alterations in connexin expression and cell...
TRANSCRIPT

Alterations in Connexin Expression and CellCommunication in Healing Corneal Epithelium
Maja Matic* Igor N. Petrov* Tovia Rosenfeld* andj. Mario Wolosin *f
Purpose. The aim of this study was to examine cell-to-cell metabolite transfer and connexindistribution in the rabbit corneal epithelium, in the stationary state, and during woundhealing.
Methods. Rabbit corneas were wounded with a surgical tool, producing a 3-mm-wide elongateddebridement. Corneas were allowed to heal in vivo for up to 45 hours. Monoclonal antibodiesagainst connexins Cx 26, Cx 32, Cx 43, and Cx 50 were used to stain cryostat sections.Cell-to-cell metabolite transfer capacity was assessed by a modification of the scrape-loadingtechnique using lucifer yellow as the organic ion tracer.
Results. The rabbit corneal epithelium contains Cx 43 and Cx 50, localized in the cell's plasmamembrane, as shown previously for other species. Cx 26 and Cx 32 are not detectable. Tracertransfer occurred in both basal and suprabasal cell layers. After wounding, the migratingepithelial monolayer lacked Cx 43 and Cx 50. This change was apparent 6 hours after injuryand persisted until complete wound closure (approximately 24 hours). The Cx 50 membranestain was increased elsewhere, in particular in the transition zone between monolayered andmultilayered epithelium. Consistent with the expression changes, migrating cells displayedno or minimal cell-to-cell tracer transfer, whereas in the periphery of the wound, tracertransfer was enhanced in comparison to the control specimen.
Conclusions. Corneal epithelial healing involves biphasic changes in the expression of connex-ins and cell-to-cell communications. These alterations may be critical for the optimization ofthe healing response. Invest Ophthalmol Vis Sci. 1997;38:600-609.
Af ter corneal injury, rapid healing of the epitheliumis essential to reestablish the ocular surface permeabil-ity barrier function. Reepithelialization is accom-plished by the migration of adjacent cells over theinjured area.12 The migrating cells undergo substan-tial phenotypic changes, particularly regarding cy-toskeletal organization3'4 and adherence to the sub-stratum and to each other. Nevertheless, sufficientcell-to-cell contacts are retained to cause the cells tomigrate as a cohesive sheet.5'6 One of the phenotypicalattributes of the stationary corneal epithelium is itsgap junction-mediated7"9 syncytial arrangement, at
From the Departments of* Ophthalmology and fPhysiology and Biophysics, MountSinai School of Medicine, New York, New York.Supported by National Institutes of Health grants EY07773 and EY01867 (corecenter grant) and by an unrestricted grant from Research to Prevent Blindness.JMW is an RPB Senior Scientific Investigator.Submitted for publication May 13, 1996; revised October 10, 1996; acceptedOctober 11, 1996.Proprietary interest category: N.Reprint requests: f. Mario Wolosin, Department of Ophthalmology, Mount SinaiSchool of Medicine, One Gustave L. Levy Place, New York, NY 10029.
least regarding electrical communications.1011 Syncy-tial arrangements may play a role in the coordinationof physiological processes involved in tissue develop-ment and regeneration.1213 Thus, the fate of thesejunctions and the cell-to-cell communication levelsduring the migratory phase of wound healing is a sub-ject that deserves investigation.
Gap junctions are made of connexins, an ex-tended family of related polypeptides.14 So far, 12 dif-ferent connexins have been described. Gap junctionsmade from different connexins may have uniqueproperties with respect to permeability, voltage, andpH dependence. Two such connexins, Cx 43, a 43-kDa protein identified initially in mammalian cardiacmyocytes,8 and Cx 50, identified initially as a lens fibergap junction polypeptide in rodents,15 have been de-scribed previously in the rat and ovine corneal epithe-lium.8'9 However, the functional consequences of thepresence of these connexins (i.e., cell-to-cell metabo-lite transfer) have not been examined yet. Accord-ingly, the current study was undertaken to establish
600Investigative Ophthalmology & Visual Science, March 1997, Vol. 38, No. 3Copyright © Association for Research in Vision and Ophthalmology
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933197/ on 06/06/2018
Gap Junctions and Epithelial Wound Healing
FIGURE 1. Schematic representation of the procedures per-formed to obtain wound-healing corneal samples. At t = 0,a 3-mm wide, 6- to 8-mm-long debridement was performedin an anesthetized rabbit as described in the Methods sec-tion. The area of this wound is represented by the horizon-tally oriented rectangular area a. Afterward, epithelial migra-tion covered some of the debrided area, leaving a smallerepithelial-free defect. This is represented by the horizontallyoriented area b. At this point, after animal euthanasia, thetracer transfer experiment {see the Methods section) wasperformed. The dye-loaded linear incision is represented bythe thick lane c. Finally, corneal segments were excised foranalysis. For the tracer transfer experiments, the segmentsextended from the center of the wound outward d. Forconnexin immunofluorescence studies (no tracer-loadingperformed), the segment extended equally to both sides ofthe wound (not shown).
both the connexin distribution and the tracer transferproperties of the rabbit corneal epithelium in its sta-tionary state and in the course of wound healing.
METHODS
Corneal Debridement
Adult albino rabbits weighing 1 to 2 kg were anesthe-tized by intravenous injection of a 1:2 mixture of keta-mine (100 mg/ml) and acepromazine (10 mg/ml).One drop of 0.5% proparacaine was applied topicallyon the eyes. Full-thickness epithelial lesions were pro-duced with a sterilized, blunt tool that was machinedand polished to minimize damage to the underlyingbasement membrane. Wounds were allowed to healin vivo for up to 45 hours. A schematic description ofthis and subsequent procedures is provided in Figure1. Unless specified otherwise, animals were killed bycarbon dioxide narcosis and asphyxiation. All experi-ments were performed in accordance with the ARVOStatement for the use of Animals in Ophthalmic andVision Research.
