an anatomical study of converging sensory pathways within the cerebral cortex of the monkey
TRANSCRIPT

IBrain (1970) 93, 793-820.
I
I
AN ANATOMICAL STUDY OF CONVERGING SENSORY PATHWAYSWITHIN THE CEREBRAL CORTEX OF THE MONKEY
BY
E. G. JONES AND T. P. S. POWELL(From the Department of Human Anatomy, University of Oxford)
INTRODUCTION
IN attempting to understand the cerebral mechanisms underlying the act of per-ception, the conclusion that such activities should necessitate a considerable degree ofconvergence and integration of all sensory systems seems inescapable (Adrian, 1949;Fessard, 1954). While the cerebral cortex is clearly not the only part of the braininvolved in the perceptive act, clinical experience argues for it having a significantrole. A portion of the parietal lobe above and behind the posterior end of the sylvianfissure, certainly of the non-dominant hemisphere, is concerned with spatial relation-ships and the body image (Critchley, 1953; Hecaen et al., 1956; Ettlinger, Warring-ton and Zangwill, 1957; Penfield, 1966; Newcombe and Russell, 1969). A functionallysimilar, though smaller, region may also be present in the dominant hemisphere(Teuber, 1965). It seems that in lesions of these areas there are defects of somatic aswell as visual function, for Teuber (1966) notes that route finding by means of mapsis equally impaired whether the map is presented visually or by tactile-kinaestheticmeans. This would presuppose an element of convergence of the two sensory systemsand probably, within a single sensory system, convergence of information relevantto the several topographic subdivisions of the body or visual fields. However, theprimary somatic, visual and auditory cortices are not interconnected, and eachprojects to very restricted and entirely separate fields chiefly in their immediatevicinity (Kuypers, Szwarcbart, Mishkin and Rosvold, 1965; Diamond, Jones andPowell, 1968; Cragg, 1969; Jones and Powell, 1969). It is apparent also from clinicaland experimental psychological studies that other quite extensive, but again discrete,parts of the frontal, temporal and parietal lobes are concerned with visual, auditoryor somatic behaviour but not with two or all three of these {see, for example, Chow,1961; Mishkin, 1966; Moffett et al., 1967; Massopust, Barnes and Verdura, 1965).It seems probable, therefore, that there is, as it were, a considerable amount of"processing" within areas of the cortex related to a single sensory system before theseveral sensory pathways come together. Moreover, within the primary corticalareas of a single sensory system it would appear that although intrinsic connexions
39 BRAIN—VOL. xcin
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

794 E. G. JONES AND T. P. S. POWELL
link all parts of the representation of one topographic subdivision (say the leg area orthe superior quadrant of the retina), there are no connexions with other subdivisions(e.g. there are none between the leg and arm area) (Jones and Powell, 1969), so thatconvergence within a system must occur at some point beyond th(T primary sensoryareas. Hence, by studying the sequence of association connexions passing outwardsfrom the primary sensory areas as though following the successive steps in a (sup-posed) sequence of cortical function, it was hoped to identify regions of convergencewithin the cortex. This should not be taken to imply, however, that the corticalassociation pathways are the only routes for sensory elaboration within the fore-brain.
The present investigation has developed from a study of the connexions of thesomatic sensory cortex (Jones and Powell, 1969). The material was, therefore,prepared with this end in view and is to some extent deficient for a detailed analysisof the connexions of the auditory and visual pathways. However, when taken inconjunction with the work of Kuypers et al. (1965) and of Pandya and Kuypers(1969), it has been possible to obtain a reasonably comprehensive coverage of allthree sensory pathways.
MATERIAL AND METHODSUnilateral lesions were placed in the cerebral cortex of 23 adult rhesus monkeys (Macaca mulatto).
Most lesions were made by careful removal under the dissecting microscope of a portion of the piamater as this usually causes avascular necrosis of the cortex and reduces the amount of damage ofunderlying white matter to a minimum; in the case of very small lesions, the white matter is oftenspared completely. Larger lesions were made with a suction aspirator and in these cases there wasinevitably some involvement of subjacent white matter. The majority of the lesions were placed interms of the architectonic subdivisions of the cortex, following the maps of Brodmann (1909),Walker (1940) and Roberts and Akert (1963) but a few made in the course of another study (Kempand Powell, 1970) were regionally located only. Seven to fourteen days after the operation, thebrains were fixed by perfusion with neutral formalin and after a further period of fixation in forma-lin they were sectioned at 25 n on a freezing microtome and stained according to the methods ofNauta and Gygax (1954) and of Fink and Heimer (1967). All brains were photographed to recordthe gyral and sulcal pattern, and the lesion and distribution of degeneration were reconstructed ontracings of the brain photograph from projection drawings of the stained sections. The sulci andgyri of the hemisphere are named following Bonin and Bailey (1947) (figs. 1 and 2).
RESULTS
The results will be presented in a sequence as though following the steps of anoutward progression from each of the main sensory areas of the cortex—somatic,visual and auditory. Some of the fields in this progression are reciprocally connected,but to preserve the sequence and to facilitate description, the returning, "inward"pathways will be dealt with separately. At certain points each of the intracorticalsensory pathways come together, so that the lesions in some experiments are relevantto two or more sensory systems. It will, therefore, be necessary at times to refer backto a preceding experiment. The site of the lesion and the distribution of axonaldegeneration in the cortex will be described as far as possible in terms of the archi-tectonic areas of Brodmann (1909) except in more rostral parts of the frontal lobe
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

CORTICAL ASSOCIATION CONNEXIONS 795
where Brodmann states that his subdivision is somewhat arbitrary; here, the descrip-tions of Walker (1940) and of Roberts and Akert (1963) will be followed (fig. 1). Theterm "motor cortex" will refer only to area 4 (Brodmann), "premotor cortex" to areas6, 8 (Brodmann, Walker) and the precentral agranular field, PrCo (Roberts andAkert); "prefrontal cortex" will refer to all other parts of the frontal lobe (Walker)except the "orbitofrontal cortex" (Roberts and Akert) which will be consideredseparately.
FIG. 1.—Reproductions of the architectonic maps of Brodmann (1909) (A), Walker (1940) (B)and Roberts and Akert (1963) (Q, and the map of Woolsey (1958) (D) showing the positions ofthe motor (MI) and supplementary motor (Mil) areas, the first (SI) and second (SII) somatic areasand the visual cortex (VI).
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from
796 E. G. JONES AND T. P. S. POWELL
The Outward Progression from the Somatic Sensory Cortex
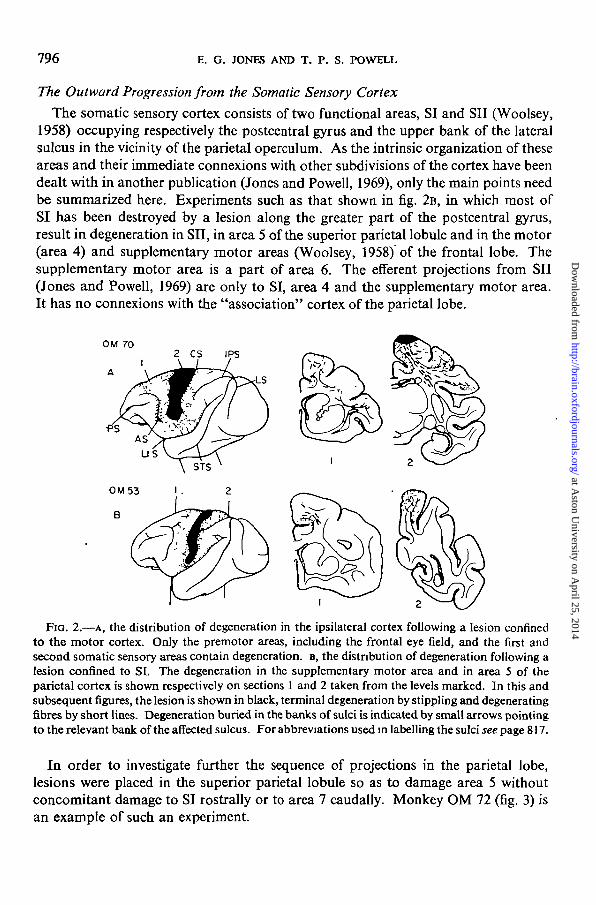
The somatic sensory cortex consists of two functional areas, SI and SII (Woolsey,1958) occupying respectively the postcentral gyrus and the upper bank of the lateralsulcus in the vicinity of the parietal operculum. As the intrinsic organization of theseareas and their immediate connexions with other subdivisions of the cortex have beendealt with in another publication (Jones and Powell, 1969), only the main points needbe summarized here. Experiments such as that shown in fig. 2B, in which most ofSI has been destroyed by a lesion along the greater part of the postcentral gyrus,result in degeneration in SET, in area 5 of the superior parietal lobule and in the motor(area 4) and supplementary motor areas (Woolsey, 1958) of the frontal lobe. Thesupplementary motor area is a part of area 6. The efferent projections from SII(Jones and Powell, 1969) are only to SI, area 4 and the supplementary motor area.It has no connexions with the "association" cortex of the parietal lobe.
OM 702 CS IPS
FIG. 2.—A, the distribution of degeneration in the ipsilateral cortex following a lesion confinedto the motor cortex. Only the premotor areas, including the frontal eye field, and the first andsecond somatic sensory areas contain degeneration, B, the distribution of degeneration following alesion confined to SI. The degeneration in the supplementary motor area and in area 5 of theparietal cortex is shown respectively on sections 1 and 2 taken from the levels marked. In this andsubsequent figures, the lesion is shown in black, terminal degeneration by stippling and degeneratingfibres by short lines. Degeneration buried in the banks of sulci is indicated by small arrows pointingto the relevant bank of the affected sulcus. For abbreviations used in labelling the sulci see page 817.
In order to investigate further the sequence of projections in the parietal lobe,lesions were placed in the superior parietal lobule so as to damage area 5 withoutconcomitant damage to SI rostrally or to area 7 caudally. Monkey OM 72 (fig. 3) isan example of such an experiment.
I
I
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

