantly peripheral areas were also la- belled (Figs. 1 and 5a). Staining was restricted to a few nuclear dots in spermatids, whereas the spermatogonia and somatic cells of the testis were not stained with the antibody (Fig. 1; see below). As demonstrated by im- munogold electron microscopy the large labelled nuclear domain of spermatocytes corresponded to the XY body (Figs. 2 and 3). Consistent with this observation, immu- nofluorescence microscopy with antibody 4EC did not recognize any comparable large nuclear domain in rat pachytene oocytes. In this case only small areas at the nuclear periphery were labelled, similar to those ob- served in spermatocytes (Fig. 4).
The identity of the small structures stained with anti- body 4EC in pachytene spermatocyte nuclei was further investigated with the aid of CREST autoantibodies. As shown by double-label immunofluorescence (Fig. 5a, a'), most, although not all, centromeres were contained in the autosomal 4EC-positive domains. The distribution of
Fig. 5a-b". Double-label immunofluorescence microscopy of cryosections through rat (a-a") and mouse testes (b-b"). Pachy- tene spermatocytes are shown after labelling with monoclonal an- tibody 4EC (a, b) and CREST autoantibodies (a', b'). The same sections were stained with the DNA-specific fluorochrome Hoe- chst 33258 (a", b"). XY bodies are denoted by arrowheads . Most autosomal centromeric regions are denoted by arrows. Bars re- present 5 ~tm
the 4EC-positive material on autosomes of the rat resem- bles that previously described fo r satellite DNA I (Mo- ens and Pearlman 1989). As has been shown by in situ hybridization on rat pachytene spermatocytes, satellite DNA I was found associated with centromeric regions of most, although not all, autosomes (Moens and Pearlman 1989). Therefore, we decided to investigate the situation in mouse, a species with acrocentric chromosomes char- acterized by the presence of large blocks of centromeric heterochromatin composed of AT-rich satellite DNA (Pardue and Gall 1970). These blocks are cytologically demonstrable by the fluorochrome Hoechst 33258 (Hil- wig and Gropp 1973). During pachytene the centromeric heterochromatic regions of different autosomal bivalents fuse to each other to form several chromocentres, each of which contain several CREST-positive dots (Brinkley et al. 1986; Fig. 5b', b"). As expected, these chromocen- tres were labelled with antibody 4EC (Fig. 5b).
Depending on the developmental stage during rat and mouse spermiogenesis, one to a few small areas were la- belled per spermatid nucleus with antibody 4EC. The lo- calization of the 4EC antigen to centromeric heterochro- matin was particularly evident in mid-stage spermatids of the mouse. Here, centromeric heterochromatin is lo- cated roughly in the centre of the nucleus (see Brinkley et al. 1986), and as shown in Fig. 6a (arrows), this chromocentre was positive with the antibody 4EC.