Immunofluorescent Staining
After enucleation, corneas were excised and cut intoappropriate segments for cryostatic processing. Thesegments were embedded immediately in Cryomatrix(Shandon, Pittsburgh, PA) and quick frozen by im-mersion in liquid nitrogen. Sixteen-micrometer-diicksections were cut orthogonally to the main cornealplane in a cryotome (International Equipment, Need-ham Heights, MA) at -20°C and deposited on sylane-coated microscope slides (New Jersey Lab and GloveSupply, Livingston, NJ). After 10 minutes, tissue fixa-tion and permeabilization in HistoChoice (Amresco,Solon, OH; complemented with 0.1% Triton X-100)for 10 minutes, and three 5-minute washes, the slideswere incubated in phosphate-buffered saline con-taining 5% bovine serum albumin (Type V; Sigma,St. Louis, MO) for 45 minutes to block nonspecificbinding sites. The sections then were overlaid witheither mouse monoclonal immunoglobulin G anti-bodies raised against Cx 26, Cx 32 (Zymed; San Fran-cisco, CA) or Cx 43 (Chemicon; Temecula, CA), ora mouse monoclonal immunoglobulin M antibody,which recognizes Cx 50 (a generous gift of Dr.Goodenough, Harvard Medical School, Boston, MA).After one 45-minute incubation and three 15-minutewashes, the corresponding tetramethylrhodamine- orfluorescein isothiocyanate-conjugated goat antimouse(Chemicon) secondary antibody was applied for 30minutes. For negative control specimens, the primaryantibody was omitted. All incubations were done atroom temperature. After three final 15-minute washes,sections were mounted with Vectashield (Vector Labo-ratories, Burlingame, CA) and photographed in anOlympus IMT-2 microscope (Tokyo, Japan) equippedwith fluorescence epiillumination. Between 90 and100 sections were cut per frozen sample. Three to foursuch sections were deposited per slide and two slideswere processed and examined per sample. Thus, de-pending on the number of control specimens carriedout, four to eight experimental sections were exam-ined per embedded sample. In some experiments,after acquisition of the immunofluorescent images,the coverslip was removed, the section was stained withhematoxylin-eosin, remounted, and photographedunder transmitted light.
Tracer Transfer Experiments
Adult albino rabbits were subjected to corneal de-bridement as described above. Eighteen hours afterthe treatment, the animals were anesthetized andkilled by an air embolism. (This method was adoptedbecause the use of carbon dioxide causes marked in-tracellular acidification, which, in turn, may blockjunctional communications.) Immediately afterward,before the removal of the cornea from the animal,
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933197/ on 06/06/2018

602 Investigative Ophthalmology & Visual Science, March 1997, Vol. 38, No. 3
FIGURE 2. Immunofluorescence micrograph of Cx 43 andCx 50 in the anterior central cornea. (A) Cx 43; note thelocalization of the staining in the basal cells with occasionalexpansion to a few stratifying cells as well as the fine, alignedpunctuate staining in the stroma (small vertical arrows). (B)Cx 50; note that the staining is present throughout the epi-thelial strata with maximal intensity in the wing cells. Bar =100 fj,m.
the scrape-loading procedure described originally byEl-Fouly et al,16 adapted to the corneal morphology,was performed. The eye surface was irrigated with a10-mM ethylene-bis (oxyethylenenitrilo) tetraaceticacid-phosphate-buffered saline solution containing3% lucifer yellow (LY; Sigma). A small surgical bladewas used to produce a nonpenetrating linear incision(referred henceforth as the cut-loading incision, orthe cut edge, as appropriate) perpendicular to, andacross the original wound (Fig. 1). In some experi-ments, two consecutive, parallel cuts were made. Dur-ing the cutting and for the subsequent 60 seconds,the tracer's solution was dripped along the incision.The ocular surface then was irrigated profusely for 10minutes with buffered formaldehyde solution. The eyewas enucleated, and the cornea was excised and post-fixed in formaldehyde solution for 2 to 4 hours. Ap-propriate 3- to 4-rnm-long tissue segments then wereprocessed for either sectioning or confocal micros-copy. Serial cryostatic sectioning was performed or-thogonally to the dye-loading cut starting from thecenter of the wound zone. The extent of spreading ofLY then was determined by epifluorescent microscopywith the proper filter set. The approximate distanceof a given section from die leading edge of die healingepithelium (i.e., the first section showing epithelium-covered stroma) was determined from the number ofsections cut (e.g., the thirtieth subsequent section waslocated at 16 mm X 30 = 0.48 mm away from theedge).
Confocal MicroscopyElongated corneal segments, extending from the orig-inal central wound toward the corneal periphery and
incorporating the cut-loading incision, were incu-bated in glycerol for 48 to 96 hours. A 400-g weightwas applied after the first 24 hours to eliminate tissuecurvature. The glycerol-embedded segment was flat-tened and placed epithelial side down on a coverslipset on a Leica LSCM inverted confocal microscope.The left or the right edge of the intensely fluorescentcut-loading incision was placed near the left or theright end of the observation field, respectively. Thesample then was scanned upward, in 2-/j,m increments,from the corneal surface to the stroma (which easilywas identifiable as the area yielding diffuse fluores-cence) using a 10X objective (numerical aperture =0.3). Finally, a plane of focus lying 10 /im away fromthe first plane of focus-insinuating stromal-derivedfluorescence was selected as the basal cell plane.
RESULTS
Immunofluorescent staining of adult rabbit cornealepithelium showed the presence of Cx 43 epitopes inthe basal cell layer (Fig. 2A; n = 6). Well-defined,punctuate staining could be observed along the apicaland lateral membranes. The suprabasal cells were de-void of such an epitopej except for cells that wereclearly in the early stratification stage. This distribu-
FIGURE3. Representative images of lucifer yellow dye spread-ing in the unwounded anterior central cornea. (A) Lowmagnification view of the cut zone showing dye spreadinginto both ends of the cut epithelium and into the subadja-cent stroma. (B,C) Representative profiles derived from twodistinct corneas. Dye spreading is evident into basal andsuprabasal cells. In most samples, however, spreading wasslightly more efficient within basal cells (arrows pointing todye excluding suprabasal cells in B). Asterisks indicate thecut-loading incision. Bar = 400 p,m (A); = 100 /xm (B,C).
