an experimental study of the thalamo-cortical projection of the macaque monkey
TRANSCRIPT

Iicconsiclcrat ion of tlic hasic aiiatomiciil studies of the cwitral iiervoiis system has btwi giveii inipetus in the last f t w ycni's by tlic atlvanccs iiiaclc in ~icuropliysiology and in ncuro- sii I*% i cal t cc l i~ i i quc. 'Ph ese invest i g:i t ion s have cleared many funclniiicntal 1)roblems. 'l'lic knowlcdgcl of the t l d a r n u s and it:, coiinections has heen augmented hy the researches of Riocli ( '%I), 1'oli:ik ( ':<a), Xett ler (%), Le Giws Clark and 12oggoii ( '32, ';XI, aiitl 'X), Sager ( '3R), and ( ' ro~icli ( '34). 111 sl)itcl of t l i c s c h cwiitributions thcl 1 ~ i ~ ) l ) l ~ ~ of the cortical I.ci)i.csciiliitioii of tlic tlialamus still remains. Hccanse of this 1ac.k tile 1)rcsent essay tries to slicd some light on tlie afferent pathways fi-oiii tlie tlialanius to t l i r h ccwhl*al cortex.
Much of the 1)revious work on this problem has been clone using the hLar*c*hi tidiiiicliie. The dificulty in making minute lesions witliiii tlic llialtimus, and in tlctermining the exact origin of the fi1wi.s i n t c w u p t t d , has led to considerable con- fusion i n aiidyziiig tlic results of siicli investigations. T o avoit1 tliosc obstacles the. rnetliocl of tlie retrograde cell cleyeii- chixtion Iias 1)ccil usrd iii this work. I t is h ~ d upoil the i'ilct ol)stw-ctl long ago by Sissl tliat wlieii ail axis cylinder of a neiiron mywhere t110ii.g i ts Course is injured distinct changes
Aidetl i iy a graiit f'roin tlic 1)ougl:is Rinitli 1'ountl:itioii.
I
TIO. . I I J L R N A I , O F i o u t ' t ~ \ ' r i ~ t . S F . I M J X O ( & ~ , 101. (2, so 1 JL'NI, 1'1%6

2 A. EARL WALKER
occur within its cell body. In neuropathological texts these are described as ‘primary irritation’ of the ganglion cell. In many neuronic systems the cell dies and is removed by the phagocytes of the central nervous system-the microglia. If a large number of cells of a compact nucleus undergo de- generation they produce certain microscopic changes that are readily recognized. These may be an absence or a decreased number of nerve cells with or without a gliosis. I n the pulvinar and in the nucleus medialis dorsalis the reaction is character- istic : the complete disappearance of the normal nerve cells fol- lowed by a marked gliosis ; in the posterior half of the lateral and ventral nuclei the gliosis is usually less pronounced. I n the anterior half of the ventral and lateral nuclei the gliosis is frequently quite marked-even though all of the nerve cells in that region have not disappeared (fig. 1). This variation is probably due to some biological properties of the cell specific for each region and cell variety; the difference in reaction certainly is not due to the length of the time allowed for degeneration nor to the severity of the lesion.
As Bielschowsky ( ’32) and others have pointed out, the term retrograde cell degeneration is not quite accurate; it should be known as retrograde cell reaction because in many in- stances, for as yet unknown reasons, the cells may not degen- erate completely and a reversal of the process may occur. This fact should be kept in mind for it may explain otherwise incompatible findings. Hence, the absence of retrograde cell degeneration in a nuclear mass following a cortical lesion is not an absolute proof of a lack of corticipetal fibers.
As almost all of the research upon the thalamo-cortical connections has been done on the Carnivora, it was thought advisable to study the entire problem in one of the primates. As the macaque monkey is the most accessible and cheapest of the primates, it was chosen as the subject for this investigation. Over forty monkeys have been used in this research and many of these were utilized for two different problems, one on each hemisphere. The use of both hemispheres of an animal may be subject to criticism, as it may be contended that some of

thc thalaniic nuclei may be coiiiiected with the cortex in both Iicmisphei*es. However, the above procedure was not adopted until such a possibility was fully iiivestigatetl. I n no case in the present series has a retrograde degeneration been found in the thalamus opposite the side of a lesioii, although ex- amiiiatioiis of such thalairii have been made in many cases some of wliich had been prnetieally completely heinidecorti- caatctl. Then, too, I’oliak ( ’32 ) has never found Sfarchi granules going across the corpus callosum following thalamic lesions. Fo r that reason it is unlikely that an error is intro- d u c ~ d ~v.hc11 both hemispheres are used. I n the protocols,
A, nucleus angularis A.D., ~iurleus aiitero-dorsalis A.M., nurleus nntero-nieclialis Aq, aqueductus s j lrii A S . , lateral parictal occipital sulcus
A.V., nucleus antero-ventralis (’.t’., corpus callosuni (“.T,., nucleus rrntralis lateralis C‘.M,, iiucleus relitruin mediaiiuni (’.Min., corpus in:iniill:irc C’.S., crntral sulcus I ) , tlcntatc g j rus E.C., external calrarinr fissure F, fornix (;.I)., glohus pallidus I{, h? potlialamus Ila., habenula fl.P.S., horizontal linib of th r inferior
prcwntral sulcus 1, inferior nurleus of the pulrinar I.(”., internal capsule I . F . , inferior frontal sulcus T . H . , inferior horn T.K., nuclcus interstitialis (of Cajal) T.O., inferior occipital sulcus T.P. , infrrior precentral suleus (arcuntc
IS., intraparietal sulcus T,, nucleus limitaus lA.T)., rtucleus lateralis dorsalis L.G., lateral gciiiculate body L.P., nurleus lateralis posterior
(simim )
sulcus)
TAV., 1ater:il ventricle M.D., nucleus nirdi:iliu dorsalis >I . (> . , medial gcnirulate body M.T., superior teniporal sulcus S.C., nurlcus caudatus 0, nucleus oculoniotorius OI.T., olfactorv t ra r t O.T., optic tract I’, nucleus parafascicularis I’a, nucleus paracentralis Pi, pineal body I ’ L , lateral nuclc%us of the pulvinnr P..M., medial nurleus of the pulvinar Pu, piit:imcn H, nuelcus reticularis S, subtlialamic body S.C., callosal sulcus S.(‘a., calcarine sulcus S.S. , suhstaiitia nigra SP., suprrior prccentral gyms Y.Po., superior postcentral suleus S.S., sp1ri:tn sulcus S.T., superior temporal sulrus Su., nuclrus submedius T, trctuni nirsenwphali T.M., Mrvnert ’s tract T.M.T., tr:ictus nlarnillo-tlialamirus V.A., nuclcus ventralis anterior V.L., nucleus ventralis laterdis V.P.L., nucleus ventralis postero-
V.P.M., nucleus rcmtralis postero- lateralis
mrdialis
4 A. EARL WALKER
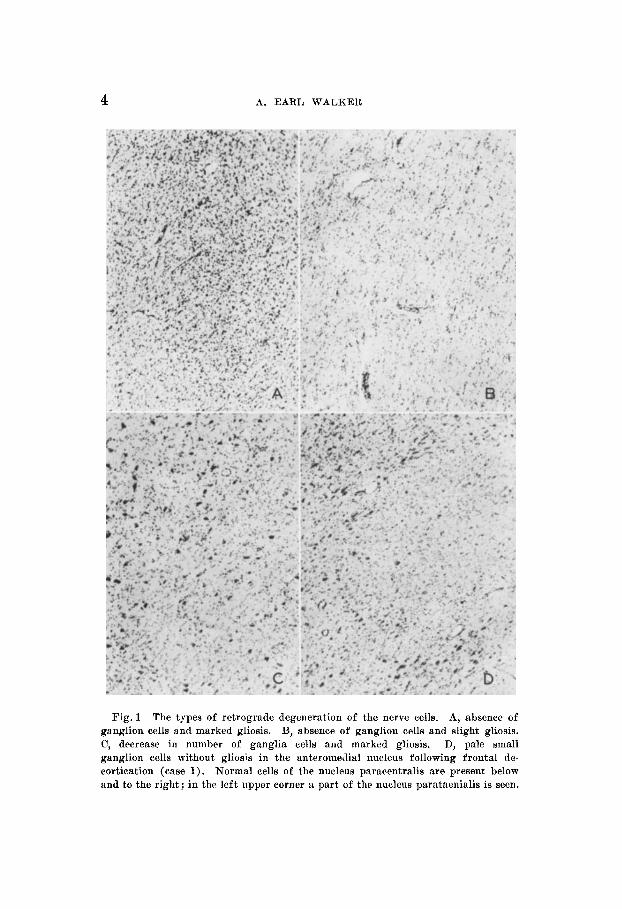
Fig. 1 The types of retrograde degeneration of the nerve cells. A, absence of ganglion cells and marked gliosis. B, absence of ganglion cells and slight gliosis. C, decrease in number of ganglia cells and marked gliosis. D, pzle small ganglion cells without gliosis in the anteromedial nucleus following frontal de- cortication (case 1). Normal cells of the nucleus paracentralis are present below and to the right; in the left upper corner a part of the nucleus parataenialis is seen.

