an examination of white-nose syndrome occurrence and
TRANSCRIPT

Western Kentucky UniversityTopSCHOLAR®
Masters Theses & Specialist Projects Graduate School
8-2012
An Examination of White-Nose SyndromeOccurrence and Dispersal Patterns: UtilizingGlobal and Local Moran's I Analysis to Evaluate anEmerging PathogenCelia M. DavisWestern Kentucky University, [email protected]
Follow this and additional works at: http://digitalcommons.wku.edu/theses
Part of the Environmental Health and Protection Commons
This Thesis is brought to you for free and open access by TopSCHOLAR®. It has been accepted for inclusion in Masters Theses & Specialist Projects byan authorized administrator of TopSCHOLAR®. For more information, please contact [email protected].
Recommended CitationDavis, Celia M., "An Examination of White-Nose Syndrome Occurrence and Dispersal Patterns: Utilizing Global and Local Moran's IAnalysis to Evaluate an Emerging Pathogen" (2012). Masters Theses & Specialist Projects. Paper 1194.http://digitalcommons.wku.edu/theses/1194


AN EXAMINATION OF WHITE-NOSE SYNDROME OCCURRENCE AND DISPERSAL PATTERNS: UTILIZING GLOBAL AND LOCAL MORAN’S I
ANALYSIS TO EVALUATE AN EMERGING PATHOGEN
A Thesis Presented to
The Faculty of the Geography and Geology Department Western Kentucky University
Bowling Green, Kentucky
In Partial Fulfillment Of the Requirements for the Degree
Master of Science
By Celia M. Davis
August 2012


iii
ACKNOWLEDGMENTS Many thanks to the numerous researchers and agencies instrumental in providing
the White-Nose Syndrome confirmation dates that made this research possible and whose
thoughtful critiques were intrinsic to its development. Particular thanks are due to Cal
Butchkoski with the Pennsylvania Game Commission and Carl Herzog with the New
York State Department of Conservation for their assistance in assembling the final
dataset and invaluable input to the project. Thanks are also due to Dr. Jason Polk and Dr.
Rick Toomey at Western Kentucky University for their tremendous input in the design,
implementation, and realization of this project. As always, my eternal gratitude goes out
to Pima, Tassie, Tupelo, and the Davis clan for their moral and financial support and
patience.

iv
TABLE OF CONTENTS
LIST OF ILLUSTRATIONS……………………………….…………………...…..... v
ABSTRACT..………………………………………………..………………………... vi
INTRODUCTION ………………………………………………………..…………. 1
LITERATURE REVIEW...……………………………….………...…....................... 5
METHODS...……………………………….………...………………………………. 11
RESULTS…………………………….…………………..…….................................. 15
DISCUSSION...……………………………………….……………………………... 22
APPENDIX: Geomyces destructans Confirmation Date...………………………….. 31
REFEREENCES....………………………………………………………..………...... 35

v
LIST OF ILLUSTRATIONS
Equation 1 Local Moran’s I …………...…………..………………………….……. 13
Figure 1 Frequency Distribution of Month Data………………..…………….......... 14
Figure 2 Spatial Autocorrelation Report, Global Moran’s I....…………….….……. 16
Figure 3 Local Moran’s I p-scores……………………………………….…..……... 17
Figure 4 Local Moran’s I CO-type………………….…….…………………....…… 18
Figure 5 Spatial Autocorrelation Report, Global Moran’s I Variable Date………… 19
Figure 6 Local Moran’s I CO-type, Variable Date…………………………………. 20
Figure 7 Local Moran’s I p-scores, Variable Date……………....………..…….….. 21

vi
AN EXAMINATION OF WHITE-NOSE SYNDROME OCCURRENCE AND DISPERSAL PATTERNS: UTILIZING GLOBAL AND LOCAL MORAN’S I
ANALYSIS TO EVALUATE AN EMERGING PATHOGEN
Celia M. Davis August 2012 38 Pages
Directed By: Dr. Jason Polk, Dr. Chris Groves, Dr. Jun Yan, and Dr. Rickard Toomey
Department of Geography and Geology Western Kentucky University
In this research, a novel approach that utilizes Moran’s I statistical analyses to
examine the spatio-temporal dispersal patterns of the White-Nose Syndrome currently
affecting North American bat species is undertaken to further understand the disease
transmission mechanism(s) of this emerging wildlife epidemic. White-Nose Syndrome
has been responsible for in excess of five million bat deaths to date and has the potential
to alter the ecological landscape significantly; however, due to a variety of factors, little
research has been conducted into the patterns of infection on a national scale. Global and
Local Moran’s I analyses were performed on the spatial-temporal variable of month and
location from the initial outbreak site in order to address the spread of the Geomyces
destructans fungus that causes White-Nose Syndrome. A comprehensive dataset of
outbreak confirmation sites has been compiled and statistical analysis using ArcGIS
reveals a complex pattern of disease dispersion since initial discovery of the disease, and
shows important policy and management implications, in particular the need for more
standardized and rigorous data collection and reporting procedures.

1
Introduction
The small blue-green planet on which we live is a closed system and, within this
closed system, each component part has its role to play in the continued functioning of
the whole. The demands human beings place on the myriad interdependent eco-systems
that surround and envelop us have far-reaching implications not only for our own
survival, but for the almost infinite number of plant and animal species upon which we
depend for the most basic of necessities. As global population growth continues
unchecked, the rate at which finite natural resources are being consumed becomes an
almost exponential equation, and anthropogenic consequences are the inevitable end-
result. In light of this disturbing trend, it seems self-evident that any developments
having the potential to disrupt or diminish existing agricultural production should be of
great concern.
Worldwide population growth and an increasing standard of living in the United
States place an unprecedented strain on available food resources for both domestic
consumption and global export. The Food and Agriculture Organization of the United
Nations estimates that in the past year alone the global economic crisis and continued
population expansion will cause approximately 1.02 billion world citizens to be
undernourished (FAO, 2009). American farmers and agri-business companies meet this
increased demand in food production in a variety of ways, resulting in the use of higher
yield genetically modified crops, increased pesticide usage, and alterations in crop-
growing methods. A significant concern is that pesticide applications have long been
linked to undesirable side-effects, such as contamination of the water table, along with
severe short-term and long-term risks to humans directly exposed to the chemicals

2
currently available for crop pest control. Along with the costs (both financial and human-
health related) of extensive pesticide use, there is also a well-demonstrated tendency for
natural pests to develop resistance to chemicals over ever-shorter periods of time. In
light of these significant drawbacks to the use of chemicals in controlling the populations
of crop-destroying pests, it is imperative to continue research into alternative schemes to
reduce dependence on chemical pesticides.
The concept of “keystone” species is not novel, and the wide-spread attention
drawn to serious population declines of the American honey bee and amphibian species
in the latter half of the twentieth century indicates a dawning awareness on the part of the
scientific community that the demands being placed on this closed system have
consequences (Meteyer et al., 2000; Johnson, 2007). These consequences may not be
readily apparent to the average world citizen; nevertheless, the decimation of seemingly
peripheral species should serve as a warning that all is not well. In the same way other
species as diverse as bees and frogs have sounded this warning, bats also function as bio-
indicators for the environments surrounding their various species (Jones et al., 2009). As
in most cases of this sort, the connections between human activities and broad-ranging
declines amongst the flora and fauna surrounding person-kind are complex, nuanced, and
multi-faceted.
Over time, nature has evolved a system of checks and balances between predator
species and the insect populations that target the most prominent food crops grown
domestically. In the case of corn, soybeans, wheat, and rice, a great number of bird and
bat species have developed into extremely effective ‘bug killing machines.’ It is a well-
documented fact that individual North American bats readily devour up to 2000 insects

3
each night and crops without these nightly visits show over a 150% increase in pest load
(Kalka et al., 2008). In addition to feasting on specific crop pests, bats are also believed
responsible for significant control of mosquito populations in states like Texas and
Florida, where millions of dollars are spent annually in efforts to control the spread of
West Nile Disease (Cleveland et al., 2006). Given the financial and environmental
impact of controlling insect populations by chemical means, it should be of particular
concern when an emerging infectious disease decimates over 90% of affected bat
populations and results in an even greater use of pesticides that eventually make their
way downstream into the environment (Blehert et al., 2009).
The so-called “White-Nose Syndrome,” where bat populations affected by the
Geomyces destructans fungus leave hibernation early and experience high mortality rates
due to insufficient wintertime food resources, has become rampant along the eastern
seaboard since its initial discovery in Albany, NY in 2006 (Blehert et al., 2009), and
confirmation of the fungus itself being present occurred as far west as Missouri (Wibbelt
et al., 2010) and Oklahoma, although there is no report of resulting mortality in those
western-most states as of now.
Since its initial discovery in Howe’s Cave outside Albany, New York in the latter
months of 2006 and subsequent confirmation in March of 2007, White-Nosed Syndrome
(WNS) and its causative Geomyces destructans fungus have been responsible for
decimating populations of Myotis Lucifugus (Little Brown bats) and Myotis sodalis (the
already critically endangered Indiana bat). Based on census numbers from the United
States Fish and Wildlife Service (FWS) and state-level agencies, the trend of mortality is
apparent with greater than 90% declines in populations of both species at surveyed sites

4
and predictive population models indicate regional extinction of Myotis lucifigus within
shockingly short time frames (Frick et al., 2010). Despite intensive efforts underway in
a variety of fields to understand the mechanisms and dispersal factors of WNS,
researchers have yet to conclusively identify the pattern of the disease spread and are thus
significantly handicapped in their attempts to predict where and when the next outbreaks
will occur. This inability to pinpoint even approximate spread patterns leaves state and
federal efforts lacking at being able to mitigate the devastating effects of this disease
upon one of our most economically valuable wildlife groups.
The goal of this project is two-fold; firstly, to gather all existing WNS outbreak
data by county level location and outbreak date at the level of month, and secondly, to
utilize the statistical tools available within the ArcMap GIS software to perform an initial
analysis of the disease dispersion patterns present as of April 2012 in an effort to better
understand the mechanism(s) behind disease dispersion. Should outlier values be yielded
by such a statistical analysis of confirmed WNS sites and their infection dates, it would
be an indication that a host capable of acting as vector to the fungus over significant
distances could be implicated in contributing to the disease dispersal.