In order to identify the antigen recognized by anti- body 4EC, protein fractions of purified rat pachytene
595
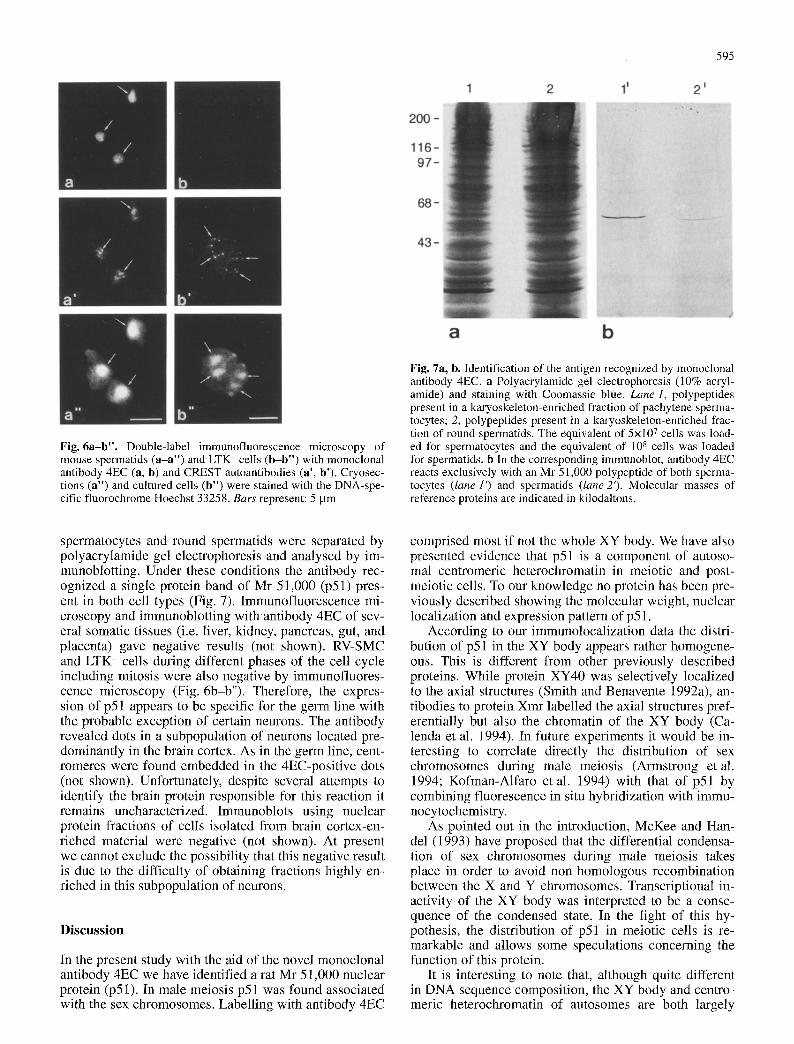
Fig. 6a-b". Double-label immunofluorescence microscopy of mouse spermatids (a-a") and LTK- cells (b-b") with monoclonal antibody 4EC (a, b) and CREST autoantibodies (a', b'). Cryosec- tions (a") and cultured cells (b") were stained with the DNA-spe- cific fluorochrome Hoechst 33258. Bars represent: 5 gm
Fig. 7a, b. Identification of the antigen recognized by monoclonal antibody 4EC. a Polyacrylamide gel electrophoresis (10% acryl- amide) and staining with Coomassie blue. Lane 1, polypeptides present in a karyoskeleton-enriched fraction of pachytene sperma- tocytes; 2, polypeptides present in a karyoskeleton-enriched frac- tion of round spermatids. The equivalent of 5x107 ceils was load- ed for spermatocytes and the equivalent of 108 cells was loaded for spermatids, b in the corresponding immunoblot, antibody 4EC reacts exclusively with an Mr 51,000 polypeptide of both sperma- tocytes (lane 11 and spermatids (lane2'). Molecular masses of reference proteins are indicated in kilodaltons.
spermatocytes and round spermatids were separated by polyacrylamide gel electrophoresis and analysed by im- munoblotting. Under these conditions the antibody rec- ognized a single protein band of Mr 51,000 (p51) pres- ent in both cell types (Fig. 7). Immunofluorescence mi- croscopy and immunoblotting with antibody 4EC of sev- eral somatic tissues (i.e. liver, kidney, pancreas, gut, and placenta) gave negative results (not shown). RV-SMC and LTK- cells during different phases of the cell cycle including mitosis were also negative by immunofluores- cence microscopy (Fig. 6b-b"). Therefore, the expres- sion of p51 appears to be specific for the germ line with the probable exception of certain neurons. The antibody revealed dots in a subpopulation of neurons located pre- dominantly in the brain cortex. As in the germ line, cent- romeres were found embedded in the 4EC-positive dots (not shown). Unfortunately, despite several attempts to identify the brain protein responsible for this reaction it remains uncharacterized. Immunoblots using nuclear protein fractions of cells isolated from brain cortex-en- riched material were negative (not shown). At present we cannot exclude the possibility that this negative result is due to the difficulty of obtaining fractions highly en- riched in this subpopulation of neurons.
Discussion
In the present study with the aid of the novel monoclonal antibody 4EC we have identified a rat Mr 51,000 nuclear protein (p51). In male meiosis p51 was found associated with the sex chromosomes. Labelling with antibody 4EC
comprised most if not the whole XY body. We have also presented evidence that p51 is a component of autoso- mal centromeric heterochromatin in meiotic and post- meiotic cells. To our knowledge no protein has been pre- viously described showing the molecular weight, nuclear localization and expression pattern of p51.
