age-related association of rdna and telomeres with the
TRANSCRIPT

Age-related association of rDNA and telomeres with thenuclear matrix in mouse hepatocytesAlberto S. Moraes12*,1, Mateus Mondin1{, Marcelo E. Beletti11, Margarida L.R. Aguiar-Perecin{, Ana M.A. Guaraldo{ andMaria Luiza S. Mello** Department of Cell Biology, Institute of Biology, State University of Campinas (UNICAMP), 13083-863 Campinas, Sao Paulo, Brazil{ Department of Parasitology, Institute of Biology, State University of Campinas (UNICAMP), 13083-863 Campinas, Sao Paulo, Brazil{ Department of Genetics, Superior Agriculture School Luiz de Queiroz, University of Sao Paulo, 13418-900 Piracicaba, SP, Brazil1
Department of Morphology, Institute of Biomedical Sciences, Federal University of Uberlandia, 38405-320 Uberlandia, MG, Brazil
AbstractTranscribed sequences have been suggested to be associated with the nuclear matrix, differing from non-transcribing
sequences, which have been reported to be contained in DNA loops. However, although a dozen of genes have their
expression level affected by aging, data on chromatin–nuclear matrix interactions under this physiological condition are still
scarce. In the present study, liver imprints from young, adult and old mice were subjected to FISH (fluorescence in situ
hybridization) for 45S rDNA and telomeric sequences, with or without a lysis treatment to produce extended chromatin
fibres. There was an increased amount of 45S rDNA sequences located in DNA loops as the animals grow older, while
telomeric sequences were always observed in DNA loops irrespective of the animal age. We assume that active rRNA
genes associate with the nuclear matrix, while DNA loops contain silent sequences. Transcription of each 45S rDNA repeat
unit is suggested to be dependent on its interaction with the nuclear matrix.
Keywords: aging; chromatin extensibility; development; nuclear matrix; rDNA; telomere
1. Introduction
A prevailing theory on aging states that it results from an
accumulation of tissue damage, similar to rust. This presumes
that aging results inevitably from accumulated damage to cells as
promoted by toxins, free-radical molecules, DNA-damaging
radiation, disease and stress (Harman, 1998). Nevertheless, there
is increasing evidence that specific genetic instructions and
structural modifications drive the process making it an active
process rather than a passive one (Oberdoerffer and Sinclair,
2007).
Studies by several authors, conducted in yeast, worms, flies
and mice, have confirmed substantial changes in gene expression
with aging (Burzynski, 2005). Among most important changes
reported is the silencing of tumour suppressors and other genes
involved in the control of cell cycle, apoptosis, detoxification and
cholesterol metabolism (Cao et al., 2001).
The activity of rRNA genes in mouse hepatocytes gradually
increases after birth, reaches a peak at 14 days of postnatal age
and then decreases drastically as the animals grow older (Ma and
Nagata, 1990). These changes are accompanied by an increase in
protein synthesis at about 1 month postnatally, followed by a
decrease in this synthesis along the adult life and low synthesis
levels in aged animals (Ma et al., 1991).
The rRNA genes, which in most eukaryotic species are
repeated ,100–5000 times per haploid genome, are located at
one or a few chromosomal sites called NORs (nucleolar organizer
regions) (Gaubatz et al., 1976; Long and Dawid, 1980). The rRNA
coding regions are organized as head-to-tail repeats, with the
transcribed regions separated by segments of non-transcribed
spacers (Fedoroff, 1979; Pan and Zhang, 2008). In rat hepato-
cytes, these sequences appear enriched in the purified nuclear
matrices after DNase I digestion, thus indicating that the high
transcription rates of rRNA genes and nuclear matrix interaction
are interconnected (Pardoll and Vogelstein, 1980).