CORTICAL ASSOCIATION CONNEXIONS 797
OM 72 3 4
FIG. 3.—The lesion and consequent degeneration in Experiment OM 72 in which area 5 wasdamaged. The top row shows, from left, medial, dorsal and lateral views of the hemisphere. Thereare forward projections to the supplementary motor area and to an adjoining part of area 6, and alocal projection to area 7. Note sparing of the fundus of the caudal part of the intraparietal sulcusin sections 3 and 4 (buried part of area 19).
The lesion is largely confined to the cortex with only minimal invasion of underlying white matterat about its middle. Coarse degenerating axons descend into the white matter and are distributedto an adjacent area in the parietal lobe and to a second area in the frontal lobe. In the superiorparietal lobule, terminal degeneration occupies the cortex behind the lesion including the rostralbank of the sulcus formed by the union of the lunate, intraparietal and superior temporal sulci, butit does not reach the fundus of this sulcus so that area 19 of the visual cortex is spared. Lateral to thelesion, the degeneration in the cortex spreads rostrally along the superficial half of the medial bankof the intraparietal sulcus, sparing the fundus (area 19) except in the rostral one-third of the sulcuswhere thedegenerationcrosses the fundus toreachthelateral bank of thesulcus and the inferior parietallobule. The inferior parietal lobule contains sparse degeneration which extends into the caudal fewmillimetres of the lateral sulcus and barely into the upper bank of the superior temporal sulcus.All the degeneration in the parietal lobe is thus confined to areas 5 and 7,-the latter being the onlynew projection area in the parietal lobe. In the frontal lobe, degeneration is confined to the cortexforming the upper part of area 6 (Brodmann, 1909); most of the fine axonal fragments lie dorsal andmedial to the upper limb of the arcuate sulcus and spread on to the medial surface of the hemisphereto involve the whole supplementary motor area. Hence, area 5 projects to the same part of thepremotor cortex, the supplementary motor area, as the primary somatic sensory areas, plus oneother part —the part of area 6 lying above the arcuate sulcus. The remainder of area 6 lying on thelateral surface below and behind the lower limb of the arcuate sulcus is unaffected. There is noprojection from area 5 back into the somatic sensory areas, for no degeneration is found in thepostcentral gyms or opercular region.
As area 5 projects locally to area 7, the next step in the sequence was to damage area 7 selectively.OM 75 (fig. 4) is an example of such an experiment. The lesion is in the inferior parietal lobule wellbehind the somatic sensory cortex and there is only marginal involvement of underlying white
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

798 E. G. JONES AND T. P. S. POWELL
matter. There is degeneration in three main parts of the cortex: a local region in the parieto-temporal cortex, regions on the medial and inferior aspects of the hemisphere, and other regionsin the frontal lobe. Adjacent to the lesion, axonal degeneration involves the parts of area 7unaffected by direct damage. Medially.it extends into the lateral bank of the intraparietal sulcus andpasses through the rostral part of the fundus to reach the medial bank in which it ascends to thecaudal part of the superior parietal lobule and thence on to the medial surface as a small area aroundthe upturned caudal end of the cingulate sulcus. Area 5 is, therefore, uninvolved. Lateral to thelesion, fragments extend through the cortex lining the most caudal part of the lateral sulcus near thepoint at which it joins the superior temporal sulcus. This degeneration continues downwards in thecortex of the upper bank of the superior temporal sulcus to about its middle; there, it crosses thebroad, insula-like area of cortex in the fundus of the sulcus to occupy the rostral part of the lowerbank only, not quite reaching its anterior end (i.e. in order, sections 5, 4 and 3 in fig. 4). The
OM75
Fio. 4.—The lesion and degeneration in Experiment OM 75 in which area 7 was damaged.There is a local projection to the fundus of the superior temporal sulcus (sections 3-5), forwardprojections to the upper part of area 46 and to area 45, and medial projections to the cingulategyms and to area 27 of the parahippocampal gyrus.
majority of the degenerating fibres passing to this region in the superior temporal sulcus does so byway of the white matter. The region has not been mapped architectonically. Other degeneratingaxons are distributed to two disparate regions on the medial surface of the hemisphere: many of theaxons directed downwards towards the superior temporal sulcus by-pass the cortex therein andcontinue to the inferomedial aspect of the temporal lobe where they distribute terminal degenera-tion to the parahippocampal gyms. Most of the degeneration is in a strip medial and parallel tothe occipito-temporal sulcus, and stretching from the caudal end of the rhinal fissure to a pointjust anterior to the rostral tip of the calcarine sulcus. In architectonic terms, it corresponds mostclosely to area 35 of Brodmann and to field TH of Bonin and Bailey (interpreted in a preliminarycommunication (Jones, 1969) as area 27. Area 27 is, in fact, the subiculum and lies medial toarea 35). The other area of degeneration on the medial surface is in the cingulate gyms; fibresreach it from the lesion by passing in the white matter deep to the intraparietal sulcus. Most of theterminal degeneration is concentrated on the summit and lower part of the cingulate gyrus approxi-mately opposite the rostral half of the corpus callosum. It is difficult to assign this degeneration toany particular architectonic field as it appears to fall within parts of both areas 23 and 24 of
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from
CORTICAL ASSOCIATION CONNEXIONS 799
Brodmann. In the frontal lobe there is heavy terminal degenerationinthecortexliningthefundusandupper bank of the principal sulcus, especially about its middle one-third and chiefly occupying thedorsal half of area 46 of Walker (1940). Sparse degeneration, however, extends on to the dorsalsurface of the brain into Walker's area 9, in front of the upper end of the arcuate sulcus; a fewfragments even reach the medial surface around the rostral end of the cingulate sulcus. A secondfocus of degeneration is found in the lower part of the lateral surface of the frontal cortex, justrostra] to the lower end of the arcuate sulcus and probably occupying area 45 of Walker.
Frontal Connexions of the Somatic Sensory Cortical Pathway
It has been shown that SI and SII send fibres to the motor cortex (area 4) and to the supplementarymotor area of the premotor cortex; the next step in the succession of somatic sensory connexions inthe parietal lobe, area 5, projects to the supplementary motor area and to a further part of area 6;then, the third parieto-temporal step, area 7, projects to the prefrontal cortex—to a part of area 46,to area 9 and to area 45. Some of these regions in the frontal lobe have been damaged in a separateseries of experiments. After large lesions placed in the precentral gyrus so as to damage area 4(fig. 2A) the only degeneration found in the parietal lobe is in SI and SII (Jones and Powell, 1969).In the frontal lobe, lesions of area 4 cause terminal degeneration in the whole premotor cortex,including the whole of area 6 (Brodmann), the supplementary motor area and the precentral agranu-lar field (PrCo, Roberts and Akert, 1963), together with the frontal eye field, area 8A (Walker), inthe rostral bank of the arcuate sulcus and area 8B above it (fig. 2A). Hence, the somatic sensoryareas and area 4 are interlocked by reciprocal pathways across the central sulcus; then SI projectsbackwards to another area in the parietal lobe (area 5) and area 4 forwards to new areas (area 6 etc.)in the frontal lobe.
Lesions affecting the other frontal or premotor projection of the somatic sensory areas, the supple-mentary motor area (OM 71, not illustrated), do not cause degeneration in the somatic areas(Jones and Powell, 1969) so that, like the projection from SI to area 5, this is a non-reciprocalconnexion.
I
I
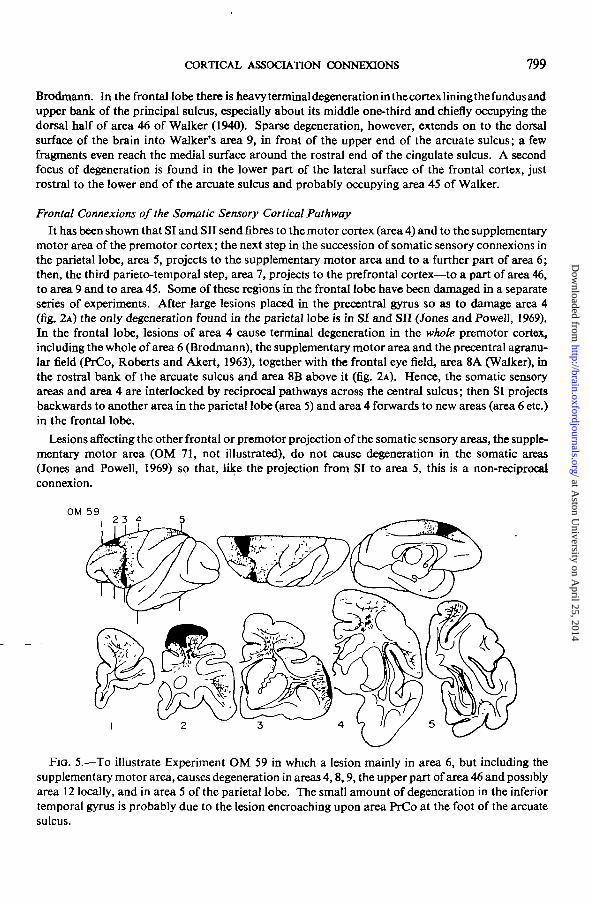
OM 59I 2 3 4
FIG. 5.—To illustrate Experiment OM 59 in which a lesion mainly in area 6, but including thesupplementary motor area, causes degeneration in areas 4,8,9, the upper part of area 46 and possiblyarea 12 locally, and in area 5 of the parietal lobe. The small amount of degeneration in the inferiortemporal gyrus is probably due to the lesion encroaching upon area PrCo at the foot of the arcuatesulcus.
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