According to our immunolocalization data the distri- bution of p51 in the XY body appears rather homogene- ous. This is different from other previously described proteins. While protein XY40 was selectively localized to the axial structures (Smith and Benavente 1992a), an- tibodies to protein Xmr labelled the axial structures pref- erentially but also the chromatin of the XY body (Ca- lenda et al. 1994). In future experiments it would be in- teresting to correlate directly the distribution of sex chromosomes during male meiosis (Armstrong et al. 1994; Kofman-Alfaro et al. 1994) with that of p51 by combining fluorescence in situ hybridization with immu- nocytochemistry.
As pointed out in the introduction, McKee and Han- del (1993) have proposed that the differential condensa- tion of sex chromosomes during male meiosis takes place in order to avoid non-homologous recombination between the X and Y chromosomes. Transcriptional in- activity of the XY body was interpreted to be a conse- quence of the condensed state. In the light of this hy- pothesis, the distribution of p51 in meiotic cells is re- markable and allows some speculations concerning the function of this protein.
It is interesting to note that, although quite different in DNA sequence composition, the XY body and centro- meric heterochromatin of autosomes are both largely

596
transcriptionally inactive. Furthermore, it is known that recombinat ion in heterochromatic regions is rare during meiosis. This is also true for the sex chromosomes in the heterogametic sex o f mammals , in which recombinat ion is restricted to a small domain o f the pseudoautosomal region (Solari 1989). On the other hand, proteins in- volved in long term supression of transcription have been previously described. Interestingly, the two best known examples in mammals , i.e. the ch romobox pro- tein M31 (a homologue of Drosophila HP1; Eissenberg et al. 1990; Wreget t et al. 1994) and the methyl-binding protein MeCP2 (Lewis et al. 1992), were shown to be as- sociated with centromeric heterochromatin. Taking to- gether, it is tempting to speculate that p51, because o f its selective association with the X Y body and centromeric heterochromatin, may be involved in a mechanism ren- dering wide ch romosome regions inaccessible for tran- scription and/or recombinat ion events during gameto- genesis.
Acknowledgements. We thank Ulrich Scheer for generous support, Gustavo Folle, Dagmar Gebauer, Robert Hock, Georg Krohne, David Lourim and Peter Vogt for stimulating discussions, Ute Vinzens for excellent technical assistance and Lynn Leverenz for correcting the manuscript. This work was supported by a grant of the Deutsche Forschungsgemeinschaft to R.B. (Be 1168/1-3).
References
Armstrong SJ, Kirkham AJ, Hulttn MA (1994) XY chromosome behaviour in the germ line of the human male: FISH analysis of spatial orientation, chromatin condensation and paMng. Chromosome Res 2:445-452
Brinkley BR, Brenner SL, Hall JM, Tousson A, Balczon RD, Val- divia MM (1986) Arrangements of kinetochores in mouse cells during meiosis and spermiogenesis. Chromosoma 94:309-317
Calenda A, Allenet B, Escalier D, Bach JF, Garchon HJ (1994) The meiosis-specific Xmr gene product is homologous to the lymphocyte XIr protein and is a component of the XY body. EMBO J 13:100-109
Driscoll DJ, Migeon BR (1990) Sex differences in methylation of single-copy genes in human meiotic germ cells: Implica- tion for X chromosome inactivation, parental imprinting, and origin of CpG mutations. Somatic Cell Mol Genet 16:267- 282
Eissenberg JC, James TC, Foster-Hartnett DM, Ngan Y, Elgin SCR (1990) Mutation in a heterochromatin-specific chromo- somal protein is associated with supression of position-effect variegation in Drosophila melanogaster. Proc Natl Acad Sci USA 87:9923-9927
Franke WW, Schmid E, Vandekerkhove J, Weber K (1980) A per- manently proliferating rat vascular smooth muscle cell with maintained expression of smooth muscle cell characteristics, including actin of the vascular smooth muscle type. J Cell Biol 87:594-600
