an interplay between two egf-receptor ligands, vein and ... · an interplay between two...
TRANSCRIPT

An interplay between two EGF-receptor ligands, Vein and Spitz, is requiredfor the formation of a subset of muscle precursors inDrosophila
Talia Yarnitzky, Li Min, Talila Volk*
Department of Molecular Genetics, The Weizmann Institute of Science, 76100 Rehovot, Israel
Received 13 August 1998; accepted 21 September 1998
Abstract
Activation of theDrosophilaEGF-receptor (DER) is spatially and temporally controlled by the release of its various ligands. DER and itsligand Spitz mediate the formation of specific somatic muscle precursors. We show that a second DER ligand, Vein, complements theactivity of Spitz in the development of various somatic muscle precursors. Invnmutant embryos, the DER-dependent muscle precursors donot form in some of the segments. This phenotype is significantly enhanced in embryos carrying only one copy of wild typespitz. Ouranalysis suggests that Vein activation of DER differs qualitatively from that of Spitz in that it does not lead to the expression of theinhibitory protein Argos, possibly leading to a continuous activation of the DER signaling pathway. 1998 Elsevier Science Ireland Ltd.All rights reserved
Keywords:Muscle;Drosophila; Mesoderm; Receptor tyrosine kinase; Egfr/DER; Vein; Spitz; RTK
1. Introduction
Locomotion of theDrosophila larva is achieved throughcoordinated contractions of an array of somatic musclesattached to the larval epidermis (Bate, 1990, 1993). Theidentity of each of the ~30 types of somatic muscle fibersis determined during early embryonic stages from a set ofmuscle founder cells (Rushton et al., 1995). Each foundercell defines a specific muscle fiber identity, characterized bya unique size, position, and orientation for reviews see(Bate, 1993; Abmayr et al., 1995). Muscle differentiationis achieved when myoblasts expressing high Twist levelsfuse to a given founder myoblast and follow its specificpattern of gene expression (Baylies and Bate, 1996). Differ-ent founder myoblasts express a specific repertoire of genescharacteristic of their distinct fate, e.g.S59(Dohrmann etal., 1990),even-skipped(Frasch et al., 1987),vestigial(Wil-liams et al., 1991),apterous(Bourgouin et al., 1992),nau-tilus (Michelson et al., 1990),connectin(Nose et al., 1992),andkruppel (Gaul et al., 1987).
The founder myoblasts are the product of distinctlineages that are initiated by the segregation of a specialclass of muscle progenitors. Each progenitor divides asym-metrically to give rise to two distinct founder myoblasts, ora founder and the precursor of an adult muscle (Carmena etal., 1995; Baylies et al., 1998). The activity ofinscutableandnumbgenes in the progenitor asymmetric cell divisionhas been recently described (Ruiz Gomez and Bate, 1997;Carmena et al., 1998). Muscle progenitors are selected in aprocess reminiscent of neuroblast segregation which simi-larly requires the neurogenic genes (Hartenstein et al.,1992). A single progenitor is selected from a cluster ofcells expressinglethal of scute(l’scute) to retain high levelsof L’scute (Ruiz Gomez et al., 1997) and in addition itexpresses a particular subset of identity genes (mentionedabove). Specification of the progenitor cell is influenced byan extrinsic mechanism involving intercellular signalingmolecules such as those encoded by thewingless(wg) anddecapentaplegic(dpp) genes (Staehling-Hampton et al.,1994; Baylies et al., 1995; Frasch, 1995; Lawrence et al.,1995; Park et al., 1996; Ranganayakulu et al., 1996).
Another mechanism essential for the specification of asubset of muscle progenitors is theDrosophilaEGF-recep-tor pathway (Buff et al., 1998). In the absence of the recep-
Mechanisms of Development 79 (1998) 73–82
0925-4773/98/$ - see front matter 1998 Elsevier Science Ireland Ltd. All rights reservedPII S0925-4773(98)00175-0
* Corresponding author. Tel.: +972-8-934-2426; fax: +972-8-934-4108;e-mail: [email protected]

tor (DER/Egfr), or its ligand, Spitz, specific muscle foundercells, including the precursor of DA1 (dorsal acute muscleno. 1), VA2 (ventral acute muscle no. 2) and LL1 (laterallongitudinal muscle no. 1), are missing, leading to a loss ofthese specific muscle fibers. The loss of DA1 muscle foun-der myoblast inDERor spitzmutant embryos results from adefected determination of its specific muscle progenitorP15. In addition, it was suggested that the development ofindividual muscles is differentially sensitive to variations inthe level of signaling by DER (Buff et al., 1998). DER is areceptor tyrosine kinase, essential for the determination of awide array of cell fates during embryonic development (Rayand Schupbach, 1996; Perrimon and Perkins, 1997;Schweitzer and Shilo, 1997). The regulation of the activityof the DER-signaling cascade is thought to be accomplishedby localized release of its ligands, since DER is widelyexpressed in all germ layers throughout embryonic devel-opment. Among DER ligands, Spitz, a TGFa homologue, isa major ligand active in a number of tissues (Rutledge et al.,1992). Spitz is produced as a membrane-bound protein andis processed to produce an extracellular, active form(Schweitzer et al., 1995b). A function for Rhomboid inthe regulation of Spitz processing has been suggested(Golembo et al., 1996a).
Vein, an additional ligand for DER (Schnepp et al., 1996;Yarnitzky et al., 1997), is a secreted growth factor contain-ing Ig-like and EGF-like domains at its C-terminal. Theembryonic expression ofvein mRNA is dynamic andmarks a number of tissues where the Spitz ligand wasreported to be the active ligand. These tissues include theventral ectoderm (Schnepp et al., 1996), tracheal pits(Schnepp et al., 1996), and somatic muscle precursors(Buff et al., 1998; and see also the present work). Despiteits restricted embryonic expression pattern, no obvious phe-notype in these tissues has been reported invein mutantembryos. During late stages of embryonic development,Vein is expressed in all the somatic muscles, and is required,independently of Spitz, for the muscle-dependent differen-tiation of tendon cells (Yarnitzky et al., 1997).
Genetic evidence described in Schnepp et al. (1996) sug-gests a functional link between both DER ligands. Forexample, reducing Vein levels in aspitznull genotype wor-sened the phenotype to produce a collapsed embryo with anextruded head skeleton. In addition, ventral denticles andhead skeleton defects inspitz;vein double mutants weremore severe than the predicted additive phenotype of thesingle mutants. This led to the hypothesis that both ligandsfunction in a synergistic manner in certain tissues. Recently,both Vein and Spitz were found to be required for specifica-tion of neuroblasts along the dorsal-ventral axis during earlyembryonic development (Skeath, 1998).