THALAMO-CORTICAL PROJECTIOS 5
I ~ o w v c r , all proccdiires on any animal a re recorded. Practi- cally all of the experiments reported here h a w been repeated or duplicated as a precaution against chance experimental errors. J~’h~never , discrepancies occurred betweeii the re- siilts of the present work and those of previous investigators, tlie ex1)wirneiit TWS repeated to be certain that the findings were accura t c.
As this report deals with a strictly anatomical problem, any physiological observation on the experimental animals has hcen clelc~ted from the protocols. Because the nomenclature cm1)lopcd in descriptions of the tlialaniic nuclei is at present in a confused state, the author has made extensive use of illustrations for it is believed that these will add more in- forniation than a lengthy narrative using unfamiliar terms. All of the sketches of the thalamus were drawn from pro- jections of the sections (using an Eclinger projector) arid the dctails wcre filled in by microscopic control. The camera lucida m w used in sketching tlie lesions in the cerebral cortex. In every case representative serial sections of the cortex have 1)cen reprochcetl to show the situation and extent of the original lesioii, and in the same case representative serial sections of the thalamus are presented to enable the reader nc~ui*;itcly to locate the retrograde cell degeneration. The number nnclcr the drawing of each section is the number of the section froni which the sketch was made. The nomencla- ture adopted in this paper for tlie description of tlie cGnfigura- tion of the cci*cI~ral cortex is that presented by Nettler (’33). The terniinology used in discussing the nuclear configuration of the thalamus with a few exceptions is that of Crouch ( ’34). He divides the lateral and ventral nuclei on a morpho- logical basis into anterior and posterior halves at about tlie level of the rostra1 end of the median central nucleus. Since there is no clear morphological distinction between the an- terior moieties of these nuclei and they appear to have similar fiber connections, they have been grouped together in this in- vestigation uiicler the name nucleus ventralis lateralis. The most anterior part of this nucleus is distinct from the re-

6 A. EARL WALKET;
inainder on the grounds of both its morphology and fiber coli- nections; i t has been cialletl ( n f t o r Aroiison and Papez, ’34) the nuclens vmtralis anterior. Except for the above, and the rccogriitioii of the nucleus siibmcdins as a distinct cell group apart from the nucleus paracwitralis, the iiomciiclaturc and tlclimitatioii of the tlialamic nuclci f‘ollotm t 7roucli ’s ( ’34) terminology (fig. 21).
The operative and histological techiiiqucs have been de- scribed previously, but will be briefly reviewed here. Either ether or sodium amytal admiiiistcred intraperitoneally were used to obtain surgical ancsthesia. k’ollowing the operation the animals were allowetl to live from G weeks to 3 months. The Ic~igth of time within the above limits seeriied to makv little tliflcreiice i n the cleariicw aiid degree of retrograde cell (legeneration. The animals were killed by cliloroform and tlie brains removed in toto. blidsagittal sections were made and in some cases the thalanii were removed. A4fter fixation in 95 per cent alcoliol for a few days, the hemispheres were examined and the cortex drawn to show the lesion with the aid of a camera lucida. The parts were then further fixetl in 95 per cent nlcohol, alcohol aiid ether mid emheddcd in celloidin. The hemispheres were cut upon a celloidiii micro- tome at 30 p and every twenty-fifth wetion stained by Nissl’s toludine hlne metlioci. The thalainus was cut a t 25 p and de- pending upon the case every sectiori or every second, third, or in sonic cases every tenth section was stained by X’issl’s method.
The problem of determining the thalamic projection may be attacked, using the principle of retrograde cell clegenera- tion, in one of two ways. Either small cortical lesions may he made in many animals, o r large ablations performccl on a few animals. The first method gives the minute or point to point projection, while the second shows more gi’oss feat wcs of the thalaniic connections. In our present state of confusion, a general conception of the thalaniic projection is essential lie- fore a finer pattern is even considered. Hence, in this study attention will be largclly coiifinecl to the second method. Re- fore tlie results obtained by that means can be interpreted i t is necessary to know the slim total of the tlialamic cells sending fibers to the cerebral cortex.

THALAMO-CORTICAL PROJECTION 7
Through the courtesy of Dr. John F. Fulton of the Yale Medical School, the thalami of two hemidecorticated Macaca mulatta monkeys were studied. These have been reported fully in an earlier paper, and only the conclusions will be discussed here. When the cortex of one hemisphere is re- moved and the thalamus examined, extensive degeneration is seen. All three anterior nuclei are devoid of normal cells and there is a moderate gliosis at the site of the nucleus. The parvicellular part of the dorsal medial nucleus is likewise com- pletely degenerated, and shows a marked gliosis. The large- celled portion of this nucleus remains intact. The nuclei of the midline, the nuclei parataenialis medialis and lateralis, interparataenialis, paraventricularis anterior and posterior, rhomboideus, paracentralis, centralis medialis, and the central gray mass including the nucleus reuniens posterior show no alteration. The intralaminar nuclei, the nucleus centrum medianum, nucleus centralis lateralis and parafascicularis, if this can be accurately described as an intralaminar nucleus, are normal. True, the nucleus centralis lateralis does not contain as many cells as in a normal preparation but this is due to degenerated misplaced cells from the adjoining nuclei. The pulvinar is completely degenerated, and shows a marked gliosis. Both the ventral and lateral nuclei are completely changed, no normal cells being present except in the nucleus ventralis anterior which is practically normal, and the nucleus lateralis dorsalis which is also preserved, although there are a number of degenerated cells on its ventral border, which probably belong to the subjacent nuclei. The nucleus reticularis is also degenerated except at its oral extremity.
It is then apparent that the main thalamo-cortical fibers arise from the cells within the pulvinar, median, ventral and lateral nuclei. Former investigators have found connections from the center median but in not one of the cases of the present series has there been degeneration within this nucleus.
TEALAMIC PROJECTION UPON THE FRONTAL LOBE
The thalamic projection to the frontal cortex has occasioned much discussion. To determine the total connections, the cor- tex of the frontal lobe was removed and the resultant retro- grade cell changes in the thalamus were studied.
TEE JOURNAL OF COMPARATIVE NBUBOLOQY. VOL. 64, NO. 1

8 A, EARL WALKER
Case 1 (E-1-35L). On January 7, 1935, the cortex of the left frontal lobe was removed from an immature male Macaca mulatta. One week later several small lesions were made in the posterior portion of the right parietal lobe. On February 14, 1935, the animal wag sacrificed and the brain removed.
Grossly the cortex of the left frontal lobe anterior to the central fissure is completely ablated. The posterior portion of the orbital surface of the frontal lobe remains intact. The extent of the lesion is verified by study of serial sections of the left hemisphere (fig. 2). The cortex of the frontal lobe over the convexity of the hemisphere has been completely ablated to the central sulcus. On the anterior wall of the superior portion of that sulcus in a few sections can be
Fig.2 The hemiephere and serial eectiona of the cerebral cortex of cam 1 (E-1-35) to ahow the sitnation and extent of the frontal decortication.
seen large Betz’s cells of the area 4. Medially the injury extends down the paracentral lobule to the surface of the corpus callosum which is damaged on its superior surface. A thrombotic lesion ex- tends posteriorly in the paracentral lobule to the anterior part of area 3 of the parietal lobe. This thrombotic lesion does not involve the eallosal gyrus, injuring only the cortex of the paracentral lobule. A considerable portion of the frontal cortex on the orbital surface, extending anterior to the tip of the temporal lobe for approximately 1 cm. is free from injury.
Examination of the le f t thalamus. A study of serial sections of the left thalamus reveals a marked retrograde cell degeneration in the anterior half of the thalamus (fig. 3 ) . The parvicellular part of the nucleus medialis dorsalis is completely degenerated in its entire extent.