5
Literature Review
The full economic impact of bat predation on pest insect populations is poorly
documented; however, studies by Wickramasinghe et al. (2004), Cleveland et al. (2006),
Kalka et al. (2008), and Boyles et al. (2011) indicate that the predatory role of bats on
food pests is considerable. In the case of the Brazilian free-tailed bat, Cleveland et al.
(2006) found that there was a 12% overall decrease in corn crop infestation by larval
worm pests directly attributable to bat predation in south-central Texas. Although studies
have not been conducted to confirm the effect across other parts of the country, this
figure is likely to be similar in states that also have insectivorous bat populations whose
range extends over agricultural areas. Given that insectivorous bat species function as a
natural pest-control mechanism with not insignificant financial and environmental
benefits, the loss of large portions of indigenous bat populations as seen in White-Nose
Syndrome is especially troubling.
The first reports of this mysterious disease that resulted in high bat mortality were
centered at Howe’s Cave of Albany, New York, in the early spring of 2006 (Blehert et
al., 2009) when recreational cavers noticed high numbers of dead bats immediately
outside of the cave. Upon examination by local fish and wildlife employees, it became
apparent that not only was the local population of little brown bats (Myotis lucifugus)
experiencing mortality of greater than 90%, but that significantly abnormal behavior in
flying and feeding patterns was observed in surviving individuals. Losses of greater than
90% were then discovered in nearby hibernacula during winter of 2006-2007 and the
condition rapidly spread along the United States eastern seaboard. As of spring 2012,
nine insectivorous bat species in nineteen states had experienced Geomyces destructans

6
infection (USFWS, July 2012d) and research into the cause of WNS-associated mortality
was well underway. Despite the identification of the Geomyces destructans fungus
responsible for WNS (Lorch et al., 2011), and the physiological symptoms of the
syndrome (Reichard and Kunz, 2009), the exact mechanism of the resulting fatality of
infected individuals is still poorly understood. It is of particular interest that the presence
of bats exhibiting White-Nose fungus symptoms has been well documented in a majority
of European countries with no resulting mortality (Wibbelt et al., 2010). Equally
noteworthy is the correlation existing research has found between body fat ratios and
mortality rates (Geluso et al., 1976; Clark et al., 1983), since metabolic malfunction is
one possible culprit behind White-Nose Syndrome deaths. Starvation after early awaking
from hibernation has been implicated (Turbill and Geiser, 2008; Reichard and Kunz,
2009) and further research speculates that evaporative water loss leading to dehydration
is the motivating force behind early awakening (Cryan et al., 2010; Willis et al., 2011).
Advances in computing power have allowed for increasingly rigorous
investigations of Tobler’s First Law of Geography, which states, “Everything is related to
everything else, but near things are more related than distant things” and this most
fundamental statement of autocorrelation or the reality that, “any set of spatial data is
likely to have characteristic distances or lengths at which is it correlated with itself”
(Sullivan and Unwin, 2003; 180) and can now be readily applied to real world
phenomena. Natural world occurrences, like those of White-Nose Syndrome, can be
analyzed statistically to reveal patterns within the dataset that reflect autocorrelations and
these patterns can be compared to a randomly generated matching dataset to establish the
degree of non-randomness present in the phenomenon being examined.

7
One such analysis that examines spatial autocorrelation across a dataset is the
Moran’s I statistic which is typically applied to areal units that possess interval data
characteristics and compares the relationship of observed data to that of randomly
generated data in order to establish a measure of positive autocorrelation (clustered),
negative autocorrelation (dispersed), or no autocorrelation (random) (O’Sullivan and
Unwin, 2003; 197). Although useful for determining overall patterns of a particular
dataset, the Global Moran’s I statistic falls short when examining the relationships
between sites of that dataset, and this shortcoming was addressed with the development
of a Local Moran’s I analysis that yields further information about where the patterns of
autocorrelation exist within the occurrences of interest (Anselin, 1995). An important
distinction between the analyses is that Local Moran’s I is disaggregate and therefore
examines the degree to which neighboring data points are similar or dissimilar.
Despite the significant volume of research that has been conducted around the
physiological consequences of WNS, characterization of the fungus itself, and controlled
examinations of transmission factors, less emphasis has been placed on understanding the
patterns within the disease dispersal of existing sites. In part, the reluctance to undertake
these sorts of analyses may be due to a perceived lack of tools to address the nature and
scope of the epidemic. However, since the development of ArcGIS’s sophisticated
statistics software package and the inclusion of tests such as Global and Local Moran’s I,
Geary’s Function, K Function, these statistics-driven analyses have been utilized by
numerous researchers from very different fields of study to analyze similar phenomena
and depict results in a concise, visually compelling manner.

8
In their research tracking shigellosis, McKee et al. (2000) employed a simple k-
nearest neighbor calculation to explore infection patterns on a military base at Fort Bragg,
North Carolina and established that the infection is indeed locally contagious. Prior to
the shigellosis research, K-function analysis had also been effectively utilized by
Morrison et al. (1998) in their work with tracking the spatial and chronological analysis
of dengue cases in Florida, Puerto Rico, and this analysis also revealed a distinctive
“hotspot” pattern. To address the shortcomings of k-nearest neighbor analysis, Local
Moran’s I analysis was used by Yeshiwondim et al. (2009) to further explicate spatial
patterns of malaria incidents in Ethiopia and this increasingly rigorous analysis revealed
that the CO-type (cluster/outlier type; referring to high-high, low-low, etc. data)
“outliers” that result from Local Moran’s I analysis do indeed offer a more nuanced,
detailed examination of the phenomenon than that found by nearest neighbor statistics
alone. Another study focused on Dengue fever in Northern Thailand by Nakhapakorna
and Jirakojohnkoolb in 2006 used the more specific Local Moran’s I and Geary’s
analysis available within the ArcMap platform to provide a detailed analysis of outlier
regions within the epidemic study area. Again, this Local Moran’s I analysis was utilized
more recently in research by Sugumaran et al. (2009) that examined the spread and
distribution of West Nile virus incidence in the United States to reveal areas in which
neighboring counties showed significantly different values of viral outbreaks. West Nile
Virus was also the focus of further research utilizing Local Moran’s I analysis to examine
spatial autocorrelations of hotspot activity over time (Carnes and Ogneva-Himmelberger,
2011).
In contrast to the dispersal patterns from auto-migration of zoonotic agents,

9
communicable human-borne diseases have been shown to demonstrate a distinct pattern
of dispersal that includes outlier values when analyzed statistically using Local Moran’s
I. In a study of meningococcal meningitis occurrences found in Niger, outlier values
were found and illustrate how movement characteristic of human vectors and the
respiratory and throat secretions accompanying such movements are reflected in the
analysis output (Paireau et al., 2012).
The aforementioned studies illustrate an important distinction that should be made
between the application of the Moran’s I analysis on diseases like West Nile and
meningitis, in which the single vector is known, and White-Nose Syndrome, in which
multiple species with differing abilities to spread the causative fungus over distance are
potentially acting in a vector capacity. Within the local statistical analysis, both a typical
“hotspot” pattern as well as what can be considered an “outlier” pattern are reported and
yield important information about the vectors responsible. Since the z-score results
reported from Moran’s I analysis are in essence the slope of a regression line derived
from the scatter plot of the differences between data points and the CO-type reported is
determined by which quadrant (e.g. “high-high”, “low-low”, “high-low”, or “low-high”)
a particular relationship is found, these results contain telling information about the data.
Because any CO-type outlier reported from the Local Moran’s I statistical analysis
represents an instance in which a single data point has a significantly different value from
that of its neighbors, a pattern characterized by an outlier nature is indicative of a
phenomenon in which a vector (most typically humans) carries the causative agent over
greater distances than would be expected from the normal migration patterns of a wildlife
vector given their normal range.

10
Thus, what could be clearly shown in the CO-type outlier results generated by
Local Moran’s I statistical analysis, and particularly relevant to an understanding of
White Nose Syndrome spread patterns, is what might be considered a “jump-gap” pattern
indicating new infection of a site geographically (or in this case, temporally) distant from
previous infection sites. Given the current lack of statistical analyses performed on
potentially multi-vector diseases, this study shows promise in addressing a broader range
of applications in epidemiological examinations.