In the present work, we show that Vein is required tocomplement Spitz in the development of a subset of muscleprecursors whose differentiation depends on DER activity.In veinmutant embryos these muscle precursors do not formin some of the segments. This phenotype is not fully pene-
trant, but is significantly enhanced in embryos carrying onlyone copy of wild typespitz, or in embryos in which theactivity of DER is reduced by expressing a dominant-nega-tive form of the receptor. Ectopic expression of Vein in themesoderm is sufficient to induce supernumerary DER-dependent muscle founders in wild type,spitz, or rhomboidmutant embryos. Unlike Spitz, Vein activation of DER doesnot lead to the expression of the inhibitory protein, Argos.These results led us to suggest a model in which Vein isrequired to complement Spitz activity in the formation ofspecific muscle precursors by inducing a continuous activa-tion of the DER signaling pathway, following the activationof this receptor by Spitz.
2. Results
2.1. Vein contributes to the specification of the DER-dependent muscle precursors
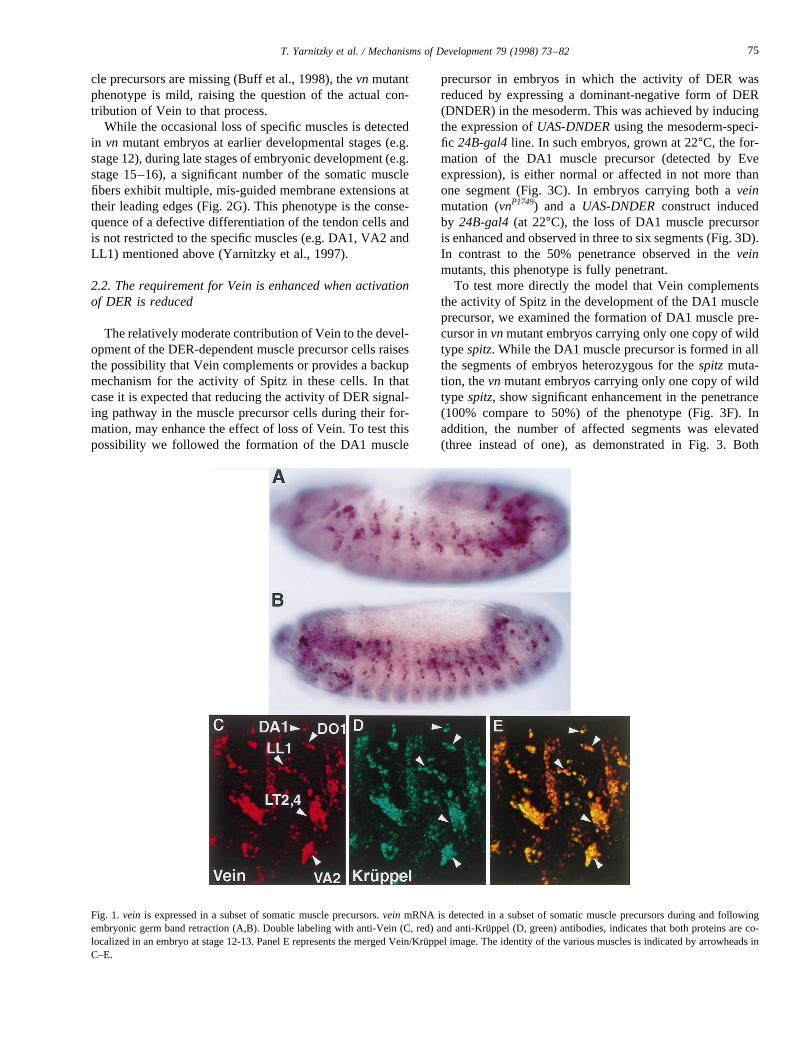
vein mRNA expression is detected in the ventral ecto-derm, somatic mesoderm, tracheal pits and chordotonalorgans (Schnepp et al., 1996). During, and following germband retraction,vein mRNA expression is observed in areiterated segmental pattern in a number of muscle precur-sor cells (Fig. 1A,B). Double labeling with anti-Vein andanti-Kruppel antibodies reveals that the Kru¨ppel-positivecells also express Vein (an overlap between Kru¨ppel andVein staining in various muscle precursors is demonstratedin Fig. 1C–E).
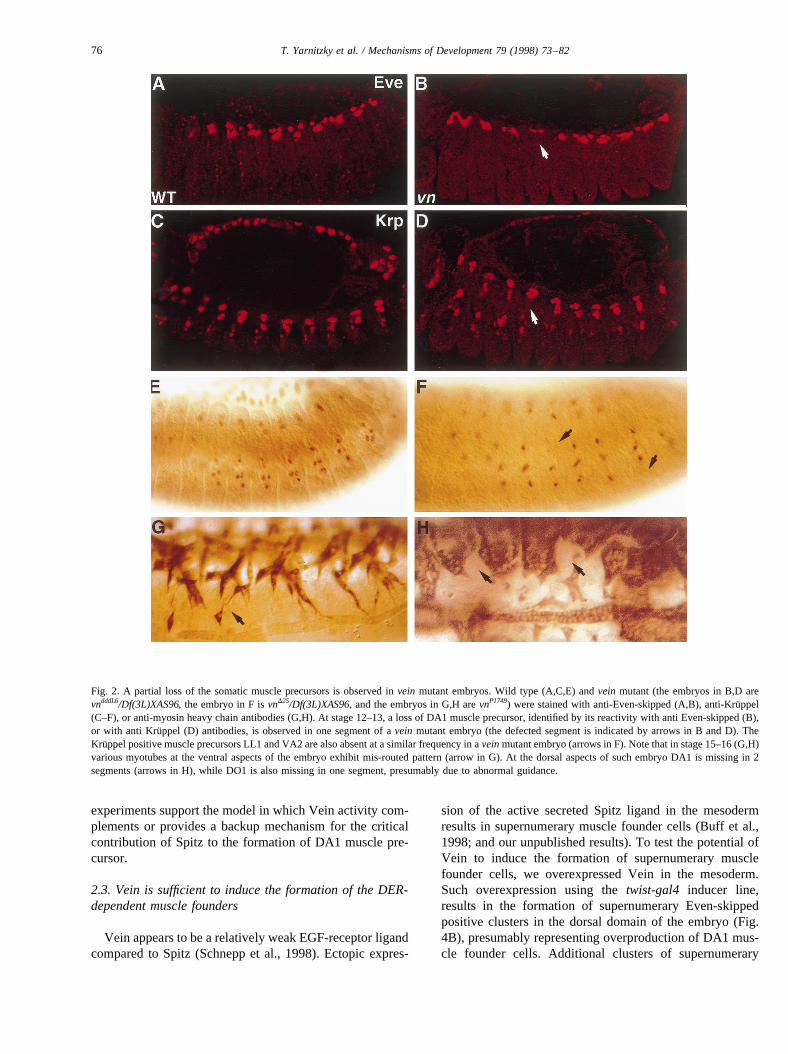
Buff et al. (1998), showed that development of theKruppel-expressing muscle founders DA1, LL1 and VA2,depends on DER activity. In the absence of Spitz, or inembryos expressing dominant negative DER (DNDER),these muscle founders are missing from all the segments.At stages 12–13 of embryonic development, the DA1 mus-cle precursor is identified at the dorsal region of the meso-derm by the common expression of Eve and Kru¨ppel in acluster of several nuclei, representing myoblasts fused to theDA1 founder cell. The precursors of muscles LL1 and VA2express Kru¨ppel, and can be identified by their relative loca-tions in an embryo at stages 12–13. Invein (vn) mutantembryos at stages 12–13 of embryonic development, theDA1 muscle precursor is missing in one (or occasionallytwo) segments of mutant embryos (figurehere>Fig. 2B,D).The penetrance of this phenotype is ~50% (out of 100embryos scored). The Kru¨ppel-positive muscle precursorsLL1 and VA2 are also missing inveinmutant embryos in asimilar frequency as observed for DA1 (Fig. 2F). No altera-tions in the severity of the phenotype were noticed in var-ious vn heteroallelic combinations, includingvnD25/Def(3L)XAS96, or vnP1749/vndddL6. Thus, we concluded thatthe loss of the various muscle precursor cells observed invein mutant embryos is in line with the expression of Veinin these cells. In contrast to the phenotype observed inspitzmutant embryos, in which all the DA1, LL1 and VA2 mus-
74 T. Yarnitzky et al. / Mechanisms of Development 79 (1998) 73–82
cle precursors are missing (Buff et al., 1998), thevn mutantphenotype is mild, raising the question of the actual con-tribution of Vein to that process.