800 E. G. JONES AND T. P. S. POWELL
The next experiment (OM 59, fig. 5) shows that areas 5 and 6 are interconnected in the samemanner as are the somatic sensory areas and area 4. In this brain there is a double lesion mainlyconfined to area 6; its upper part stretches from the supplementary motor area at the anterior endof the marginal gyrus medially, to the upper limb of the arcuate sulcus on the dorsal aspect of thehemisphere laterally. The second half of the lesion lies below and behind the lower limb of thearcuate sulcus and probably just encroaches upon area PrCo at the lower end of the sulcus. Thereare, in addition, two tiny adhesions damaging the most superficial part of the molecular layer of thecortex over area 8A in front of the arcuate sulcus. These appear to be so superficial as to beunimportant. In this brain there is degeneration throughout the cortex of areas 6 and 4 and there aretwo other main areas of terminal degeneration in the cortex—in the frontal and in the parietal lobes.A bundle of coarse degenerating fibres sweeps posteriorly under the central sulcus in the whitematter deep to the postcentral gyrus and distributes heavy degeneration to the cortex of area 5 inthe superior parietal lobule. In the frontal lobe, degeneration extends forwards from the upperlesion into areas 8B and 9 (Walker) and is especially heavy in the upper part of area 46 in the upperbank and adjoining cortex above the principal sulcus—in the same area affected by lesions of area 7(cf. figs. 4 and 5). There is also degeneration in area 8A and further degeneration extends forwardsfrom the lower lesion to involve mainly area 45 at the lower end of the arcuate sulcus, although afew fragments encroach upon the part of area 46 lying below the principal sulcus and there may bea few in area 12 (Walker). A small number of degenerating axons are also seen in area 20 of thetemporal lobe; these are probably caused by the lesion encroaching upon area PrCo {see below).
The next experiment (OM 78, fig. 6) shows that as with areas 6 and 5, so the upper part of area46 and area 7 are interconnected. In this brain, the lesion is primarily in areas 8B and 9 but thefrontal projection of area 7 in and adjacent to the upper bank of the principal sulcus is undercut byan extension of the lesion into the white matter. Degenerating axons can be traced through thewhite matter deep to the central sulcus into the caudal parts of the superior and inferior parietal
FIG. 6.—A lesion involving mainly area 9 but with undercutting of the upper part of area 46 andcausing degeneration in area 45 and the frontal pole locally, in areas 7 and 22 in the parietal an'dtemporal lobes, and in the anterior cingulate and retrosplenial areas on the medial surface. Thedegeneration in area 7 is due solely to undercutting of area 46.
I
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

CORTICAL ASSOCIATION CONNEXIONS 801
lobules in which they distribute terminal degeneration to area 7. Although there is degenerationelsewhere in this brain, the degeneration in the parietal lobe is due to undercutting of the upper partof area 46, as shown by other brains such as OM 69 (fig. 7) in which areas 8B and 9 but not area 46are affected. These cause degeneration in the same regions as OM 78 with the exception of area 7.Lesions affecting area 9 such as OM 78 (fig. 6) and OM 69 (fig. 7) in which the frontal pole (area 10)is also slightly damaged, cause additional degeneration in parts of the cingulate cortex, particularlyin area 24 about the genu of the corpus callosum and in the retrosplenial cortex, i.e. mainly inregions in front of and behind the cingulate projection of area 7. Fibres reaching the retrosplenialcortex pass backwards in the cingulum. The main rostral projection of areas 9 and 46 appears tobe to the frontal pole (area 10 of Walker) and to area 45 at the foot of the arcuate sulcus (figs. 6 and7) although our material is insufficient for analysing this in detail. There is no brain with a lesionin the other rostral projection of area 7, area 45.
OM 69
FIG. 7.—A lesion of area 9 with additional involvement of area 10 at the frontal pole. Thedegeneration on the orbital surface of the frontal lobe and at the temporal pole is apparently dueto involvement of area 10, while that in area 45 and in the superior temporal gyms (area 22) ispresumably caused by the damage of area 9 (cf. fig. 6).
The Outward Progression from the Visual Cortex
As this has been described in some detail by Kuypers et al. (1965), Cragg (1969) and Zeki (1969).only certain significant points will be mentioned. Lesions confined to the area striata (area 17) suchas that in experiment OM 62 (fig. 8A) cause degeneration in a circumstriate belt incorporating areas18 and 19 (Brodmann) and extending for some distance along the fundus of the intraparietal sulcus(area 19); there is also an additional focus of heavy and coarse degeneration in the middle portionof the caudal bank of the superior temporal sulcus, and in the frontal lobe there are a few degenerat-ing fibres in the cortex of area 8A immediately in front of the arcuate sulcus. When a lesion such asthat in OM 66 (fig. 8B) involves areas 18 and 19 as well as area 17, there is an additional localprojection to area 20 (Brodmann) so that the inferior temporal gyrus becomes filled with degenerat-ing fragments from the anterior end of the inferior occipital sulcus on the lateral surface to theoccipito-temporal sulcus on the inferior surface. Furthermore, a definite focus of degeneration
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

802 E. G. JONES AND T. P. S. POWELL
I
FIG. 8.—A, The lesion and degeneration in Experiment OM 62. Here, the striate cortex (area 17)only, has been damaged. Note the degeneration in areas 18 and 19 and the additional dense focusin the middle of the caudal bank of the superior temporal sulcus. B, The lesion and degeneration inOM 66. In this case, there is destruction of parts of areas 17, 18 and 19. Degeneration spreadslocally into area 20 and there is a projection to area 8A in the frontal lobe. The sections relate toOM 66 only.
now becomes obvious in the frontal lobe, filling the frontal eye field, area 8A, in the rostral bankof the concavity of the arcuate sulcus.
Area 20 was damaged by lesions of the inferior temporal gyrus such as that in OM 68. Thelesion (fig. 9) which is situated between the anterior middle temporal sulcus and the rostral end of
0M68
I
I
I 2
FIG. 9.—Experiment OM 68 in which a part of area 20 was damaged. Local degenerationinvolves area 21 and spreads medially into the lateral part of the entorhinal cortex but there isno projection back to the visual areas. Frontal degeneration affects areas 8A and PrCo. In thisbrain there is heavy degeneration in the basolateral group of amygdaloid nuclei (section 3).
I
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

CORTICAL ASSOCIATION CONNEXIONS 803
the occipito-temporal sulcus is confined to the cortex except at its middle, where there is slightinvasion of white matter. From this lesion, degeneration fills area 20 and spreads locally into area 21of the middle temporal gyrus but not back into areas 17,18 or 19, and there is a projection forwardsto the frontal lobe. This latter includes area 8A and one new premotor field, area PrCo, immediatelyposterior to the lower end of the arcuate sulcus. A further striking feature of brains with lesions ofarea 20 is the presence of fine, but quite dense degeneration in the basolateral group of amygdaloidnuclei. Also in brains with lesions of area 20, a few degenerating fragments extend medially beyondthe boundanes of areas 20 and 21 to reach the cortex of what is probably area 35 or TH along thelateral margin of the entorhinal cortex at the level of the caudal end of the rhinal fissure.
The next step in the local progression from the visual cortex, area 21, was damaged withoutinvolvement of white matter in experiments such as OM 67 (fig. 10). This shows no projection backto area 20 (cf. area 7 which does not project back to area 5), but instead sends fibres to the tip of thetemporal lobe and to a part of the cortex buried in the superior temporal sulcus, as well as to theprefrontal cortex. The tip of the temporal lobe contains fragmented axons in a region incorporatingthe rostral few millimetres of the superior temporal gyrus, the degeneration becoming continuouscaudally with that in area 21 itself. The tip of the temporal lobe was not recognized as a separatefield in the monkey by Brodmann but is called area TG by Bonin and Bailey (1947). In the superiortemporal sulcus, the region affected is largely co-extensive with that which contains degenerationafter a lesion of area 7 (cf. fig. 4) although it is heaviest in the lower and posterior parts of the sulcuswhere the buried insula-like cortex of the fundus meets the caudal bank. This region is deeper in thesulcus than the direct projection of areas 17, 18 and 19 to the caudal bank; the latter region is freeof degeneration in this brain (large arrow in fig. 10). Thus, somatic and visual cortical pathwayscome together and overlap in the depths of the superior temporal sulcus. In all brains with lesions
OM 67
FIG. 10.—The lesion and the extent of the degeneraton in Experiment OM67. Area 21 is affectedand degeneration spreads locally into the temporal pole, the lateral pan of the entorhinal cortexand particularly into the fundus of the superior temporal sulcus where it overlaps with the regionshowing degeneration after lesions of area 7 (cf. fig. 4). The unaffected projection area of the visualcortex is shown by the large arrow in section 4. In the frontal lobe, the degeneration is confined tothe lower part of area 46 on the lateral surface and to area 25 on the medial surface.
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