T H A U M O - C O R T I C A L PROJECTION 9
The gliosis in this nuclear mass is well marked throughout. In the lateral portion of the ventral and lateral nuclei at the level of the rostral end of the center median nucleus is a linear zone of retrograde cell degeneration which increases in size as it extends anteriorly. Just in front of the rostral end of the center median nucleus the de- generation extends medially to involve almost the entire nucleus ventralis lateralis pars dorsalis. Still more anteriorly, the entire
. . . . . , , . . ! . . . . . . . .
, .. .. . : . . . . .. . . '. . . . . . . .
.. ~
Fig. 3 Serial section8 of the thalamus to show the retrograde cell degeneration in case 1. The stippling in this and all the following drawings of the thalamne repreaents degeneration.
ventral portion shows retrograde cell degeneration. A few normal cells may be found in any section in this nucleus and more anteriorly these are grouped in clumps. Toward the rostral end of the thalamus the degeneration is less severe and near its anterior extremity the cellular architecture is practically normal. The anterior thalamic nuclei show some cellular change. The anterodorsal nucleus is normal throughout and the anteroventral shows few changes in its dorsal

10 A. EARL WALKER
moiety, but the cells of the ventral part of that nucleus and the antero- medial nucleus are very pale staining and small, although there is no gliosis present.
It is evident from this experiment that the fwntal lobe receives fibers mainly from two nuclei of the thalamus, the nucleus medialis dorsalis and the nucleus ventralis lateralis which constitutes most of the anterior half of the lateral nuclear group. To determine the projection from each of these nuclei another case is presented in which there was complete degeneration of the parvicellular part of the medial dorsal nucleus and very little degeneration within the nucleus ven- tralis lateralis.
65V 575 Z!O 425 350 275 200 125
Fig. 4 The hemisphere and serial sections of the cerebral cortex of case 2 (A) to show the site and amount of cortical damage.
Case 2 ( A ) . On April 15, 1932, the cortex of the frontal lobe an- terior to the left precentral gyrus was removed with a sharp knife in an immature macaque monkey (species unknown). The animal was sacrificed July 10, 1932, and the brain removed.
Grossly the lesion involves the lower port,ion of the frontal cortex extending from 4 mm. anterior to the central sulcus (just posterior to the arcuate) to the frontal pole. It reaches from the horizontal limb of the precentral sulcus to the lateral margin of the hemisphere. The lesion appears as several shallow grooves, the main one outlining the arcuate sulcus, and another within the angle of its limbs. Anterior to the horizontal limb of the precentral sulcus, the lesion follows the superior margin of the inferior frontal sulcus, its upper edge being about 7 mm. from the sagittal fissure. The lesion appears superficial throughout (fig. 4) . Microscopically i t extends from the inferior surface of the frontal pole to just behind and above the arcuate sulcus.

THALAMO-CORTICAL PROJECTION 11
Anteriorly the lesion involves the white matter for 3 to 4 mm. but in its posterior half it is practically confined to the cortex. It involves most of the cortex on the lateral surface of the hemisphere inferior to the inferior frontal and horizontal limb of the precentral sulci.
............ ....... I
;,; :;. .; . .:.,: .*:!. . .... ........ . . .
: ....... -f;v L GM ', ., . . . . . . . .
. . . ........ . . . . . . .
. . . . . . . . .
a k;
. .
..... .? .. .Y@ r . . . . . . . . . . . . . . . . . . . . ' , .. '.. ....... i ...... .I 3 j
v,. ;
Fig. 5 Serial sections of the thalamus to show the extent of the cell degeneration in caee 2.
The lower part of area 9, the upper part of area 10, area 8, and a small portion of the middle of areas 6 and 4 are destroyed by the lesion.
Examinatwrr of the l e f t thalamus (fig. 5 ) . Beginning in the center of the nucleus dorsalis at its caudal extremity there is an area of well marked and almost complete neuronic degeneration. Anteriorly

12 A. EARL WALKER
the degeneration involves the entire nucleus leaving only a row of normal ganglion cells a t the periphery. Only a very occasional normal cell is present in the degenerated area. The retrograde cell change continues into the anterior portion of the nucleus, still involving all of the nucleus medialis dorsalis but becoming somewhat more lateral.
At the rostra1 extremity of the nucleus medialis dorsalis there is a considerable glial increase in the nucleus ventralis lateralis pars dorsalis along its medial border with the nucleus paracentralis. Al- though a few ganglion cells show degenerative changes, there are many normal neurons present and nowhere is there an area of com- ple te degeneration.
This experiment has shown the projection of the medial dorsal nucleus to the frontal lobe. The nucleus ventralis lateralis must then project to the remainder of the frontal cortex. This is in keeping with the experimental work of Le Gros Clark and Boggon ( '35), and with the findings of other cases in this series. One other case with a frontal lobe lesion is presented because of an accidental injury to the hypo- thalamus, which produced retrograde cell degeneration in the thalamus.
Case 3 ( N - 1 4 ) . On April 6, 1934, on an immature male Macaca mulatta, an attempt was made to make a small lesion in the optic chiasm. In so doing the orbital surface of the right frontal lobe was contused considerably. Following this the cortex in the right central sulcus was removed with the electric cautery. The animal was sacri- ficed May 24, 1934.
The first occupies the site of the central sulcus, and extends downward to within 3 mm. of the sylvian fissure. It is 1 cm. in width and involves a portion of both the pre- and postcentral gyri. Superiorly it does not quite reach the midline and appears to be superficial. The second lesion occupies the lateral and orbital surface of the frontal lobe. It is 2.5 em. in length and extends from 1 to 2 mm. of the sylvian' fissure well anteriorly. A considerable amount of the posterior portion of the orbital surface of the frontal lobe appears to have been damaged.
Microscopic examination of serial sections of the cerebral cortex reveals that the cortex of the central sulcus has been removed almost from the sylvian fissure to within 1 em. of the midline. The lesion extends anteriorly into area 4 and into area 5 posteriorly, but areas 1, 2 and 3 have been completely removed from the lower two-thirds of the central sulcus. It reaches 5 mm. into the white matter in the depth of the sulcus but does not involve any subcortical gray mass. On the inferior surface of the frontal lobe is another extensive injury. Be- ginning anteriorly with a small area of damage to the lateral margin of the frontal lobe it becomes larger to involve most of the cortex
There are two gross lesions of the cerebral cortex (fig. 6).

THALAMO-CORTICAL PROJECTION 13
on the lateral surface of the frontal lobe at the level of the rostrum of the corpus callosum. Posterior to this, the lesion is located only on the base of the frontal lobe. Here it involves the orbital convolution and extends somewhat deeply into the subcortical gray matter. The anterior part of the caudate nucleus has been damaged considerably by the lesion which extends for 0.7 cm. into its substance. A com- munication by way of the damaged area is present between the an- terior horn of the right lateral ventricle and the subarachnoid space.
475-5.50- 625 700 775 050
I 0 0
Fig. ti The hemisphere and serial wetions of the cerebral cortex to show the extent and situation of the lesion in ease 3 (N-14).
More anteriorly the lesion extends in a linear manner vertically to the anterior limb of the internal capsule which lies between the head and body of the caudate nucleus. The antero-inferior part of the body of the capsule has been injured, but the damage is slight. There is no injury to the temporal lobe. If the lesion is followed to its posterior extremity it is seen to end a t the anterior end of the hypothalamus. This region is damaged both anterior and posterior to the optic chiasm. The anterior lesion is a softening involving the preoptic nucleus and extending to the anterior commissure on the right side. At the level of the chiasm the hypothalamus is free from injury, but just posterior to this there is a lesion in the floor of the third ventricle just to the

14 A. EARL WALKER
right of the midline. It is a small injury and does not reach to the rostral extremity of the mammillary bodies.
There is a very ex- tensive degeneration in the thalamus. The nucleus ventralis posterior is almost completely changed except in its most medial and lateral
Examination of the right thalamus (fig. 7 ) .
Fig. 7 Serial sections of the thalamue to show the retrograde cell degeneration in case 3. The insert is a drawing of the orbital eurface of the frontal lobe to show the extent of the lesion.
parts where the cells are normal. The degeneration begins near its caudal extremity and continues past the rostral extremity of the center median into the ventral part of the nucleus ventralis lateralis. The ventral part of the nucleus lateralis posterior shows some slight changes, the dorsal being quite normal. The degeneration there adjoins that in the ventral nucleus.

THALAMO-CORTICAL PROJECTION 15
There is well marked degeneration in the nucleus medialis dorsalis. Beginning in the center of that nucleus at its posterior extremity, it becomes larger anteriorly, although a fringe of normal cells is present laterally until the anterior third of the nucleus is reached, at which point the degeneration involves the entire nucleus. I t continues thus throughout the remainder of both the magno- and parvicellular parts of the nuclear mass.
The nuclei of the midline show evidence of retrograde cell de- generation. In most of its extent the nucleus paraventricularis pos- terior is slightly degenerated. More anteriorly the posterior part of the nucleus centralis medialis shows degeneration. In the posterior part of the parataenial nuclei the cells show a simple atrophy. The nucleus medialis ventralis shows a marked degeneration in its entire extent. The nucleus paraventricularis anterior shows no change.
The medial group of nuclei also shows changes. The center median is stained palely, but there is no definite evidence of cell degeneration. The nuclei parafascicularis, paracentralis and centralis lateralis ap- pear normal but the nucleus submedius is markedly degenerated. Ex- cept the medial part of the anteromedial nucleus, the cells of which are pale and small, the anterior nuclei appear normal.
The interpretation of the retrograde cell degeneration in this case requires some consideration. The changes in the ventral and lateral nuclei are due to the injury to the cortex of the central sulcus (see the discussion of the projection of the cortex to the parietal lobe). From the preceding experiments the damage to the cortex in the region of the inferior precentral and inferior frontal sulci may be considered the cause of the cellular changes in the nucleus medialis dorsalis pars parvicellularis. The degeneration in the nuclei of the midline remains to be explained. This is the only case in which such degeneration was found. That it is not due to the injury of the caudate nucleus may be assumed, since the head of that nuclear mass was damaged in both decortications without causing changes in these nuclei. It seems certain then that the retrograde cell degeneration in the nuclei of the midline is due to the involvement of the hypothalamus; and hence that the projection of these nuclei is to the hypothalamus.
Summary
It is apparent, then, that the thalamic projection to the frontal lobe is derived from the nucleus medialis dorsalis pars parvicellularis, the anteromedial nucleus, and the nucleus ventralis lateralis. The projection from the medial nucleus must be to the cortex in the region of the inferior frontal and