While the occasional loss of specific muscles is detectedin vn mutant embryos at earlier developmental stages (e.g.stage 12), during late stages of embryonic development (e.g.stage 15–16), a significant number of the somatic musclefibers exhibit multiple, mis-guided membrane extensions attheir leading edges (Fig. 2G). This phenotype is the conse-quence of a defective differentiation of the tendon cells andis not restricted to the specific muscles (e.g. DA1, VA2 andLL1) mentioned above (Yarnitzky et al., 1997).
2.2. The requirement for Vein is enhanced when activationof DER is reduced
The relatively moderate contribution of Vein to the devel-opment of the DER-dependent muscle precursor cells raisesthe possibility that Vein complements or provides a backupmechanism for the activity of Spitz in these cells. In thatcase it is expected that reducing the activity of DER signal-ing pathway in the muscle precursor cells during their for-mation, may enhance the effect of loss of Vein. To test thispossibility we followed the formation of the DA1 muscle
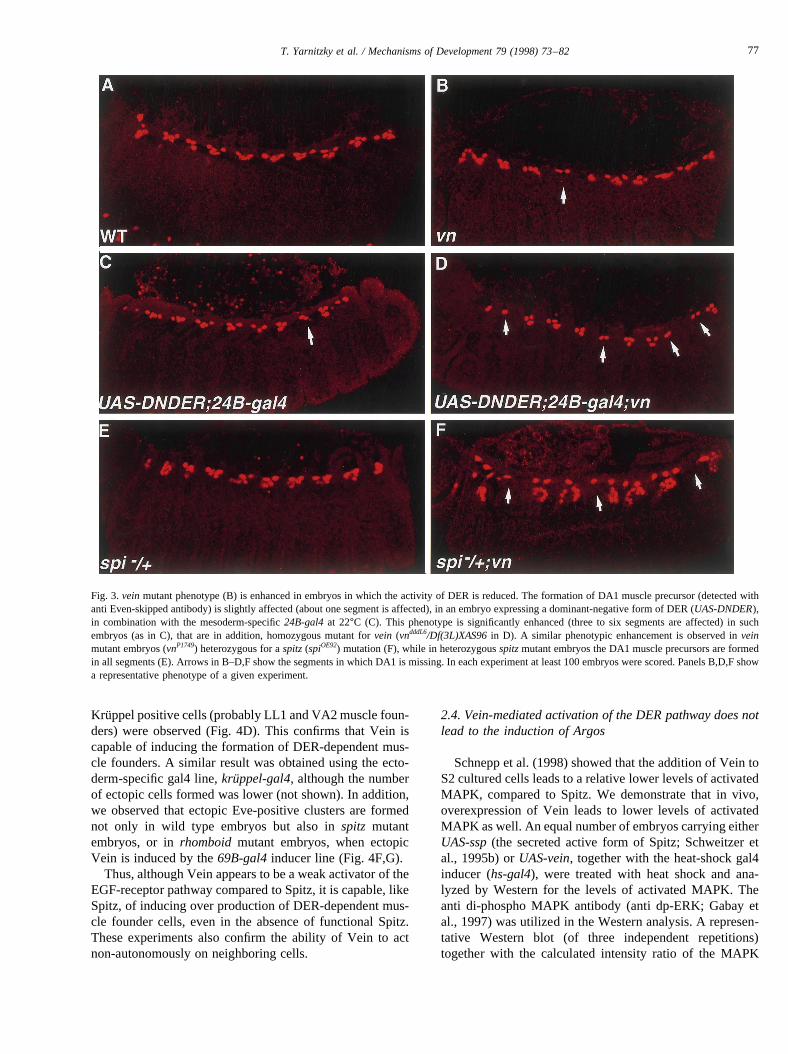
precursor in embryos in which the activity of DER wasreduced by expressing a dominant-negative form of DER(DNDER) in the mesoderm. This was achieved by inducingthe expression ofUAS-DNDERusing the mesoderm-speci-fic 24B-gal4line. In such embryos, grown at 22°C, the for-mation of the DA1 muscle precursor (detected by Eveexpression), is either normal or affected in not more thanone segment (Fig. 3C). In embryos carrying both aveinmutation (vnP1749) and a UAS-DNDERconstruct inducedby 24B-gal4(at 22°C), the loss of DA1 muscle precursoris enhanced and observed in three to six segments (Fig. 3D).In contrast to the 50% penetrance observed in theveinmutants, this phenotype is fully penetrant.
To test more directly the model that Vein complementsthe activity of Spitz in the development of the DA1 muscleprecursor, we examined the formation of DA1 muscle pre-cursor invnmutant embryos carrying only one copy of wildtypespitz. While the DA1 muscle precursor is formed in allthe segments of embryos heterozygous for thespitzmuta-tion, thevn mutant embryos carrying only one copy of wildtype spitz, show significant enhancement in the penetrance(100% compare to 50%) of the phenotype (Fig. 3F). Inaddition, the number of affected segments was elevated(three instead of one), as demonstrated in Fig. 3. Both
Fig. 1. vein is expressed in a subset of somatic muscle precursors.vein mRNA is detected in a subset of somatic muscle precursors during and followingembryonic germ band retraction (A,B). Double labeling with anti-Vein (C, red) and anti-Kru¨ppel (D, green) antibodies, indicates that both proteins are co-localized in an embryo at stage 12-13. Panel E represents the merged Vein/Kru¨ppel image. The identity of the various muscles is indicated by arrowheads inC–E.