The nuclear matrix is an operationally defined nuclear skeletal
structure assumed to be involved in many nuclear functions,
including DNA replication, transcription and repair and pre-mRNA
processing/transport. It is also a passenger concourse for
proteins going in and out of the nucleus through the nuclear pore
complex (Tsutsui et al., 2005). The nuclear matrix comprises a
protein framework formed by the nuclear pore/nuclear lamina
complex, a residual nucleolus and a residual internal fibrillar–
globular mesh (Berezney and Coffey, 1974; Gasser and Laemmli,
1986; Berezney, 1991). Until relatively recently, the nuclear matrix
was considered a rigid and static structure, but new theories now
consider it as a dynamic structure (Tsutsui et al., 2005; Barboro
et al., 2009).
The isolation of the nuclear matrix requires nuclease and high-
salt treatments that remove chromatin and other soluble nuclear
proteins from the cell nuclei (Nickerson, 2001). An ordinary method
for nuclear matrix isolation involves the extraction of nuclei with
highly concentrated solutions of monovalent salts such as 2 M NaCl
(Berezney and Coffey, 1977). After that, the nuclei adopt a so-called
nuclear halo configuration, in which nuclear DNA sticks out from the
residual structure (nuclear matrix) as negatively supercoiled loops
attached at their base by MARs (matrix attachment regions). If
1 These authors contributed equally to this work.2 To whom correspondence should be addressed (email [email protected]).Abbreviations: DAPI, 49,6-diamidino-2-phenylindole; e, erythroblast; ECF, extended chromatin fibre; FISH, fluorescence in situ hybridization; MAR, matrixattachment region; n, nucleoli.
Cell Biol. Int. (2010) 34, 925–931 (Printed in Great Britain)
Research Article
E The Author(s) Journal compilation E 2010 Portland Press Limited Volume 34 (9) N pages 925–931 N doi:10.1042/CBI20090457 N www.cellbiolint.org 925

nuclei are treated vertically, under the action of gravity, formation of
ECFs (extended chromatin fibres) may occur, as they are strongly
affected by nuclear matrix proteins, thus contributing to the higher-
order chromatin structure (Gerdes et al., 1994; Haaf and Ward,
1994; Davie, 1995; Cremer et al., 2000; Vidal, 2000). An increased
number of interactions between chromatin and nuclear matrix can
render or even prevent the ECF formation, since there are salt-
resistant interactions of MARs with the nuclear matrix (Moraes et
al., 2005, 2007). After this extraction, the nuclear DNA can be
fractionated into loop DNA and matrix-associated DNA; the latter
accounts for 1% of total nuclear DNA and is enriched in rRNA
sequences (Pardoll and Vogelstein, 1980). When this treatment is
followed by FISH (fluorescence in situ hybridization) assay,
transcriptionally inactive sequences produce long strings of signal
extending out on to the DNA halo or loop, whereas active
sequences remain tightly condensed as single spots within the
residual nucleus, thus suggesting that the association of genes with
the nuclear matrix may be a factor responsible for transcription
activation (Gerdes et al., 1994).
Taking into account that differences have previously been
reported for the changes in the activity of rRNA genes with
advancing age, in the present work, we aimed at comparing FISH
signals of active and silenced 45S rDNA and repetitive silenced
telomeric sequences in relation to their attachment to nuclear
matrix after ECF formation was induced in young, adult and old
mice hepatocytes.
2. Experimental procedures
2.1. Animals
Male mice from the inbred strain A/Uni, obtained from the
Multidisciplinary Center for Biological Investigation (CEMIB) of
the State University of Campinas were reared under normal
conditions and fed extruded chow (Purinaj) ad libitum. The
animals used were 1–2 (young), 15–20 (adult) and 61–100 (aged)
weeks old. At least three animals of each group were used.
Animals were killed by decapitation. Their livers were then
immediately removed and placed in cold 0.9% NaCl solution.
Liver slices were imprinted on histological slides. All protocols
involving animal care and use were approved by the Institutional
Committee for Ethics in Animal Experimentation (CEEA/IB/
UNICAMP – 378-1).
2.2. Treatments
Freshly prepared imprints were fixed in a mixture of absolute
ethanol and acetic acid (3:1, v/v, respectively) for 1 min and then
rinsed in 70% ethanol for 5 min. The preparations were positioned
vertically using a protocol previously described for ECF invest-
igation (Moraes et al., 2005). Briefly, the lysis solution consisted of
2 M NaCl plus 1% Triton X-100 in Tris/HCl buffer (25 mM, pH 7.4).
Treatment lasted for 5 h at 25uC, after which, the volume of
solution was completed with absolute ethanol to a final concen-
tration of 50%, and treatment was prolonged for another 10 min.