804 E. G. JONES AND T. P. S. POWELL
of area 21 there is a little degeneration in the cortex of the walls of the rhinal sulcus immediatelyadjoining the lateral part of the entorhinal cortex and a very occasional fragmented fibre in thebasolateral nuclei of the amygdala. In the frontal lobe, there is heavy degeneration in the part ofarea 46 (Walker) lying below and in the lower bank of the principal sulcus. The fibres reach thisarea by way of the uncinate fasciculus. The degeneration in this area does not overlap with theupper part of area 46 which receives fibres from area 7. Also, on the medial aspect of the frontallobe there are a few degenerating fibres within area 25 of the cortex about the rostral sulcus.
Frontal Connexions of the Visual Cortical Pathway
It has been shown that as with the somatic sensory projections to the frontal lobe, so in theconnexions of the visual areas there are two premotor and prefrontal "steps." Thus, areas 17, 18and 19 project to area 8A, area 20 to areas 8A and PrCo, and area 21 to the lower part of area 46and to area 25. It has not been possible to damage area 8A selectively so that the further connexionsof this area are to some extent unknown. Large lesions of areas 4 and 6 (not illustrated) whichundercut it have not been seen to cause degeneration in the temporal or occipital lobes so that itmay not project back to areas 18 and 19 or to area 20.
There is, similarly, no experiment with a lesion strictly confined to area PrCo, but it may bededuced that it sends fibres back to area 20, for marginal involvement of area PrCo by the lesionin experiment OM 59 (fig. 5) caused sparse degeneration in area 20. If this is so, areas 20 and PrComay be akin to areas 5 and 6 in the somatic system. The part of area 46 lying below the principalsulcus does not receive degenerating fibres after lesions of area 6 proper but the few fragments whichit contains in experiment OM 59 suggests that it may receive from field PrCo.
Lesions in the lower, "visual" part of area 46 project back primarily to area 21. This is obviousfrom the work of Kuypers et al. (1965) and of Pandya and Kuypers (1969) and can be demonstratedby exclusion from experiments OM 78, 69 and 61 (figs. 6, 7 and 11). All have lesions involving
0M6II
I 2FIG. 11.—The lesion and degeneration in Experiment OM 61. Most areas on the dorsolateral
aspect of the frontal lobe in front of the arcuate sulcus have been destroyed and there is degenerationin their parieto-temporal projection areas: areas 7, 22, 21", the temporal pole and in the depths ofthe superior temporal sulcus.
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

CORTICAL ASSOCIATION CONNEXIONS 805
I
I
I
several areas of the frontal cortex but only that of OM 61 (fig. 11) affects the lower part of area 46and only this lesion causes degeneration in area 21 of the temporal lobe. In the frontal lobe thefurther connexions of the lower part of area 46 appear to be to the frontal pole and to the cortex atthe foot of the arcuate sulcus which includes area 45 and the orbito-frontal cortex of Roberts andAkert (1963) (see fig. 11, and fig. 1 of Pandya and Kuypers, 1969).
The Outward Progression from the Auditory Cortex
It has not been possible to damage the auditory cortex (areas 41 and 42) without concomitantdamage to either the superior temporal gyms (area 22) or the parietal operculum (SII). There issome evidence, however, that the auditory areas project to the supratemporal plane (i.e. the upper"surface of the temporal operculum, or area TB of Bonin and Bailey, 1947) (Hurst, 1959; Pandya,Hallett and Mukherjee, 1969). Moreover, auditory evoked potentials are obtained from both thesupratemporal plane and from area 22 (Ades and Felder, 1942; Pribram, Burton and Rosenblith,1954) and the supratemporal plane is said to "fire" area 22 (Bonin and Bailey, 1947). It appearspossible, therefore, that the auditory cortex sends fibres to the supratemporal plane and that thelatter in turn projects to area 22. A lesion such as that in OM 82 (fig. 12) which involves a portion of
OM 82
FIG. 12.—The lesion affecting the supratemporal plane and area 22 of the para-auditory cortexin Experiment OM 82. The ensuing degeneration in the frontal lobe involves areas 8B, 9, 10 and12.
the supratemporal plane together with area 22 and possibly encroaches on the primary auditorycortex causes degeneration throughout area 22 locally and in three regions of the frontal lobe. Thereis degeneration at the frontal pole (area 10 of Walker), extending back on the medial surface intoarea 25; other degeneration fills area 12 where the lateral and orbital surfaces of the frontal lobebecome continuous; the third focus of degeneration occupies a region around the rostral end of theupper limb of the arcuate sulcus including area 8B, possibly affecting area 9 as well and extendingrostrally to reach the anterior end of the principal sulcus.
The superior temporal gyms (area 22) was damaged alone in experiments such as OM 65 (fig. 13).In this brain there is degeneration throughout area 22 locally and this extends into the small regionat the tip of the temporal lobe as well as area 35 or TH along the lateral margin of the entorhinalcortex. In addition there is terminal degeneration in the cortex lining the fundus of the superiortemporal sulcus approximately co-extensive with the region affected after lesions of areas 7 and21 but heaviest inferiorly. In the frontal lobe, only areas 10, 25 and 12 contain degeneration so thatit may be concluded that it is the supratemporal plane which is projecting to area 8B and possibly toarea 9 in the frontal lobe and from the literature cited above it is probable that it sends fibres locallyto area 22. Area 22, therefore, is probably the next step, projecting to two new prefrontal areas and
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

806 E. G. JONES AND T. P. S. POWELL
0M65
Fia. 13.—The lesion ccnfined to area 22 in Experiment OM 65. The degeneration in the temporallobe affects the fundus of the supenor temporal sulcus in a region overlapping with that seen afterlesions of areas 7 and 21 so that there is a convergence of auditory, somatic and visual pathways inthis region. A little further local degeneration affects the lateral part of the entorhinal cortex. Inthe frontal lobe, areas 10, 25 and 12 are affected and the degeneration in the latter may extend intoarea 45. There is no degeneration in areas 8A or 9 (cf. fig. 12).
to new local areas at the tip of the temporal lobe and in the fundus of the superior temporal sulcus.Thus, somatic, visual and auditory paths are brought together in the depths of the superior temporalsulcus. In the frontal lobe of this brain with a lesion of area 22, it is difficult to exclude that thedegeneration in area 12 does not encroach upon the orbito-frontal cortex although the rostral partof the fronto-parietal operculum which contains the greater part of the orbito-frontal cortex, isclear of axon fragments.
Frontal Connexions of the Auditory CortexThe material is not adequate for describing the further connexions of the auditory pathway in the
frontal lobe. It can be shown from experiments OM 78 (fig. 6), OM 69 (fig. 7) and OM 61 (fig. 11),however, that lesions involving the more rostral parts of the frontal lobe and including area 9 causedegeneration in area 22 but not those involving area 10 (OM 81, fig. 14).
Regions of Convergence and Further Connexions of the Frontal and Temporal LobesThe small amount of evidence available from the large lesions of the frontal lobe suggests that
areas in front of the arcuate sulcus project forwards to the frontal pole (area 10) and downwards to aregion including area 45 and the orbito-frontal cortex of Roberts and Akert (1963) at the lower endof the arcuate sulcus and frontal operculum. Therefore, these may also be regions of convergenceof all three intracortical pathways. When lesions such as that in OM 81 (fig. 14) involve area 10(in this case together with area 11) projections can be demonstrated back to areas 9,45,46 and 12 ofthe frontal lobe, but with the possible exception of area 8A, not to the premotor or motor cortex.In addition, there are certain new fields in the frontal lobe which contain degeneration, notably thewhole orbito-frontal cortex and all areas (11, 13 and 14) on the orbital surface, together with areas24 and 25 of the rostral cingulate cortex. The degeneration in the orbito-frontal cortex is the
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

CORTICAL ASSOCIATION CONNEXIONS 807
I heaviest and most extensive seen in this area in all the available brains and reaches back to the rostralend of the insula (fig. 14). Degenerating fibres can also be traced through the uncinate fasciculus tothe tip of the temporal lobe, and a more dorsally situated group in the external capsule passes to thefundus of the superior temporal sulcus where they distribute degeneration to the upper part of therostral bank and to the lower parts of the fundus and caudal bank in a region approximatelyco-extensive with that affected after lesions of areas 7, 21 and 22, although most of the heavydegeneration is in the upper half.
OM ei
I
2 3 4
FIG. 14.—The lesion affecting area 10 at the frontal pole and the ensuing degeneration inExperiment OM 81. There is local degeneration throughout the lateral, orbital and medial surfacesof the frontal lobe. That on the frontal surface extends into the anterior limbic cortex, while thaton the lateral surface spreads into area 45 and the orbito-frontal cortex of the frontal operculum.In the parieto-temporal region, there is degeneration at the temporal pole and in the region ofconvergence in rhe depths of the superior temporal sulcus.
I
lesions at the temporal pole (area TG of Bonin and Bailey, 1947) (fig. 15) such as that in OM 80send degenerating fibres into many of the same regions as the frontal pole. Degenerating fibres"passing forwards in the uncinate fasciculus reach the frontal lobe and distribute terminal degenera-tion to area 10 at the frontal pole, to area 24 on the medial surface, to areas 11, 13 and 14 on theorbital surface, and to the orbito-frontal cortex and area 45 on the lateral surface and frontaloperculum. It is difficult to exclude that some of the latter degeneration mayencroach upon area 12.Locally, in the temporal lobe all degeneration is confined to the immediate vicinity of the lesion.There are a few fragments in the cortex of area 35 (TH) adjoining the lateral part of the entorhinalcortex but areas 21 and 22 are free, as also is the region of convergence in the depths of the superiortemporal sulcus. This latter region is so deeply situated that it was not possible to place a lesion init and nothing can be said about its efferent connexions. Similarly, area 45, the orbito-frontalcortex and the areas on the orbital surface of the frontal lobe have not been damaged selectively.Nauta (1964) notes that fibres pass from the orbital surface to the rostral part of the temporal lobeand that in addition many reach the lateral hypothalamus.
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