75T. Yarnitzky et al. / Mechanisms of Development 79 (1998) 73–82
experiments support the model in which Vein activity com-plements or provides a backup mechanism for the criticalcontribution of Spitz to the formation of DA1 muscle pre-cursor.
2.3. Vein is sufficient to induce the formation of the DER-dependent muscle founders
Vein appears to be a relatively weak EGF-receptor ligandcompared to Spitz (Schnepp et al., 1998). Ectopic expres-
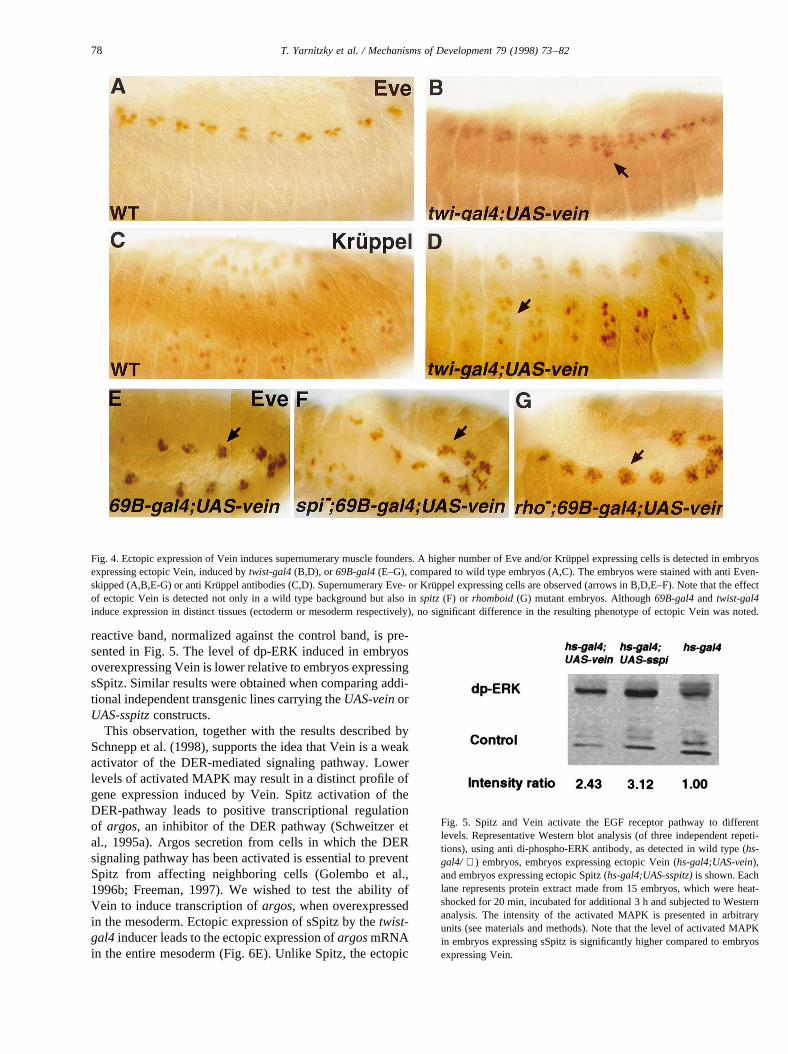
sion of the active secreted Spitz ligand in the mesodermresults in supernumerary muscle founder cells (Buff et al.,1998; and our unpublished results). To test the potential ofVein to induce the formation of supernumerary musclefounder cells, we overexpressed Vein in the mesoderm.Such overexpression using thetwist-gal4 inducer line,results in the formation of supernumerary Even-skippedpositive clusters in the dorsal domain of the embryo (Fig.4B), presumably representing overproduction of DA1 mus-cle founder cells. Additional clusters of supernumerary
Fig. 2. A partial loss of the somatic muscle precursors is observed invein mutant embryos. Wild type (A,C,E) andvein mutant (the embryos in B,D arevndddL6/Df(3L)XAS96, the embryo in F isvnD25/Df(3L)XAS96, and the embryos in G,H arevnP1749) were stained with anti-Even-skipped (A,B), anti-Kru¨ppel(C–F), or anti-myosin heavy chain antibodies (G,H). At stage 12–13, a loss of DA1 muscle precursor, identified by its reactivity with anti Even-skipped (B),or with anti Kruppel (D) antibodies, is observed in one segment of avein mutant embryo (the defected segment is indicated by arrows in B and D). TheKruppel positive muscle precursors LL1 and VA2 are also absent at a similar frequency in aveinmutant embryo (arrows in F). Note that in stage 15–16 (G,H)various myotubes at the ventral aspects of the embryo exhibit mis-routed pattern (arrow in G). At the dorsal aspects of such embryo DA1 is missing in 2segments (arrows in H), while DO1 is also missing in one segment, presumably due to abnormal guidance.
76 T. Yarnitzky et al. / Mechanisms of Development 79 (1998) 73–82
Kruppel positive cells (probably LL1 and VA2 muscle foun-ders) were observed (Fig. 4D). This confirms that Vein iscapable of inducing the formation of DER-dependent mus-cle founders. A similar result was obtained using the ecto-derm-specific gal4 line,kruppel-gal4, although the numberof ectopic cells formed was lower (not shown). In addition,we observed that ectopic Eve-positive clusters are formednot only in wild type embryos but also inspitz mutantembryos, or inrhomboid mutant embryos, when ectopicVein is induced by the69B-gal4inducer line (Fig. 4F,G).
Thus, although Vein appears to be a weak activator of theEGF-receptor pathway compared to Spitz, it is capable, likeSpitz, of inducing over production of DER-dependent mus-cle founder cells, even in the absence of functional Spitz.These experiments also confirm the ability of Vein to actnon-autonomously on neighboring cells.
2.4. Vein-mediated activation of the DER pathway does notlead to the induction of Argos
Schnepp et al. (1998) showed that the addition of Vein toS2 cultured cells leads to a relative lower levels of activatedMAPK, compared to Spitz. We demonstrate that in vivo,overexpression of Vein leads to lower levels of activatedMAPK as well. An equal number of embryos carrying eitherUAS-ssp(the secreted active form of Spitz; Schweitzer etal., 1995b) orUAS-vein, together with the heat-shock gal4inducer (hs-gal4), were treated with heat shock and ana-lyzed by Western for the levels of activated MAPK. Theanti di-phospho MAPK antibody (anti dp-ERK; Gabay etal., 1997) was utilized in the Western analysis. A represen-tative Western blot (of three independent repetitions)together with the calculated intensity ratio of the MAPK
Fig. 3.veinmutant phenotype (B) is enhanced in embryos in which the activity of DER is reduced. The formation of DA1 muscle precursor (detected withanti Even-skipped antibody) is slightly affected (about one segment is affected), in an embryo expressing a dominant-negative form of DER (UAS-DNDER),in combination with the mesoderm-specific24B-gal4at 22°C (C). This phenotype is significantly enhanced (three to six segments are affected) in suchembryos (as in C), that are in addition, homozygous mutant forvein (vndddL6/Df(3L)XAS96in D). A similar phenotypic enhancement is observed inveinmutant embryos (vnP1749) heterozygous for aspitz(spiOE92) mutation (F), while in heterozygousspitzmutant embryos the DA1 muscle precursors are formedin all segments (E). Arrows in B–D,F show the segments in which DA1 is missing. In each experiment at least 100 embryos were scored. Panels B,D,F showa representative phenotype of a given experiment.