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933197/ on 06/06/2018
Gap Junctions and Epithelial Wound Healing 603
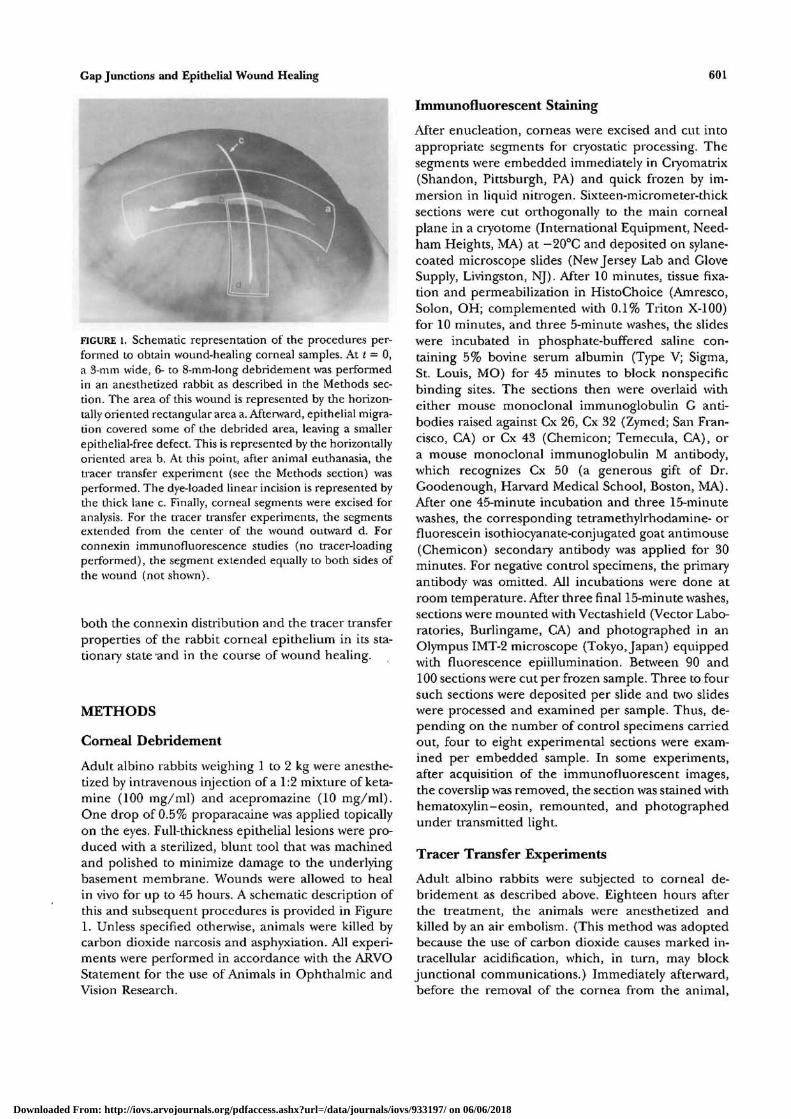
FIGURE 4. Micrographic details of a cornea 6 hours after epithelial debridement. (A) Hema-toxylin-eosin staining; note the swelling of the superficial stroma in the debridement area.(B,C) Immunostaining for Cx 43 of the left and right wound edge, respectively. The tip ofthe leading cells in each side is shown {arroioheads). Some of the mobilized cells (mroxos)exhibit diffuse fluorescence above the background level. Bar = 600 //m (A); = 100 /im
tion was invariant throughout the cornea, from theperiphery to the center. Cx 43 also was abundantlypresent in the stroma, where the parallel, lamellar-like pattern of staining (Fig. 2A, vertical arrows) wasconsistent with the expected distribution of keratocyte
17,18
basal layer. There was minimal staining of the squa-mous-like outer layers and no stromal staining (Fig.2B, n = 6). Antibodies against two other well-charac-terized connexins, Cx 26 and Cx 32, fail to stain anycorneal component in four different mammalian spe-
junctions. Cx 50 was present primarily in the su- cies.prabasal wing cells, with weaker staining seen in the To establish whether the presence of Cx 43 or Cx
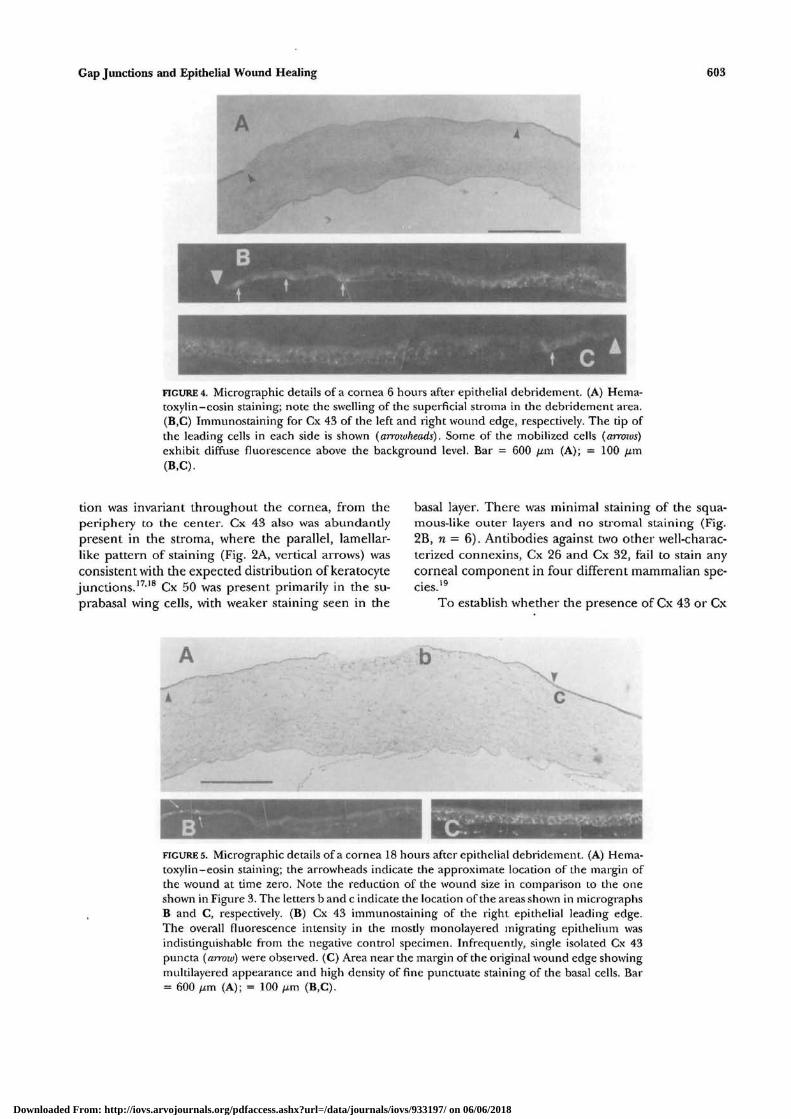
FIGURES. Micrographic details of a cornea 18 hours after epithelial debridement. (A) Hema-toxylin-eosin staining; the arrowheads indicate the approximate location of the margin ofthe wound at time zero. Note the reduction of the wound size in comparison to the oneshown in Figure 3. The letters b and c indicate the location of the areas shown in micrographsB and C, respectively. (B) Cx 43 immunostaining of the right epithelial leading edge.The overall fluorescence intensity in the mostly monolayered migrating epithelium wasindistinguishable from the negative control specimen. Infrequently, single isolated Cx 43puncta (arrozv) were observed. (C) Area near the margin of the original wound edge showingmultilayered appearance and high density of fine punctuate staining of the basal cells. Bar= 600 fim (A); = 100 ^m (B,C).
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933197/ on 06/06/2018

604 Investigative Ophthalmology & Visual Science, March 1997, Vol. 38, No. 3
50 or both confers to the system junctional permeabil-ity toward typical cell metabolites, the cell-to-cell trans-fer capacity was assayed by the cut-loading techniquedescribed in the Methods section. Figure 3 depicts thetypical spreading of LY during a 60-second incubation.Along the basal cell axes, LY diffused approximately10 cells inward from the cut edge {n = 6). Dye spread-ing also was patent in the suprabasal, intrastratal celllayers. Frequently, however, LY was excluded fromwing cells overlying dye-loaded basal cells Fig. 3B). LYalso was seen in the superficial cells. However, dyeaccumulation in these cells also was observed in testexperiments in which the corneal surface was pressed(rather than cut) using a blunted blade to generatelateral stretch tension (data not shown). Thus, thepossibility that, during cutting, these mature cells un-dergo mechanical damage leading to dye penetrationthrough disrupted apical surfaces can not be dis-counted. Additionally, some of the older superficialcells, in their final aging stages, may be permeablespontaneously.