11
Methods
To accomplish the first goal of gathering WNS outbreak data from the seventeen
states affected by WNS as of March 29, 2012 into a single data set that was appropriate
for importing into the ArcGIS platform, a time-intensive process of approaching
individual researchers, research groups, private and university labs, conservation groups
at the state and federal level, and members of the National Speleological Society to
request all available data was necessary. Due to the various methods being used in
existing studies for data collection purposes, the dataset used for this project was an
amalgamation of data from multiple sources and the criteria used to determine what sites
and dates were judged sufficiently rigorous to appear in the underlying dataset for this
project deserve explication. The counties in the final dataset were provided by Cal
Butchkoski of the Pennsylvania Game Commission and Carl Herzog of the New York
Department of Conservation and were initially cross-checked against posted dates found
from the USGS National Wildlife Health Center (NWHC) and work done by researchers
at the United States Geologic Survey. An important detail to note, however, is that the
dates of WNS confirmation in the NWHC dataset are frequently limited to seasons (i.e.
“Winter, 2007) and thus are not particularly useful for analysis purposes.
From this initial dataset, monthly occurrence of first WNS confirmations were
established from primary research sources and from the “suspicious mortality” listings of
the NWHC for all data points when possible. Counties in which no published
confirmation date could be used to establish month-level outbreak dates were assigned a
month date that reflected the median date within the three month range provided by
NWHC; for example, “Winter 2009” value was set to January 1, 2009 within the

12
functional dataset used for analysis. Although this method is not without drawbacks, it
was used consistently and notes were made within the attribute table to reflect points in
which the monthly dates are approximated. The potential concern with using the
aforementioned method to establish dates from the NWHC database was that it might
artificially affect the results reported from a temporally sensitive analysis like the Local
Moran’s I (LMI) as it is being used in this context. To address this potential issue of date
bias within the NWHC derived dates, an additional analysis was carried out on the
dataset in which the confirmation date of Geomyces destructans detection was randomly
altered by up to 3 months (either earlier or later by a computerized random algorithm) to
reflect the seasonal reporting nature of these dates and the results of this analysis were
compared to the unaltered analysis results. It is important to note that the data compiled
for this research is comprised of sites in which the fungus itself has been confirmed rather
than the appearance of WNS symptoms since there are sites such as Kentucky and
Oklahoma which have confirmed fungus sightings without associated mortality or
physiological symptoms of the disease.
A deficiency of the dataset that was a potential impediment to conducting an
analysis of the WNS distribution pattern was that point data for most karst and cave
areas, and thus hibernaculum areas, is not available for public use. A novel method was
utilized in this analysis by using the compiled WNS confirmation month to establish
interval data points within the attribute table upon which Moran’s I global and local
statistical and pattern analyses were based. Due to the nature of extensive cave systems,
it is possible that county level data could be more representative of the distribution within
a naturally occurring area and within ArcGIS analysis, it is possible to perform spatial

13
autocorrelation statistical tests at this level. To avoid zero values and the inherent
difficulties of mapping the results of mathematical analysis of these zero values, time
zero (that of the first confirmed outbreak in Schoharie County, New York) was set to “1”
and each successive outbreak month was then assigned a value relative to this first
outbreak date. A clear understanding that the attribute being examined here is time
interval from initial outbreak rather than the geographic location of the outbreak itself is
crucial in any interpretation. The complete dataset on which final analyses were
performed can be found in Appendix I. An additional note of importance is that the
analyses carried out here were conducted only upon the sites listed and not on
surrounding counties with no Geomyces destructans confirmation.
The assembled dataset as completed above was imported into ESRI’s ArcMap
GIS and both Global and Local Moran’s I analysis was performed using the variable of
time (in this case, months from initial outbreak) with inverse Euclidean distance
parameters. The Local Moran’s I formula is shown below in Equation 1, which
illustrates its utility in analyzing the deviation from global mean to establish the
similarity of weighted neighbors that are reported as CO-type and associated degrees of
significance.
An additional factor in the decision to utilize Global and Local Moran’s I
statistical analyses rather than similar statistical methods was the non-normative
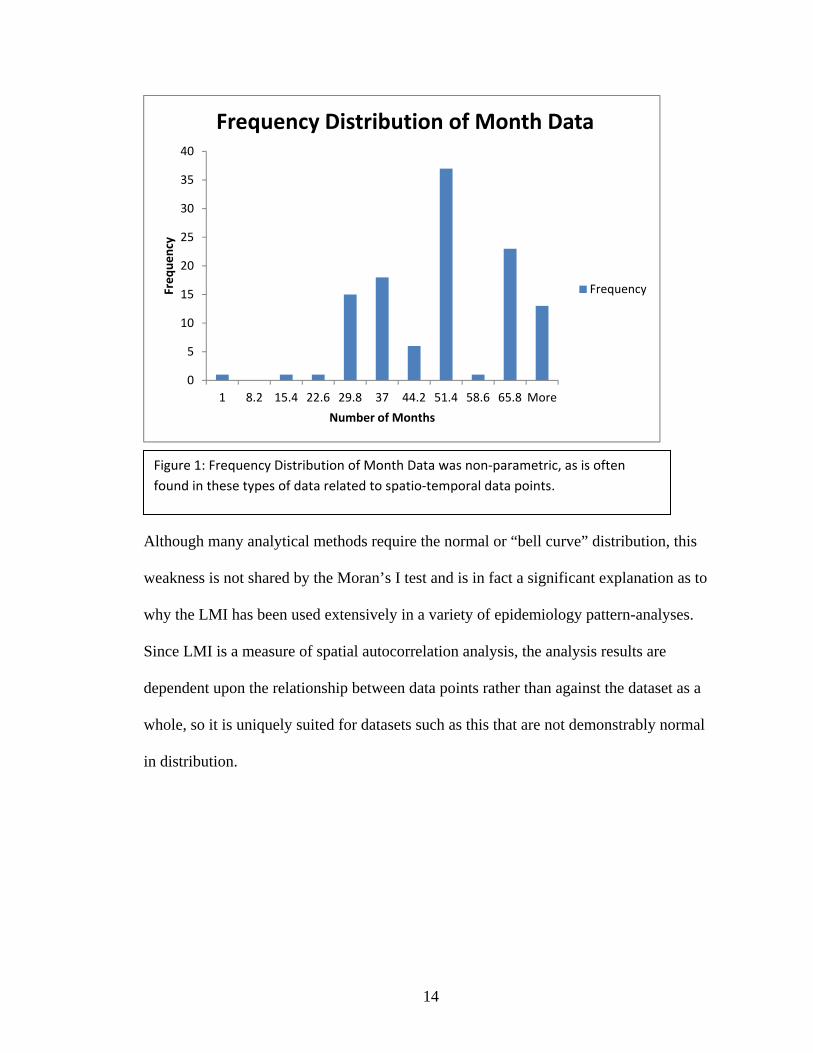
distribution of data points (Figure 1) within the completed dataset.
Equation 1: Local Moran’s I
14
Although many analytical methods require the normal or “bell curve” distribution, this
weakness is not shared by the Moran’s I test and is in fact a significant explanation as to
why the LMI has been used extensively in a variety of epidemiology pattern-analyses.
Since LMI is a measure of spatial autocorrelation analysis, the analysis results are
dependent upon the relationship between data points rather than against the dataset as a
whole, so it is uniquely suited for datasets such as this that are not demonstrably normal
in distribution.
0
5
10
15
20
25
30
35
40
1 8.2 15.4 22.6 29.8 37 44.2 51.4 58.6 65.8 More
Frequency
Number of Months
Frequency Distribution of Month Data
Frequency
Figure 1: Frequency Distribution of Month Data was non‐parametric, as is often
found in these types of data related to spatio‐temporal data points.