77T. Yarnitzky et al. / Mechanisms of Development 79 (1998) 73–82
reactive band, normalized against the control band, is pre-sented in Fig. 5. The level of dp-ERK induced in embryosoverexpressing Vein is lower relative to embryos expressingsSpitz. Similar results were obtained when comparing addi-tional independent transgenic lines carrying theUAS-veinorUAS-sspitzconstructs.
This observation, together with the results described bySchnepp et al. (1998), supports the idea that Vein is a weakactivator of the DER-mediated signaling pathway. Lowerlevels of activated MAPK may result in a distinct profile ofgene expression induced by Vein. Spitz activation of theDER-pathway leads to positive transcriptional regulationof argos, an inhibitor of the DER pathway (Schweitzer etal., 1995a). Argos secretion from cells in which the DERsignaling pathway has been activated is essential to preventSpitz from affecting neighboring cells (Golembo et al.,1996b; Freeman, 1997). We wished to test the ability ofVein to induce transcription ofargos, when overexpressedin the mesoderm. Ectopic expression of sSpitz by thetwist-gal4 inducer leads to the ectopic expression ofargosmRNAin the entire mesoderm (Fig. 6E). Unlike Spitz, the ectopic
Fig. 4. Ectopic expression of Vein induces supernumerary muscle founders. A higher number of Eve and/or Kru¨ppel expressing cells is detected in embryosexpressing ectopic Vein, induced bytwist-gal4(B,D), or 69B-gal4(E–G), compared to wild type embryos (A,C). The embryos were stained with anti Even-skipped (A,B,E-G) or anti Kru¨ppel antibodies (C,D). Supernumerary Eve- or Kru¨ppel expressing cells are observed (arrows in B,D,E–F). Note that the effectof ectopic Vein is detected not only in a wild type background but also inspitz (F) or rhomboid(G) mutant embryos. Although69B-gal4and twist-gal4induce expression in distinct tissues (ectoderm or mesoderm respectively), no significant difference in the resulting phenotype of ectopic Vein wasnoted.
Fig. 5. Spitz and Vein activate the EGF receptor pathway to differentlevels. Representative Western blot analysis (of three independent repeti-tions), using anti di-phospho-ERK antibody, as detected in wild type (hs-gal4/ + ) embryos, embryos expressing ectopic Vein (hs-gal4;UAS-vein),and embryos expressing ectopic Spitz (hs-gal4;UAS-sspitz)is shown. Eachlane represents protein extract made from 15 embryos, which were heat-shocked for 20 min, incubated for additional 3 h and subjected to Westernanalysis. The intensity of the activated MAPK is presented in arbitraryunits (see materials and methods). Note that the level of activated MAPKin embryos expressing sSpitz is significantly higher compared to embryosexpressing Vein.
78 T. Yarnitzky et al. / Mechanisms of Development 79 (1998) 73–82
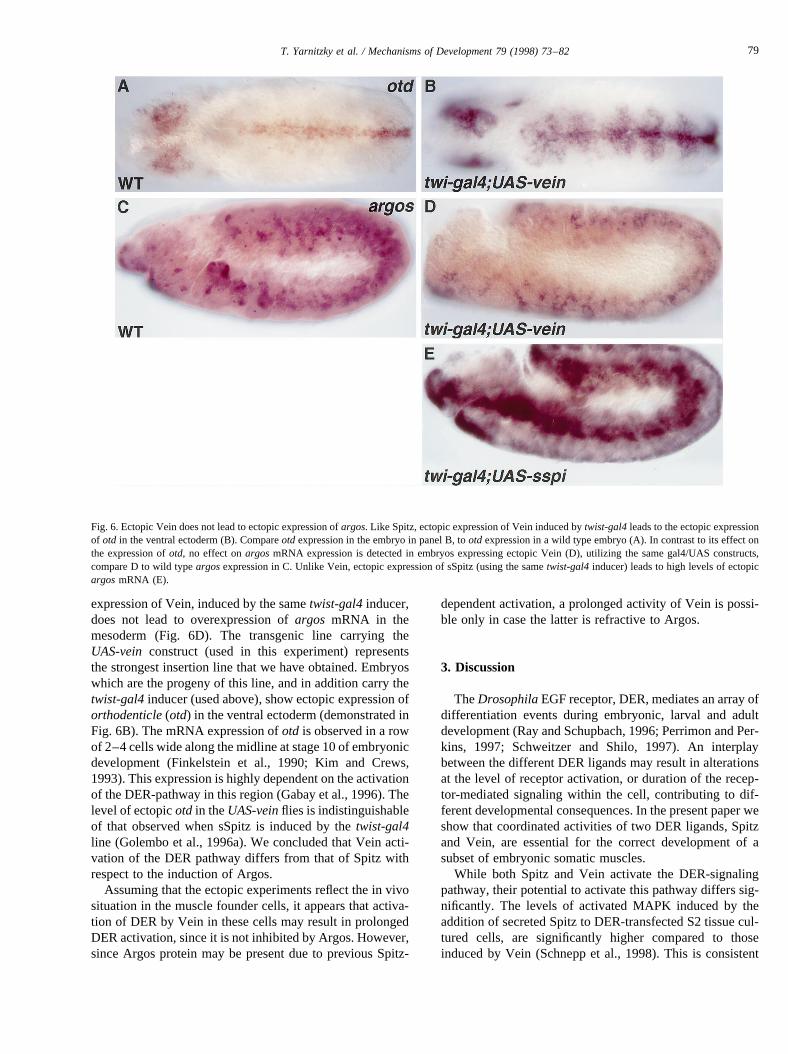
expression of Vein, induced by the sametwist-gal4inducer,does not lead to overexpression ofargos mRNA in themesoderm (Fig. 6D). The transgenic line carrying theUAS-vein construct (used in this experiment) representsthe strongest insertion line that we have obtained. Embryoswhich are the progeny of this line, and in addition carry thetwist-gal4inducer (used above), show ectopic expression oforthodenticle(otd) in the ventral ectoderm (demonstrated inFig. 6B). The mRNA expression ofotd is observed in a rowof 2–4 cells wide along the midline at stage 10 of embryonicdevelopment (Finkelstein et al., 1990; Kim and Crews,1993). This expression is highly dependent on the activationof the DER-pathway in this region (Gabay et al., 1996). Thelevel of ectopicotd in theUAS-veinflies is indistinguishableof that observed when sSpitz is induced by thetwist-gal4line (Golembo et al., 1996a). We concluded that Vein acti-vation of the DER pathway differs from that of Spitz withrespect to the induction of Argos.