808 E. G. JONES AND T. P. S. POWELL
0M80,4 5 .6
FIG. 15.—The lesion at the temporal pole in Experiment OM 80 and the distribution of theresultant degeneration. Apart from local degeneration in the immediate vicinity of the lesion, alldegeneration is in the frontal lobe, particularly in areas 10-14 on the orbital surface, area 24 on themedial surface and the orbito-frontal cortex on the lateral aspect and in the frontal operculum.There is a small amount of degeneration in the basolateral amygdaloid nuclei (section 5).
DISCUSSION
Association connexions joining the frontal, parietal and temporal lobes have beenknown for many years from work with the Marchi technique (Mettler, 1935; Jansen,1937; Krieg, 1963). Recently there has been renewed interest in such connexionsand a number of studies have appeared in which the more sensitive Nauta techniquehas been used (Nauta, 1964; Kuypers et ai, 1965; Pandya and Kuypers, 1969;Jones, 1969) and there has been at least one evoked potential study of the frontalprojections of area 22 (Bignall, 1969). As these authors, and particularly Geschwind(1965) have reviewed the earlier literature in detail, a further summary does not seemwarranted.
The present material is clearly unsatisfactory for a total analysis of the intracorticalpaths of the three sensory systems because there is a lack of experiments with smalllocalized lesions in separate fields of the frontal lobe. Although not impossible, suchlesions would be difficult to obtain because of the variability in different brains oflandmarks such as sulci, and because of the problem of undercutting fibres passing toand from adjoining areas. However, certain general principles do seem to emergeand we are putting these forward provisionally, as possibly forming a backgroundfor one of the likely mechanisms of sensory integration within the cerebral hemi-sphere. The main points which appear to emerge are as follows: in each of the threesystems studied there is a stepwise, outward progression from the main sensoryareas within both the parieto-temporal and frontal lobes, with an interlocking of eachnew parieto-temporal and frontal step. In the somatic sensory and visual systems,
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

CORTICAL ASSOCIATION CONNEXIONS 809
I and probably also in the auditory system, each primary area projects to a local area inthe same lobe and to a portion of the premotor cortex in the frontal lobe. For example,in the somatic sensory pathway, the primary cortex projects locally to area 5 andforwards to the supplementary motor area. The local projection area then sendsfibres to a new local area and forwards to the previous premotor area, as well as to anew part of the premotor cortex. The frontal areas are then interlocked by reciprocalconnexions with the first local projection area of the primary sensory cortex. Thus,in the somatic system, area 5 projects to area 7 in the parietal lobe, and to the supple-mentary motor area and an additional part of area 6 in the frontal lobe; areas 5 and6 are then interconnected. The sequence is now repeated by projections to furtherlocal and prefrontal fields which are again interconnected. Hence, in the somaticsystem, area 7 projects locally to the floor of the superior temporal sulcus androstrally to the upper part of area 46; area 46 then returns fibres to area 7. The newprefrontal fields also receive from the premotor fields of the previous step: thus,area 46 receives fibres from area 6. Finally, when fibres cease being sent back directlyto the motor and premotor areas from the frontal and parieto-temporal cortex, the
SOMATIC
TG
AUDITORY
FIG. 16.—A schematic diagram summarizing the outward progression of connexions from theprimary somatic, visual and auditory areas of the cortex. Each new local step is shown in blackand the further connexions of the new areas by light stippling or hatching. Notice that allsensory pathways converge in the depths of the superior temporal sulcus (STS).
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

8 1 0 E. G. JONES AND T. P. S. POWELL
three sensory pathways commence sending fibres into the limbic cortex, from boththeir frontal and parieto-temporal components; they now begin to converge withinthe cortex of both the frontal and parieto-temporal lobes. Most of the findings are ingeneral agreement with those of Pandya and Kuypers (1969), but we have beenunable to demonstrate a projection from the supplementary motor area back to thesomatic sensory cortex. As Pandya and Kuypers have discussed the organization ofthe association connexions with particular reference to the origin of the projectionto the precentral gyms and premotor area, the present discussion will concern itselfmainly with the possible significance of the association connexions in relation tointracortical convergence of the sensory pathways.
The three main sensory areas (and here SI and SII will be considered to form thesomatic cortex, areas 17, 18 and 19 together as visual cortex and areas 41 and 42 asauditory cortex) have certain similarities and one important difference. Each has alocal and a premotor projection, neither of which is a reciprocal one. But the visualand auditory areas are isolated from the rest of the cortex in not receiving any fibresreturning from elsewhere, whereas the somatic sensory cortex has a heavy input fromthe motor cortex (area 4); the two-way connexions between the somatic and motorcortices are probably a reflection of the close integration of somatic sensory andmotor activities from the level of the simple spinal reflex upwards.
Apart from the possibility of a degree of sensory interaction occurring in the motorcortex, the three cortical sensory pathways are kept entirely separate across a numberof synaptic linkages in both the parieto-temporal and frontal lobes. The presentobservations, together with those made in other studies (Kuypers et al., 1965;Diamond et al., 1968; Cragg, 1969; Jones and Powell, 1969), showing the restrictionof the initial projections from the primary areas to distinctly separate fields, issupported by clinical and experimental psychological observations. It is known, forexample, that the initial local projection areas of the visual cortex, areas 20 and 21,are concerned largely if not exclusively with visual discriminations {see Chow, 1961;Mishkin, 1966; Weiskrantz, 1968). Similarly, that of the auditory cortex, area 22, isconcerned with auditory discriminations (Massopust et al., 1965; Dewson and Cowey,1969) and more caudal parts of the parietal lobes (the initial projections of thesomatic cortex) with somatic sensory activities (Moffett et al., 1967). The differencesin the nature of the visual discrimination defect resulting from anterior as opposed toposterior lesions of the inferotemporal cortex {see Weiskrantz, 1968) suggests thatfurther functional subdivision of the local projection areas could depend upon ananalysis of the deficits accruing from lesions of the individual architectonic fields;lesions of the anterior inferotemporal region should affect primarily area 21 whilethose of the posterior inferotemporal region should affect mainly area 20. Each ofthese areas, although strongly connected with the visual cortex, has a different patternof cortical connexions.
There has been a considerable number of behavioural studies devoted to elucidat-ing the part played by the prefrontal granular cortex in discrimination and learning.The current status of these studies is reviewed by Gross and Weiskrantz (1964),
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

CORTICAL ASSOCIATION CONNEXIONS 811
I Mishkin (1964, 1966) and Rosvold and Szwarcbart (1964). Of significance to thepresent study are the observations that bilateral lesions involving the banks of theprincipal sulcus lead to severe impairment in tests of the acquisition and retentionof a delayed response, irrespective of the sensory modality employed (Rosvold andMishkin, 1961; Gross and Weiskrantz, 1964). In the present study, the banks of theprincipal sulcus have been shown to receive fibres of auditory (rostrally), somatic(dorsally) and visual (ventrally) provenance, so that any lesions of this region mightbe expected to cause deficits relating to all three sensory systems. Gross and Weis-krantz (1964) found that bilateral lesions about the principal sulcus also produceeffects upon certain visual discriminations which may be attributed to an inability toutilize recent information. They found, further, that a defect of auditory discrimina-tion can be produced from bilateral lesions which spare the principal sulcus, suggest-ing that some other region is more critical for this. Finally, Mishkin (1966) notes
Fio. 17.—A schematic diagram showing the stepwise progression of connexions of the somatic(above) and visual (below) pathways in the frontal lobe and the reciprocal connexions withequivalent steps in the parietal and temporal lobes (cf. fig. 16). Note the regions of convergence inarea 10 and the orbito-frontal cortex.
I
that bilateral lesions of the cortex ventrolateral to the principal sulcus are the mostefficacious in producing defective visual discrimination. If the present observationshave any bearing upon this subject, then it might be possible to dissociate defects ofsomatic and visual discrimination by lesions respectively in the upper and lowerbanks of the principal sulcus and auditory defects by lesions of area 12, for theseare the equivalent steps in the chain of frontal projections from each of the threeintracortical sensory pathways.
The defects resulting from lesions of the local and frontal projection areas suggestthat they are involved in more integrative functions than the primary sensory areas,for the presence of the former is necessary for the performance of more subtle andcomplex tasks than the primary cortex. This is not to say that the primary cortex may
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