15
Results
One of the greatest advantages of graphically displaying the results of the
Moran’s I analyses conducted over the course of this study is that the underlying patterns
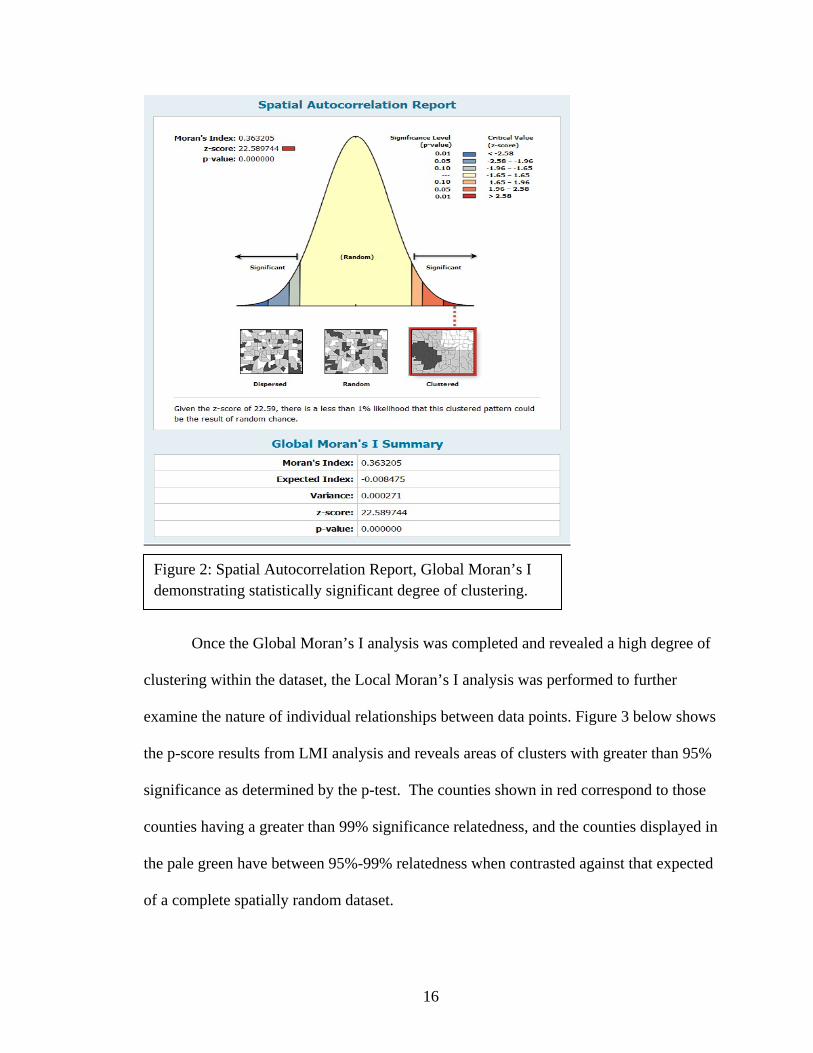
make themselves readily apparent. Results from the Global Moran’s I analysis
demonstrate conclusively that there is indeed an overall clustered pattern upon
examination of the entire dataset as shown below in Figure 2. The large z-score and
extremely small p-value meaningfully show the clustered pattern to have a less than
0.01% chance of being due to random occurrence and strongly indicate that the real
world phenomenon being examined does indeed have a spatiotemporally significant
correlation. Put simply, the overall pattern of infection exhibits a significant grouping of
similar values due primarily to the large number of sites found early on in the epidemic
and this clustered effect is due to the occurrence of Geomyces destructans infection itself
rather than random chance.
16
Once the Global Moran’s I analysis was completed and revealed a high degree of
clustering within the dataset, the Local Moran’s I analysis was performed to further
examine the nature of individual relationships between data points. Figure 3 below shows
the p-score results from LMI analysis and reveals areas of clusters with greater than 95%
significance as determined by the p-test. The counties shown in red correspond to those
counties having a greater than 99% significance relatedness, and the counties displayed in
the pale green have between 95%-99% relatedness when contrasted against that expected
of a complete spatially random dataset.
Figure 2: Spatial Autocorrelation Report, Global Moran’s I demonstrating statistically significant degree of clustering.

17
Notably, the areas of significance depicted in Figure 4 correspond to those areas
shown to be clustered under the CO-type analysis reported from LMI conducted on the
temporal assignments of WNS associated fungus confirmation, with those counties with
“early-early” and “late-late” designations having >99% significance (Figure 4). In the
context of this analysis, “early-early” is defined as a data point having low number of
months after initial infection that is surrounded by neighbors of similar values and “late-
late” is defined conversely. The presence of mixed relationships, “late-early” and “early-
late” as shown in dark green and magenta, respectively, is significant based on the
Figure 3: Local Moran’s I p‐score demonstrating areas of significant clustering.

18
robustness of the analysis. The CO-Type results can be thought of as a measure of
similarity to nearby sites, with “early-early” CO-Type signifying an early confirmation
site that is effectively surrounded by other sites with similarly early confirmation dates.
As mentioned previously, in order to test the robustness of the dataset against date
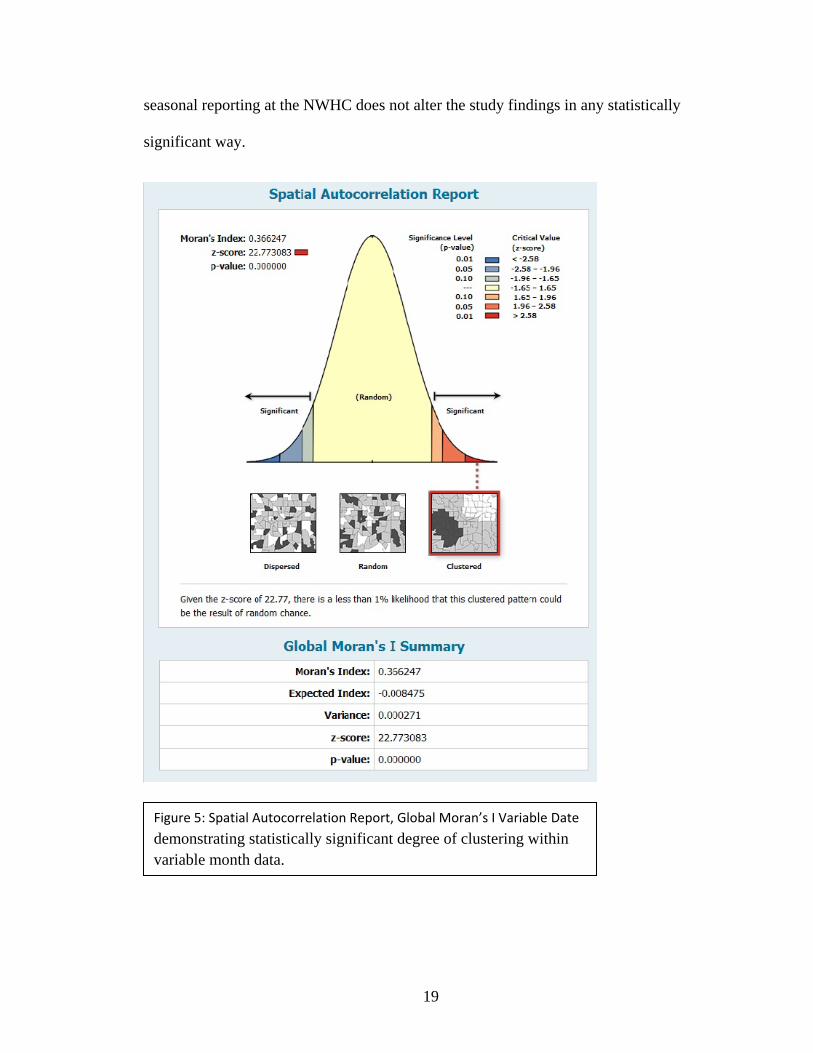
bias, an analysis with identical parameters to that of the original was performed upon the
alternate dataset that varied each confirmation date within a range of three months and
the results are depicted in Figures 5, 6, and 7. It is possible to understand from the
observed results that the possible imprecision introduced into the dataset from the
Figure 4: Local Moran’s I CO‐type reported for analysis of temporal distance from
initial confirmation date showing Early‐Late outlier (Pendleton County, WVA) and
Late‐Early outliers (Fairfield County, CT and Oxford County, ME).
19
seasonal reporting at the NWHC does not alter the study findings in any statistically
significant way.
Figure 5: Spatial Autocorrelation Report, Global Moran’s I Variable Date
demonstrating statistically significant degree of clustering within variable month data.

20
Figure 6: Local Moran’s I CO‐type, Variable Date reported for analysis of temporal
distance from initial confirmation date showing Early‐Late outlier (Pendleton
County, WVA) and Late‐Early outliers (Fairfield County, CT and Oxford County, ME).

21
Figure 7: Local Moran’s I p‐scores, Variable Date demonstrating areas of significant
clustering.

22
Discussion
Once the labor intensive process of assembling the necessary data into a format
conducive to analysis within the ArcGIS software platform was completed, it became
clear that Moran’s I global and local analysis yielded the most useful information in
determining whether a pattern exists within the data itself (Global Moran’s I, Figures 2
and 5) and, if such a pattern was present, what characterized the nature of that pattern
(Local Moran’s I, Figures 3, 4, 6, and 7). Although a statistically significant clustered
pattern of WNS outbreak sites and dates is present, this is not unexpected due to the
nature of typical disease outbreak and hotspot tendencies as illustrated by the West Nile
virus, meningitis, and Dengue fever outbreaks discussed previously. Of more particular
interest and relevance to the matter at hand is the precise nature of the pattern within the
larger, global clustered pattern, and it is this question that the Local Moran’s I (LMI)
analysis is best suited to answer.
The most unique ability of Moran’s I analysis in this context is the determination
of the presence of so-called “outlier values,” which would indicate a jump-gap
transmission pattern despite the limitations of currently available data (the lack of cave
specific latitude and longitude coordinates and thus “point” data). The aforementioned
limitations of the dataset that make analysis by other methods inappropriate in this
instance are not shared by LMI analysis and this makes it particularly useful in coming to
an understanding of whether the Geomyces destructans fungus believed to be responsible
for WNS follows the roughly linear spread pattern that would be expected from a bat-to-
bat transmission mechanism (single vector with multiple reservoirs) like that shown by
mosquito transmission of West Nile Virus (Sugumaran et al., 2009) or is instead being