The objective of this study was to determine theeffect of epithelial wounding on both connexin ex-pression and cell-to-cell solute transfer capacity. Thesize of the wound was chosen on consideration of anumber of factors, including the size of the rabbitcornea and the wound closure kinetics. The use of a3-mm wide, linear-like (>8-mm long) debridementwas found highly convenient from an experimentalviewpoint. Wounds were allowed to heal for differentperiods, up to 2 days. Animals then were killed andthe corneas processed for immunostaining, or scrape-loading.
Six hours after the debridement, the epitheliumwas in its early stage of mobilization (Fig. 4A). Immu-nostaining with Cx 43 showed that the cells at theleading edge lost their punctuate membranous stain-ing, (Figs. 4B, 4C), although diffuse staining in excessof background fluorescence could be seen in some ofthese cells (n = 1). After a healing period of 18 hours,the wound approached full closure (Fig. 5A). Themonolayer of basal cells that had migrated over thepreviously denuded area (1.0 to 1.5 mm in length)lacked (or has minimal levels of) Cx 43 (Fig. 5B) andCx 50 (Figs. 6A, 6B). In contrast, the two connexinsabundantly were present in die transition zone locatedbetween the advancing monolayer and the fully stra-tified area (Figs. 5C, 6B). In particular, Cx 50, whichbarely is expressed in the basal cells of the stationeryepithelium, displayed strong expression in basally lo-cated cells adjacent to the overtly monolayered zone.Furthermore, the Cx 50 staining in fully stratifiedareas 2 to 3 mm away from the wound center exhibitedlevels that clearly were higher than those observedanywhere in any of the six control specimens studied(Figs. 6C, 6.D). This enhanced staining was not uni-
FIGURE 6. Cx 50 epithelial distribution at various times aftercorneal debridement. (A to D) Details pertinent to samplesthat have undergone 18 hours of healing. (A,B) Nomarskicontrast (A) and hnmunofluorescent (B) micrographs ofthe multilayered-to-monolayered transition zone. Note in-tense Cx 50 staining on the multilayered side and sharpdisappearance of the staining as the epithelium becomesmonolayered. (C) Cx 50 distribution in a control specimen,unwounded cornea. (D) Cx 50 expression in a fully stratifiedarea of a wounded sample. Samples C and D were embeddedand frozen side by side in a single block to ensure completelyidentical processing from embedding to section mountingin Vectashield (Vector Laboratories, Burlingame, CA), Sub-sequently, all photographic procedures needed to generatemicrographs were performed under identical conditions.(E) The multilayered-to-monolayered transition zone of asample that has undergone 24 hours of healing and reachedfull wound seal. Note the intense, disorganized staining onthe multilayered side of the tissue and the rapid transitionto a stain-free condition as the epithelium becomes mono-layered. Asterisks indicate the side proximal to the wound.Bar = 100 /j,m (A,B,E); = 60 /xm (C,D).
form; rather, it occurred in discrete patches. Four sep-arate experiments yielded comparable results.
