alfalfa roots induce nod genes of rhizobium - plant physiology
TRANSCRIPT

Plant Physiol. (1989) 91, 842-8470032-0889/89/91 /0842/06/$01 .00/0
Received for publication May 10, 1989and in revised form June 22, 1989
A Chalcone and Two Related Flavonoids Released fromAlfalfa Roots Induce nod Genes of Rhizobium meliloti'
Carl A. Maxwell, Ueli A. Hartwig, Cecillia M. Joseph, and Donald A. Phillips*Department of Agronomy and Range Science, University of California, Davis, California 95616
ABSTRACT
Flavonoid signals from alfalfa (Medicago sativa L.) induce tran-scription of nodulation (nod) genes in Rhizobium melioti. Previousinvestigations identified the flavone luteolin as an active inducerin alfalfa seed extracts, but the nature of nod inducers releasedfrom roots has not been reported. Root exudate from 3-day-oldalfalfa seedlings was purified and then assayed for biologicalactivity with a nodABC-IacZ fusion in R. meliloti. Indentities ofmajor nod inducers were established by spectroscopic analyses(ultraviolet/visible, proton nuclear magnetic resonance, and massspectroscopy) and comparison with authentic standards. Majornod inducers, which were identified as 4',7-dihydroxyflavone, 4'-7-dihydroxyflavanone, and 4,4'-dihydroxy-2'-methoxychalcone,were released from seedling roots at 54, 22, and 20 picomole.plantr'. day-1, respectively. Luteolin was not found in these rootexudates. The 4,4'-dihydroxy-2'-methoxychalcone induced nodgenes at a concentration one order of magnitude lower thanluteolin and is the first naturally released chalcone reported tohave this function. Moderate and weak nod-inducing activity wasassociated, respectively, with 4',7-dihydroxyflavone and 4',7-dihydroxyflavanone.
Alfalfa (Medicago sativa L.), an important leguminousforage crop throughout the world, forms N2-fixing root nod-ules in association with the soil bacterium Rhizobium meliloti.The earliest events of alfalfa nodule formation require expres-sion ofthe nodulation (nod) DABC genes on the megaplasmid(pSym) of R. meliloti (8, 16). Transcription of nodABC isinduced through the cooperative action of the constitutivenodD product and components of root and seed exudates(22). Luteolin, 3',4',5,7-tetrahydroxyflavone, was isolatedfrom alfalfa seed extracts and shown to participate in nodinduction (23), while different, commercially obtained, fla-vonoids had weaker inducing capabilities (23, 24). Closelyrelated compounds have similar functions with other Rhizo-bium- and Bradyrhizobium-legume associations (1, 9, 17, 25,27, 31).The availability of nod-inducing flavonoids in the rhizo-
sphere may limit alfalfa seedling growth. Kapulnik et al. (14)observed significant increases in nodulation, N2 fixation, andseedling growth of 'Hairy Peruvian' alfalfa when the rhizo-sphere was supplemented with luteolin. This is consistent with
' Supported in part by U.S. Department of Agriculture CRGOgrant 87-CRCR-1-2552 and grant IS-1348-87 from BARD, the U.S.-Israel Binational Agricultural Research and Development Fund. U.A. H. was supported by the Swiss National Research Foundation.
evidence that some N2-dependent alfalfa seedlings are initiallyN-limited due to insufficient root nodule formation (10) andsuggests that more nodules might be formed ifmore rhizobialcells are induced to initiate the infection process.The presence of active nod inducers in plants does not
guarantee release into the rhizosphere. Yelton et al. (30)observed that extracts from some plants induced nod tran-scription in R. meliloti even when exudates ofthe same plantsdid not. In contrast, both extracts and exudates of alfalfainduced nod genes, but it is unclear if luteolin was solelyresponsible for nod induction.
In order to better understand the process of nod-inducerrelease, it is necessary to identify active compounds actuallyexuded into the rhizosphere. The purpose of this study was toidentify, quantify, and characterize the major nod inducersexuded by roots of young, unnodulated alfalfa seedlings.
MATERIALS AND METHODS
Plant Culture
One g of alfalfa (Medicago sativa L.) seed (cv 'Moapa 69')containing about 400 seeds (94% viable) was scarified, surface-sterilized 3 min with 70% ethanol, rinsed with sterile water,and imbibed in sterile, aerated water. Imbibing solutions werechanged after 4 and 8 h to remove seed-derived compounds.At the end of 8 h, seeds were placed on a sterile cheesecloth-covered screen over 300 mL of sterile, aerated, N-free nutrientsolution (6; with sodium ferric diethylenetriamine pentaace-tate as the iron source, same concentration of iron) in aplastic, 473 mL, 'freezer' container. A sterile clear plastic sheetwas positioned over the assembled container and held in placewith a rubber band. Containers were maintained under anirradiance of 320 ,uE m22s-' (400-700 nm), 16/8 h light/dark, 25/20°C, and 50% RH. Solutions containing root exu-date were changed every 24 h for 8 d, sampled for nod-inducing activity, and stored at -20°C. At each sampling timefour replicates consisting of solutions from three containerswere collected.