Assuming that the ectopic experiments reflect the in vivosituation in the muscle founder cells, it appears that activa-tion of DER by Vein in these cells may result in prolongedDER activation, since it is not inhibited by Argos. However,since Argos protein may be present due to previous Spitz-
dependent activation, a prolonged activity of Vein is possi-ble only in case the latter is refractive to Argos.
3. Discussion
TheDrosophilaEGF receptor, DER, mediates an array ofdifferentiation events during embryonic, larval and adultdevelopment (Ray and Schupbach, 1996; Perrimon and Per-kins, 1997; Schweitzer and Shilo, 1997). An interplaybetween the different DER ligands may result in alterationsat the level of receptor activation, or duration of the recep-tor-mediated signaling within the cell, contributing to dif-ferent developmental consequences. In the present paper weshow that coordinated activities of two DER ligands, Spitzand Vein, are essential for the correct development of asubset of embryonic somatic muscles.
While both Spitz and Vein activate the DER-signalingpathway, their potential to activate this pathway differs sig-nificantly. The levels of activated MAPK induced by theaddition of secreted Spitz to DER-transfected S2 tissue cul-tured cells, are significantly higher compared to thoseinduced by Vein (Schnepp et al., 1998). This is consistent
Fig. 6. Ectopic Vein does not lead to ectopic expression ofargos. Like Spitz, ectopic expression of Vein induced bytwist-gal4leads to the ectopic expressionof otd in the ventral ectoderm (B). Compareotd expression in the embryo in panel B, tootd expression in a wild type embryo (A). In contrast to its effect onthe expression ofotd, no effect onargosmRNA expression is detected in embryos expressing ectopic Vein (D), utilizing the same gal4/UAS constructs,compare D to wild typeargosexpression in C. Unlike Vein, ectopic expression of sSpitz (using the sametwist-gal4inducer) leads to high levels of ectopicargosmRNA (E).
79T. Yarnitzky et al. / Mechanisms of Development 79 (1998) 73–82

with our in vivo analysis of the levels of activated MAPKinduced by either secreted Spitz or Vein. The lower activa-tion of DER by Vein may lead to qualitatively differentconsequences in terms of gene expression. For examplewe have shown that unlike Spitz, Vein overexpressiondoes not lead to Argos expression. A prolonged expressionof Vein in the muscle founders, thus, may lead to a contin-uous activation of the EGF-receptor pathway in these cells.
Since Vein is a secreted protein, while Spitz is producedas a membrane-bound, non-active DER ligand, distinctmechanisms control their mode of DER activation. Regula-tion of Vein activity appears to be at the transcriptionallevel, since its ectopic expression in various tissues (e.g.ectoderm or mesoderm) leads to measurable biological con-sequences (Schnepp et al., 1998; and our results). Spitzactivity is controlled by the expression of Rhomboid orStar, presumably by its processing into an active secretedform (Schweitzer et al., 1995b).
3.1. Different modes of Vein activity during embryonicdevelopment
Although Vein is a less potent activator of DER, it func-tions as a single ligand in certain tissues, such as in tendoncell differentiation (Yarnitzky et al., 1997). In this tissue,the limited activation of DER by Vein may be compensatedby concentrating it at the site of activity, i.e. at the muscle-tendon junctional site. The accumulation of Vein at this sitemay be also essential to restrict the number of affected cells.
Spitz functions in a wide array of embryonic tissueswhere Vein mRNA is also detected, e.g. the somatic meso-derm, ventral mesectodermal cells, tracheal pits, and chor-dotonal organs. The biological contribution of Vein intissues where Spitz appears to be the major ligand (accord-ing to the resulting mutant phenotype), could be explainedby two alternative mechanisms: Vein could act synergisti-cally with Spitz, or, Vein may act sequentially to Spitz toprovide a low, but continuous level of DER activationwhich is required to backup, or complement Spitz activity.Simultaneous expression of Vein together with sSpitz inembryos, using heat-shock-gal4, did not lead to higherlevels of activated MAPK (our unpublished results). There-fore we do not favor the model of synergistic activation ofDER by both ligands. To this end, our results support theidea that the role of Vein in tissues where Spitz is the majorligand is to complement Spitz activity. This explains theextremely weakvein phenotype observed in these tissues,in comparison to a significant and measurable phenotypeobtained in tissues where Vein functions as a single ligand.It is also consistent with the genetic interaction betweenveinandspitz, described by Schnepp et al. (1996).
veinmRNA is also observed in domains in which Spitzor Rhomboid contribution has not yet been described, e.g.the amnioserosa, the dorsal most row of ectodermal cellsduring germ band retraction and head structures. Noobvious phenotype is observed in these tissues invein
mutant embryos and the role of Vein in these tissues isyet to be elucidated.
3.2. Vein functions sequentially to Spitz
Vein appears to act sequentially to Spitz sinceveinmRNA in the somatic mesodermal cells follows Rhomboidexpression in the progenitors (Buff et al., 1998). Rhomboidexpression appears to represent the local activation of Spitz(Schweitzer et al., 1995b). The delayed expression of veinmRNA, is consistent with the suggestive function of Vein asa ligand that complements Spitz activity. It is possible thatSpitz activation of DER positively regulates the transcrip-tion of vein in the founder cells. In line with this possibilityis our observation that overexpression of Spitz leads to theexpression of Vein (Golembo et al., unpublished).
The nature of the inter-relationship between Vein andArgos is yet to be elucidated. Since activation of DER bySpitz leads to Argos expression (Golembo et al., 1996b), itmay inhibit Vein activity. Vein activity in the muscle foun-ders suggests that these cells are either refractive to Argos orthat Argos does not inhibit Vein activity.
In summary, an interplay between the temporal expressionand differential activities of two EGF-receptor ligands des-cribed in this work, may be essential for a robust receptoractivation required to accomplish specific differentiation pro-grams.