The metabolite transfer capacity in cells of theepithelium after 18 hours of healing was studied byscrape-loading. Samples were examined by either sa-gittal sequential sectioning (n — 4) or en face by con-focal microscopy (n = 2), as described in the Methodssection. Figure 7 depicts LY transference profiles near(Figs. 7B, 7D) and away (Figs. 7A, 7C) from the ad-vancing edge of the wound, respectively. In the migrat-ing epithelium, LY was present only in the cells adja-cent, to the cut edge. For the most part, the intensityof the fluorescence in these cells was uniform andhigh and ended abruptly rather than decreasing grad-ually away from the wound, suggesting that it reflectsonly entry into cells damaged by the incision. Never-
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933197/ on 06/06/2018
Gap Junctions and Epithelial Wound Healing 605
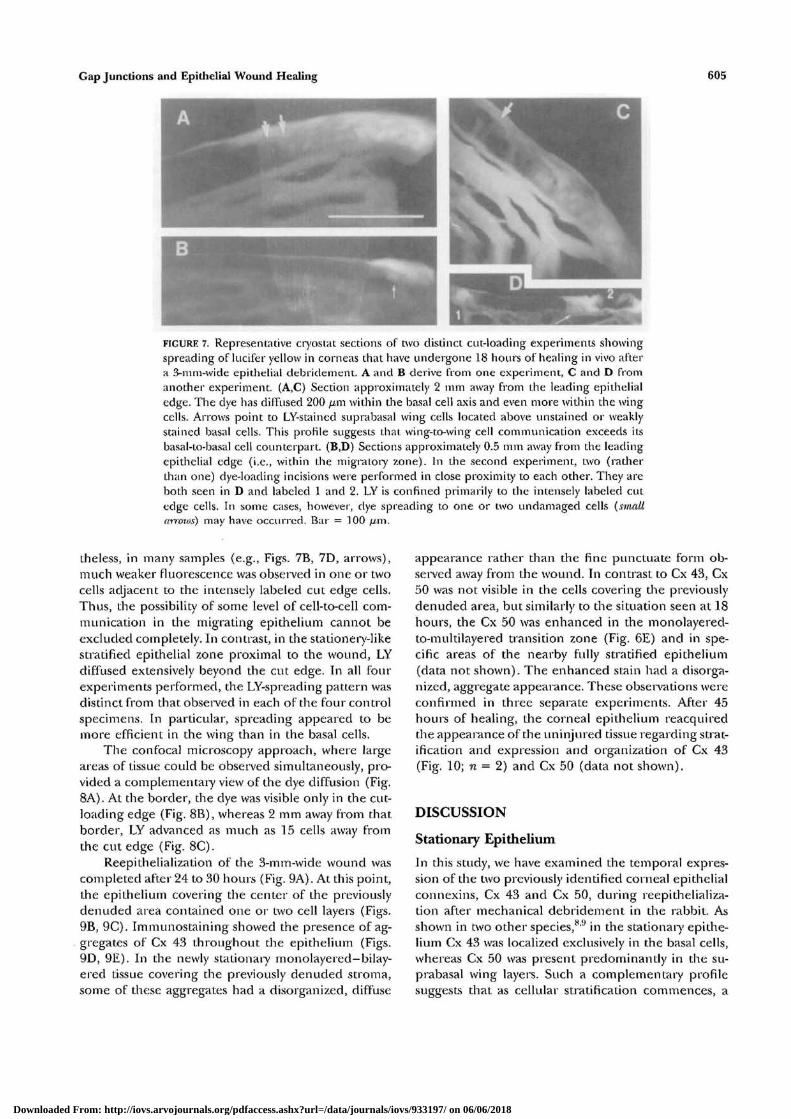
FIGURE 7. Representative cryostat sections of two distinct cut-loading experiments showingspreading of lucifer yellow in corneas that have undergone 18 hours of healing in vivo aftera 3-mm-wide epithelial debridement. A and B derive from one experiment, C and D fromanother experiment. (A,C) Section approximately 2 mm away from the leading epithelialedge. The dye has diffused 200 fim within the basal cell axis and even more within the wing-cells. Arrows point to LY-stained suprabasal wing cells located above unstained or weaklystained basal cells. This profile suggests that wing-to-wing cell communication exceeds itsbasal-to-basal cell counterpart. (B,D) Sections approximately 0.5 mm away from the leading-epithelial edge (i.e., within tbe migratory zone). In the second experiment, two (ratherthan one) dye-loading incisions were performed in close proximity to each other. They areboth seen in D and labeled 1 and 2. LY is confined primarily to the intensely labeled cutedge cells. In some cases, however, dye spreading to one or two undamaged cells (smaltarrows) may have occurred. Bar = 100 fj.m.
theless, in many samples (e.g., Figs. 7B, 7D, arrows),much weaker fluorescence was observed in one or twocells adjacent to the intensely labeled cut edge cells.Thus, the possibility of some level of cell-to-cell com-munication in the migrating epithelium cannot beexcluded completely. In contrast, in the stationery-likestratified epithelial zone proximal to the wound, LYdiffused extensively beyond the cut edge. In all fourexperiments performed, the LY-spreading pattern wasdistinct from that observed in each of the four controlspecimens. In particular, spreading appeared to bemore efficient in the wing than in the basal cells.
The confocal microscopy approach, where largeareas of tissue could be observed simultaneously, pro-vided a complementary view of the dye diffusion (Fig.8A). At the border, the dye was visible only in die cut-loading edge (Fig. 8B), whereas 2 mm away from thatborder, LY advanced as much as 15 cells away fromthe cut edge (Fig. 8C).