Biological Activities
The nod-inducing capacity of root exudate and specificcompounds was assayed as f3-galactosidase activity from tran-scription of the nodC-lacZ fusion on plasmid pRmM57 inRhizobium meliloti strain 1021 (22), obtained from Dr. S. R.Long (Stanford University). Cultures were maintained onLMB (29) slants with yeast extract as the nitrogen source and10.8 ,ug tetracycline-mL-', before transfer to LMB liquid
842
Dow
nloaded from https://academ
ic.oup.com/plphys/article/91/3/842/6084565 by guest on 22 February 2022

ALFALFA ROOT FLAVONOIDS INDUCE RHIZOBIAL nod GENES
media, containing NH4NO3, at least 12 h prior to use in theassay.The i3-galactosidase assay was modified from Miller (21) as
previously reported (22). Volumes were adjusted to 100 ,uLwith 0.1 M sodium phosphate buffer (pH 7.0), and 375 gL ofR. meliloti culture in midlog phase (0.2-0.3 A60o) was added.Samples were incubated at room temperature with shakingfor 3 h, and 50 ,uL each of 0.1% (w/v) SDS and chloroformwere added.
Purification of nod Inducers
Root exudate samples were thawed, centrifuged at 6200gfor 15 min, and passed through 0.8 and 0.2 ,um polycarbonatefilters (Nuclepore Corp., Pleasanton, CA). Subsamples (50mL) were adsorbed to 900 mg C18 Maxi-Clean cartridges(Alltech Associates, Inc., Deerfield, IL). Flavonoids from eachcartridge were eluted with acetone, dried with a N2 gas stream(25°C), and dissolved in 400 ,uL of 50% methanol. Aliquots(100 ,gL) were loaded onto a Waters HPLC system (MilliporeCorp., Milford, MA) fitted with a 250 x 4.6 mm LichrosorbSRP 18 column (Phenomenex, Rancho Palos Verdes, CA) andeluted at 0.5 mL-min-' from 0 to 20 min with an isocraticsolvent system of 52.5:45:2.5 (v/v/v) water:methanol:aceticacid. From 20 to 30 min a linear gradient to 60:37.5:2.5 (v/v/v) methanol:water:acetic acid was applied, and the analysiscontinued isocratically at that concentration for another 30min. Eluting compounds were monitored with a Waters 990photodiode array detector, which measured absorbance (230-400 nm) every second with 1.4 nm resolution. Eluant fractionswere collected every 30 s, combined when associated withabsorbance peaks, dried under N2 (45C), redissolved in 500)L of 0.1 M sodium phosphate buffer (pH 7.0), and assayedfor nod induction. The quantity of nod-inducing activityassociated with a subsample of each HPLC absorbance peakwas related to the concentration of crude root exudate byassuming the peak represented the total amount ofa particularcompound injected into the HPLC.
Flavonoid nod inducers detected after initial HPLC analysesof root exudate were purified to homogeneity from exudateproduced between 48 and 72 h after seeds were exposed towater. Exudate was partitioned against hexane to removelipid, and flavonoids in the aqueous fraction were adsorbedto C18 cartridges. Acetone eluants from C18 cartridges werecombined and dried under N2 gas at 45°C. Flavonoids weresolubilized in 500 gL of 50% methanol and separated byHPLC on a 250 x 10mm Lichrosorb SRP18 semi-preparativecolumn (Alltech Associates, Inc.) with an elution rate of 2mL.min-'. HPLC solvents and gradients were identical tothose described above. Eluant fractions were collected every30 s, combined when associated with absorbance peaks, anddried either under N2 gas or with an Evapotec (Haake Buchler,Saddle Brook, NJ). Closely eluting contaminants were re-moved from nod inducers by a second HPLC separation onthe semipreparative column with appropriate concentrationsof methanol.
Fractions of interest were further purified by chromatogra-phy in 100% methanol on a column (1.5 x 25 cm) ofSephadex LH20 (Sigma Chemical Co., St. Louis, MO). Frac-
tions containing nod inducers were identified by spectralanalysis (200-500 nm) and dried under N2 gas.Some compounds were purified further by normal phase
HPLC with a column (250 x 4.6 mm) containing 5 umHypersil (Alltech Associates, Inc.) Following isocratic chro-matography in 100% chloroform for the first 10 min, a lineargradient to 90:9.5:0.5 (v/v/v) chloroform:methanol:water at30 min was produced. Compounds of interest were trackedby spectral analysis and dried under N2 gas.
Identification of nod Inducers
UV/visible spectral shift analyses (19) were done with aLambda 6 dual beam spectrophotometer (Perkin Elmer, Nor-walk, CT). Authentic standards were compared after tentativeidentifications. Standards were obtained from the followingsources: 4',7-dihydroxyflavone, 4',7-dihydroxyflavanone,and luteolin (Spectrum Chemical MFG Corp., Gardena, CA);2',4,4'-trihydroxychalcone (L. Jurd, USDA, Albany, CA);4,4'-dihydroxy-2'-methoxychalcone (R. Carlson, EcochemResearch, Inc., Chaska, MN).COSY2 and one-dimensional proton NMR experiments
were done in [U-2H]methanol on a Nicolet NT-360 spectrom-eter at the NMR Facility, University of California, Davis.One study of the 'strong' inducer was done, courtesy of Dr.J. Dallas, on a GE-NMR GN-500 Omega spectrometer at theGeneral Electric Co., Fremont, CA. Spectra were referencedto the methyl peak (3.30 ppm) of methanol.