16 A. EARL WALKER
inferior precentral sulci. The anteromedial nucleus probably has connections to the base of the frontal lobe and to the gyrus cinguli, although a definite statement on this point is not war- ranted from the evidence presented here. The nucleus ven- tralis lateralis, which consists of the anterior half of the lateral and ventral nuclear groups, excepting the most rostra1 portion, the nucleus ventralis anterior, sends fibers to the remainder of the frontal cortex.
THALAMIC PROJECTION UPON THE PARIETAL LOBE
Since the demonstration by Hitzig and Fritsch (1870) of the electrical excitability of the precentral gyrus, it has been assumed that the cortex posterior to the central fissure is somato-sensory in function. However, the exact thalamic connections of the postcentral gyrus and of the parietal lobe have never been clearly demonstrated. To do this using the method of retrograde cell degeneration is rather simple. First, to determine the sum total thalamic fibers to the parietal lobe a case is presented in which the parietal cortex was com- pletely removed.
On January 7, 1935, the cortex of the left frontal lobe of a Macaca mulatta monkey was removed with the electric cautery. Exactly 1 month later the right parietal lobe waa entirely removed. The animal was sacrificed March 28, 1935.
Grossly there is a large lesion of the right parietal lobe extending from the central sulcus posteriorly and inferiorly to the sylvian fissure. Superiorly it extends 5 mm. downward on the medial surface of the hemisphere. There is also a small lesion in the precentral gyrus in the lower arm area. Microscopically the latter lesion is seen to be relatively small and in many sections not even to extend through the cortex. Beginning near the inferior extremity of the central sulcus is the larger lesion which undercuts that sulcus and destroys the cortex of the postcentral gyrus. This lesion extends from the posterior lip of the cehtral sulcus to the upper lip of the sylvian &sure, and be- tween these two it extends into the underlying white matter approxi- mately 5 mm. The lesion extends superiorly to the sagittal sulcus but does not involve the cortex of the medial surface of the hemisphere. The apparent gross involvement was due merely to the ablation of the cortex on the convexity (fig. 8).
Case 4 (E-2-35-22).

THALAMO-CORTICAL PROJECTION 17
Fig.8 "he hemisphere and serial sections of the cerebral cortex in w e 4 (E-2-36 R) to show the situation and extent of the damage.
Fig. 9 Serial sections of the thalamus to show the situation and extent of the retrograde cell degeneration in case 4.

18 A. EARL WALKER
Exa.minatwn of the right thalamus. There is a very marked de- generation of the lateral and ventral thalamic nuclei (fig. 9) . The remainder of the thalamic nuclei are normal. Beginning at the rostral extremity of the pulvinar at about its center, the degeneration rapidly becomes larger to involve the lower two thirds of the lateral nuclear mass (mainly ventral nucleus). Shortly after the median center is reached the degeneration extends upward to include the entire nucleus lateralis posterior and downward to include all of the ventral nu- cleus other than the nucleus ventralis posteromedialis pars medialis. Followed more anteriorly the degeneration becomes smaller again, and at the rostral extremity of the median center it occupies the middle two-thirds of the lateral nuclear mass. Just anterior to this it dis- appears, the last vestige being present in the lateral part of the nucleus ventralis lateralis.
The retrograde cell degeneration in the nucleus lateralis posterior and posterior half of the ventral nucleus must be due to the interrup- tion of fibers from these nuclei. It is therefore evident that their con- nections are to the cortex of the postcentral gyms and the parietal lobe. There is, however, a much more localized projection, which will be demonstrated in the following case.
SO I25 200 250 300 350 400 450
Fig. 10 The hemisphere and surial sections of the cortex to show the situation and extent of the lesion in case 5 (N-13R).
Case 5 ( N - 1 3 R ) . On March 29,1934, on a Macaca mulatta monkey, bilateral bone flaps were reflected. A single linear incision was made along the posterior border of the right central suleus in a n effort to damage area 3 more or less selectively. On the opposite hemisphere the cortex of the temporal lobe was removed. The animal was sacri- ficed May 24, 1934.
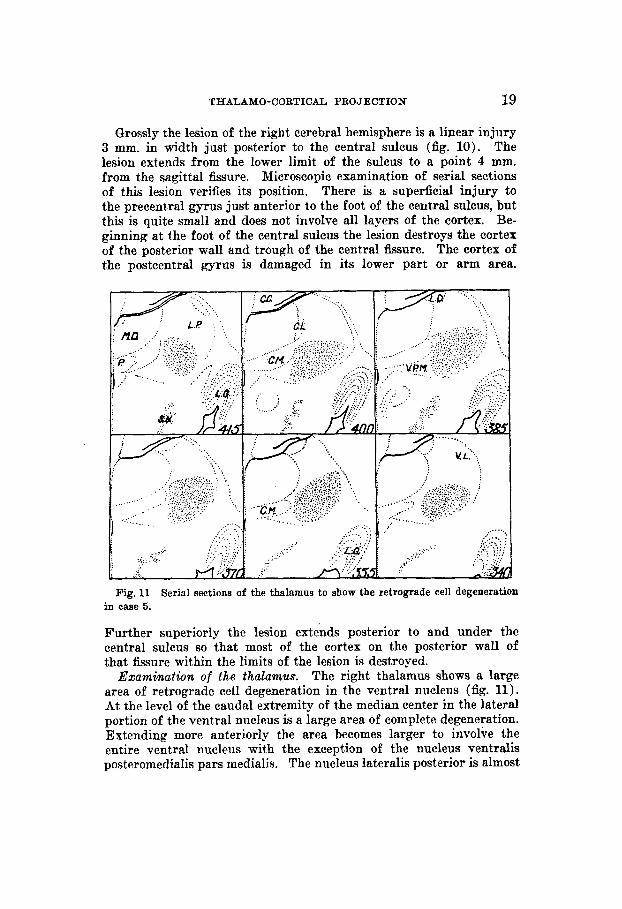
THALAMO-CORTICAL PROJECTION 19
Grossly the lesion of the right cerebral hemisphere is a linear injury 3 mm. in width just posterior to the central sulcus (fig. 10). The lesion extends from the lower limit of the sulcus to a point 4 mm. from the sagittal fissure. Microscopic examination of aerial sections of this lesion verifies its position. There is a superficial injury to the precentral gyrus just anterior to the foot of the central sulcus, but this is quite small and does not involve all layers of the cortex. Be- ginning at the foot of the central sulcus the lesion destroys the cortex of the posterior wall and trough of the central fissure. The cortex of the postcentral gyrus is damaged in its lower part or arm area.
Fig. 11 in case 5.
Further superiorly the lesion extends posterior to and under the central sulcus so that most of the cortex on the posterior wall of that fissure within the limits of the lesion is destroyed.
Examination of the thalamus. The right thalamus shows a large area of retrograde cell degeneration in the ventral nucleus (fig. 11). At the level of the caudal extremity of the median center in the lateral portion of the ventral nucleus is a large area of complete degeneration. Extending more anteriorly the area becomes larger to involve the entire ventral nucleus with the exception of the nucleus ventralis posteromedialis pars medialis. The nucleus lateralis posterior is almost
Serial sections of the thalamus to show the retrograde cell degeneration

20 A. EARL WALKER
entirely spared. At about the middle of the median center the de- generation is most extensive and there are practically no normal cells to be found in the ventral nucleus. The degeneration decreases further anteriorly and disappears at the level of the rostra1 extremity of the median center. There is no degeneration in the lateral nucleus, except perhaps at its ventral margin, nor in any other thalamic nucleus.
From a perusal of this case it is apparent that the cortex of the trough and posterior lip of the rolandic fissure receives practically all of the fibers from the nucleus ventralis posterior. In this case the medial portion of this nucleus was intact. What, then, is its pro- jection? In a previous paper evidence was adduced that this part of the thalamus projects to the face area of the cerebral cortex, which
150 22.5 300 375HO Us bm 673 Is0 a 5
Fig. I2 The hemhphere and serial sectiona of the cortex to ahow the situation and extent of the lesion in caae 6 (N-12 a).
in the present case was not injured. To show this the following case is presented.
On June 1, 1933, bilateral bone flaps were re- flected on a Macaca mulatta monkey. On the right hemisphere the frontal and parietal operculum region and a small area in the pos- terior temporal region were damaged with a sharp scalpel. A small lesion was made in the first temporal convolution on the left side. The animal was sacrificed July 28, 1933.
Gross examination of the cortex of the right hemisphere reveals lesions of the operculum and second temporal convolution (fig. 12). That of the opercular region extends from a point 3 to 4 mm. posterior to the inferior precentral sulcus at the level of the anterior central sulcus posteriorly to a point just caudad to the anterior extremity of
Case 6 (N-12-R).