Reepithelialization of the 3-mm-wide wound wascompleted after 24 to 30 hours (Fig. 9A). At this point,the epithelium covering the center of the previouslydenuded area contained one or two cell layers (Figs.9B, 9C). Immunostaining showed the presence of ag-gregates of Cx 43 throughout the epithelium (Figs.9D, 9E). In the newly stationary monolayered-bilay-ered tissue covering the previously denuded stroma,some of these aggregates had a disorganized, diffuse
appearance rather than the fine punctuate form ob-served away from the wound. In contrast to Cx 43, Cx50 was not visible in the cells covering die previouslydenuded area, but similarly to the situation seen at 18hours, the Cx 50 was enhanced in the monolayered-to-multilayered transition zone (Fig. 6E) and in spe-cific areas of the nearby fully stratified epithelium(data not shown). The enhanced stain had a disorga-nized, aggregate appearance. These observations wereconfirmed in three separate experiments. After 45hours of healing, the corneal epithelium reacquiredthe appearance of the uninjured tissue regarding strat-ification and expression and organization of Cx 43(Fig. 10; n = 2) and Cx 50 (data not shown).
DISCUSSION
Stationary Epithelium
In this study, we have examined the temporal expres-sion of the two previously identified corneal epithelialconnexins, Cx 43 and Cx 50, during reepithelializa-tion after mechanical debridement in the rabbit. Asshown in two other species,8'9 in the stationary epithe-lium Cx 43 was localized exclusively in the basal cells,whereas Cx 50 was present predominantly in the su-prabasal wing layers. Such a complementary profilesuggests that as cellular stratification commences, a
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933197/ on 06/06/2018

606 Investigative Ophthalmology & Visual Science, March 1997, Vol. 38, No. 3
FIGURE 8. Diffusion of lucifer yellow (LY) in wounded tissue18 hours after epithelial debridement viewed by en faceconfocal microscopy. (A) Nomarski-contrast micrograph ofthe cut-loading incision. The large arrow points to the de-marcation generated by the wound margin (i.e., the leadingedge of the migrating epithelium); the remaining denudedstroma lies above this demarcation. The cut-loading incisionis seen as the horizontal fissure bisecting the segment fromend to end. The letters b and c indicate the areas imagedin the B and C micrographs, respectively. (B) LY fluores-cence at the wound margin (\vn). The intense fluorescenceat the top of the image reflects the penetration of LY intothe denuded stroma during the dye exposure period. Notethe complete absence of dye spreading away from the load-ing incision (li; seen as the vertically oriented fluorescence).(C) LY fluorescence at a location approximately 2 mm awayfrom the wound margin. Note that LY uniformly has spreadapproximately 200 fj,m away from the incision edge through-out the imaged area.
coordinated switch from Cx 43 to Cx 50 expressionoccurs. Additionally, Cx 50 was present minimally inthe flattened superficial cell layers, consistent with thepaucity of gap junction freeze fracture profiles in thesecells.7
The stratal distribution profiles may be importantfor the ionic and metabolic activities of the tissue.The diffusion of LY confirmed the functionality ofthe syncytium regarding metabolites; during the 60-second incubation, this 443-Da divalent anion spreadto a substantial number of cells away from the cut
edge. In microelectrophysiological studies, Klyce10 hasshown that the basal cells maintain limited ionic com-munications with the overlying wing cells (e.g., a basal-wing cell junctional resistance in the order of 300 to400 ft, [or 20% of the total transcellular epithelialresistance] could be measured). Using the paired oo-cyte expression assay,20 White et al21 have shown thatCx 50 and Cx 43 hemiconnexins do not form func-tional channels. Thus, unless connexins other thanCx 43 and Cx 50 are present in the tissue, the wing-to-basal cell-to-cell communication can be expectedto be dependent exclusively on the Cx 50 distribution;Cx 43 may be involved exclusively in communicationsbetween basal cells. The weak expression of Cx 50 inbasal cells then may underpin the partial basal-wingcommunication discontinuity shown by the microelec-trophysiological study. The inability of LY to diffuseinto some of the wing cells from their underlying basalcounterparts also is fully consistent with this notion.
Healing Epithelium
The immunostaining profiles 6, 18, and 24 hours aftera 3-mm-wide elongated debridement indicate clearlythat at certain times of the wound healing process, Cx43 and Cx 50 are downregulated in the basal cellsas they migrate over denuded basement membrane.Additionally, the Cx 50 staining patterns in Figure 6suggest either transient increases in the expression orchanges in the organization of this connexin in boththe monolayered-to-multilayered transition zone andthe fully stratified nearby area. With respect to Cx 43,we could not discern from the current histologic datawhether its level of expression in the transition zonewas modified from those of the unwounded controlspecimens. Although there is not previous pertinentdata for the cornea, Gabbiani et al22 found a fourfoldincrease in gap junctional area in the outer marginsof epidermal wounds. Additionally, Goliger and Paul23
made similar observations regarding enhanced ex-pression of Cx 26, the main suprabasal epidermal con-nexin,24 during wound healing of rat epidermis.