Electron ionization MS data were collected by staff at theFacility for Advanced Instrumentation, University of Califor-nia, Davis using a solids probe to introduce the samples on aTrio-2 MS (VG Masslab, Altrincham, UK).
Flavonoid Concentrations
After identities of nod inducers were verified by compari-sons with authentic standards, their concentrations in rootexudate and in nod-induction assays were determined spec-trophotometrically in methanol from the following log e val-ues: 4,4'-dihydroxy-2'-methoxychalcone, 4.25 at 349 nm (4);4',7-dihydroxyflavone, 4.52 at 328 nm (15); 4',7-dihydroxy-flavanone, 4.14 at 275 nm (13); 2',4,4'-trihydroxychalcone4.20 at 370.5 nm (13). Luteolin concentrations were deter-mined in ethanol from a log c of 4.17 at 350 nm (19).Amounts of the nod inducers in root exudate were deter-
mined by HPLC with known quantities ofauthentic standardsby applying the integrator function within the Waters 990software package. Values were corrected for percent germi-nation and flavonoid recovery, as determined with knownamounts of 4',7-dihydroxyflavone processed in the normalmanner used for root exudate.
RESULTS
The nod-inducing activity of root exudate reached a maxi-mum during the third day after initiation of imbibition (Fig.
2Abbreviations: COSY, two-dimensional homonuclear shift cor-relation spectroscopy; 6H, chemical shift of proton; 2H, two protons,etc.; C-1, carbon one, etc.; d, doublet; dd, double doublet; s, singlet;m, multiplet; J, coupling constant; Hz, Hertz; m/z, mass charge ratio.
843
Dow
nloaded from https://academ
ic.oup.com/plphys/article/91/3/842/6084565 by guest on 22 February 2022
Plant Physiol. Vol. 91,1989
I*-ca
D1000C° 800
0 600
0° 400
la
0 2000.2 00
0 0 1 2 3 4 5 6 7 8Seedling Age (d)
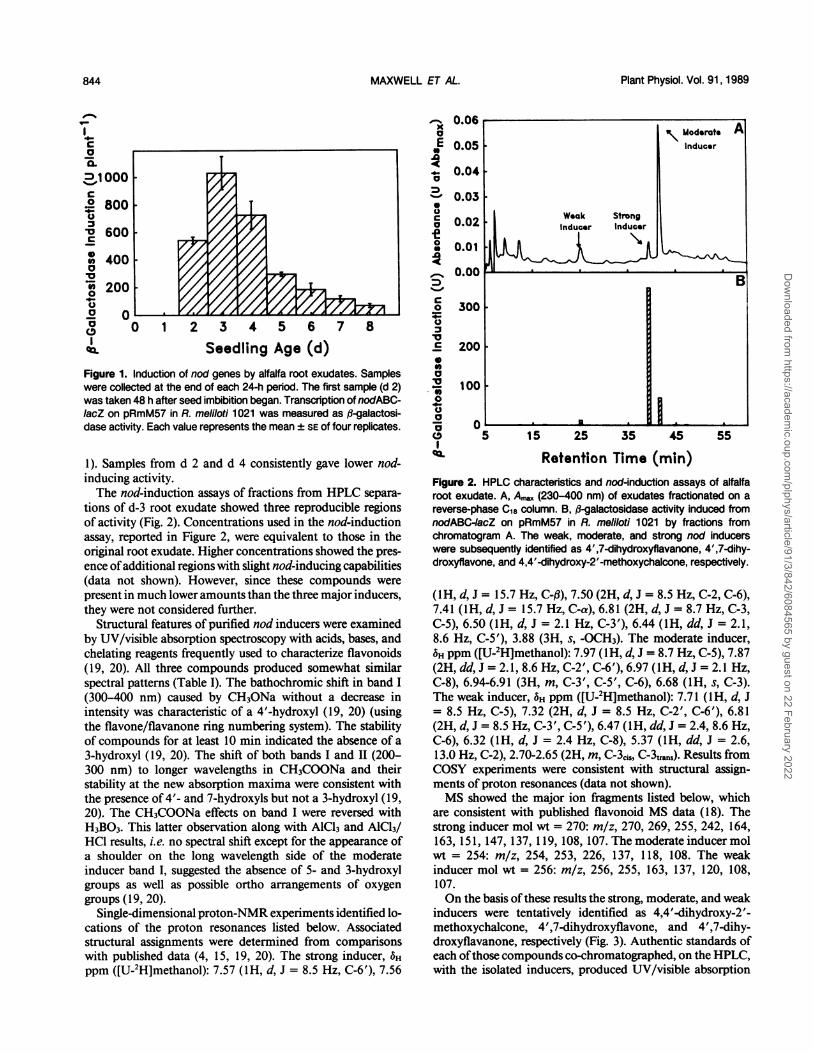
Figure 1. Induction of nod genes by alfalfa root exudates. Sampleswere collected at the end of each 24-h period. The first sample (d 2)was taken 48 h after seed imbibition began. Transcription of nodABC-/acZ on pRmM57 in R. meliloti 1021 was measured as jl-galactosi-dase activity. Each value represents the mean ± SE of four replicates.