The slides were then carefully removed from the lysis solution and
transferred to 70% ethanol for 30 min. Fixed preparations not
subjected to the lysis protocol were used as controls.
2.3. FISH
The plasmid HM 456, which contains part of the 18S and 28S
rDNA of Xenopus laevis (Meunier-Rotival et al., 1979) was kindly
provided by Dr A.M. Vianna-Morgante (IB-USP, Sao Paulo, Brazil),
and the telomere-like oligonucleotide DNA sequence was syn-
thesized by Invitrogen. Probes were labelled by nick translation
with biotin-16dUTP (Bionick Labeling Kit, Life Technologies, Inc.).
The protocol used was previously described by Mondin et al.
(2007). Briefly, the preparations were treated with RNase A (Sigma
Chemical Co.), followed by treatment with 0.01 M HCl for 2 min,
washed in 26 SSC (16 SSC is 0.15 M NaCl/0.015 M sodium
citrate), treated with 5 mg/ml pepsin for 10 min at 37uC and fixed in
4% paraformaldehyde for 5 min, followed by denaturation with
26 SSC/50% formamide for 10 min at 80uC. Heat-denatured
probes were hybridized at 37uC for 20 h. After hybridization and
washes with 26 SSC/50% formamide and 26 SSC at 42uC,
probe detection used a two-step procedure for amplification.
Biotin was revealed using mouse anti-biotin and rabbit anti-mouse
TRITC conjugated antibodies. Coverslips were mounted in
Vectashield (Vector Laboratories) containing 0.5 mg/ml DAPI
(49,6-diamidino-2-phenylindole).
2.4. Microscopy
DNA FISH preparations were analysed under a Zeiss Axiophot 2
epifluorescence microscope equipped with an HBO-100 W
stabilized mercury lamp as the light source and appropriate
filters. Zeiss Plan 625/0.50, 640/0.75 and 6100/1.4 oil-
immersion objectives were used. Digital images were obtained
using a Sony CCD-IRIS/RGB Hyper HAD colour video camera and
ISIS (MetaSystems, Altlussheim) software. Selected images were
processed using CorelDrawj12 software.
2.5. Image processing
Images of DAPI-stained nuclei were segmented using supervised
threshold to mark the nuclear region. Areas with average
intensities above this threshold were set to 1 (nuclear region),
and all other areas were set to 0. The resulting images were called
DAPI mask.
FISH images for rDNA were converted into greyscale format
and were segmented using supervised threshold to determine
the region positive for this marker. The sum of pixel values of this
region was calculated and was called IGL. The sum of pixel values
with overlapping DAPI mask of value 1 was calculated, and its
relative IGL percentage was called %IGLin and corresponds to the
percentage of integrated grey levels of FISH signals inside nuclear
remnants. The sum of pixel values with overlapping DAPI mask with
value 0 was called %IGLout and corresponds to the percentual of
integrated grey levels of FISH signals outside nuclear remnants
(extended chromatin fibres and nuclear halos).
Aging and DNA/nuclear matrix
926 www.cellbiolint.org N Volume 34 (9) N pages 925–931 E The Author(s) Journal compilation E 2010 Portland Press Limited
2.6. Statistics
All calculations and statistical analyses were done with Minitab 14
(State College, PA). The Mann–Whitney test was used.