The LY spreading patterns in the samples thatunderwent 18 hours of healing were consistent fullywith the connexin staining patterns described above.At the wound and near the wound site, LY could notbe detected beyond the cut edge, although our studycould not discern between a total absence of commu-nication and a drastic reduction of it. In contrast, inthe proximity of the transition zone, LY spread pro-fusely away from the cut-loading edge. Furthermore,consistent with the apparent increases in Cx 50 expres-sion during this period, dye spreading in this areaappears to be both more extensive than that observedin the uninjured cornea and weighed toward the su-prabasal cell layers rather than to the basal cells. Inter-estingly, in contrast with the results in the epithelium,
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933197/ on 06/06/2018

Gap Junctions and Epithelial Wound Healing
A
607
FIGURE 9. Micrographic details of a cornea 24 hours after epithelial debridement. (A) Lowmagnification hematoxylin-eosin staining shows complete reepithelialization. The approxi-mate locations of Lhe margins of the original wound are shown {arrowheads). (B and C)Higher magnification details of A showing that the newly formed epithelium is monolayeredor bilayered. (D and E) Cx 43 immunostaining 2 mm away from (D), and within thereepithelialized area (E), Note that, whereas in the fully stratified area, the staining exclu-sively is punctuate, and in the newly formed tissue, the staining displays a diffuse or disorga-nized profile as well. The asterisks indicate the side proximal to the wound. Bar = 700 ,̂m(A); = 200 fj,m (B); - 50 fxm (C.D.E).
using simple-cell microinjections, Watsky18 has docu-mented the preservation of gap junctional communi-cations between corneal keratocytes migrating into awounded stromal area. However, the data provided inthat study suggest that a substantial reduction in theextent of communication occurs 24 hours post-wounding.
After reepithelialization of the denuded areas(i.e., on the reestablishment of the continuous mono-layer), Cx 43 staining appeared in the monolayeredcell zone. Given the reported fast turnover rate of Cx43,25l2h vis-a-vis the length of the healing process, andthe absence of Cx 43 staining in the migrating cellsbefore wound seal, it is logical to assume that thisvigorous staining represents de novo expression of Cx
FIGURE 10. Immunofluorescent photomicrograph of cornealepithelium 45 hours postdebridement showing complete re-stratification of the epithelium and the expression of Cx 43comparable to the unwounded tissue. Bar = 100 /urn.
43. Because this rapid reexpression occurs without anovert change in either the cell environment or shape,it suggests that the previous connexin downregulationmay result directly from the migratory state of thecells. For example, the cytoskeletal reorganization as-sociated with rapid migration3'"1^ may be incompati-ble with intracellular transport functions that are es-sential to sustain certain constitutive structures, suchas gap junctions, in the plasma membrane. Autocrini-cally derived cytokines27 or the absence of close range,stable interactions of the migrating cells with base-ment membrane elements affecting cell phenotype28
also could be involved in this downregulation. At vari-ance with the preceding analysis, Goliger and Paul"9
had suggested that the loss of intracellular communi-cations may be a causal factor for cellular activationrather than a consequence of it. Wound healing stud-ies with a recently developed Cx 43 knockout micemight provide the means to address this intriguingissue. The elimination (or drastic reduction) of cell-to-cell communications may be advantageous for cellsundergoing rapid migration because such migrationinvolves coordinated spatiotemporal fluctuation insecond messengers (e.g., Ca2+) within each singlecell30; cell-to-cell communications may be incompati-ble or detrimental to these intracellular rhythms.
The changes in the Cx 50 staining away from the
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933197/ on 06/06/2018

608 Investigative Ophthalmology 8c Visual Science, March 1997, Vol. 38, No. 3
wound margin area likely are to be mediated by diffus-ible signals able to reach beyond the original debride-ment zone. Because epithelial wound may cause therelease of a large variety of cytokines and growth fac-tors,31 further elaboration at this point is unwarranted.Nevertheless, it is noteworthy that enhanced cell com-munications will improve the metabolic status of thecells at the wound periphery. Takahashi et al32 recentlyhave shown rapid upregulation of the facilitated glu-cose transporter in the basal cells during wound heal-ing. The enhanced cell-to-cell communications mayextend the putative metabolic benefits of such upregu-lation to the suprabasal cell compartment during cer-tain periods of the wound healing response.
Key Words
connexin expression, corneal epithelium, gap junctions,metabolic cooperation, wound healing
Acknowledgments
The authors thank Dr. Scott Henderson for his help withconfocal microscopy, Dr. Zeev Stegman for image pro-cessing, and Christine Wang for her help with manuscriptpreparation.
References
1. Hanna C. Proliferation and migration of epithelialcells during corneal wound repair in the rabbit andthe rat. AmJOphthalmol. 1966;61:55-63.
2. Pfister RR. The healing of corneal epithelial abrasionsin the rabbit: A scanning electron microscope study.Invest Ophthalmol Vis Sci. 1975; 14:648-661.
3. Takeuchi S. The rearrangement of cytoskeletal sys-tems in epithelial cells accompanying the transitionfrom a stationary to a motile state at the start of epithe-lial spreading. / Cell Sci. 1987;88:109-119.
4. Gipson IK, Anderson RA. Actin filaments in normaland migrating corneal epithelial cells. Invest Ophthal-mol Vis Sci. 1977; 16:161-166.
5. Krawczyk WS, Wilgram GF. Hemidesmosome and des-mosome morphogenesis during epidermal woundhealing. / Ultrastruct Res. 1973;45:93-101.
6. Chan KY, Patton DL, Cosgrave YT. Time-lapse video-microscopic study of in vitro wound closure in rabbitcorneal cells. Invest Ophthalmol Vis Sci. 1989;30:12-19.
7. McLaughlin B, Caldwell B, Sasaki Y, Wood TO. Freeze-fracture quantitative comparison of rabbit corneal epi-thelial and endothelial membranes. Curr Eye Res.1985;4:951-961.
8. Beyer EC, Kistler J, Paul DL, Goodenough DA. Anti-sera directed against Connexin 43 peptides react witha 43 kD protein localized in gap junctions in myocar-dium and other tissues. / Cell Biol. 1989; 108:595-605.
9. Dong Y, Roos M, Gruijters T, et al. Differential expres-sion of two gap junction proteins in corneal epithe-lium. Eur J Cell Biol. 1994; 64:95-100.
10. Klyce SD. Electrical profiles in the corneal epithelium.JPhysiol (Lond). 1972; 226:407-429.
11. Wolosin JM. Gap junctions in rabbit corneal epithe-
lium: Limited permeability and inhibition by cAMP.Am JPhysiol. 1991; 261 :C857-C864.