1). Samples from d 2 and d 4 consistently gave lower nod-inducing activity.The nod-induction assays of fractions from HPLC separa-
tions of d-3 root exudate showed three reproducible regionsof activity (Fig. 2). Concentrations used in the nod-inductionassay, reported in Figure 2, were equivalent to those in theoriginal root exudate. Higher concentrations showed the pres-ence ofadditional regions with slight nod-inducing capabilities(data not shown). However, since these compounds werepresent in much lower amounts than the three major inducers,they were not considered further.
Structural features of purified nod inducers were examinedby UV/visible absorption spectroscopy with acids, bases, andchelating reagents frequently used to characterize flavonoids(19, 20). All three compounds produced somewhat similarspectral patterns (Table I). The bathochromic shift in band I(300-400 nm) caused by CH3ONa without a decrease inintensity was characteristic of a 4'-hydroxyl (19, 20) (usingthe flavone/flavanone ring numbering system). The stabilityof compounds for at least 10 min indicated the absence of a3-hydroxyl (19, 20). The shift of both bands I and II (200-300 nm) to longer wavelengths in CH3COONa and theirstability at the new absorption maxima were consistent withthe presence of 4'- and 7-hydroxyls but not a 3-hydroxyl (19,20). The CH3COONa effects on band I were reversed withH3BO3. This latter observation along with AlCl3 and AlCl3/HCI results, i.e. no spectral shift except for the appearance ofa shoulder on the long wavelength side of the moderateinducer band I, suggested the absence of 5- and 3-hydroxylgroups as well as possible ortho arrangements of oxygengroups ( 19, 20).
Single-dimensional proton-NMR experiments identified lo-cations of the proton resonances listed below. Associatedstructural assignments were determined from comparisonswith published data (4, 15, 19, 20). The strong inducer, 6Hppm ([U-2H]methanol): 7.57 (1H, d, J = 8.5 Hz, C-6'), 7.56
x 0.06aEc 0.05
<0.040
.00.02°0.04a
<
_. 0.00-
-C 200
204
cs.X 300
o0
la
_ Moderate A
Inducor
Weak StrongInducer Inducor
B
5 15 25 35 45 55
Retention Time (min)Figure 2. HPLC characteristics and nod-induction assays of alfalfaroot exudate. A, A, (230-400 nm) of exudates fractionated on areverse-phase C18 column. B, ,-galactosidase activity induced fromnodABC4acZ on pRmM57 in R. meliloti 1021 by fractions fromchromatogram A. The weak, moderate, and strong nod inducerswere subsequently identified as 4',7-dihydroxyflavanone, 4',7-dihy-droxyflavone, and 4,4'-dihydroxy-2'-methoxychalcone, respectively.
(1H, d, J = 15.7 Hz, C-,B), 7.50 (2H, d, J = 8.5 Hz, C-2, C-6),7.41 (lH, d, J = 15.7 Hz, C-a), 6.81 (2H, d, J = 8.7 Hz, C-3,C-5), 6.50 (1H, d, J = 2.1 Hz, C-3'), 6.44 (1H, dd, J = 2.1,8.6 Hz, C-5'), 3.88 (3H, s, -OCH3). The moderate inducer,6H ppm ([U-2H]methanol): 7.97 (1H, d, J = 8.7 Hz, C-5), 7.87(2H, dd, J = 2.1, 8.6 Hz, C-2', C-6') 6.97 (IH, d, J = 2.1 Hz,C-8), 6.94-6.91 (3H, m, C-3', C-5', C-6), 6.68 (IH, s, C-3).The weak inducer, 5H ppm ([U-2H]methanol): 7.71 (1H, d, J= 8.5 Hz, C-5), 7.32 (2H, d, J = 8.5 Hz, C-2', C-6'), 6.81(2H, d, J = 8.5 Hz, C-3', C-5'), 6.47 (IH, dd, J = 2.4, 8.6 Hz,C-6), 6.32 (1H, d, J = 2.4 Hz, C-8), 5.37 (IH, dd, J = 2.6,13.0 Hz, C-2), 2.70-2.65 (2H, m, C-3cis, C-3ans,). Results fromCOSY experiments were consistent with structural assign-ments of proton resonances (data not shown).MS showed the major ion fragments listed below, which
are consistent with published flavonoid MS data (18). Thestrong inducer mol wt = 270: m/z, 270, 269, 255, 242, 164,163, 151, 147, 137, 119, 108, 107. Themoderateinducermolwt = 254: m/z, 254, 253, 226, 137, 118, 108. The weakinducer mol wt = 256: m/z, 256, 255, 163, 137, 120, 108,107.On the basis of these results the strong, moderate, and weak
inducers were tentatively identified as 4,4'-dihydroxy-2'-methoxychalcone, 4',7-dihydroxyflavone, and 4',7-dihy-droxyflavanone, respectively (Fig. 3). Authentic standards ofeach ofthose compounds co-chromatographed, on the HPLC,with the isolated inducers, produced UV/visible absorption
844 MAXWELL ET AL.
Dow
nloaded from https://academ
ic.oup.com/plphys/article/91/3/842/6084565 by guest on 22 February 2022

ALFALFA ROOT FLAVONOIDS INDUCE RHIZOBIAL nod GENES
Table I. UVIVisible Absorption Maxima of nod InducersPurified nod inducers were treated with shift reagents in methanol, using published procedures (19).