3. Results
3.1. rDNA sequences detach from the nuclear matrixwith development and aging
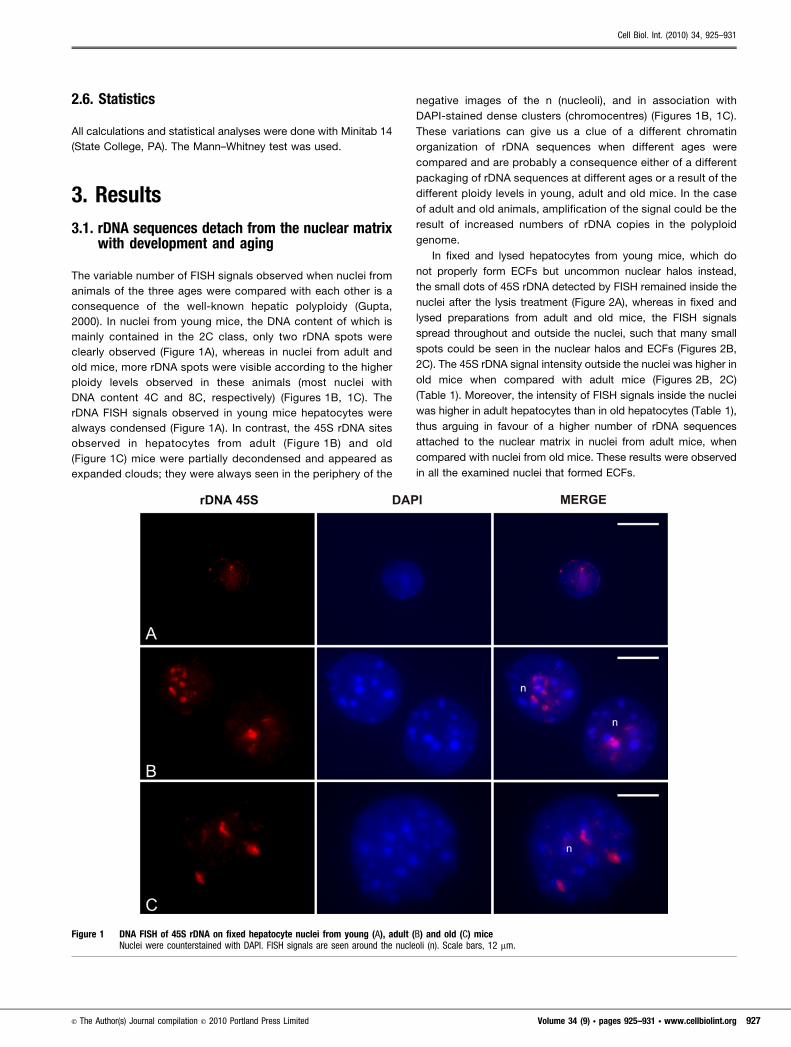
The variable number of FISH signals observed when nuclei from
animals of the three ages were compared with each other is a
consequence of the well-known hepatic polyploidy (Gupta,
2000). In nuclei from young mice, the DNA content of which is
mainly contained in the 2C class, only two rDNA spots were
clearly observed (Figure 1A), whereas in nuclei from adult and
old mice, more rDNA spots were visible according to the higher
ploidy levels observed in these animals (most nuclei with
DNA content 4C and 8C, respectively) (Figures 1B, 1C). The
rDNA FISH signals observed in young mice hepatocytes were
always condensed (Figure 1A). In contrast, the 45S rDNA sites
observed in hepatocytes from adult (Figure 1B) and old
(Figure 1C) mice were partially decondensed and appeared as
expanded clouds; they were always seen in the periphery of the
negative images of the n (nucleoli), and in association with
DAPI-stained dense clusters (chromocentres) (Figures 1B, 1C).
These variations can give us a clue of a different chromatin
organization of rDNA sequences when different ages were
compared and are probably a consequence either of a different
packaging of rDNA sequences at different ages or a result of the
different ploidy levels in young, adult and old mice. In the case
of adult and old animals, amplification of the signal could be the
result of increased numbers of rDNA copies in the polyploid
genome.