12. Larson DM. Junctional communications and thewound healing response. In: DeMello WC, ed. CellIntercommunications. Boca Raton: CRC Press; 1990.
13. Guthrie SC, Gilula NB. Gap junctional communicationand development. Trends Neurosci. 1989; 12:12-16.
14. Kumar NM, Gilula NB. The gap junction communica-tion channel. Cell. 1996;84:381-388.
15. White TW, Bruzzone R, Goodenough DA, Paul DL.Mouse Cx50, a functional member of the connexinfamily of gap junction proteins, is the lens fiber pro-tein MP70. MolBiol Cell. 1992;3:711-720.
16. El-Fouly MH, Trosco JE, Chang CC. Scrape-loadingand dye transfer: A rapid and simple technique tostudy gap junctional intercellular communication. ExpCell Res. 1987; 168:422-430.
17. Klyce SD, Beuerman RW. Structure and function ofthe cornea. In: Kaufman HE, Barron BA, McDonaldMB, Waltman SR, eds. The Cornea. New York: ChurchillLivingstone; 1988:3-54.
18. Watsky MA. Keratocyte gapjunctional communicationin normal and wounded rabbit corneas and humancorneas. Invest Ophthalmol Vis Sci. 1995; 36:2568-2576.
19. Matic M, Petrov IN, Chen S, et al. Stem cells of thecorneal epithelium lack connexins and metabolictransfer capacity. Differentiation. 1997; in press
20. Swenson KI, Jordan JR, Beyer EC, Paul DL. Formationof gap junctions by expression of connexins in Xenopusoocyte pairs. Cell. 1987;57:145-155.
21. White TW, Bruzzone R, Wolfram S, Paul DL,Goodenough DA. Selective interactions among themultiple connexin proteins expressed in the verte-brate lens: The second extracellular domain is a deter-minant of compatibility between connexins. JCell Biol.1194; 125:879-892.
22. Gabbiani G, Chaponnier C, Huttner I. Cytoplasmicfilaments and gap junctions in epithelial cells and my-ofibroblasts during wound healing. JCell Biol. 1978; 76:561-568.
23. Goliger JA, Paul DL. Wounding alters epidermal con-nexin expression and gap junction-mediated intercel-lular communication. Mol Biol Cell. 1995; 6:1491-1501.
24. Goliger JA, Paul DL. Expression of gap junction pro-teins Cx26, Cx31.1, Cx37 and Cx43 in developing andmature epidermis. Dev Dyn. 1994; 20:1-13.
25. Berthoud VM, Ledbetter MLS, Hertzberg EL, Saez JC.Connexin 43 in MDCK cells: Regulation by a tumor-promoting phorbol ester and Ca2+. Eur J Cell Biol.1992; 57:40-50.
26. Laird DW, Puranam KL, Revel JP. Turnover and phos-phorylation of connexin 43 gap junction protein incultured cardiac myocytes. BiochemJ. 1991; 273:67-72.
27. Wilson SE, He YG, Lloyd SA. EGF, EGF receptor, basicFGF, TGF beta-1 and interleukin-1 alpha mRNA inhuman corneal epithelial cells and stromal fibroblasts.Invest Ophthalmol Vis Sci. 1992;33:1756-1765.
28. Roskelley CD, Desprez PY, Bissell MJ. Extracellularmatrix-dependent tissue-specific gene expression inmammary epithelial cells requires both physical and
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933197/ on 06/06/2018

Gap Junctions and Epithelial Wound Healing 609
biochemical signal transduction. Proc Natl Acad SciUSA. 1994;26:12378-12382.
29. Reaume AG, deSousa PA, Kulkarn S, et al. Cardiacmalformation in neonatal mice lacking connexin-43.Science. 1995; 267:1831-1834.
30. Hendy B, Maxfield FR. Regulation of neutrophil motility and adhesion by intracellular calcium transients.Blood-Cells. 1993; 19:143-161.
31. Wilson SE, Shultz GS, Chegini N, Weng J, He YG.
32.
Epidermal growth factor, transforming growth factoralpha, transforming growth factor beta, acidic fibro-blast growth factor, basic fibroblast growth factor, andinterleukin-1 proteins in the cornea. Exp Eye Res.1994; 59:63-70.Takahashi H, Kaminsky AE, Zieke JD. Expressionof glucose transporter is enhanced during cornealepithelial migration. Invest Ophthalmol Vis Sci. 1995;36(suppl):S699.
Announcement
JIN H. KINOSHITA LECTURESHIP
Call for Nominations
The National Foundation for Eye Research, an organization dedicated to the supportof cataract research, is pleased to announce a call for nominations for die 4di Jin H.Kinoshita Lectureship. The Trustees of die Foundation have approved the followingstatement concerning the Lectureship and die qualifications of nominees:
The Kinoshita Lectureship was established in 1989 by the National Foundation forEye Research to honor Dr. Jin H. Kinoshita for his outstanding contributions to visionresearch. It is given for meritorious research which contributes to the understanding ofthe etiology and prevention of cataract development. The recipient presents a lecture ofhis or her research at the biennial international U.S.-Japan Cooperative CataractResearch Meeting and is awarded an honorarium of $15,000 and a plaque.
A committee has been established to evaluate all nominees and to recommend twotop candidates to the Trustees of the Foundation, who will make the final selection.The committee consists of Leo T. Chylack, MD, James F. Hejtmancik, MD, PhD,andj. Samuel Zigler,Jr, PhD (Chair) with the President of the Foundation, VenkatN. Reddy, PhD, as an ex officio member without a vote. To nominate someone forthis award, send a letter outiining that person's relevant research accomplishmentsand a copy of his or her curriculum vitae to Samuel Zigler, 6 Center Drive MSC-2735, National Eye Institute, Bethesda, MD 20892-2735. The deadline for receiptof nominations is June 15, 1997.
Downloaded From: http://iovs.arvojournals.org/pdfaccess.ashx?url=/data/journals/iovs/933197/ on 06/06/2018