InducerShift Reagent
Strong Moderate Weak
Xmax, nm
Methanol 230, 348 228sha, 252sh, 312sh, 327 213, 230, 275, 313CH3ONa 247, 418 251, 264sh, 327, 387 249, 299sh, 335AICI3 233, 349 228sh, 252sh, 313sh, 327, 382sh 213, 231, 275, 311AICl3/HCI 235, 351 229sh, 31 3sh, 327, 395 231, 275, 311CH3COONa 341sh, 392 263, 310, 317sh, 373 250, 286sh, 335CH3COONa/H3BO3 351 255sh, 313sh, 329 276, 312
aSpectral shoulder.
HO O 0
0
4',7-Dthydroxyf lavanone
,OH
HO 0O<
4' 7-Dihydroxyf lavone
OOH
HO nOCHi
0
4,4'-D1hydroxy-2'-methoxychalconeFigure 3. Structures of 4',7-dihydroxyflavanone, 4',7-dihydroxyfla-vone, and 4,4'-dihydroxy-2'-methoxychalcone, the weak, moderate,and strong nod inducers, respectively, isolated from alfalfa rootexudate.
spectra equivalent to those of the purified inducers within theerror of the shift reagent method (19), and provided proton-NMR spectra identical to those ofpurified nod inducers. Mostimportantly, the authentic standards showed nod-inducingactivity comparable to that of putative compounds from root
exudate when assayed at the same concentrations (Table II).Additional assays for nodC-lacZ induction with more concen-
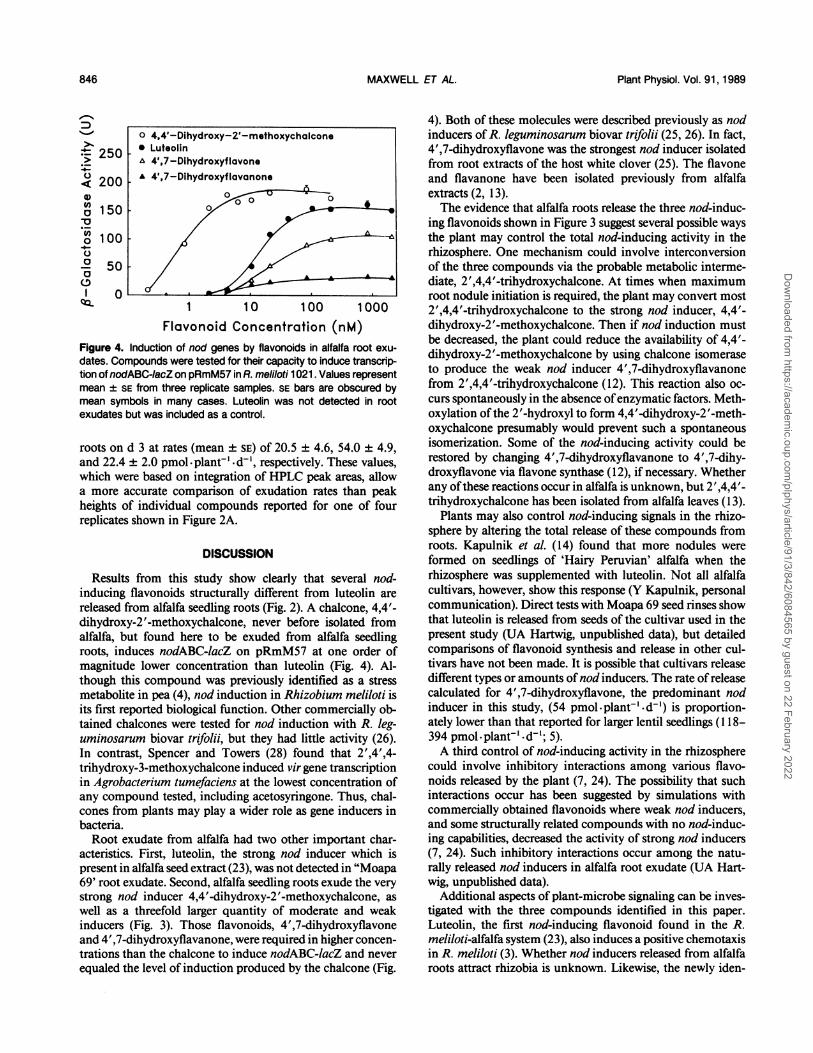
trations of the authentic compounds increased the resolutionof the biological activity (Fig. 4).