In fixed and lysed hepatocytes from young mice, which do
not properly form ECFs but uncommon nuclear halos instead,
the small dots of 45S rDNA detected by FISH remained inside the
nuclei after the lysis treatment (Figure 2A), whereas in fixed and
lysed preparations from adult and old mice, the FISH signals
spread throughout and outside the nuclei, such that many small
spots could be seen in the nuclear halos and ECFs (Figures 2B,
2C). The 45S rDNA signal intensity outside the nuclei was higher in
old mice when compared with adult mice (Figures 2B, 2C)
(Table 1). Moreover, the intensity of FISH signals inside the nuclei
was higher in adult hepatocytes than in old hepatocytes (Table 1),
thus arguing in favour of a higher number of rDNA sequences
attached to the nuclear matrix in nuclei from adult mice, when
compared with nuclei from old mice. These results were observed
in all the examined nuclei that formed ECFs.
Figure 1 DNA FISH of 45S rDNA on fixed hepatocyte nuclei from young (A), adult (B) and old (C) miceNuclei were counterstained with DAPI. FISH signals are seen around the nucleoli (n). Scale bars, 12 mm.
Cell Biol. Int. (2010) 34, 925–931
E The Author(s) Journal compilation E 2010 Portland Press Limited Volume 34 (9) N pages 925–931 N www.cellbiolint.org 927

3.2. Telomeric sequences are always located inchromatin loops
The behaviour of telomeric repetitive motifs, known to be hetero-
chromatic and non-genic was also observed, in order to test the
hypothesis that only transcribed sequences remain attached to the
nuclear matrix, after the lysis treatment here employed.
We observed that, in mouse hepatocyte nuclei, telomeres were
also detected as small dots (young and adult mice) (Figures 3A,
3C). In adult (Figure 3C) and old mice (not shown), telomeric
sequences were seen spread in all the analysed nuclei; due to the
increased extent of ploidy, several blocks could be seen, most of
them near the bright DAPI-stained chromocentres but not
associated with them. In young mice, these sequences were
Figure 2 45S rDNA differently associates with the nuclear matrix in hepatocytes of different agesrDNA FISH signals on fixed and vertically lysed hepatocyte nuclei from young (A), adult (B) and old (C) mice. Nuclei were counterstained with DAPI. Arrowsindicate direction of gravity and of ECFs. Scale bars, 30 mm.
Table 1 Image analysis of rDNA FISH signals after lysis treatment%IGLin, percentual of integrated gray levels of FISH signals inside nuclear remnants; %IGLout, percentual of integrated gray levels of FISH signalsoutside nuclear remnants (extended chromatin fibres and nuclear halos); in/out, proportion between integrated gray levels of FISH signals inside andoutside nuclear remnants; M, median.
Model Specimen %IGLin %IGLout In/out M
Adult 1 31.60 68.40 0.46 0.46*2 40.57 59.43 0.683 33.17 66.83 0.504 9.83 90.17 0.115 21.13 78.87 0.27
Old 1 7.04 92.96 0.08 0.10*2 9.40 90.60 0.103 5.77 94.23 0.064 3.72 96.28 0.045 11.68 88.32 0.136 9.09 90.91 0.107 18.43 81.57 0.238 8.69 91.31 0.10
* Statistical difference with P50.0104.
Aging and DNA/nuclear matrix
928 www.cellbiolint.org N Volume 34 (9) N pages 925–931 E The Author(s) Journal compilation E 2010 Portland Press Limited

detected only in nuclei of e (erythroblasts) (Figure 3A). In these
cells, most of the signals appeared positioned as organized
features, concentrated in one part of the cell.
After lysis treatment, the telomeric sequences extended
outside the nuclei at all the ages studied, but with less intensity
in nuclei from young mice (Figure 3B), where they appeared at the
nuclear halos. In this case, a DNA mass distributed homogenously
out and inside the nuclei could be clearly observed. A similar
pattern of telomeric sequences inside the nuclei and spreading
out of them was observed in adult mice (Figure 3D). However, the
intensity of the signals in the halo and in the ECFs of the adult
mice was significantly brighter than those observed in the nuclei of
young mice. Moreover, some dense blocks were clearly seen in
the halo, a phenomenon not observed in nuclei of young mice. The
telomeric sequences that remained in the nuclei were seen also on
the bases of the DNA loop structures located at the nuclear
periphery, which is in accordance with the notion that telomeres
are often associated with the nuclear envelope (Pandita et al.,
2007).