23
spread by human transmission from caver gear or clothing from site to site, which would
be reflected by the presence of outliers within the dataset similar to those seen within the
spread of meningitis as previously discussed (Paireau et al., 2012).
Considerable debate has ensued surrounding the nature of WNS dispersal, and
because outliers within Local Moran’s I analysis results occur when an individual data
point is judged to be significantly different in value than its surrounding neighbor
locations, the existence of numerous outliers could be interpreted as evidence of the
“jump dispersal” pattern that might implicate human transmission by visitation to
multiple caves after being in an infected cave as a culprit in the spread of the disease.
Conversely, since only three CO-type outliers were found while performing the LMI
analysis on the WNS mortality confirmation date dataset (Figure 4), the results of this
study suggest that the dominant spread of WNS can be more likely attributed to direct
contact transmission between affected individuals within the bat populations themselves
rather than a human vector model of transmission. Because annual migration of the most
severely affected bat species is limited to distances of 520 km or less for Myotis sodalis
(Kurta and Murray 2002) and 48.5 km for Myotis lucifugus (Butchkoski 2010), the
maximum annual transmission distance of Geomyces destructans by a bat carrier would
be within that range based on the current transmission data. In contrast, human carriers
could potentially travel well in excess of the above stated migration limits and thus
“seed” a location with the fungus at much greater distances. If such a long-distance
seeding were to occur by human carriers as a matter of routine transmission, the resulting
White-Nose Syndrome mortality confirmation site would be revealed as an outlier with a

24
value earlier than that of surrounding confirmed sites (or an “early-late” outlier within the
LMI analysis as performed in this study).
The two “late-early” outliers found in Fairfield County, Connecticut and Oxford
County, Maine (Figure 6) represent counties in which the Geomyces destructans fungus
was detected at a statistically significant later date than those counties nearest the sites,
and suggests that these counties may possess some topographical or ecological variation
which prevented the spread of WNS into bat populations there at earlier dates.
Alternatively, the surveying interval due to cave or mine access issues, or lack of
surveying resources or personnel, could account for the difference in detection date; the
fungus may have been present prior to reporting date but due to a delay in survey, was
not discovered until later in the epidemic. Since the emphasis on most disease
transmission research is the front edge of an epidemic, these “late-early” outliers are of
potential interest in exploring what factors may have played a role in the transmission
delay, however do little to explicate the underlying dispersal mechanism.
Of more concern and relevance to understanding the dispersal pattern of
Geomyces destructans is the Pendleton County, West Virginia (WV) site (Figure 6) that
is shown to be an “early-late” outlier by Local Moran’s I analysis. The presence of such
an outlier could be interpreted as support for a “human as vector” model of disease
transmission, and in fact, if analysis had revealed a pattern of such outliers, such a
conclusion could be meaningfully drawn. However, closer examination of the dataset,
particularly those counties surrounding Pendleton County, reveals that counties south of
the site (Giles confirmed 2/2009, Bland confirmed 5/2009) showed earlier infection. The
seven counties surrounding Pendleton County show considerable variation in reported

25
dates (Figure 6- Pocohantas WV on 3/2010, Highland VA on 4/10, Rockingham VA on
8/2009, Tucker WV on 2/2011, Grant WV on 4/2011, Hardy WV on 3/2010, and Bath
VA on 2/2009), and this constellation of occurrence dates suggests that the outlier status
of Pendleton County, WV is perhaps a reflection of reporting artifacts and the much later
reporting of Tucker and Grant counties in West Virginia is sufficiently later to alter the
statistical analysis reporting of CO-type status even when weighed against the earlier
dates of surrounding counties. Given the inherent financial, time, and access resource
constraints placed on survey efforts in the mountains of West Virginia, it is possible that
earlier appearances of the fungus were unrecorded.
When the analysis was performed again using a dataset that had been artificially
altered to reflect a degree of three month variability in either the forward or backward
temporal direction, the results did not differ in any statistically meaningful way from the
original dataset, and again revealed a pattern with two clusters (early-early and late-late)
along with the presence of outlier values for Pendleton County, WV, Fairfield County,
CT, and Oxford County, Maine (Figures 5, 6, and 7). Performing the Moran’s I analyses
on a dataset in which the data points had been randomly altered by three months
addressed a potential vulnerability of the analysis because of the possible variation in
confirmation reporting dates due to the inherent difficulty in obtaining primary data;
timing of cave surveys, difficulty in accessing hibernaculum sites, and seasonal
limitations all have the effect of introducing a possible “timing artifact” bias into the
results. It also addresses the possible issue of sensitivity of the analysis to be able to
detect outliers within a larger reporting error. The variable month dataset and analysis
results effectively demonstrate that the nature of the pattern found in this study is a

26
reflection of the actual phenomenon being examined rather than a side-effect of the
unavoidable data collection limitations. Had there been an alteration of outlier values in
the CO-type reporting by such an alteration of the dataset, the veracity of such a
statistical analysis could rightly be questioned. However, in this case, the affirmation by a
high degree of similarity of reported values further emphasizes the robust nature of the
analysis itself.
Ultimately, the results of this study, primarily the lack of multiple “early-late”
outlier values as revealed by Local Moran’s I CO-type reporting and associated
significance values (Figures 3 and 4), suggest that the dominant transmission pattern of
White-Nose Syndrome in the United States is typical of that found in diseases
characterized by direct transmission between individuals of affected bat species rather
than that of a separate vector. One would expect that if humans were acting as a vector
for Geomyces destructans, the dispersal pattern would resemble that of meningitis as
discussed above and outlier values representing “jumps” in the dispersal would be
present. This finding is contrary to much of what has been supposed about the dispersal
of WNS and further emphasizes just how critical the creation of, and continued efforts to
maintain, a comprehensive dataset of outbreak data is to appropriately address the issue at
hand and arrive at policies that more effectively mitigate White-Nose Syndrome. As any
research is limited to the quality of data upon which the analyses are performed, the
discrepancy around the Pendleton, WV confirmation dates should serve as a strong
reminder of how critically important the practice of standardized, rigorous data collection
is to our understanding of the timing and occurrence of natural phenomena like infectious
disease dispersal patterns. Additionally, because such robust data collection is made even

27
more difficult with diminished resources, it should serve as a call for increased attention
to communication and management at both the state and federal levels in order to more
promptly develop monitoring and surveying plans, and mobilize “ground troops” for
future outbreaks.
A more complete understanding of the dispersal patterns of White-Nose
Syndrome, as revealed by careful statistical evaluation of the relationships between each
confirmed site within the compiled dataset, is absolutely critical to the development of a
scientifically valid, well-reasoned approach to addressing and possibly mitigating the
devastating effects of WNS. Previously, such large-scale studies have not been
conducted due in part to the lack of a comprehensive dataset and, now that the stated
research goal of establishing such a dataset and determining its usefulness has been
accomplished here, more scientifically rigorous evaluations of the dispersal patterns
found within the epidemic can be built upon the results of this study. Of particular
interest for future mitigation efforts could be the development of a dataset that compares
Geomyces destructans infection dates against WNS associated mortality dates. Such a
dataset would allow for the detailed examination of transmission and mortality
expression rates and could potentially reveal crucial information about factors which
influence disease development, however that data is not yet available and will take
considerable cooperation and effort to develop into a meaningful investigative tool.
In addition to the chronological and temporal analysis of the disease spread itself,
perhaps one of the most powerful aspects of applying ArcMap’s graphical capabilities to
an examination of this disease spread pattern is that alternate variables, such as
topography, prevailing wind patterns, and migration paths, can be incorporated into

28
further analyses. Although it is impossible to predict at this stage which of these
variables, if any, will prove to be significant upon further examination, the fact that the
Geomyces destructans fungus is cold-loving and therefore potentially affected by factors
such as annual temperature and geomorphology of the hibernaculum themselves strongly
suggests that a correlation with these annual temperatures and dominant wind patterns
should present itself when used as a co-variable against the spread pattern and resulting
mortality. As of completion of this study, wide-ranging efforts are underway to establish
robust baseline population numbers for affected bat species. These census efforts
coupled with more sophisticated mortality counts could potentially allow further research
into how the climatic and ecological factors mentioned above play a role in individual
and population survivability.
Most current mitigation techniques have involved closing caves or mines, or
limiting access by humans, along with evolving decontamination procedures (USFWS,
2012a,b). A better understanding of the fungus itself and its survivability in various
environments from recent research also introduced continued discussion about how best
to address WNS issues (Blehert 2011). However, many of the current efforts to curtail the
spread of Geomyces destructans have been hampered by an incomplete understanding of
the underlying patterns of dispersal, combined with the lack of a standardized method of
disease discovery and reporting. Additionally, the policies in place at the federal and state
government level regarding well-intentioned cave and mine closures may have had the
unintentional effect of diminishing beneficial input from potential allies within the
outside community. Given that eventual modeling efforts will be largely dependent on
the robustness of site data (both spatial and temporal) and the large role played by non-