Luteolin was not detected in root exudate, even whenHPLC eluants were tested at higher concentrations. Neverthe-less, it was included in nod-induction assays as a standard forcomparison (Fig. 4). The stronger inducer, 4,4'-dihydroxy-2'-methoxychalcone, induced nod genes at an order of magni-tude lower concentration than luteolin. This difference was
consistent over numerous assays, although the specific con-
centrations associated with the half maximum for nod induc-tion sometimes varied slightly.When 2',4,4'-trihydroxychalcone, a compound structurally
related to the three major nod inducers but not obviouslypresent in root exudate, was assayed for nod-inducing activity,it showed less activity than 4,4'-dihydroxy-2'-methoxychal-cone but substantially more than 4',7-dihydroxyflavanone.For example, in one assay (0.2 nM-2. 1 uM) 2',4,4'-trihydrox-ychalcone had a maximum activity of 96.1 ± 3.4 (mean ±
SE) ,B-galactosidase units, while 4,4'-dihydroxy-2'-methoxy-chalcone had 110 ± 2.8 units and 4',7-dihydroxyflavanonehad 11.8 ± 0.6 units. The concentrations required for half-maximum nod induction were 9, 2, and 11 nm for 2',4,4'-trihydroxychalcone, 4,4'-dihydroxy-2'-methoxychalcone,and 4',7-dihydroxyflavanone, respectively.The 4,4'-dihydroxy-2'-methoxychalcone, 4',7-dihydroxy-
flavone, and 4',7-dihydroxyflavanone were released from
Table II. Comparison of nod-Inducing Activity for Putative andAuthentic Flavonoids
Values represent means for three replicates.4,4'-dihydroxy-2'- 4',7-dihydroxy- 4',7-dihydroxy-
Concentration methoxychalcone flavanone flavonePutative Authentic Putative Authentic Putative Authentic
nM units f-galactosidase0.2 8.8 7.72.1 73.2 61.6 1.6 0.8 2.2 1.74.2 83.7 87.9 9.7 6.7 21.0 14.810.5 103 106 8.2 7.6 27.9 27.0210 113 112 11.7 9.7 40.1 31.02100 12.9 10.1 48.7 49.5LSD (0.05) 5.6 0.9 3.2
845
Dow
nloaded from https://academ
ic.oup.com/plphys/article/91/3/842/6084565 by guest on 22 February 2022
Plant Physiol. Vol. 91,1989
:t 250-a< 2000G)a 150
-.o100
4-O
U50a
I 01 10 100 1000
Flavonoid Concentration (nM)Figure 4. Induction of nod genes by flavonoids in alfalfa root exu-dates. Compounds were tested for their capacity to induce transcrip-tion of nodABC-lacZ on pRmM57 in R. meliloti 1021. Values representmean ± SE from three replicate samples. SE bars are obscured bymean symbols in many cases. Luteolin was not detected in rootexudates but was included as a control.
roots on d 3 at rates (mean ± SE) of 20.5 ± 4.6, 54.0 ± 4.9,and 22.4 ± 2.0 pmol-plant'-dd', respectively. These values,which were based on integration of HPLC peak areas, allowa more accurate comparison of exudation rates than peakheights of individual compounds reported for one of fourreplicates shown in Figure 2A.
DISCUSSION
Results from this study show clearly that several nod-inducing flavonoids structurally different from luteolin are
released from alfalfa seedling roots (Fig. 2). A chalcone, 4,4'-dihydroxy-2'-methoxychalcone, never before isolated fromalfalfa, but found here to be exuded from alfalfa seedlingroots, induces nodABC-lacZ on pRmM57 at one order ofmagnitude lower concentration than luteolin (Fig. 4). Al-though this compound was previously identified as a stressmetabolite in pea (4), nod induction in Rhizobium meliloti isits first reported biological function. Other commercially ob-tained chalcones were tested for nod induction with R. leg-uminosarum biovar trifolii, but they had little activity (26).In contrast, Spencer and Towers (28) found that 2',4',4-trihydroxy-3-methoxychalcone induced vir gene transcriptionin Agrobacterium tumefaciens at the lowest concentration ofany compound tested, including acetosyringone. Thus, chal-cones from plants may play a wider role as gene inducers inbacteria.Root exudate from alfalfa had two other important char-
acteristics. First, luteolin, the strong nod inducer which ispresent in alfalfa seed extract (23), was not detected in "Moapa69' root exudate. Second, alfalfa seedling roots exude the verystrong nod inducer 4,4'-dihydroxy-2'-methoxychalcone, as
well as a threefold larger quantity of moderate and weakinducers (Fig. 3). Those flavonoids, 4',7-dihydroxyflavoneand 4',7-dihydroxyflavanone, were required in higher concen-trations than the chalcone to induce nodABC-lacZ and never
equaled the level of induction produced by the chalcone (Fig.