THALAMO-CORTICAL PROJECTION 21
the rolandic fissure, the cortex of the margin of which it involves. Serial sections show that it is quite superficial except for a single linear penetrating lesion just anterior to the rolandic fissure which extends 4 mm. into the white matter. The lesion involves areas 4 and 3.
In the middle third of the second temporal convolution is another lesion 5 to 6 mm. wide. Serial sections show that it is quite superficial, damaging only the cortex and not extending into the subjacent white matter.
Study of the serial sections of the right thalamus shows two definite zones of degeneration (fig. 13).
Emmination of the right thalamus.
Fig. 13 Serial sections of the thalamua to ahow the retrograde cell degeneration in cme 6.
The first i$ situated in the inferior portion of the posterior p a d of the pulvinar in the nucleus pulvinaris medialis, pars inferior. It is not very extensive and it disappears shortly after the cortico-tectal tracts appear. The second area of the retrograde cell degeneration is in the posterior portion of the nucleus ventralis posteromedialis, pars inferior. This zone of the cell change is quite extensive and more anteriorly involves the superior part of the nucleus ventralis poatero- medialis dorsalis. The degeneration is complete, that is, all ganglion cells in the area are degenerated. This zone extends practically to the rostra1 extremity of the nucleus centrum medianum.
22 A. EARL WALKER
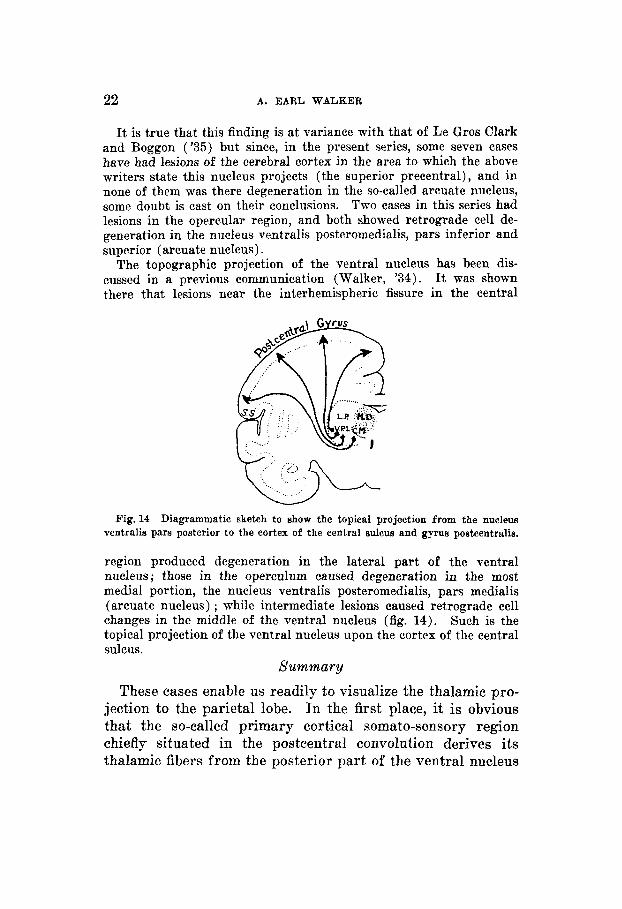
It is true that this finding is a t variance with that of Le Gros Clark and Boggon ('35) but since, in the present series, some seven cases have had lesions of the cerebral cortex in the area to which the above writers state this nucleus projects (the superior precentral) , and in none of them was there degeneration in the so-called arcuate nucleus, some doubt is cast on their conclusions. Two cases in this series had lesions in the opereular region, and both showed retrograde cell de- generation in the nucleus ventralis posteromedialis, pars inferior and superior (arcuate nucleus).
The topographic projection of the ventral nucleus has been dis- cussed in a previous communication (Walker, '34). It was shown there that lesions near the interhemispheric fissure in the central
Fig. 14 Diagrammatic sketch to show the topical projection from the nucleus ventralis pars posterior to the cortex of the central sulcus and gyrus postcentralis.
region produced degeneration in the lateral part of the ventral iiucleus; those in the operculum caused degeneration in the most medial portion, the nucleus ventralis posteromedialis, pars medialis (arcuate nucleus) ; while intermediate lesions caused retrograde cell changes in the middle of the ventral nucleus (fig. 14). Such is the topical projection of the ventral nucleus upon the cortex of the central sulcus.
Summary
These cases enable us readily t,o visualize the thalamic pro- jection to the parietal lobe. In the first place, it is obvious that the so-called primary cortical somato-sensory region chiefly situated in the postcentral convolution derives its thalamic fibers from the posterior part of the ventral nucleus

THALAMO-CORTICAL PROJECTION 23
with, perhaps, a few fibers from the ventral portion of the lateral nucleus. The projection from the medial part of this is to the cortical face area, and that from the lateral part to the leg area, with the intermediate parts of the ventral nucleus projecting to the arm and trunk areas. The parietal cortex proper (Brodmann's areas 5 and 7), on the other hand, repre- sents the nucleus lateralis, pars posterior.
THALAMIC PROJECTION W O N THE OCCIPITAL LOBE
The cortex of the occipital lobe is intimately connected with the visual projection which has been carefully worked out by Brouwer ('27) and by hie co-workers, and in detail by Poliak ( '32). While occasional lesions have been made in the occipital lobe in this work, nothing new has been found concerning the projection of the lateral geniculate body on the striate area. As the projection of the annectant cortex (areas 18 and 19) was not studied by these former workers, their thalamic connections will be presented here with that of the temporal cortex.
THALAMIC PRO.TECTION UPON THE TEMPORAL LOBE
Relatively little has been written upon the thalamic projec- tion to the temporal lobe. True, the connections of the medial geniculate body have received some attention, but they com- prise a complex system quite separate from the thalamus To determine the total number of cells projecting to the temporal lobe a complete ablation of the temporal and annec- tant cortex will be considered primarily.
Case 7 (N-13 L ) . On March 29, 1934, on an immature Macaca mulatta, bilateral bone flaps were reflected. A linear incision was made just posterior to the right central sulcus in an effort to damage area 3 more or less selectively. On the left side the cortex of the left temporal lobe was removed as far posteriorly the temporo-occipital and parieto-occipital Gssures. The animal was sacrificed May 24,1934.
The left cerebral hemisphere shows a large temporal ablation (fig. 15). It extends from the tip of the temporal lobe posteriorly to 3 mm. behind the simian sulcus and from the sylvian fissure superiorly to the
FRE JOUBXAL or c o m r m m NEUPOLOQY, VOL 64. NO. 1

24 A. EARL WALKER
second temporal convolution inferiorly. A small linear incision crosses the sylvian fissure to run into the postcentral gyrus for a distance of 10 mm. A study of serial sections of this hemisphere shows that almost all of the cortex of the first and the second temporal convolution has been removed. The posterior wall of the sylvian fissure is almost en- tirely preserved, however. The injury to the postcentral gyrus is quite small. Posteriorly the lesion extends backward into the striate area just below the external calcarine fissure for about 5 mm. It involves the subjacent white matter in most of its extent but does not damage any subcortical gray matter.
Fig. 15 The hemisphere and serial sections of the cerebral cortex to show the situation and extent of the damage in case 7 (IT-13 L).
Examination of the l e f t thalamus. The left thalamus reveals a con- siderable amount of degeneration in the pulvinar (fig. 16) . The in- ferior two-thirds of the caudal extremity of the pulvinar are coin- pletely degenerated ; further anterior this degeneration becomes limited to the nucleus pulvinaris lateralis, pars inferior. Still further anteriorly the degeneration involves the inferior nucleus, and disap- pears when the lateral geniculate body replaces the lateral portion of the pulvinar. The posterior portion of the medial geniculate body is markedly degenerated as is also the lateral part of the lateral genicu- late body. The remainder of the thalamic nuclei are intact.
In this case most of the cortex of the temporal lobe and of the periparastriate areas (fields 18 and 19 of Brodmann) has been re- moved. A very extensive retrograde cell degeneration is present in the pulvinar. To determine which portion of the ablated cortex receives the fibers from these cells, further experimentation is necessary.