29
governmental visitors in establishing outbreak sites early in the disease, the decision to
close all caves and mines on public land may have the end result of decreasing our ability
to reasonably predict where and when future outbreaks will occur.
In order to effectively collect and compile a current dataset of ongoing outbreak
sites, it will be necessary to establish a clearinghouse for suspected WNS sites at the
federal level with a correspondingly rigorous protocol to determine data accuracy and
what criteria by which to include new sites. For the purposes of this study, WNS
associated mortality was not a prerequisite for inclusion in the dataset; however, the
nature of the causative fungus and manifestation of WNS mortality itself is as of yet
incompletely understood with some species exhibiting symptoms of WNS with no
resulting mortality (USFWS, 2012b). As survey and census efforts intensify, more
comprehensive data on sites infected with Geomyces destructans prior to mortality events
may well lead to important discoveries regarding species susceptibility and survivability.
Perhaps of most urgent import to wildlife managers and conservation efforts is the
ability to more effectively predict the areas which Geomyces destructans is most likely to
infect and when it will affect those areas. The novel application of Moran’s I used in this
research suggests that, not only would it be possible to create predictive geospatial
models, but that transmission time itself can be used as a variable when constructing
these predictive models. This study provides an example or the robustness of these
techniques and their ability to play a crucial role in management activities associated with
WNS and future outbreaks, while also illustrating the limitations of the analysis given the
myriad data collection and reporting techniques used to date. This has significant