812 E. G. JONES AND T. P. S. POWELL
not itself be involved in quite complex discriminations, for Diamond and Hall (1969)have shown that the primary visual cortex of the tree shrew is essential for solving adiscriminative problem requiring the abstraction of the relevant stimulus (a triangle)from a masking circle. In the absence of the visual cortex, the remaining "associa-tion" cortex enables the animal to perform visual pattern discriminations. It may beargued, however, that the ability to discriminate a total pattern must require theintegration of many simple pattern units, whereas the ability to abstract one simplepattern from another does not. Apart from a single investigation showing auditoryevoked responses in the superior, and visual responses in the inferior temporal gyrus(Gross, Schiller, Wells and Gerstein, 1967), there have been no detailed microelectrodestudies of single neurons in the association cortex of the monkey. However, Hubeland Wiesel (1965) note that in the primary visual cortex of the cat (areas 17, 18 and19), in passing from area 17, through area 18 to area 19, the receptive fields of singleneurons become less specific in regard to stimulus placement upon the retina. Hubeland Wiesel have proposed that this indicates a convergence of axons derived frommany neurons with much simpler, though more specific receptive field properties as anecessary prerequisite for perceiving a total image. The apparent reduction of topo-graphic segregation shown by the projections passing from the primary to the associa-tion cortex in this and other studies may reflect a similar convergence, albeit at a verygross level. Thus, in the somatic sensory path, the connexions between the primarycortex and the motor and premotor areas maintain the topographic representationof the body surface exhibited by these areas. For example, the hind-limb sub-division of SI is only interconnected with the hind-limb subdivisions of SII and of themotor and supplementary motor areas (Jones and Powell, 1968, 1969). The topo-graphic parcellation tends to be maintained in the projection of SI to area 5 but heresome overlap is beginning (Jones and Powell, 1968, 1969) and it may be expected thatthis reflects the commencement of a bringing together of information from differentparts of the body surface preparatory to building up a total body scheme. In pro-gressing beyond the immediate local projection areas into the posterior parietalregion and into the prefrontal cortex, the topographic parcellation seems to breakdown and at this juncture all parts of the image of the body surface may be weldedinto one. Convergence within the somatic sensory system, therefore, seems to pre-cede convergence between it and the other two systems. Because we have no experi-ments with small lesions confined to different parts of the peripheral representationin the primary visual and auditory areas, it is not known whether, like the somaticsensory system, there is a progressive convergence of parts of the representation, butsuch a possibility seems by no means unlikely.
The significance of the double projection pattern—Ihe one local and the other tothe frontal lobe—shown so strikingly by all three sensory systems is obscure but itlends itself to certain speculative considerations. In the visual system, according toTeuber (1966) the visual cortex is responsible for the immediate presentation ofvisual space, while the frontal cortex is necessary for compensation for changes inspatial order occurring during voluntary changes of posture; the parietal cortex is
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

I
I
I
I
CORTICAL ASSOCIATION CONNEXIONS 813
responsible for representation, i.e. for dealing with spatial relations not immediatelygiven, such as route finding by means of maps. It may be, therefore, that in allsystems the frontal projection areas are more concerned with praxic functions orsensorimotor integration, and this would probably explain their connexions with themotor cortex. In adjusting to displacement of the visual field by prismatic spectacles,it is essential for the subject to walk about and manipulate his environment, for theadjustment cannot be made if he is passively pushed about in a wheelchair {seeTeuber, 1966). Moreover, the defects in form and depth discrimination exhibited bygrowing kittens passively carried about their environment appear to be at a levelhigher than the primary visual pathways (Teuber, 1966). Each of these cases couldbe an expression of a functional disconnexion of areas 46, PrCo and 4. By contrast tothe above, the local projections may have more to do with gnostic functions or theawareness of spatial and other relationships. These speculations would, of course, bein keeping with the known deficits resulting from frontal and parietal lesions in man.It must be remembered, however, that every step in the sequence of connexions inone lobe is reciprocally connected with the equivalent step in the other lobe. Thestrong interconnexions at all levels between the frontal and parieto-temporal fieldsmay explain why in many cases of parietal lobe lesion, especially if the supramarginalgyrus is involved, an apraxia as well as a complex agnosia may result (Critchley,1953; Brain, 1961). Similarly, the ataxia with postural loss which may follow lesionsof the paracentral lobule (area 5) (Critchley, 1953) could be due to disconnexion ofareas 5 and 6. In the primary sensory areas of the somatic system at least, there issome evidence that the different subdivisions may be concerned with both or withonly one of these activities. Thus, the cortical outflow from SI is to both the motorand the parietal cortex whereas that of SII is to the motor areas alone (Jones andPowell, 1968, 1969). It would be interesting to know whether the dual (or more)ascending pathways in all three sensory systems are in some way a reflection of theduality exhibited by the intracortical pathways.
The three sensory systems start to converge beyond the point at which the steps inthe sequence of cortical projections cease having immediate connexions with themotor or premotor cortex. The most obvious regions for convergence of all threesystems are in the depths, particularly of the upper two-thirds, of the superiortemporal sulcus, in orbito-frontal cortex at the lower end of the arcuate sulcus andextending into the frontal operculum, and at the frontal pole. In some areas, suchas the temporal pole, area 9 and area 25, only two paths may come together in thefirst instance, but all are ultimately interconnected with one another and with theregions of full convergence. The regions of convergence in the depths of the superiortemporal sulcus and at the lower end of the arcuate sulcus are of interest from thepoint of view of their possible homologies to regions of clinical significance in man.From its position and relation to other cortical areas, the area for convergence in thefundus of the superior temporal sulcus should represent the homologues of theangular and supramarginal gyri and contain areas 39 and 40 (Brodmann, 1909).Lesions of these areas are of course the most critical for the production of agnosias,
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

814 E. G. JONES AND T. P. S. POWELL
and particularly on the dominant side for causing dyslexias, agraphia and otherdramatic forms of aphasia (see Critchley, 1953; Brain, 1961; Geschwind, 1965). Thegeneral consensus throughout the literature is that areas 39 and 40 are rudimentaryin non-human primates (Critchley, 1953; Geschwind, 1965) although the exactevidence upon which this is based is not clear. In the absence of knowledge con-cerning the possibility of dominance in the non-human brain and with no evidencethat a Wernicke's or speech area exists in subhuman species, it would clearly beunwise to carry the argument further. However, it would appear that a region ofconvergence such as that demonstrated in the present study could furnish a necessarysubstratum out of which in the course of further cerebral evolution, a region devotedto "language and symbolic thought" (Critchley) could arise. Clearly, in the homo-logous regions of the human brain, more than one functional area may exist, forGeschwind (1965) distinguishes the effect of lesions in the angular gyms from thoseof Wernicke's area proper and Penfield (1959) notes that "speech cortex" (memoriesfor words) does not overlap with "interpretative cortex" (past similar experience).Similar considerations apply to the area of convergence at the lower end of thearcuate sulcus and orbital surface, for this region should be homologous to areas44, 45 and 47 which on the dominant side include Broca's area. As this region iswithin the projection of the mediodorsal nucleus of the thalamus (Akert, 1964), itmay be that olfactory information passing through the latter nucleus (Powell, Cowanand Raisman, 1965) is also merged with the other sensory pathways at this point.
It is noteworthy that a number of the regions of convergence in the frontal lobeappear to be identical in situation to the areas which in the squirrel monkey, andapparently also in the Macaque (Bignall and Imbert, 1969), show polysensory con-vergence of all three modalities. Bignall and Imbert (1969) have recognized foursuch areas: one in front of and one behind the arcuate sulcus which are probably theequivalents of areas 9 and 6 respectively; another in the frontal operculum andanterior insular area ("area 44"); a fourth on the orbital surface. The last two are ofinterest as lying in the vicinity of the final regions of convergence demonstrated inthe present study. If the pattern of connexions shown by our experiments has anyrelevance to the functional basis of these polysensory areas, it would be necessary topredict that the pre- and post-arcuate areas represent lower "levels" of organizationthan the fronto-opercular and orbital areas, for the former two are on more directpaths from the primary sensory areas and the latter two represent regions of laterconvergence. This is not to say that cortico-cortical connexions are the only pathwaysconcerned in the functioning of these polysensory areas, or of any of the regions ofconvergence. Indeed, Bignall and Imbert (1969) have shown that ablation of theprimary areas does not completely abolish the polysensory responses in the frontallobe.
As the three sensory pathways begin to converge within the cortex, so the outputfrom each new area becomes more and more separated from the motor cortex and isdirected into the limbic system. The third step in the chain of local projections, i.e.areas 7 (somatic), 21 (visual) and 22 (auditory), each send fibres to the cingulate
I
I
I
I
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

I
I
I
I
CORTICAL ASSOCIATION CONNEXIONS 815
cortex (areas 23, 24 and 25) and to a part of the parahippocampal gyms; the latteralso receives fibres from an earlier step in the visual pathway, area 20. In the case ofthe parahippocampal gyrus, area 7 projects most caudally and the other two sendfibres more anteriorly into the cortex of the walls of the rhinal sulcus. A comparisonof figs. 4, 10 and 13 shows that the degeneration in this region resulting from lesionsin areas 7, 21 and 22 forms a continuous, and slightly overlapping, band which liesimmediately lateral to the entorhinal area. This narrow band of degenerationcorresponds very closely to the perirhinal area, 35 of Brodmann (1909) and to TH ofBonin and Bailey (1947). Although Brodmann (1909) did not indicate this archi-tectonic area in his map of Cercopithecus he shows it on a section across the rhinalsulcus of the rhesus monkey and on the maps of all the other mammalian brainswhich he studied, including that of man. In all instances this area immediately adjoinsthe entorhinal cortex. The distribution of the degeneration in the vicinity of therhinal sulcus after damage of the inferior temporal gyrus in the monkeys studied byWhitlock and Nauta (1956) was very similar, as was that after lesions of the insulo-temporal region in the cats described by Cragg (1965) and Diamond et al. (1968).In all these earlier accounts the degeneration was considered to be in the lateral partof the entorhinal cortex, but it is probably more correct to interpret it as occupyinga separate architectonic area.
In the rat brain a projection has been found from the olfactory pathway into whathas been described as the most lateral part of the entorhinal cortex (Cragg, 1961;Powell et ah, 1965; White, 1965; Heimer, 1968) but which is now suggested to be atransition zone between the entorhinal cortex proper and the pyriform cortex (Priceand Powell, 1970). Furthermore, in the rat a projection has been found from theneocortex into area 35 which lies between the entorhinal area and the neocortex(Price and Powell, 1970). It would appear, therefore, that the entorhinal cortex maybe surrounded entirely by a transition zone, part of which adjoins the pyriform cortexand receives fibres from the olfactory pathway and the .rest of which is area 35adjoining the neocortex and receiving afferent connexions from the three mainsensory systems. In this transition region there would be a final relay of these sensorypathways before the entorhinal area proper, and thence the hippocampus, isreached.
These projections are of interest in view of Geschwind's (1965) belief that a lesionof an appropriate association area "may specifically disconnect the hippocampalregion from a single modality and produce a recent memory defect in the modalityalone." This would presuppose a non-convergent neocortical input to the hippo-campus such as that demonstrated here. A further point of interest is that at least inthe somatic sensory system, it is only from these regions in which the topographicsegregation commences breaking down that the input to the cingulate and para-hippocampal regions begins. Beyond this point, when topography is not maintainedand all systems start to converge, so most of the efferent intracortical pathways passto limbic cortex only, particularly to the cingulate and retrosplenial areas, and in thecase of the regions on the orbital surface of the frontal lobe, even to the hypothalamus
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