4. Discussion
The 45S rDNA repeats, which are responsible for the production of
a large amount of rRNAs (Wellauer et al., 1974; Perry, 1976;
Kominami et al., 1978), were expected to differently bind to the
nuclear matrix with development and aging, since decreased
transcription levels have been reported under these conditions
(Ma and Nagata, 1990). Indeed, the present results indicate that,
as the animal ages, there is an increase in the amount of 45S rDNA
Figure 3 DNA FISH of telomeric DNA on fixed non-lysed (A, C) and fixed and vertically lysed (B, D) hepatocytes nuclei from young (A, B) and adult (C, D)miceRegardless of age, telomeres do not associate with the nuclear matrix. Nuclei were counterstained with DAPI. e, erythroblast; h, hepatocyte. The arrowindicates direction of gravity and the ECF flow. Scale bars, 12 mm.
Cell Biol. Int. (2010) 34, 925–931
E The Author(s) Journal compilation E 2010 Portland Press Limited Volume 34 (9) N pages 925–931 N www.cellbiolint.org 929

repeats non-associated with the nuclear matrix and located in the
nuclear halos or ECFs, thus suggesting that the attachment of
these sequences to the nuclear matrix is related to gene
activation.
According to the model proposed by Heng et al. (2004), there
are two types of MAR sequences with ability to bind to the nuclear
matrix: (i) the constitutive MARs, which are found in the bases of
the DNA loops and outside transcribed sequences, presenting a
structural role; (ii) facultative MARs, which are located inside the
genes, being responsible for their transcriptional activity. Since
the MARs found until now on rDNA sequences are located only in
flanking sequences, it seems that they are of the constitutive type
(Chen et al., 1997). In fact, there is no description of facultative
MARs on rDNA genes.
In this regard, our present results confirm the existence of
MARs in rRNA genes, since many rDNA copies remained attached
to the nuclear matrix even after the lysis treatment. The
association of MARs with the nuclear matrix is salt-resistant
(Cook et al., 1976; Moraes et al., 2007).
Additionally, the results regarding the 45S rDNA genes suggest
that, in multicopy genes, there is a nuclear matrix-dependent
control of gene expression different from that of single copy
genes. In single copy genes, the adhesion to the nuclear matrix
determines their transcriptional level (Allen et al., 2000; Whitelaw
et al., 2000), while in the case of 45S rDNA, this interaction may
determine if each gene copy will be transcribed or not. Thus, the
amount of transcripts produced could be regulated by controlling
the production average of the thousands of gene copies rather
than by controlling the level of activity of each single repeating
unit.
In relation to the telomeric sequences, the results clearly show
that the association of the chromatin with the nuclear matrix is a
feature common to transcribing sequences. After the lysis
treatment, most telomeric sequences exit the nuclei, remaining
attached only to the nuclear envelope, and it is independent of the
animal developmental state or age. Since telomeric repetitive
motifs are devoid of genes, it is reasonable to conclude that non-
transcribing sequences do not associate with the nuclear matrix.
However, a question still remains on whether other heterochro-
matic sequences such as pericentric repeats have a similar
behaviour.
It is probable that the telomeric sequences in nuclei from
young mice are so packed that they were unavailable to telomeric
probes in fixed nuclei or that, in adult and old mice, there is
unpackaging of the telomeric structure, either as a natural
physiological event or as a by-product of the genomic instability
that is characteristic of the aging process. Nevertheless, after
lysis, which is responsible for solubilizing several histone and non-
histone proteins, the telomeres became accessible to the
telomeric probes, thus binding them and revealing the sequences
located at the nuclear halos. However, it is not to be overlooked
that the telomeric repetitive motifs were seen in control nuclei only
of polyploid adult and old hepatocytes and not in diploid nuclei
from young mice, where there are less telomeric repeats present
and, due to the sensitivity of the method, these sequences were
detectable only when stretched with the lysis treatment.