30
implications not just for the disease being examined within the context of this study, but
for the broader field of wildlife epidemiology as a whole.
31
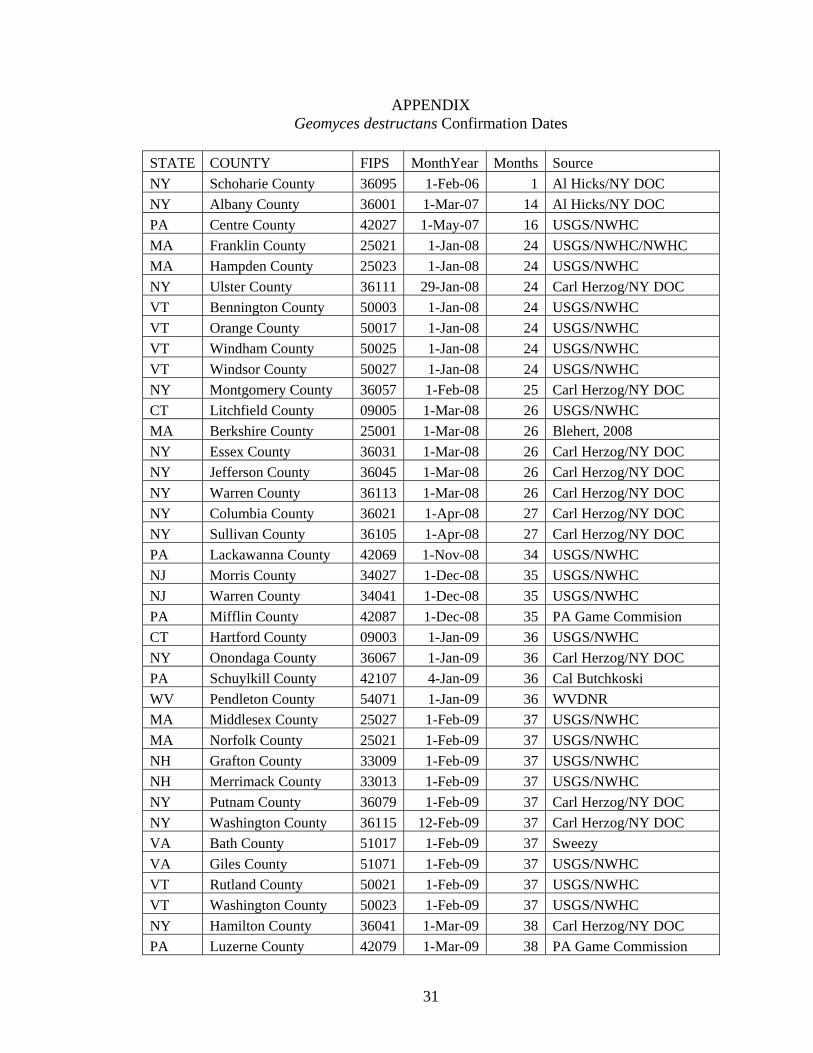
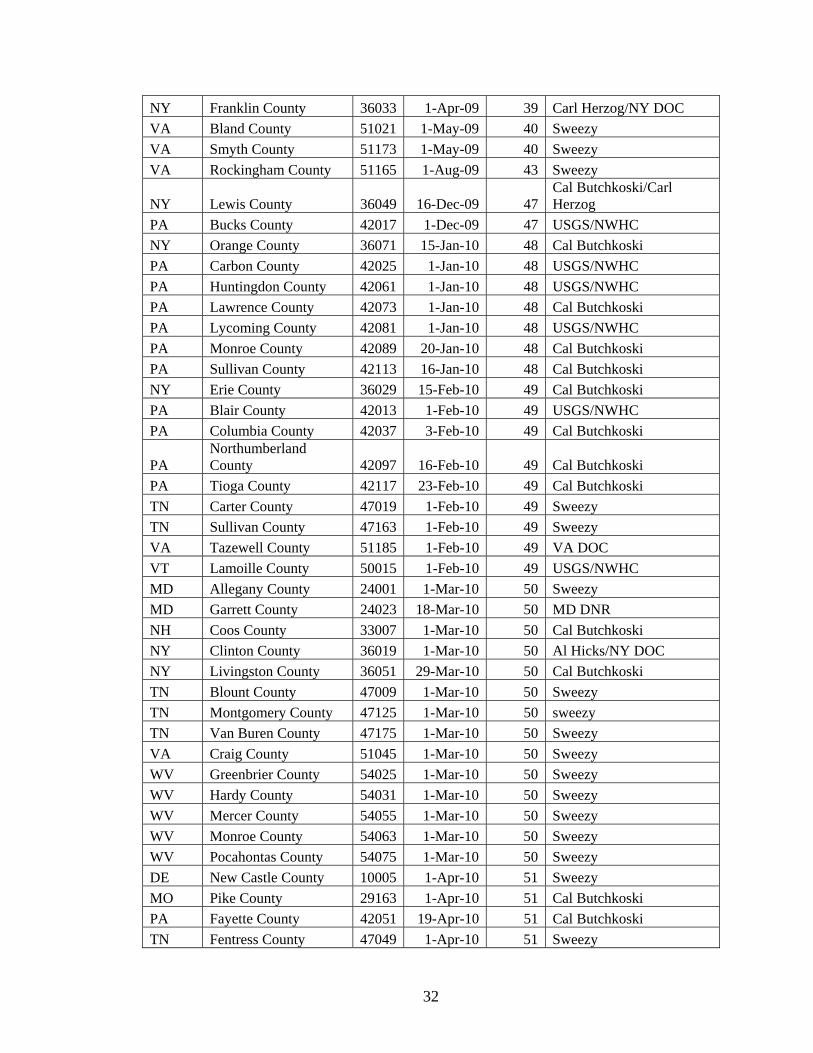
APPENDIX Geomyces destructans Confirmation Dates
STATE COUNTY FIPS MonthYear Months Source
NY Schoharie County 36095 1-Feb-06 1 Al Hicks/NY DOC
NY Albany County 36001 1-Mar-07 14 Al Hicks/NY DOC
PA Centre County 42027 1-May-07 16 USGS/NWHC
MA Franklin County 25021 1-Jan-08 24 USGS/NWHC/NWHC
MA Hampden County 25023 1-Jan-08 24 USGS/NWHC
NY Ulster County 36111 29-Jan-08 24 Carl Herzog/NY DOC
VT Bennington County 50003 1-Jan-08 24 USGS/NWHC
VT Orange County 50017 1-Jan-08 24 USGS/NWHC
VT Windham County 50025 1-Jan-08 24 USGS/NWHC
VT Windsor County 50027 1-Jan-08 24 USGS/NWHC
NY Montgomery County 36057 1-Feb-08 25 Carl Herzog/NY DOC
CT Litchfield County 09005 1-Mar-08 26 USGS/NWHC
MA Berkshire County 25001 1-Mar-08 26 Blehert, 2008
NY Essex County 36031 1-Mar-08 26 Carl Herzog/NY DOC
NY Jefferson County 36045 1-Mar-08 26 Carl Herzog/NY DOC
NY Warren County 36113 1-Mar-08 26 Carl Herzog/NY DOC
NY Columbia County 36021 1-Apr-08 27 Carl Herzog/NY DOC
NY Sullivan County 36105 1-Apr-08 27 Carl Herzog/NY DOC
PA Lackawanna County 42069 1-Nov-08 34 USGS/NWHC
NJ Morris County 34027 1-Dec-08 35 USGS/NWHC
NJ Warren County 34041 1-Dec-08 35 USGS/NWHC
PA Mifflin County 42087 1-Dec-08 35 PA Game Commision
CT Hartford County 09003 1-Jan-09 36 USGS/NWHC
NY Onondaga County 36067 1-Jan-09 36 Carl Herzog/NY DOC
PA Schuylkill County 42107 4-Jan-09 36 Cal Butchkoski
WV Pendleton County 54071 1-Jan-09 36 WVDNR
MA Middlesex County 25027 1-Feb-09 37 USGS/NWHC
MA Norfolk County 25021 1-Feb-09 37 USGS/NWHC
NH Grafton County 33009 1-Feb-09 37 USGS/NWHC
NH Merrimack County 33013 1-Feb-09 37 USGS/NWHC
NY Putnam County 36079 1-Feb-09 37 Carl Herzog/NY DOC
NY Washington County 36115 12-Feb-09 37 Carl Herzog/NY DOC
VA Bath County 51017 1-Feb-09 37 Sweezy
VA Giles County 51071 1-Feb-09 37 USGS/NWHC
VT Rutland County 50021 1-Feb-09 37 USGS/NWHC
VT Washington County 50023 1-Feb-09 37 USGS/NWHC
NY Hamilton County 36041 1-Mar-09 38 Carl Herzog/NY DOC
PA Luzerne County 42079 1-Mar-09 38 PA Game Commission
32
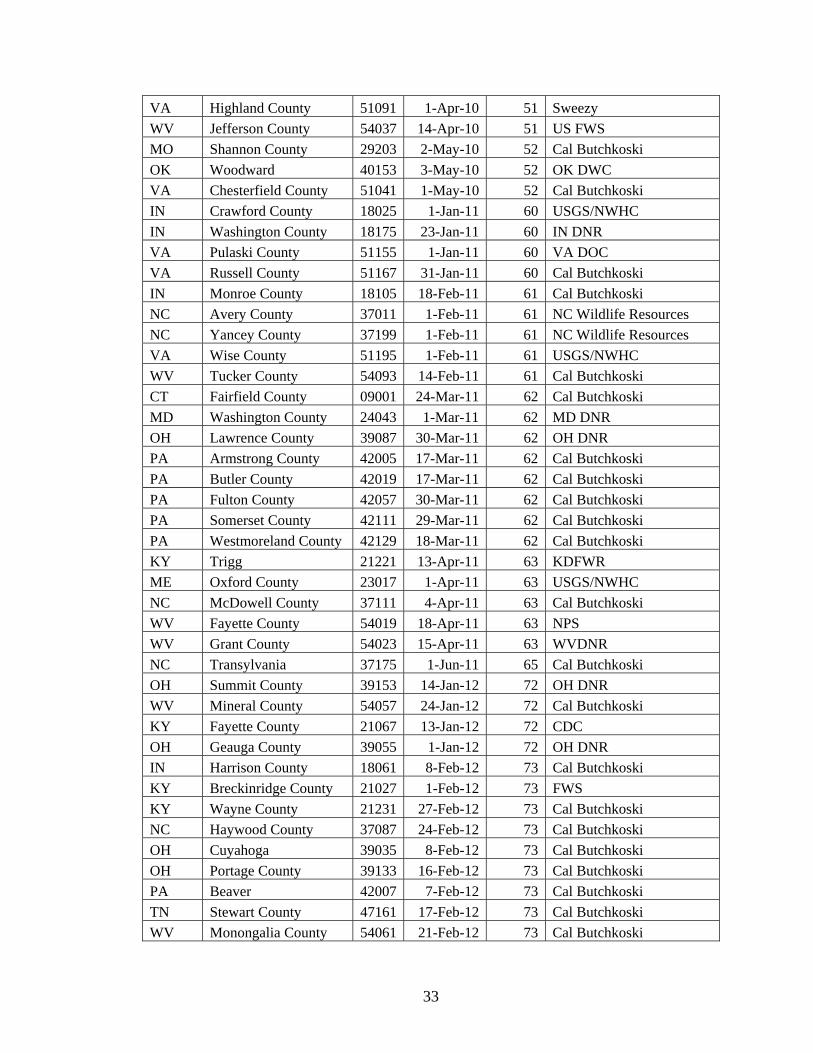
NY Franklin County 36033 1-Apr-09 39 Carl Herzog/NY DOC
VA Bland County 51021 1-May-09 40 Sweezy
VA Smyth County 51173 1-May-09 40 Sweezy
VA Rockingham County 51165 1-Aug-09 43 Sweezy
NY Lewis County 36049 16-Dec-09 47Cal Butchkoski/Carl Herzog
PA Bucks County 42017 1-Dec-09 47 USGS/NWHC
NY Orange County 36071 15-Jan-10 48 Cal Butchkoski
PA Carbon County 42025 1-Jan-10 48 USGS/NWHC
PA Huntingdon County 42061 1-Jan-10 48 USGS/NWHC
PA Lawrence County 42073 1-Jan-10 48 Cal Butchkoski
PA Lycoming County 42081 1-Jan-10 48 USGS/NWHC
PA Monroe County 42089 20-Jan-10 48 Cal Butchkoski
PA Sullivan County 42113 16-Jan-10 48 Cal Butchkoski
NY Erie County 36029 15-Feb-10 49 Cal Butchkoski
PA Blair County 42013 1-Feb-10 49 USGS/NWHC
PA Columbia County 42037 3-Feb-10 49 Cal Butchkoski
PA Northumberland County 42097 16-Feb-10 49 Cal Butchkoski
PA Tioga County 42117 23-Feb-10 49 Cal Butchkoski
TN Carter County 47019 1-Feb-10 49 Sweezy
TN Sullivan County 47163 1-Feb-10 49 Sweezy
VA Tazewell County 51185 1-Feb-10 49 VA DOC
VT Lamoille County 50015 1-Feb-10 49 USGS/NWHC
MD Allegany County 24001 1-Mar-10 50 Sweezy
MD Garrett County 24023 18-Mar-10 50 MD DNR
NH Coos County 33007 1-Mar-10 50 Cal Butchkoski
NY Clinton County 36019 1-Mar-10 50 Al Hicks/NY DOC
NY Livingston County 36051 29-Mar-10 50 Cal Butchkoski
TN Blount County 47009 1-Mar-10 50 Sweezy
TN Montgomery County 47125 1-Mar-10 50 sweezy
TN Van Buren County 47175 1-Mar-10 50 Sweezy
VA Craig County 51045 1-Mar-10 50 Sweezy
WV Greenbrier County 54025 1-Mar-10 50 Sweezy
WV Hardy County 54031 1-Mar-10 50 Sweezy
WV Mercer County 54055 1-Mar-10 50 Sweezy
WV Monroe County 54063 1-Mar-10 50 Sweezy
WV Pocahontas County 54075 1-Mar-10 50 Sweezy
DE New Castle County 10005 1-Apr-10 51 Sweezy
MO Pike County 29163 1-Apr-10 51 Cal Butchkoski
PA Fayette County 42051 19-Apr-10 51 Cal Butchkoski
TN Fentress County 47049 1-Apr-10 51 Sweezy
33
VA Highland County 51091 1-Apr-10 51 Sweezy
WV Jefferson County 54037 14-Apr-10 51 US FWS
MO Shannon County 29203 2-May-10 52 Cal Butchkoski
OK Woodward 40153 3-May-10 52 OK DWC
VA Chesterfield County 51041 1-May-10 52 Cal Butchkoski
IN Crawford County 18025 1-Jan-11 60 USGS/NWHC
IN Washington County 18175 23-Jan-11 60 IN DNR
VA Pulaski County 51155 1-Jan-11 60 VA DOC
VA Russell County 51167 31-Jan-11 60 Cal Butchkoski
IN Monroe County 18105 18-Feb-11 61 Cal Butchkoski
NC Avery County 37011 1-Feb-11 61 NC Wildlife Resources
NC Yancey County 37199 1-Feb-11 61 NC Wildlife Resources
VA Wise County 51195 1-Feb-11 61 USGS/NWHC
WV Tucker County 54093 14-Feb-11 61 Cal Butchkoski
CT Fairfield County 09001 24-Mar-11 62 Cal Butchkoski
MD Washington County 24043 1-Mar-11 62 MD DNR
OH Lawrence County 39087 30-Mar-11 62 OH DNR
PA Armstrong County 42005 17-Mar-11 62 Cal Butchkoski
PA Butler County 42019 17-Mar-11 62 Cal Butchkoski
PA Fulton County 42057 30-Mar-11 62 Cal Butchkoski
PA Somerset County 42111 29-Mar-11 62 Cal Butchkoski
PA Westmoreland County 42129 18-Mar-11 62 Cal Butchkoski
KY Trigg 21221 13-Apr-11 63 KDFWR
ME Oxford County 23017 1-Apr-11 63 USGS/NWHC
NC McDowell County 37111 4-Apr-11 63 Cal Butchkoski
WV Fayette County 54019 18-Apr-11 63 NPS
WV Grant County 54023 15-Apr-11 63 WVDNR
NC Transylvania 37175 1-Jun-11 65 Cal Butchkoski
OH Summit County 39153 14-Jan-12 72 OH DNR
WV Mineral County 54057 24-Jan-12 72 Cal Butchkoski
KY Fayette County 21067 13-Jan-12 72 CDC
OH Geauga County 39055 1-Jan-12 72 OH DNR
IN Harrison County 18061 8-Feb-12 73 Cal Butchkoski
KY Breckinridge County 21027 1-Feb-12 73 FWS
KY Wayne County 21231 27-Feb-12 73 Cal Butchkoski
NC Haywood County 37087 24-Feb-12 73 Cal Butchkoski
OH Cuyahoga 39035 8-Feb-12 73 Cal Butchkoski
OH Portage County 39133 16-Feb-12 73 Cal Butchkoski
PA Beaver 42007 7-Feb-12 73 Cal Butchkoski
TN Stewart County 47161 17-Feb-12 73 Cal Butchkoski
WV Monongalia County 54061 21-Feb-12 73 Cal Butchkoski