816 E. G. JONES AND T. P. S. POWELL
direct (Nauta, 1964; Johnson, Rosvold and Mishkin, 1968). The hypothalamus,therefore, seems to receive both "modality specific" and convergent information, thefirst through the entorhinal cortex and the hippocampus, the second both throughthis route (cingulate and retrosplenial cortex and hippocampus) and via the orbitalsurface of the frontal lobe.
Among the sensory systems, the olfactory and visual are different in having earlierinputs into the hypothalamus through the amygdala, the former as a direct pathwayfrom the olfactory tract (Powell et al., 1965) and as an indirect one from the pyriformcortex; the latter through the visual association area, area 20. A projection fromthe infero-temporal region to the basolateral group of amygdaloid nuclei has beennoted previously by Adey and Meyer (1952) and by Whitlock and Nauta (1956).Both of these projections are possibly explicable since olfactory and visual stimuliplay an important part in reproductive and other behaviour. In the case of repro-ductive behaviour, it would appear that the effective pathway for visual influencesfrom the retina reaches the hypothalamus at a pre-geniculate level (Clark,McKeown and Zuckerman, 1939). However, the visual system almost certainly hasother influences upon behaviour and Geschwind (1965) attributes the tameness tovisual stimuli of monkeys with temporal lobe lesions of the kind which induce aKlliver-Bucy syndrome (Akert, Gruesen, Woolsey and Meyer, 1961) to disconnexionof complex visual stimuli from the amygdala. In this context it may also be notedthat Walker (1957) states that the severest memory loss following unilateral temporallobe excision is experienced by those patients who use predominantly visualimagery.
The hippocampal formation has been implicated as being vital for recent memoryprocesses in man (Milner, 1958; see Brain, 1969, for other references), althoughWeiskrantz (1968) infers that the temporal cortex itself is more critical in the monkey.To some extent it might be possible to reconcile these divergent views, for inter-ference with the region of convergence in the depths of the superior temporal sulcusmight form the basis of Weiskrantz's experimental observations and if the sequenceof connexions observed in the present study is maintained, this region may beexpected to project directly or indirectly into the hippocampus. If it is connected tothe hippocampal formation, this could also explain Penfield's observations thatmemories may be evoked by stimulation of the temporal cortex (Penfield, 1958).There is, thus, a strong need for determining the further connexions of the inaccessibleGortex in the depths of the superior temporal sulcus. Other aspects of the corticalintegrative pathways which have not been dealt with in the present study but whichalso warrant further investigation are the interhemispheric connexions of the associa-tion areas for these, too, may be expected to play a part in the convergence of sensorypathways at certain levels. They may be especially important as an anatomical basisfor the unilateral dominance displayed by the human brain. Similarly, it is certainthat thalamocortical and corticothalamic pathways are involved in determining thefunctional characteristics of all areas of the cortex and there is the possibility of"further sensory convergence occurring by means of these routes.
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

CORTICAL ASSOCIATION CONNEXIONS 817
I
I
I
SUMMARY
The ipsilateral cortical connexions of the main sensory areas of the cerebral cortexhave been studied in the rhesus monkey, with a view to determining the regions inwhich the somatic, visual and auditory pathways come together.
Each primary sensory area projects to an adjacent field in the parieto-temporalcortex and to a different part of the premotor cortex. Only the somatic sensory areas,which are also reciprocally connected with the motor cortex, receive an afferentcortical input. There is next an orderly sequence of projections within each of thethree intracortical sensory paths in both the frontal and parieto-temporal lobes;each new frontal and parieto-temporal step in the sequence is reciprocally connected.Convergence of topographic subdivisions within a system appears to precede con-vergence between the three systems; when the topographic segregation within asystem can be no longer recognized, the three systems start to converge within boththe parieto-temporal and frontal regions. The most obvious regions of convergenceare in the depths of the superior temporal sulcus (probably the homologue of areas39 and 40 in man), at the frontal pole, and in orbito-frontal cortex of the frontaloperculum. As far as can be ascertained, all regions of convergence are inter-connected with one another, with the orbital surface of the frontal lobe and with thetemporal pole.
Projections into the limbic system both precede and follow the intracortical con-vergence of the three systems. Once the topographic subdivision within a systemcommences breaking down and before it meets the other two, there is a projectionboth to cingulate cortex and to a part of the parahippocampal gyrus, area 35. Thoseregions of convergence which have been studied also project to limbic cortex and thoseon the orbital surface may send fibres direct to the hypothalamus. The second step inthe sequence of visual projections (area 20) is unique in having a heavy, direct inputto the amygdala.
ACKNOWLEDGMENTSThis work was supported by grants from the Medical and Science Research Councils and was
done during the tenure by E. G. J. of a Nuffield Dominions Demonstratorship, on leave fromthe University of Otago, New Zealand.
ABBREVIATIONSA pi imary auditory cortexAm amygdalaAS arcuate sujcusCS central sulcusCal S calcarine sulcusCg S cingulate sulcusCG cingulate cortex1 PS intraparietal sulcusLS lunate sulcusLt S lateral sulcusOF orbito-frontal cortexOTS occipito-temporal sulcus
PS principal sulcusPSt prestriate cortexPrCo precentral agranular fieldRS rostral sulcusRh S rhinal sulcusS primary somatic cortexSI first somatic sensory areaSII second somatic sensory areaSM supplementary motor cortexSTP supratemporal planeSTS superior temporal sulcusTG area TG of Bonin and Bailey (1947)
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

818 E. G. JONES AND T. P. S. POWELL
REFERENCES
ADES, H. W., and FELDER, R. (1942) The acoustic area of the monkey (Macaca mulatto). J. Neurophysiol., 5,49-54.
ADEY, W. R., and MEYER, M. (1952) Hippocampal and hypothalamic connexions of the temporal lobe inthe monkey. Brain, 75, 358-384.
ADRIAN, E. D. (1949) "The Sherrington Lectures. I. Sensory Integration." Liverpool: Univ. Liverpool Press.AKERT K. (1964) Comparative anatomy of frontal cortex and thalamo-frontal connections. In: "The
Frontal Granular Cortex and Behavior." Edited by J. M. Warren and K. Akert. New York:McGraw-Hill, pp. 372-396.
, GRUESEN, R. A., WOOLSEY, C. N., and MEYER, D. R. (1961) Kluver-Bucy syndrome in monkeyswith neocortical ablations of temporal lobe. Brain, 84, 480-498.
BIGNALL, K. E. (1969) Bilateral temporofrontal projections in the squirrel monkey: origin, distribution andpathways. Brain Res., 13, 319-327.
, and IMBERT, M. (1969) Polysensory and cortico-cortical projections to frontal lobe of squirrel andrhesus monkeys. Electroenceph. din. Neurophysiol., 26, 206-215.
BONTN, G. VON, and BAILEY, P. (1947) "The Neocortex of Macaca Mulatta." Urbana: Univ. Illinois Press.BRAIN, W. R. (1961) "Speech Disorders. Aphasia, Apraxia and Agnosia." London: Butterworth.
(1969) Disorders of memory. In: "Recent Advances in Neurology and Neuropsychiatry." Editedby Lord Brain and M. Wilkinson. London: Churchill, pp. 1-12.
BRODMANN, K. (1909) "Vergleichende Lokalisationslehre der Grosshimrinde in ihren Pnnzipien dargestelltauf Grund des Zellenbaues." Leipzig: Barth.
CHOW, K. L. (1961) Anatomical and electrophysiological analysis of temporal neocortex in relation tovisual discrimination learning in monkeys. In: "Brain Mechanisms and Learning." Edited byJ. F. Delafresnaye. Oxford: Blackwell, pp. 507-525.
CLARK, W. E. LE GROS, MCKEOWN, T., and ZUCKERMAN, S. (1939) Visual pathways concerned in gonadalstimulation in ferrets. Proc. R. Soc., B, 126, 449-468.
CRAGG, B. G. (1961) Olfactory and other afferent connections of the hippocampus in the rabbit, rat, and cat.Expl Neurol., 3, 588-600.(1965) Afferent connexions of the allocortex. / . Anal., Lond., 99, 339-357.(1969) The topography of the afferent projections in the circumstriate visual cortex of the monkeystudied by the Nauta method. Vision Res., 9, 733-746.
CRITCHLEY, M. (1953) "The Parietal Lobes." London: Arnold.DEWSON, J. H. Ill, and COWEY, A. (1969) Discrimination of auditory sequences by monkeys. Nature, Lond.,
222, 695-697.DIAMOND, I. T., JONES, E. G., and POWELL, T. P. S. (1968) The association connections of the auditory
cortex of the cat. Brain Res., 11, 560-579.ETTLrNGRR, G., WARRTNGTON, E., and ZANGWILL, 0. L. (1957) A further study of visual-spatial agnosia.
Brain, 80, 335-361.FESSARD, A. E. (1954) Mechanisms of nervous integration and conscious experience. In: "Brain Mechanisms
and Consciousness." Edited by J. F. Delafresnaye. Oxford: Blackwell, pp. 200-236.FINK, R. P., and HEIMER, L. (1967) Two methods for selective silver impregnation of degenerating axons and
their synaptic endings in the central nervous system. Brain Res., 4, 369-374.GESCHWTND, N. (1965) Disconnexion syndromes in animals and man. Brain, 88, 237-294 and 585-644.GROSS, C. G., SCHILLER, P. H., WELLS, C , and GERSTHN, G. L. (1967) Single-unit activity in temporal associa-
tion cortex of the monkey. / . Neurophysiol., 30, 833-843., and WEBKRANTZ, L. (1964) Some changes in behavior produced by lateral frontal lesions in the
macaque. In: "The Frontal Granular Cortex and Behavior." Edited by J. M. Warren and K. Akert.New York: McGraw-Hill, pp. 74-101.
HECAEN, H., PENFIELD, W., BERTRAND, C , and MALMO, R. (1956) The syndrome of apractognosia due tolesions of the minor cerebral hemisphere. A.M.A. Archs Neurol. Psychiatry, 75, 400-434.
HEIMER, L. (1968) Synaptic distribution of centripetal and centrifugal nerve fibres in the olfactory system of therat. An experimental anatomical study. J. Anal., Lond., 103, 413-432.
I
I
I
I
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