TEIALAMO-CORTICAL PROJECTION 25
Case 8 (N-11 R ) . On June 1, 1933, bilateral bone flaps were re- flected on an immature male Macaca mulatta. On the left side a lesion was made in the posterior portion of the parietal lobe. The tip and the base of the right temporal lobe were then removed with the electric cautery. A small lesion was made in the posterior part of the orbital surface of the right frontal lobe. On July 21, 1933, the animal was sacrificed.
1 *,,6i I ....... .,:.;. . -. . .. .: . .. . . ... ~. ':, . .. ._ . . . ..._ i., .. . .: . ..... . . r . *
Fig. 16 The serial sections of the thalamus to show the extent of the retrograde cell degeneration in case 7.
TWO lesions are present in the right hemisphere (fig. 17). Grossly the first appears as a linear area of decortication in the inferior frontal convolution. It extends along the inferior margin of the lateral surface of the hemisphere, from the anterior tip of the inferior limb of the precentral sulcus to a point in the postcentral convolution a few millimeters behind the central sulcus. That sulcus was not in- volved by the lesion, but the lower portion of the damage involved the cortex of the sylvian fissure. Microscopic examination of serial sections of the cortex shows that in addition to this large lesion there is a smaller one in the orbital gyrus. This latter injury involves only a relatively small amount of cortex, although it extends from the anterior level of the lesion in the inferior frontal convolution to the t ip of the temporal lobe. It is a narrow lesion being only a few

26 A. EARL WALKER
millimeters in width, but extends 2 or 3 mm. into the subjacent white matter. The main damage to the frontal lobe begins anteriorly as a superficial injury to the outer two or three layers of the cortex, but soon becomes deeper destroying the cortex and a small amount of subcortical white matter of the inferior frontal gyrus. Just anterior to the tip of the temporal lobe it is 5 mm. in width and extends 3 mm. into the white substance. At the anterior extremity of the head of the caudate nucleus there is a punctured wound piercing deeply into the white matter and almost reaching the gray substance of the caudate nucleus. Further posteriorly the lesion merges with the injury in the temporal lobe. The posterior portion involves the cortex of the sub- central gyrus, but does not damage the walls of the central sulcus. Areas 6 and 10 have been injured.
JD ub3Do 27s 35D 415 SO0 575 CSQ XU 775
Fig.17 The hemisphere and serial sections of the cerebral cortex in case 8 (N-11 R) to show the extent of the damage.
The second large lesion has injured the t ip of the temporal lobe and the superior and middle temporal gyri. It extends along the in- ferior margin of the sylvian fissure to the anterior extremity of the posterior marginal gyrus. It is a broad deep damage to the first and second temporal convolutions, but posteriorly tapers to a shallow and narrow lesion in the first temporal convolution. Study of serial sections shows that the lesion has destroyed the cortex of the entire t ip of the temporal lobe and several millimet.ers of the underlying white matter in the first and second temporal convolutions. The lesion ex- tends to the wall of the sylvian fissure but does not damage that struc- ture. Inferiorly it involves t.he anterior or upper half of the middle temporal gyrus. Posteriorly the surface of the first temporal convo- lution including its upper or sylvian margin has been ablated. Areas 21 and 22 and a small part of area 19 are involved by the lesion.

THALAMO-CORTICAL PROJECTION 27
Examination of the right thalamus. Areas of cell degeneration are found in the pulvinar only (fig. IS). Beginning near the posterior extremity of the pulvinar in the nucleus pulvinaris medialis, pars intermedia, is a broad, oblique, linear zone of complete degeneration paralleling the nucleus reticularis. Followed more anteriorly this zone is seen to become broken into several discrete areas lying more superiorly and medially in the pars intermedia of the medial nucleus of the pulvinar. Followed still more anteriorly these areas, now two discrete masses of the degenerated cells, are seen to lie just lateral and
Fig. 18 in caae 8.
superior to the nucleus limitans, the one above the other. The de- generated area ends at the caudal end of the nucleus medialis dorsalis, and just anterior to the point where the cortico-tectal tract divides pulvinar into a superior and an inferior portion. There is also an irregular dif€usely scattered degeneration in the nucleus ventralis lateralis, pars lateralis, beginning at the level of approximately the rostra1 end of the median center. In no place is the gliosis intense, nor is there a marked decrease in the number of cells. The changes extend forward to approximately the middle of this nucleus.
In this experiment there is an extensive injury to the temporal cortex, but a very small amount of degeneration in the thalamus. It would appear, then, that the thalamic projection to the temporal lobe
Serial sections of the thalamus to show the retrograde cell degeneration

28 A. EARL WALKER
proper is quite small, and that the greatest part of the fibers from the pulvinar pass to the periparastriate areas. A s a further proof of this a case with an occipital lobectomy is included in this series. This animal was used by Dr. Heinrich Kliiver in his study of the physiology of vision. It is through his kindness that the brain was made available for the present investigation.
On March 30, 1933, the occipital lobe was removed from the right hemisphere of a Macaca rnulatta monkey. The animal was sacrificed May 16, 1933.
Grossly the entire right occipital cortex has been removed posterior to the first temporal sulcus on the convexity of the hemisphere and
Case 9 ( M 37).
625 6 7 5 725 77.5 850
Fig. 19 The hemisphere and serial aections of the cerebral cortex to show the extent and situation of the occipital decortication in case 9 (N Y).
behind the temporo-occipital fissure on the lateral surface (fig. 19). On the medial surface the line of ablation extends inferiorly from the lateral parietal occipital sulcus (simian) to the anterior portion of the calcarine &sure, leaving intact only its anterior portion.
Microscopic examination of serial sections of the cortex confirms the gross findings. A small part of the anterior portion of area 17 lying within the ealcarine fissure remains intact, but this is only present in about fifty sections. The lesion has removed most of Brod- mann’s fields 17 and 18, and a considerable portion of the field 19.
The degeneration is confined to the pulvinar and the lateral geniculate body, which is completely
Exa.mination of the right thdamus.

THALAMO-CORTICAL PROJECTION 29
degenerated (fig. 20). In the lateral portion of the pulvinar it ex- tends anteriorly from the caudal extremity. Although the degenera- tion is more extensive inferiorly, it reaches almost to the superior angle of the pulvinar. The inferior nucleus is completely degenerated. At the level of the caudal extremity of the lateral geniculate body the degeneration ends. A marked gliosis and a complete absence of normal nerve cells is noticeable in the involved areas.
Fig. 20 in case 9.
Serial sections of the thalamus to show the retrograde cell degeneration
If the retrograde cell degeneration in this case be compared with that in the case 7 it will be clear that the amount and site of the de- generation is quite similar--except that the changes are more extensive in case 9. Therefore, it must be evident that the degeneration in case 7 is the result of the injury to the cortex of periparastriate areas, and not to the damage done to the temporal lobe proper. This confirms the impression gained from the case 8.
Summary
It is plausible from the above cases that the afferent pro- jection of the thalamus upon the temporal cortex plroper (areas 20,21 and 22) is small or none at all. (This, naturalb,

30 A. EARL WALKER
does not concern the medial geniculate body.) The annectant cortical areas (fields 18 and 19 of Brodmann) receive many fibers from the pulvinar, and in consequence of the occipital injury an extensive retrograde cell degeneration is found in the pulvinar. The medial geniculate body showed degenera- tion only in case 7, but from the cases here its projection cannot be positively asserted. However, from the investigations of Poliak ( '32) and from my own .unpublished investigation, where the method of the retrograde cell degeneration was ap- plied, the medial geniculate body in Macaca mulatta is found to have a point to point projection to an area of koniocortex situated on the superior surface of the first temporal con- volution within the sylvian fissure.
PROJECTION OF THE THALAMIC NUCLEI UPON THE CEREBRAL CORTEX
The afferent connections of the thalamus with the different cortical areas have been presented. From these data it is possible to analyze the projection of each thalamic nucleus. The results of this analysis should allow of a division of the thalamus on a basis of its fiber connections. Conversely it will serve as a means of separating morphologically similar nuclear groups, having different cortical connections.
Anterior group. That the anterior thalamic nuclei have a cortical projection is attested by the degeneration which occurs following hemidecortication. This fact previously has not been unanimously accepted, Dejerine ( '01)' Flechsig ( '20) and Sachs ( '09) denying any cortical connections from this nuclear group. Those who have described such pathways (Monakow, 1895 ; Rlingazzini '13 ; Minkowski, '23-'24 ; and Roussy, '07) mention the frontal lobe as the terminal station of the fibers arising from the anterior nuclei. Le Gros Clark and Boggon ( '32) have studied the projection of these nuclei both with the Marchi and retrograde cell degeneration tech- niques in rats and cats. They found that the anterodorsal and anteroventral nuclei are projected upon the gyrus cinguli, and suggested that the anteromedial cell group might have connections to the cortex of the medial surface of the frontal

THALAMO-CORTICAL PROJECTION 31
lobe, such as has been shown by these experiments. Following a complete ablation of the frontal cortex only the anteromedial nucleus shows changes and, as this nucleus shows very slight alterations following an injury to the orbital surface of the frontal lobe, it would appear likely that it has a small pro- jection to that portion of the frontal cortex. The remainder of its connections are probably with the medial surface of the frontal lobe (gyrus cinguli). In none of the experiments was there any change in the anteroventral or anterodorsal nuclei; these probably project upon the callosal gyrus posterior to the paracentral lobule, as that was the only region not injured in our series of experiments excepting the two hemidecorti- cated cases. This assumption is quite compatible with the findings in the lower mammals (Waller, '34; Le Gros Clark, '32 a).
The nuclei of the midline. There is no evidence that the nuclei of the midline are represented in the cerebral cortex; on the contrary all experimental studies point to their sub- cortical, efferent connections (Munzer and Wiener, '02 ; Waller, '34; Le Oros Clark and Boggon, '33). I n only one case in this series (case 3) was there any retrograde cell de- generation in the mentioned nuclei. In this case a lesion was present in the supraoptic nuclei and retrograde cell degenera- tion was found in the large-celled portion of the nucleus medialis dorsalis, nuclei submedius, paracentralis, posterior centralis medialis, and medialis ventralis. Such connections in lower mammals have been described by many investigators (Rioch, '29; Le Gros Clark, '32 b ; Huber and Crosby, '29). The projection of the remainder of the nuclei of this group cannot be determined from this study.
The medial nuclei. This group consists of the nuclei medialis dorsalis, parafascicularis, paracentralis, centralis lateralis, and median center. For the time being afferent cortical connections can only be established for the first of these. Since degeneration in the nucleus medialis dorsalis, pars parvicellularis, occurred in all those cases in which the frontal lobe was injured and the area common to the three