4). Both of these molecules were described previously as nodinducers of R. leguminosarum biovar trifolii (25, 26). In fact,4',7-dihydroxyflavone was the strongest nod inducer isolatedfrom root extracts of the host white clover (25). The flavoneand flavanone have been isolated previously from alfalfaextracts (2, 13).The evidence that alfalfa roots release the three nod-induc-
ing flavonoids shown in Figure 3 suggest several possible waysthe plant may control the total nod-inducing activity in therhizosphere. One mechanism could involve interconversionof the three compounds via the probable metabolic interme-diate, 2',4,4'-trihydroxychalcone. At times when maximumroot nodule initiation is required, the plant may convert most2',4,4'-trihydroxychalcone to the strong nod inducer, 4,4'-dihydroxy-2'-methoxychalcone. Then if nod induction mustbe decreased, the plant could reduce the availability of 4,4'-dihydroxy-2'-methoxychalcone by using chalcone isomeraseto produce the weak nod inducer 4',7-dihydroxyflavanonefrom 2',4,4'-trihydroxychalcone (12). This reaction also oc-curs spontaneously in the absence ofenzymatic factors. Meth-oxylation ofthe 2'-hydroxyl to form 4,4'-dihydroxy-2'-meth-oxychalcone presumably would prevent such a spontaneousisomerization. Some of the nod-inducing activity could berestored by changing 4',7-dihydroxyflavanone to 4',7-dihy-droxyflavone via flavone synthase (12), if necessary. Whetherany ofthese reactions occur in alfalfa is unknown, but 2',4,4'-trihydroxychalcone has been isolated from alfalfa leaves (13).
Plants may also control nod-inducing signals in the rhizo-sphere by altering the total release of these compounds fromroots. Kapulnik et al. (14) found that more nodules wereformed on seedlings of 'Hairy Peruvian' alfalfa when therhizosphere was supplemented with luteolin. Not all alfalfacultivars, however, show this response (Y Kapulnik, personalcommunication). Direct tests with Moapa 69 seed rinses showthat luteolin is released from seeds of the cultivar used in thepresent study (UA Hartwig, unpublished data), but detailedcomparisons of flavonoid synthesis and release in other cul-tivars have not been made. It is possible that cultivars releasedifferent types or amounts ofnod inducers. The rate of releasecalculated for 4',7-dihydroxyflavone, the predominant nodinducer in this study, (54 pmol.plant-' .d') is proportion-ately lower than that reported for larger lentil seedlings (1 18-394 pmol-plantr' d-'; 5).A third control of nod-inducing activity in the rhizosphere
could involve inhibitory interactions among various flavo-noids released by the plant (7, 24). The possibility that suchinteractions occur has been suggested by simulations withcommercially obtained flavonoids where weak nod inducers,and some structurally related compounds with no nod-induc-ing capabilities, decreased the activity of strong nod inducers(7, 24). Such inhibitory interactions occur among the natu-rally released nod inducers in alfalfa root exudate (UA Hart-wig, unpublished data).
Additional aspects of plant-microbe signaling can be inves-tigated with the three compounds identified in this paper.Luteolin, the first nod-inducing flavonoid found in the R.meliloti-alfalfa system (23), also induces a positive chemotaxisin R. meliloti (3). Whether nod inducers released from alfalfaroots attract rhizobia is unknown. Likewise, the newly iden-
846 MAXWELL ET AL.
Dow
nloaded from https://academ
ic.oup.com/plphys/article/91/3/842/6084565 by guest on 22 February 2022

ALFALFA ROOT FLAVONOIDS INDUCE RHIZOBIAL nod GENES
tified nod-inducing flavonoids from alfalfa root exudate mayhelp to clarify the apparent specificity of the three nodDalleles of R. meliloti for different flavonoids (1 1).
ACKNOWLEDGMENTS
The authors thank Drs. M. Berhow, S. Cunningham, R. Horowitz,and L. Jurd and associates for helpful discussions on flavonoidpurification and identification. Dr. J. DeRopp and staff of the UCDavis NMR Facility provided instruction and helpful discussions onNMR. We are especially indebted to Dr. R. Carlson for his expeditiousgift of 4,4'-dihydroxy-2'-methoxychalcone.