4. Materials and methods
4.1. Fly stocks
The following gal4 inducers were used:69B-gal4, twist-gal4, 24B-gal4 (A. Brand, Cambridge, UK),kruppel-gal4(M. Leptin, Koln, Germany),hs-gal4 (K25-2Xsev hsp70-GAL4, obtained from E. Hafen, Zu¨rich, Switzerland). Inaddition the following strains were used:y w (wild-typestrain); Df(3L)XAS96 (Dr. W.A. Johnson, Iowa, WI);spit-zoe92(N. Perrimon, MA, USA);veinD25 (created in our lab byexcision of the P-element invnP1749), veindddL6 (A. Simcox,OH, USA), vnP1749 (A.C. Spradling, MD, USA);UAS-DNDER, UAS-Secreted-Spitz 17a; UAS-Secreted-Spitz 4b(B. Shilo, Rehovot, Israel).UAS-vein flies were producedin our lab (Yarnitzky et al., 1997). TheUAS-vein110 inser-tion induces a high expression level and UAS-vein 98repre-sents a weaker line. Mutant embryos were identified using‘blue balancers’ and anti-b-galactosidase staining.
4.2. Immunochemical reagents
Vein was visualized either by in situ hybridization with adigoxigenin-labeled 3.4 kbveincDNA fragment, or by anti-body against GST-Vein fusion protein, raised in rats. Argosexpression was monitored by in situ hybridization usingargos RNA as probe (M. Freeman, MRC, Cambridge). An
80 T. Yarnitzky et al. / Mechanisms of Development 79 (1998) 73–82

otd clone was obtained from R. Finklestein (University ofPennsylvania, PA). Anti Kru¨ppel antibody was obtainedfrom C. Rushlow (NYU, New York, NY) and anti Even-skipped antibody was obtained from M. Frasch (MountSinai Hospital, New York, NY). Anti-myosin heavy chain(MHC) antibody was obtained from P. Fisher (Stony Brook,NY). The level of activated MAPK was monitored by antidiphospho-ERK (Sigma). Anti-b-galactosidase antibodieswere purchased from Cappel (USA).
Secondary antibodies included HRP, Fluorescein, Rhoda-min or Cy3-conjugated Goat or Donkey anti Rabbit or antiRat IgG (Jackson, USA), and anti-dig-AP antibody (Boeh-ringer Mannheim, Germany).
4.3. Whole mount embryonic staining
Staining was performed essentially as described (Ashbur-ner, 1989). In brief, embryos were collected and incubatedas indicated, dechorionated and fixed in a mixture of 3%paraformaldehyde and heptane. Following two washes,embryos were stained with X-gal staining solution, permea-bilized, and stained with primary antibody, and second anti-body. Embryos labeled with fluorescent antibody weredouble-labeled with anti-b-galactosidase antibody to iden-tify mutant embryos in situ hybridization was performed bythe method of Tautz and Pfeiffle (1989).
Stained embryos were examined under a Zeiss Axioscopemicroscope.
4.4. Flat preparation of embryos
Flat preparations (used for fluorescent staining in Fig. 1),were prepared essentially according to Bate (1990); liveembryos were dechorionated and the vitteline membranewas removed by hand. The embryos were opened and flat-tened on poly-l-Lysine covered coverslips, fixed with 3%paraformaldehyde in PBS, and stained.
4.5. Western analysis
Embryos containing eitherUAS-veinor UAS-sspitz, inaddition tohs-gal4 inducer and alsohs-gal4/+ embryos,were collected for 2 h, incubated for 3 h at 25°C, heat-shocked at 37°C for 20 min, and returned to 29°C for 3 h.Embryos were boiled in sample buffer and subjected to SDS-PAGE and Western analysis. HRP-conjugated-antibody wasused and detected by Super signal substrate (Pierce). Eachlane represents 15 embryos. The intensity of the bands wasanalyzed using NIH Image version 1.55 (NIH, MD, USA).The intensities of the MAPK reactive band and of a controlband, were calculated for each cross, and the ratio betweenthese numbers was compared to the corresponding ratio ofthe control experiment (hs-gal4/+ embryos).
4.6. Confocal microscopy
Fluorescent labeled preparations were imaged using a
BioRad MRC 1024 confocal microscope coupled to aZeiss Axiovert 135M microscope. Bright field and fluores-cent digital images were processed using Photoshop version3.0 (Adobe Systems Inc., CA, USA).
Acknowledgements
We thank A. Brand, M. Leptin, N. Perrimon, E. Hafen, B.Shilo, A. Simcox and W.A. Johnson for various fly strains;C. Rushlow, M. Frasch, M. Freeman, P. Fisher, and B. Shilofor antibodies and cDNA probes; B. Shilo, and E. Schejterfor critical reading of the manuscript; B. Shilo, M. Golemboand A. Michelson for fruitful discussions and suggestions.This work was supported by a grant from the Israel ScienceFoundation (T.V.).
References
Abmayr, S.M., Erickson, M.S., Bour, B.A., 1995. Embryonic developmentof the larval body wall musculature ofDrosophila melanogaster. TrendsGenet. 11, 153–159.
Ashburner, M., 1989.Drosophilaa Laboratory Manual. Cold Spring Har-bor Laboratory Press, Cold Spring Harbor, NY, pp. 44–49.
Bate, M., 1990. The embryonic development of larval muscles inDrosophila. Development 110, 791–804.
Bate, M., 1993. The mesoderm and its derivatives. In: Bate, M., MartinesArias, A. (Eds.), The Development ofDrosophila Melanogaster. ColdSpring Harbor Laboratory Press, Cold Spring Harbor, NY, pp. 1013–1090.
Baylies, M.K., Bate, M., 1996.twist: a myogenic switch inDrosophila.Science 272, 1481–1484.
Baylies, M.K., Bate, M., Ruiz Gomez, M., 1998. Myogenesis: a view fromDrosophila. Cell 93, 921–927.
Baylies, M.K., Martinez Arias, A., Bate, M., 1995.winglessis required forthe formation of a subset of muscle founder cells duringDrosophilaembryogenesis. Development 121, 3829–3837.
Bourgouin, C., Lundgren, S.E., Thomas, J.B., 1992.apterousis a Droso-phila LIM domain gene required for the development of a subset ofembryonic muscles. Neuron 9, 549–561.
Buff, E., Carmena, A., Gisselbrecht, S., Jimenez, F., Michelson, A.M.,1998. Signalling by theDrosophila epidermal growth factor receptoris required for the specification and diversification of embryonic muscleprogenitors. Development 125, 2075–2086.
Carmena, A., Bate, M., Jimenez, F., 1995.Lethal of scute, a proneuralgene, participates in the specification of muscle progenitors duringDro-sophilaembryogenesis. Genes Dev. 9, 2373–2383.
Carmena, A., Murugasu-Oei, B., Menon, D., Jimenez, F., Chia, W., 1998.Inscuteableandnumbmediate asymmetric muscle progenitor cell divi-sions duringDrosophilamyogenesis. Genes Dev. 12, 304–315.
Dohrmann, C., Azpiazu, N., Frasch, M., 1990. A newDrosophilahomeo-box gene is expressed in mesodermal precursor cells of distinct musclesduring embryogenesis. Genes Dev. 4, 2098–2111.