32 A. EARL WALKER
lesions was the cortex between the inferior limb of the pre- central and the inferior prefrontal sulci, it would seem logical to assume that area as the main cortical projection of the nu- cleus dorsalis medialis. This finding is quite in accord with both the earlier (Monakow, 1895 ; Probst, '00 ; Dejerine, '01 ; Mingazzini, '13 ; Fukuda, '19 ; and Minkowski, '23-'24) and more recent workers (Le Gros Clark and Boggon, '35). 0. Sager ('33) on the basis of four cases with lesions of the fronta lobe concluded that this nucleus did not have much pro- jection to the frontal cortex. However, none of his lesions more than touched the cortex anterior to the inferior limb of the precentral sulcus. The large-celled part of the nucleus medialis dorsalis, which does not have cortical connections, de- generated in consequence of a lesion in the hypothalamic region (case 3) . This &ding is in accord with Le Gros Clark's ('32 b) conceptions of the significance of that nucleus.
The intralaminar nuclei do not show appreciable degenera- tion following hemidecortication. It is true that there are fewer cells in these nuclei after such a procedure than in normal preparations, but there is always a considerable number of normal cells. This probably means that these intralaminar nuclei have interspersed cells, which really be- long to adjoining nuclei. Be it as it may, following hemide- cortication may normal cells are found in these nuclear groups. Particularly is this true of the median center, which appears unchanged in these preparations. From the present work, little can be said regarding the projection of these nudei; they probably have connections with the ventral, lateral and medial thalamic nuclei.
This group is composed of the lateral and ventral nuclei. The lateral nucleus has a distinct dorsal part, the nucleus lateralis dorsalis (La, or disseminated nuclei of Obersteiner ) which does not completely degenerate in consequence of lesions of the cerebral cortex, and should be considered an intralaminar nucleus, probably having intra- thalamic connections.
The lateral group o f nuclei.

THALAMO-CORTICAL PROJECTION 33
The remainder of the lateral and ventral nuclei will be re- ferred to collectively, as the lateral nuclear mass. This may be divided into anterior and posterior moieties. The anterior half has no clear morphological division into ventral and lateral portions and so has been called the nucleus ventralis lateralis (Walker, in press). Following a complete hemide- cortication the rostra1 extremity of the lateral nuclear mass shows little alteration, while the more posterior portions are markedly degenerated. Hence, it is logical to assume that the anterior extremity does not send fibers to the cortex. This part also can be distinguished on morphological grounds and is termed the nucleus ventralis anterior (Griinthal, '34 ; Aronson and Papez, '34). The projection from this part of the thalamus cannot be determined from the present ex- perimentation. The nucleus ventralis lateralis, however, de- generates following the removal of the frontal cortex, but not after an ablation of the cortex anterior to the arcuate nucleus (case 2). It is very likely, then, that the projection from these nuclei is to the cortex of the motor and premotor areas. This is quite in agreement with the findings of Le Cros Clark and Boggon ('35), who found degeneration in these nuclei following lesions of Brodmann's areas 4 and 6. Such an assumption is further corroborated by several unpublished cases with lesions similarly situated in which retrograde cell degeneration was present in the nucleus ventralis lateralis. The significance of this thalamic projection to the motor cortex is apparent when the termination of the braohium conjunc- tivum in the thalamus is considered. In the macaque monkey a study of Marchi degeneration after section of the superior cerebellar peduncle shows that this tract ends predominately in the nucleus ventralis lateralis.
The main lateral nucleus (nucleus lateralis posterior) de- generated in consequence to an ablation of the parietal lobe (case 4), but not when only the postcentral gyrus was injured (case 5) . Its projection must then be to the cortex of the areas 5 and 7. This finding is quite in accord with that of Le Gros Clark and Boggon ( '35).

34 A. EARL WALKER
The posterior half of the ventral nucleus, the nucleus ven- tralis posterior, degenerates in consequence of a lesion of the posterior lip of the central sulcus or of the postcentral gyrus. It has not shown any change as the result of a frontal lobe injury, although Le Gros Clark and Boggon (’35) in their case (J) describe marked degeneration in the ‘arcuate’ nucleus from a lesion of the superior precentral convolution. The evidence that the nucleus ventralis posterior, is the termination of the medial fillet and spino-thalamic tracts has been discussed in several recent papers (Walker, ’34 ; Le Gros Clark and Boggon, ’35), and need not be reviewed here ex- cept to say that the consensus of opinion of the investigators and the weight of evidence is in favor of this nucleus receiving the endings of these tracts. There is also evidence of a topical distribution within this nuclear mass. The studies of Wallen- berg (1896) and of Probst (’00) suggest that the central sensory pathways of the trigeminal nerve ends in approxi- mately the medial part of this nucleus. 0. Sager (’33) con- cluded that this part, the ‘arcuate nucleus,’ is mainly con- nected by ascending fibers with the face region of the post- central cortex and slightly with the arm area. More con- clusive evidence of a topical distribution was presented in a previous communication of mine, when it was shown that the cells from the medial to the lateral side of the nucleus pro- jected to the central gyri from the inferior to the superior portions (fig. 14). Le Gros Clark and Boggon (’35) came to a somewhat similar conclusion in spite of the fact that one of their experiments (J) suggested a projection from the medial part of this nucleus (arcuate) to the superior precentral gyrus.
Since the pulvinar entirely degenerates following a complete cortical ablation, one may consider that this large nuclear mass projects entirely to the cerebral cortex. It has also been noted that the pulvinar shows no degeneration after the lesions of the frontal lobe and very slight degeneration when the parietal lobe is extirpated (a degeneration probably due to damage to adjacent cortical areas or to its afferent fibers passing underneath the parietal
The posterior nuclear group.

THALAMO-CORTICAL PROJECTION 35
cortex on their way to the periparastriate area). Lesions confined to the tip of the temporal lobe also cause little or no retrograde cell changes in the thalamus. Hence, the assump- tion that the pulvinar projects upon the periparastriate area (fields 18 and 19 of Brodmann) would seem justified, this more so since Poliak ('32) has shown that lesions confined to the striate area produce retrograde cell degeneration only in the lateral geniculate body. This conclusion is in general accord with the findings of both the more recent and older investiga- tors (Monakow, 1895 ; Minkowski, '23-'24 ; Dejerine, '01 ; Le Gros Clark and Boggon, '35 ; Mettler, '35 ; Poliak, '32).
Summary
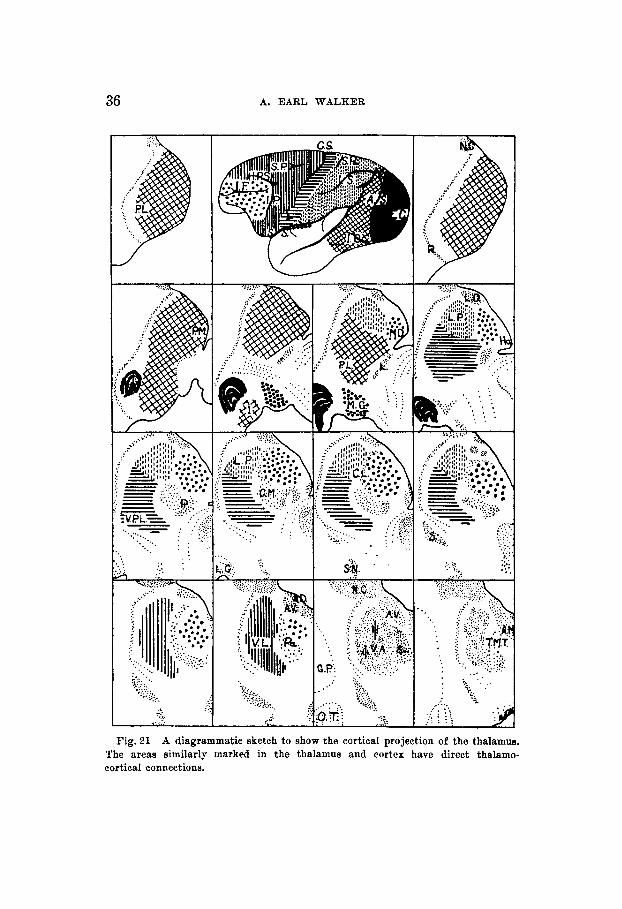
The retrograde cell degeneration in the thalamus following the removal of the cortex of each lobe of the cerebral hemi- sphere in Macaca mulatta has been presented (fig. 21). Fol- lowing a frontal lobe decortication the small-celled part of the medial dorsal nucleus and the nucleus ventralis lateralis degenerated. A lesion between the limbs of the arcuate sulcus caused practically complete degeneration in the dorsomedial nucleus thus demonstrating its cortical projection. A parietal decortication caused retrograde cell degeneration in the nuclei ventralis, pars posterior, and nucleus lateralis, pars posterior. A lesion of the postcentral convolution undermining the central sulcus produced a change in the posterior ventral nucleus, from which it may be deduced that the latter is represented in the cortex of the postcentral gyri ; whereas the posterior lateral nucleus is represented in the parietal cortex proper. A decortication of the temporal lobe caused very slight degeneration in the pulvinar. An occipital lobectomy removing the striate area, simultaneously considerably dam- aging the annectant periparastriate areas, produced com- plete degeneration of the lateral geniculate body and a marked degeneration in the pulvinar. That the latter changes were due to the involvement of the periparastriate cortex alone was shown by another case in which there was but a slight injury to the striate area (field 17).
36 A. EARL WALKER
... 4.'
Fig. 31 A diagrammatic sketch to show the cortical projection of the thalamus. The areas similarly marked in the thalamus and cortex have direct thalamo- cortical connections.