34
WV Preston County 54077 27-Feb-12 73 Cal Butchkoski

35
REFERENCES
Anselin L. 1995. Local Indicators of Spatial Association—LISA. Geographical Analysis. 27(2):93-115. Blehert D.S., A.C. Hicks, M. Behr, C.U. Meteyer, B. Berlowski-Zier, E.L. Buckles, J.T.H. Coleman, S.R. Darling, A. Gargas, R. Niver, J.C. Okoniewski, R.J. Rudd, and W.B. Stone. 2009. Bat White-Nose syndrome: an emerging fungal pathogen? Science. 323(5911):227 Boyles J.G., Cryan P.M., McCraken G.F., and Kunz T.H. 2011. Economic Importance of Bats in Agriculture. Science. 332:41-42. Butchkoski C. 2010. Indiana Bat (Myotis sodalis) Summer Roost Investigations. Pennsylvania Game Commission Bureau Of Wildlife Management Project Annual Job Report. February 25, 2010. Carnes A. and Ogneva-Himmelberger Y. 2011. Temporal Variations in the Distribution of West Nile Virus Within the United States; 2000-2008. Applied Spatial Analysis and Policy. DOI: 10.1007/s12061-011-9067-7 Clark D.R. Jr., C.M. Bunck, and E. Cromartie. 1983. Year and age effects on residues of Dieldrin and Heptachlor in dead gray bats, Franklin County, Missouri-1976, 1977, and 1978. Environmental Toxicology and Chemistry. 2:387-393 Cleveland C.J., M. Betke, P. Federico, J.D. Frank, T.G. Hallam, J. Horn, J.D. Lopez Jr., G.F. McCracken, R.A. Medillin, A. Moreno-Valdez, C.G. Sansone, J.K. Westbrook, and T.H. Kunz. 2006. Economic value of the pest control service provided by Brazilian free-tailed bat in south-central Texas. Frontiers in Ecology and Environment 4:238–243. Cryan P., Meteyer C., Boyles J., and Blehert D. 2010. Wing pathology of White-Nose syndrome in bats suggests life-threatening disruption of physiology. BMC Biology.8(1):135. FAO. (Food and Agriculture Organization of the United Nations). 2009. The State of Food Insecurity in the World: Economic crises – impacts and lessons learned. Accessed 10/20/10. http://www.fao.org/docrep/012/i0876e/i0876e00.htm
Frick W.F., Pollock J.F., Hicks A.C., Langwig K.E., Reynolds D.S., Turner G.G., Butchkoski C.M., and Kunz T, H. 2010. An Emerging Disease Causes Regional Population Collapse of a Common North American Bat Species. Science. 329:679-682. Gargas A., M.T. Trest, M. Christensen, T.J. Volk, and D.S. Blehert. 2009. Geomyces destructans sp. Nov. associated with bat White-Nose syndrome. Mycotaxon. 108:147-154

36
Geluso K.N., J.S. Altenbach and D.E. Wilson. 1976. Bat mortality: pesticide poisoning and migratory stress. Science. 194:184–186 Johnson, R. 2007. Honey Bee Colony Collapse Disorder. Congressional Research Service, The Library of Congress. Washington, D.C. Report date June 20, 2007. Jones G., D.S Jacobs, T.H. Kunz, M.R. Willig, and P.A. Racey. 2009. Carpe noctem: The importance of bats as bioindicators. Endangered Species Research. 8:93-115 Kalka Margareta B., Smith A.R, and Kalko, E.K.V. 2008. Bats limit arthropods and herbivory in a tropical forest. Science. 320(5872):71 Kurta A., and Murray S.W. 2002. Philopatry and migration of banded Indiana bats (Myotis sodalis) and effects of using transmitters. Journal of Mammalogy.83:585-589. Lorch J.M., Meteyer C.U., Behr M.J., Boyles J.G., Cryan P.M., Hicks A.C., Ballmann A.E., Coleman J.T.H., Redell D.N., Reeder D.M, and Blehert D.S. 2011. Experimental infection of bats with Geomyces destructans causes white-nose syndrome. Nature. October 26, 2011. doi:10.1038/nature10590.
McKee Jr., K.T., Shields, T.M., Jenkins, P.R., Zenilman, J.M., and Glass, G.E. 2000. “Application of a Geographic Information System to the Tracking and Control of an Outbreak of Shigellosis.” Clinical Infectious Diseases. 31(3):728-733. Meteyer, C.U., Loeffler, I.K., Fallon, J.F., Converse, K.A., Green, E., Helgen, J.C., Kersten, S., Levey, R., Eaton-Poole, L., Burkhart, J,G,. 2000. Hind Limb Malformations in Free-Living Northern Leopard Frogs (Rana pipiens) From Maine, Minnesota, and Vermont Suggest Multiple Etiologies. Teratology. 62:151-171. Morrison, A.C., Getis, A., Santiago, M., Rigua-Perez, J.G., and Reiter, P. 1998. “Exploratory space-time analysis of reported dengue cases during an outbreak in Florida, Puerto Rico, 1991-1992.” American Journal of Tropical Medicine and Hygiene. 58(3):287-298 Nakhapakorna,K. and Jirakajohnkoolb, S. 2006. “Temporal and Spatial Autocorrelation Statistics of Dengue Fever.” Dengue Bulletin. Volume 30:177-183. Published by World Health Organization. National Wildlife Health Center. Division of the United States Geological Survey. http://www.nwhc.usgs.gov/publications/quarterly_reports. Accessed 3/2011-5/2011. O’Sullivan D., and Unwin D.J. 2003. Geographic Information Analysis. John Wiley & Sons. Hoboken, New Jersey.

37
Paireau J., Girond F., Collard, J-M., Maïnassara H.B., Jusot J.F. 2012. Analysing Spatio-Temporal Clustering of Meningococcal Meningitis Outbreaks in Niger Reveals Opportunities for Improved Disease Control. PloS Neglected Tropical Diseases. March; 6(3):e1577. Reichard, J.D., and T.H. Kunz. 2009. White-Nose Syndrome inflicts lasting injuries to the wings of little brown myotis (Myotis lucifugus). Acta Chiropterologica. 11:457-464. Sugumaran, R., Larson, S.R., and DeGroote, J.P. 2009. “Spatio-temporal cluster analysis of county-based human West Nile virus incidence in the continental United States.” International Journal of Health Geographics. 8:43. Swezey, C. and Garrity, C. 2010. “Notes On The Geology And Meteorology of Sites Infected With White-Nose Syndrome Before July 2010 in the Southeastern United States”. U.S. Geological Survey. Reston, VA. Turbill, C., and F. Geiser. 2008. Hibernation in tree-roosting bats. Journal of Comparative Physiology. 178B:597–605. United States Fish and Wildlife Service. 2012a. White-Nose Syndrome Decontamination Protocol-Version 03.15.2012. www.fws.gov/WhiteNoseSyndrome/pdf/National _WNS_Decontamination _Protocol_v03.15.2012.pdf. Last accessed 7/3/2012.
Unites States Fish and Wildlife Service. 2012b. May 12 News Release. www.caves.org/WNS/Gray_bats_2012_NR_Final.pdf. Last accessed 7/5/12. United States Fish and Wildlife Service. 2012c. White-Nose Syndrome Message to Cavers. www.fws.gov/WhiteNoseSyndrome/cavers.html. Last accessed 7/3/12. United States Fish and Wildlife Service. 2012d. U.S. Fish and Wildlife Service Awards Grants to 30 States for White-Nose Syndrome Work. www.batcon.org/pdfs/whitenose/2012%20WNS%20grant. Last accessed 7/26/12. Wibbelt G., A. Kurth, D. Hellmann, M. Weishaar, A. Barlow, M. Veith, J. Prüger, T. Görföl, L. Grosche, F. Bontadina, U. Zöphel, H.P. Seidl, P.M. Cryan, and D.S. Blehert. 2010. White-Nose Syndrome Fungus (Geomyces destrucatns) in Bats, Europe. Emerging Infectious Diseases. 16(8):1237-1242.
Wickramasinghe L.P., S. Harris, G. Jones, and N.V. Jennings. 2004. Abundance and species richness of nocturnal insects on organic and conventional farms: effects of agricultural intensification on bat foraging. Conservation Biology. 18:1283–1292. Willis C.K.R., Menzies A.K., Boyles J.G., and Wojciechowski M.S. 2011. Evaporative Water Loss Is a Plausible Explanation for Mortality of Bats from White-Nose Syndrome. Integrative and Comparative Biology. 51:6.

38
Yeshiwondim, A.K., Gopal, S., Hailemariam, A.T., Dengela, D.O., and Patel, H.P. 2009. “Spatial analysis of malaria incidence at the village level in areas with unstable transmission in Ethiopia.” International Journal of Health Geographics. 8:5.Ope