LITERATURE CITED
1. Bassam BJ, Djordjevic MA, Redmond JW, Batley M, Rolfe BG(1988) Induction of a nodD-dependent locus in the Rhizobiumstrain NGR234 activated by phenolic factors secreted by soy-beans and other legumes. Mol Plant-Microb Interact 1:16 1-168
2. Bickoff EM, Livingston AL, Witt SC (1965) Identification of7,4'-dihydroxyflavone in forages. Phytochemistry 4: 523-524
3. Caetano-Annolles G, Christ-Estes DK, Bauer WD (1988)Chemotaxis ofRhizobium meliloti to the plant flavone luteolinrequires functional nodulation genes. J Bacteriol 170:3164-3169
4. Carlson RE, Dolphin DH (1982) Pisum sativum stress metabo-lites: two cinnamylphenols and a 2'-methoxychalcone. Phy-tochemistry 21: 1733-1736
5. D'Arcy-Lameta A (1986) Study of soybean and lentil root exu-dates. II Identification of some polyphenolic compounds, re-lation with plantlet physiology. Plant Soil 92: 113-123
6. Dejong TM, Phillips DA (1981) Nitrogen stress and apparentphotosynthesis in symbiotically grown Pisum sativum L. PlantPhysiol 68: 309-313
7. Djordjevic MA, Redmond JW, Batley M, Rolfe BG (1987)Clovers secrete specific phenolic compounds which either stim-ulate or repress nod gene expression in Rhizobium trifolii.EMBOJ6: 1173-1179
8. Dudley ME, Jacobs TW, Long SR (1987) Microscopic studiesof cell divisions induced in alfalfa roots by Rhizobium meliloti.Planta 171: 289-301
9. Firmin JL, Wilson KE, Rossen L, Johnston AWB (1986) Fla-vonoid activation of nodulation genes in Rhizobium reversedby other compounds present in plants. Nature 324: 90-92
10. Fishbeck KA, Phillips DA (1982) Combined nitrogen and vege-tative regrowth of symbiotically-grown alfalfa. Agron J 73:975-978
1 1. Gyorgypal Z, Iyer N, Kondorosi A (1988) Three regulatory nodDalleles of diverged flavonoid-specificity are involved in host-dependent nodulation by Rhizobium meliloti. Mol Gen Genet212: 85-92
12. Heller W, Forkmann G (1988) Biosynthesis. In JB Harborne, ed,The Flavonoids. Advances in Research Since 1980. Chapmanand Hall Ltd, London, pp 399-425
13. Ingham JL (1979) Isoflavonoid phytoalexins of the genus Medi-cago. Biochem Syst Ecol 7: 29-34
14. Kapulnik Y, Joseph CM, Phillips DA (1987) Flavone limitationsto root nodulation and symbiotic nitrogen fixation in alfalfa.Plant Physiol 84: 1193-1196
15. Kobayashi A, Yata S, Kawazu K (1988) A ,B-hydroxychalconeand flavonoids from alfalfa callus stimulated by a fungalnaphthoquinone, PO-1. Agric Biol Chem 52: 3223-3227
16. Kondorosi E, Banfalvi Z, Kondorosi A (1984) Physical and geneticanalysis of a symbiotic region of Rhizobium meliloti: Identi-fication of nodulation genes. Mol Gen Genet 193:445-452
17. Kosslak RM, Bookland R, Barkei J, Paaren HE, Appelbaum ER(1987) Induction of Bradyrhizobiumjaponicum common nodgenes by isoflavones isolated from Glycine max. Proc NatlAcad Sci USA 84: 7428-7432
18. Mabry TJ, Markham KR (1975) Mass spectrometry of flavo-noids. In JB Harborne, TJ Mabry, H Mabry, ed, The Flavo-noids, Ed 1, Vol 1. Academic Press, New York, pp 78-126
19. Mabry TJ, Markham KR, Thomas MB (1970) The SystematicIdentification of Flavonoids. Springer-Verlag, New York
20. Markham KR, Mabry TJ (1975) Ultraviolet-visible and protonmagnetic resonance spectroscopy of flavonoids. In JB Har-borne, TJ Mabry, H Mabry, eds, The Flavonoids, Ed 1, Vol 1.Academic Press, New York, pp 45-77
21. Miller JH (1972) Experiments in Molecular Genetics. ColdSpring Harbor Laboratory, New York, pp 352-355
22. Mulligan JT, Long SR (1985) Induction of Rhizobium melilotinodC expression by plant exudate requires nodD. Proc NatlAcad Sci USA 82: 6609-6613
23. Peters NK, Frost JW, Long SR (1986) A plant flavone, luteolin,induces expression of Rhizobium meliloti nodulation genes.Science 233: 977-980
24. Peters NK, Long SR (1988) Alfalfa root exudates and compoundswhich promote or inhibit induction of Rhizobium melilotinodulation genes. Plant Physiol 88: 396-400
25. Redmond JW, Batley M, Djordjevic MA, Innes RW, KuempelPL, Rolfe BG (1986) Flavones induce expression ofnodulationgenes in Rhizobium. Nature 323: 632-635
26. Redmond JW, Batley M, Innes RW, Kuempel PL, DjordjevicMA, Rolfe BG (1986) Flavones induce expression of the nod-ulation genes in Rhizobium. In B Lugtenberg, ed, Recognitionin Microbe-Plant Symbiotic and Pathogenic Interactions,NATO ASI Series, Vol H4. Springer-Verlag, Berlin, pp115-121
27. Sadowsky MJ, Olson ER, Foster VE, Kosslak RM, Verma DP(1988) Two host-inducible genes of Rhizobium fredii andcharacterization of the inducing compound. J Bacteriol 170:171-178
28. Spencer TA, Towers GHN (1988) Specificity of signal com-pounds detected by Agrobacterium tumefaciens. Phytochem-istry 27: 2781-2785
29. Vincent JM (1970) A Manual for the Practical Study of Root-Nodule Bacteria, IBP Handbook, Vol 15. Blackwell ScientificPublications, Oxford
30. Yelton MM, Mulligan JT, Long SR (1987) Expression of Rhi-zobium meliloti nod genes in Rhizobium and Agrobacteriumbackgrounds. J Bacteriol 169: 3094-3098
31. Zaat SA, Wijffelman CA, Spaink HP, Van Brussel AAN, OkkerRJH, Lugtenberg BJJ (1987) Induction of the nodA promoterof Rhizobium leguminosarum Sym plasmid pRLlJI by plantflavanones and flavones. J Bacteriol 169: 198-204
847
Dow
nloaded from https://academ
ic.oup.com/plphys/article/91/3/842/6084565 by guest on 22 February 2022