THALAMO-CORTICAL PROJECTION 37
CONCLUSIONS
1. The anterior thalamic nuclei project to a small part of the orbital surface of the frontal lobe, and to the gyrus cinguli posterior to the paracentral lobule.
2. The small-celled part of the medial dorsal nucleus sends fibers to the cortex around the inferior precentral and inferior prefrontal sulci. The large-celled portion connects with the hypothalamus.
3. The nuclei of the midline have connections with the supra- optic nuclear groups, and are not likely to possess afferent connections with the cerebral cortex.
4. The intralaminar nuclei have no afferent cortical con- nections. This includes also the median center.
5. The nucleus ventralis, pars anterior, has no afferent cortical connections, but the remainder of the anterior half of the lateral nuclear mass projects upon the frontal cortex exclusive of the area anterior to the arcuate sulcus.
6. The nncleus ventralis posterior, projects entirely upon the cortex of the central sulcus and the postcentral convolu- tion (areas 1, 2, 3 of Brodmann).
7. The nucleus lateralis posterior, sends fibers to the parietal cortex proper (areas 5 and 7 of Brodmann).
8. The pulvinar projects upon the periparastriate cortex (areas 18 and 19 of Brodmann).
9. The temporal cortex (proper) has few or no thalamic con- nections except from the, medial geniculate body.
The stimulus for this work was given by Dr. Stephen Poljak to whom I wish to express my gratitude for his many kind criticisms and assistance, also to Dr. Percival Bailey for helpful suggestions and constant encouragement. I would also like to make grateful acknowledgment of technical as- sistance to Miss Ruth Erichson and Miss Gladys Ruhland, but for whose perseverance in cutting and staining thousands of sections, this work would have been impossible.

38 A. EAFtL WALKER
LITERATURE CITED
AEONSON, L. R. AXD J. W. PAPEZ Thalamic nuclei of Pithecus (Macacus) rhesus. 11. Dorsal thalamus. Arch. Neur. and Psych., POI. 32, pp. 27-44.
BIELSCEOWfiKY, M. 1932 Histopathology of nerve cells; in “Cytology and cellular pathology of the nervous syNtem,” edited by Wilder Penfield. New York. Paul B. Hoeber, vol. 1, pp. 145-188.
Anatomical, phylogenetical, and clincal studies on the central nervoue system. Baltimore, Williams and Wilkina, 67 pp.
1932 a An experimental study of the thalamic connec- tions in the rat. Phil. Trans. Roy. 8oc. London, aeries B, vol. 232,
1932b The structure and connections of the thalamus. Brain, vol. 65, pp. 408-470.
CLARK, W. E. LE QEO6 AND H. R. B o a a o ~ 1932 On the connections of the an- tenor nucleus of the thalamus. J. Anat., vol. 67, pp. 215-226. 1933 On the connections of the medial cell groups of the thalamus.
Brain, vol. 56, pp. 83-98. 1935 The thalamic connections of the parietal and frontal lob-
of the brain in the monkey. Phil. Trans. Roy. Soe. London, Series B,
CROUCH, R. L. 1934 The nuclear configuration of the thalamus of Macacus rhesus. J. Comp. Neur., vol. 59, pp. 451-485.
D E J ~ N E , J. 1901 Anatomie des eentrea nerveur. Paris, J. Rueff, 720 pp. FLECHBIG, P. 1920 Anatomie des menscblichen Qehirns und Ruekenmarks auf
myelogenetiseher Grundlage. Leipzig, Q. Thieme, 121 pp. FEZTfiCH, 0. AND E. H~Imro 1870 Ueber die elektrische Erregbarkeit des Groee-
hirns. Arch. f. Anat. Physiol. u. wisaen. Med., Bd. 37, 8. 300-332. I%JxUDA, T s ~ w E 1919 Ueber die faseranatomischen Beziehungen zwischen
den Kernen dea Thalamus OptiCUB und den frontalen Windungen (Froqtalregion) dea Menwhen. Schweiz. Arch. f . Neurol. u. Peyeh., Bd. 5, 8. 325-377.
GB~~NTHAL, E. 1934 Der Zellbau im Thalamus der &uger und dee Menschen. J. f. Paych. u. Neurol., Bd. 46, S. 41-112.
HUBE~, G. C. AKD E. C. WBBY 1929 Somatio and visceral connections of the diencephalon. Arch. Neur. and Psych., vol. 22, pp. 187-229.
METFLEE, F. A. 1933 The brain of Pitliecua rheeus (M. rheeus). Am. J. Phys. Anthrop., vol. 17, pp. 309-331.
Corticifugal fiber connections of the cortex of Macaca mulatta. The occipital region. J. Comp. Neur., vol. 61, pp. 221-256.
MINGAZZINI, G. 1913 Ueber die Beteiligung beider Hirnhemispharen an der Funktion der Bpraehe (gleichzeitig ein pathologiech-anatomischer Beitrag zum Btudium einiger Stirnformationen). Folia Neurobiologica, Bd. 7, 8. 1-66.
MINKOWSKI, M. 1923-1924 Etude Bur les connexions anatomjques dea circonvo- lutions rolandiques, paribtales, et frontales. Bchweiz. Arch. f. Neur. u. Psych., Bd. 12, 8. 22-268; Bd. 14, 8. 255-278; Bd. 15, 8. 97-132.
1934
BEOUWER, B. 1927
CLABK, W. E. LE Qws
pp. 1-28.
V O ~ . 224, pp. 313-369.
1935

THALAMO-CORTICAL PROJECTION 39
MONAKOW, C. 1895 Erperimentelle und pathologinch-anatomische Unterauchung iiber die Haubenregion, den Sehhiigel und die Regio eubthalamica. Arch. f. Paych., Bd. 27, €3. 1-128, 386-478.
1902 Das Zwiachen- und Mittelhim dee Kaninchene und die Beeiehungen dieeer Teile zum iibrigen Centralnervenayattern. Monateachr. f. Paych., Bd. 12, S. 241-279.
POLXAK, 8. 1932 The main afferent fiber ayetema of the cerebral cortex in primates.
PROSST, M. 1900 Experimentelle Unterauchungen iiber die Schleifenendigung, die Haubenbahn, das doraale Iiingabiindel und die hintere Kommiseur. Arch. f. Peych., Bd. 33, 8. 147.
RXOCE, D. M. 1929 8tudiea in the diencephalon of the Carnivora. Part I. The nuclear configuration of the thalamue, epithalamne, .and hypothalamus of the dog and cat. J. Comp. Neur., vol. 49, pp. 1-120.
ROUSSY, 0. 1907 La couche optique (etude anatomique, phymologiqne, et clinique) ; le syndrome thalamique. ThBse, Paria, 349 pp.
SACES, E. 1909 On the structure and functional relations of the optic thalabua. Brain, vol. 32, pp. 95-186.
SAGIEB, 0. 1933 Recherchcm eur la somatotopie sensitive dane le thalamua dea singee, Btudiben par la d t h o d e de la dbgih6reeeence r6trograde. Haarlem de Erven F. Bohn, 139 pp.
WALK- A. E. 1934 The thalamic projection to the central gyri in Macacue rhewe. J. Comp. Neur., vol. 60, pp. 161-184. 1935 The retrograde cell degeneration in the thalamue of Macacw
rhewe following hemidecortication. J. Comp. Neur., vol. 62, pp. 407-419.
M~~NZER, E. AND H. WIENEB
The University of California Preee, Berkeley, 370 pp.
A note on the thalamic nuclei of Macaw mulatta. (In preae.) W-m, A. 1896 Die eecundiire Bahn dea nenmblen Trigeminua. Anat.
WULER, W. H. 1934 Topographical relatione of cortical leeiona to the thalamic Am., Bd. 12, €4. 95-110.
nuclei in the albino rat. J. a m p . Neur., vol. 60, pp. 237-267.
THL JOCIIRTAL OW COXPABA'FXVZ XSUPOMOY. V O L 84, XO. 1