Handel MA, Park C, Kot M (1994) Genetic control of sex-chro- mosome inactivation during male meiosis. Cytogenet Cell Genet 66:83-88
Harlowe E, Lane D (1988) Antibodies. A laboratory manual. Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY
Heyting C, Dietrich AJJ (1991) Meiotic chromosome preparation and protein labeling. Methods Cell Biol 35:177-202
Hilwig I, Gropp A (1973) Decondensation of constitutive hetero- chromatin in L cell chromosomes by a benzimidazole com- pound ("33258 Hoechst"). Exp Cell Res 81:474-477
Kaufmaun SH, Coffey DS, Shaper JH (1981) Considerations in the isolation of rat liver nuclear matrix, nuclear envelope, and pore complex-lamina. Exp Cell Res 132:105-123
Kay GF, Penny GD, Patei D, Ashworth A, Brockdorff N, Rastan S (1993) Expression of Xist during mouse development suggests a role in the initiation of X chromosome inactivation. Cell 72:171-182
Khyse-Andersen J (1984) Electroblotting of multiple gels: a sim- ple apparatus without buffer tank for rapid transfer of proteins from polyacrylamide to nitrocellulose. J Biochem Biophys Methods 10:203-209
Kofman-Alfaro S, Speed RM, Boyle S, Chandley AC (1994) Con- densation of the human X chromosome in male germ cells and Sertoli cells examined by fluorescence in situ hybridization. Chromosome Res 2:439-444
Lammers JHM, Offenberg HH, van Aalderen M, Vink ACG, Die- trich AJJ, Heyting C (1994) The gene encoding a major com- ponent of lateral elements of synaptonemal complexes of the rat is related to X-linked lymphocyte-regulated genes. Mol Cell Biol 14:1137-1146
Lewis JD, Meehan RR, Henzel WJ, Maurer-Fogy I, Jeppesen P, Klein F, Bird A (1992) Purification, sequence, and cellular lo- calization of a novel chromosomal protein that binds to meth- ylated DNA. Cell 69:905-914
Lifschytz E, Lindsley DL (1972) The role of X-chromosome inac- tivation during spermatogenesis. Proc Natl Acad Sci USA 69:182-186
McCarrey JR, Dilworth DD (1992) Expression of Xist in mouse germ cells correlates with X-chromosome inactivation. Nature Genet 2:200-203
McKee BD, Handel MA (1993) Sex chromosomes, recombina- tion, and chromatin conformation. Chromosoma 102:71-80
Meistrich ML (1977) Separation of spermatogenic cells and nu- clei of rodent testes. Methods Cell Biol 15:15-54
Meuwiessen RL, Offenberg HI-I, Dietrich AJJ, Riesewijk A, van Iersel M, Heyting C (1992) A coiled-coil related protein spe- cific for synapsed regions of meiotic prophase chromosomes. EMBO J 11:5091-5100
Moens PB, Pearlman RE (1989) Satellite DNA I in chromosomal loops of rat pachytene chromosomes and spermatids. Chromo- soma 98:287-294
Pardue ML, Gall JG (1970) Chromosomal localization of mouse satellite DNA. Science 168:1356-1358
Richler C, Soreq H, Wahrman J (1992) X inactivation in mamma- lian testis is correlated with inactive X-specific transcription. Nature Genet 2:192-195
Salido EC, Yen PH, Mohandas TK, Shapiro LJ (1992) Expression of X-inactivation-associated gene XIST during spermatogene- sis. Nature Genet 2:196-199
Smith A, Benavente R (1992a) Meiosis-specific protein selective- ly associated with sex chromosomes of rat pachytene sperma- tocytes. Proc Natl Acad Sci USA 98:6938-6942
Smith A, Benavente R (1992b) Identification of a structural pro- tein component of rat synaptonemal complexes. Exp Cell Res 198:291-297
Solari AJ (1974) The behaviour of the XY pair in mammals. Int Rev Cytol 38:273-317
Solari AJ (1989) Sex chromosome pairing and fertility in the heterogametic sex of mammals and birds. In: Gillies CB (ed) Fertility and chromosome pairing: recent studies in plants and animals. CRC, Boca Raton, pp 77-107
Wregett KA, Hill F, James PS, Hutchings A, Butcher GW, Singh PB (1994) A mammalian homologue of Drosophila hetero- chromatin protein 1 (HP1) is a component of constitutive het- erochromatin. Cytogenet Cell Genet 66:99-103