I
I
I
I
CORTICAL ASSOCIATION CONNEXIONS 819
HUBEL, D. H., and WIESEL, T. N. (1965) Receptive fields and functional architecture in two nonstriate visualareas (18 and 19) of the cat. J. Neurophysiol, 28, 229-289.
HURST, E. M. (1959) Some cortical association systems related to auditory functions. / . comp. Neurol., 112,103-119.
JANSEN, J. (1937) Experimental investigations of the associational connections of the cerebral cortex, withspecial reference to the conditions in the frontal lobes. Avhandl. Norsk. Videnskaps-Akad. 1. Mat.Naturv., 1-21.
JOHNSON, T. N., ROSVOLD, H. E., and MISHKIN, M. (1968) Projections from behaviorally-defined sectors ofthe prefrontal cortex to the basal ganglia, septum, and diencephalon of the monkey. Expl Neurol.,21, 20-34.
JONES, E. G. (1969) Interrelationships of parieto-temporal and frontal cortex in the rhesus monkey. BrainRes., 13, 412^115.
, and POWELL, T. P. S. (1968) The ipsilateral cortical connexions of the somatic sensory areas inthe cat. Brain Res., 9, 71-94.
, (1969) Connexions of the somatic sensory cortex of the rhesus monkey. I. Ipsilateralcortical connexions. Brain, 92, 477-502.
KEMP, J. M., and POWELL, T. P. S. (1970) The cortico-striate projection in the monkey. Brain, 93, 525-546.KRIEO, W. J. S. (1963) "Connections of the Cerebral Cortex." Evanston: Brain books.KUYPERS, H. G. J. M., SZWARCBART, M. K., MISHKIN, M., and ROSVOLD, H. E. (1965) Occipitotemporal
corticocortical connections in the rhesus monkey. Expl Neurol., 11, 245-262.MASSOPUST, L. C , Jr., BARNES, H. W., and VERDURA, J. (1965) Auditory frequency discrimination in
cortically ablated monkeys. / . audit. Res., 5, 85-93.METTLER, F. A. (1935) Corticifugal fiber connections of the cortex of Macaca mulatta. The temporal region.
J. comp. Neurol., 63, 25-47.MTLNER, B. (1958) Psychological defects produced by temporal lobe excision. Res. Publs Ass. Res. nerv.
ment. Dis., 36, 244-257.MISHKTN, M. (1964) Perseveration of central sets after frontal lesions in monkeys. In: "The Frontal Granular
Cortex and Behavior." Edited by J. M. Warren and K. Akert. New York: McGraw-Hill, pp. 219-241.(1966) Visual mechanisms beyond the striate cortex. In: "Frontiers in Physiological Psychology."Edited by R. W. Russell. New York Academic Press, pp. 93-119.
MOFFETT, A., ETTLTNGER, G., Morton, H. B., and PERCY, M. F. (1967) Tactile discrimination performancein the monkey: the effect of ablation of various subdivisions of posterior parietal cortex. Cortex, 3,59-96.
NAUTA, W. J. H. (1964) Some efferent connections of the prefrontal cortex in the monkey. In • "The FrontalGranular Cortex and Behavior." Edited by J. M. Warren and K. Akert. New York. McGraw-Hill,pp. 397-407.
, and GYGAX, P. A. (1954) Silver impregnation of degenerating axons in the central nervous system:a modified technic. 5mm Technoi, 29, 91-93.
NEWCOMBE, F., and RUSSELL, W. RITCHIE (1969) Dissociated visual perceptual and spatial deficits in focallesions of (he right hemisphere. J. Neurol. Neurosurg. Psychiat., 32, 73-81.
PANDYA, D. K , HALLETT, M., and MUKHERJEE, S. K. (1969) Intra- and inter-hemispheric connections of theneocortical auditory system in the rhesus monkey. Brain Res., 14,49-65.
, and KUYPERS, H. G. J. M. (1969) Cortico-cortical connections in the rhesus monkey. Brain Res.,13, 13-36.
PENFELD, W. (1958) The role of the temporal cortex in recall of past experience and interpretation of thepresent. In • "Ciba Foundation Symposium on the Neurological Basis of Behaviour." Edited byG. E. W. Wolstenholme and C. M. O'Connor. London: Churchill, pp. 149-174.(1959} The interpretive cortex. Science, N. Y., 129, 1719-1725.(1966) Speech, perception and the uncommitted cortex. In: "Brain and Conscious Experience."Edited by J. C. Eccles. Berlin: Springer, pp. 217-237.
POWELL, T. P. S., COWAN, W. M , and RAISMAN, G. (1965) The central olfactory connexions. / . Anat., Lond.,99, 791-813.
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

820 E. G. JONES AND T. P. S. POWELL
PRIBRAM, K. H., BURTON, S. R., and ROSENBUTH, W. A. (1954) Electrical responses to acoustic clicks inmonkey: extent of cortex activated. / . Neurophysiol., 17, 336-344.
PRICE, J. L., and POWELL, T. P. S. (1970) Further observations on the central olfactory connexions. Inpreparation.
ROBERTS, T. S.( and AKERT, K. (1963) Insular and opercular cortex and its thalamic projection inMacaca mulatta. Schweizer Arch. Neurol. Neurochir. Psychiat., 92, 1-43.
ROSVOLD, H. E., and MISHKIN, M. (1961) Non-sensory effects of frontal lesions on discrimination learningand performance. In: "Brain Mechanisms and Learning." Edited by J. F. Delafresnaye. Oxford:Blackwell, pp. 555-576.
ROSVOLD, H. E., and SZWARCBART, M. K. (1964) Neural structures involved in delayed-responseperformance, pp. 1-15. In: "The Frontal Granular Cortex and Behaviour " Edited by J. MWarren and K. Akert. New York: McGraw-Hill.
TEUBER, H. L. (1965) Postscript: some needed revisions of the classical views of agnosia. Neuropsychologia,3, 371-378.
• (1966) Alterations of perception after brain injury. In • "Brain and Conscious Experience." Editedby J. C. Eccles. Berlin: Springer, pp. 182-216.
WALKER, A. E. (1940) A cytoarchitectural study of the prefrontal area of the macaque monkey. J. comp.Neurol., 73, 59-86.(1957) Recent memory impairment in unilateral temporal lesions. A.M.A. Archs Neurol. Psychiatry,78, 543-552.
WEISKRANTZ, L. (1968) Experiments on the r.n.s. (real nervous system) and monkey memory. Proc. R. Soc ,B, 171, 335-352.
WHITE, L. E., Jr. (1965) Olfactory bulb projections of the rat. Anat. Rec, 152, 465-480.WHTTLOCK, D. G., and NAUTA, W. J. H. (1956) Subcortical projections from the temporal neocortex in
Macaca mulatta. / . comp. Neurol., 106, 183-212.WOOLSEY, C. N. (1958) Organization of somatic sensory and motor areas of the cerebral cortex. In-
"Biological and Biochemical Bases of Behavior." Edited by H. F. Harlow and C. N. Woolsey.Madison: Univ. Wisconsin Press, pp. 63-81.
ZEKJ, S M. (1969) Representation of central visual fields in prestriate cortex of monkey. Brain Res., 14,271-291.
LEGEND FOR PLATE
PLATE LXFIG. 18.—Both photomicrographs are of frozen sections stained by the Nauta-Gygax (1954)
technique, A, to show degeneration in the cortex of area 46 of the frontal lobe twelve days aftera lesion was placed in area 21 of the temporal lobe, x 680. B, The fibre and terminal degenerationin the cortex of the superior temporal sulcus eleven days after extensive damage of the frontal lobe.x97O.
(Received 25 February 1970)
I
I
I
I
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from

PLATE LX
I
I
i FIG 18.
7c> illustrate article by E. G. Jones and T. P. S. Powell.
at Aston U
niversity on April 25, 2014
http://brain.oxfordjournals.org/D
ownloaded from