Finkelstein, R., Smouse, D., Capaci, T.M., Spradling, A.C., Perrimon, N.,1990. Theorthodenticlegene encodes a novel homeo domain proteininvolved in the development of theDrosophila nervous system andocellar visual structures. Genes Dev. 4, 1516–1527.
Frasch, M., 1995. Induction of visceral and cardiac mesoderm by ectoder-mal Dpp in the earlyDrosophilaembryo. Nature 374, 464–467.
Frasch, M., Hoey, T., Rushlow, C., Doyle, H., Levine, M., 1987. Char-acterization and localization of the even-skipped protein ofDrosophila.EMBO J. 6, 749–759.
81T. Yarnitzky et al. / Mechanisms of Development 79 (1998) 73–82

Freeman, M., 1997. Cell determination strategies in theDrosophilaeye.Development 124, 261–279.
Gabay, L., Scholz, H., Golembo, M., Klaes, A., Shilo, B.Z., Klambt, C.,1996. EGF receptor signaling inducespointedP1 transcription and inac-tivates Yan protein in theDrosophila embryonic ventral ectoderm.Development 122, 3355–3362.
Gabay, L., Seger, R., Shilo, B.Z., 1997. In situ activation pattern ofDro-sophilaEGF receptor pathway during development. Science 277, 1103–1106.
Gaul, U., Seifert, E., Schuh, R., Ja¨ckle, H., 1987. Analysis of Kruppelprotein distribution during earlyDrosophiladevelopment reveals post-transcriptional regulation. Cell 50, 639–647.
Golembo, M., Raz, E., Shilo, B.Z., 1996. TheDrosophilaembryonic mid-line is the site of Spitz processing, and induces activation of the EGFreceptor in the ventral ectoderm. Development 122, 3363–3370.
Golembo, M., Schweitzer, R., Freeman, M., Shilo, B.Z., 1996.Argostranscription is induced by theDrosophila EGF receptor pathway toform an inhibitory feedback loop. Development 122, 223–230.
Hartenstein, A.Y., Rugendorff, A., Tepass, U., Hartenstein, V., 1992. Thefunction of the neurogenic genes during epithelial development in theDrosophilaembryo. Development 116, 1203–1220.
Kim, S.H., Crews, S.T., 1993. Influence ofDrosophilaventral epidermaldevelopment by the CNS midline cells andspitz class genes.Development 118, 893–901.
Lawrence, P.A., Bodmer, R., Vincent, J.P., 1995. Segmental patterning ofheart precursors inDrosophila. Development 121, 4303–4308.
Michelson, A., Abmayr, S., Bate, M., Martinez Arias, A., Maniatis, T.,1990. Expression of aMyoD family member prefigures muscle patternin Drosophilaembryos. Genes Dev. 4, 2086–2097.
Nose, A., Mahajan, V.B., Goodman, C.S., 1992. Connectin: a homophiliccell adhesion molecule expressed on a subset of muscles and the moto-neurons that innervate them inDrosophila. Cell 70, 553–567.
Park, M., Wu, X., Golden, K., Axelrod, J.D., Bodmer, R., 1996. Thewingless signaling pathway is directly involved inDrosophila heartdevelopment. Dev. Biol. 177, 104–116.
Perrimon, N., Perkins, L.A., 1997. There must be 50 ways to rule thesignal: the case of theDrosophila EGF receptor. Cell 89, 13–16.
Ranganayakulu, G., Schulz, R.A., Olson, E.N., 1996. Wingless signalinginducesnautilusexpression in the ventral mesoderm of theDrosophilaembryo. Dev. Biol. 176, 143–148.
Ray, R.P., Schupbach, T., 1996. Intercellular signaling and the polariza-tion of body axes duringDrosophilaoogenesis. Genes Dev. 10, 1711–1723.
Ruiz Gomez, M., Bate, M., 1997. Segregation of myogenic lineages inDrosophilarequiresnumb. Development 124, 4857–4866.
Ruiz Gomez, M., Romani, S., Hartmann, C., Ja¨ckle, H., Bate, M., 1997.Specific muscle identities are regulated by Kruppel duringDrosophilaembryogenesis. Development 124, 3407–3414.
Rushton, E., Drysdale, R., Abmayr, S.M., Michelson, A.M., Bate, M.,1995. Mutations in a novel gene,myoblast city, provide evidence insupport of the founder cell hypothesis forDrosophila muscledevelopment. Development 121, 1979–1988.
Rutledge, B.J., Zhang, K., Bier, E., Jan, Y.N., Perrimon, N., 1992. TheDrosophila spitz gene encodes a putative EGF-like growth factorinvolved in dorsal-ventral axis formation and neurogenesis. GenesDev. 6, 1503–1517.
Schnepp, B., Donaldson, T., Grumbling, G., Ostrowski, S., Schweitzer, R.,Shilo, B.Z., Simcox, A., 1998. EGF domain swap converts aDrosophilaEGF receptor activator into an inhibitor. Genes Dev. 12, 908–913.
Schnepp, B., Grumbling, G., Donaldson, T., Simcox, A., 1996. Vein is anovel component in theDrosophila epidermal growth factor receptorpathway with similarity to the neuregulins. Genes Dev. 10, 2302–2313.
Schweitzer, R., Howes, R., Smith, R., Shilo, B.Z., Freeman, M., 1995.Inhibition of DrosophilaEGF receptor activation by the secreted proteinArgos. Nature 376, 699–702.
Schweitzer, R., Shaharabany, M., Seger, R., Shilo, B.Z., 1995. SecretedSpitz triggers the DER signaling pathway and is a limiting component inembryonic ventral ectoderm determination. Genes Dev. 9, 1518–1529.
Schweitzer, R., Shilo, B.Z., 1997. A thousand and one roles forDrosophilaEGF receptor. Trends Genet. 13, 191–196.
Skeath, J.B., 1998. TheDrosophila EGF receptor controls the formationand specification of neuroblasts along the dorsal-ventral axis of theDrosophilaembryo. Development 125, 3301–3312.
Staehling-Hampton, K., Hoffmann, F.M., Baylies, M.K., Rushton, E.,Bate, M., 1994. dpp induces mesodermal gene expression inDrosophila. Nature 372, 783–786.
Tautz, D., Pfeiffle, C., 1989. A nonradioactive in situ hybridization methodfor the localization of specific RNAs inDrosophilaembryos reveals atranslational control of the segmentation genehunchback. Chromosoma98, 81–85.
Williams, J.A., Bell, J.B., Carroll, S.B., 1991. Control ofDrosophilawingand haltere development by the nuclearvestigial gene product. GenesDev. 5, 2481–2495.
Yarnitzky, T., Min, L., Volk, T., 1997. TheDrosophilaneuregulin homo-log Vein mediates inductive interactions between myotubes and theirepidermal attachment cells. Genes Dev. 11, 2691–2700.
82 T. Yarnitzky et al. / Mechanisms of Development 79 (1998) 73–82