In the present study, we showed that, when 45S rDNA genes
are being highly transcribed, as in the case of young mice (Ma and
Nagata, 1990), all of these genes were found completely attached
to the nuclear matrix, while with development and aging, their
copies are progressively switched off and consequently detached
from the nuclear matrix. At the same time, the non-coding
telomeric sequences were found detached from the nuclear
matrix irrespective of the age or developmental state of the
animal, but associated with the nuclear envelope, thus supporting
the idea that non-coding regions probably do not associate with
the nuclear matrix.
Author contribution
Alberto Moraes carried out FISH experiments, microscopy, image
processing, statistics and drafted the manuscript. Mateus Mondin
and Margarida Aguiar-Perecin carried out FISH experiments and
drafted the manuscript. Marcelo Beletti carried out image
processing and image analysis and drafted the manuscript. Ana
Guaraldo provided animal facilities and carried out bioterism
services. Maria Luiza Mello co-ordinated the study and drafted the
manuscript.
Acknowledgements
We are grateful to Dr Nick Gilbert from the Cancer Research
Centre (Edinburgh, U.K.) for the critical reading of the manuscript
prior to submission.
Funding
This study was part of a thesis presented by A.M. to the Institute of
Biology, UNICAMP, in partial fulfillment of the requirements for a
Ph.D. degree and was supported by grants from the Brazilian
National Council for Research and Development (CNPq, grant
number. 470587/03-2), the Sao Paulo State Research Foundation
(FAPESP, grant number 2006/00066-8) and Brazilian Research
and Teaching Foundation (CAPES). A.M. and M.L.M. received
Ph.D. and research fellowships respectively, from CNPq. M.M.
received a Pro-Doc fellowship from CAPES.
ReferencesAllen GC, Spiker S, Thompson WF. Use of matrix attachment regions MARs
to minimize transgene silencing. Plant Mol Biol 2000;43:361–76.
Barboro P, D’Arrigo C, Repaci E, Bagnasco L, Orecchia P, Carnemolla B,Patrone E, Balbi C. Proteomic analysis of the nuclear matrix in theearly stages of rat liver carcinogenesis: identification of differentiallyexpressed and MAR-binding proteins. Exp Cell Res 2009;315:226–39.
Berezney R. The nuclear matrix: a heuristic model for investigatinggenomic organization and function in the cell nucleus. J CellBiochem 1991;47:109–23.
Berezney R, Coffey DS. Identification of a nuclear protein matrix.Biochem Biophys Res Commun 1974;60:1410–7.
Berezney R, Coffey DS. Nuclear matrix. Isolation and characterization ofa framework structure from rat liver nuclei. J Cell Biol 1977;73:616–37.
Aging and DNA/nuclear matrix
930 www.cellbiolint.org N Volume 34 (9) N pages 925–931 E The Author(s) Journal compilation E 2010 Portland Press Limited

Burzynski SR. Aging: gene silencing or gene activation?Med Hypotheses 2005;64:201–8.
Cao SX, Dhahbi JM, Mote PL, Spindler SR. Genomic profiling of short-and long-term caloric restriction effects in the liver of aging mice.Proc Natl Acad Sci USA 2001;98:10630–5.
Chen Y, Zhang B, Li X, Zhai Z. Association of DNA with nuclear matrix inin vitro assembled nuclei induced by rDNA from Tetrahymenashanghaiensis in Xenopus egg extracts. FEBS Lett 1997;413:449–52.
Cook PR, Brazell I, Jost E. Characterization of nuclear structurescontaining superhelical DNA. J Cell Sci 1976;22:303–24.
Cremer T, Kreth G, Koester H, Fink RH, Heintzmann R, Cremer M,Solovei I, Zink D, Cremer C. Chromosome territories,interchromatin domain compartment, and nuclear matrix: anintegrated view of the functional nuclear architecture. Crit RevEukaryot Gene Expr 2000;10:179–212.
Davie JR. The nuclear matrix and the regulation of chromatinorganization and function. Int Rev Cytol 1995;162A:191–250.
Fedoroff NV. On spacers. Cell 1979;16:697–710.Gasser SM, Laemmli UK. Cohabitation of scaffold binding regions with
upstream/enhancer elements of three developmentally regulatedgenes of D. melanogaster. Cell 1986;46:521–30.
Gaubatz J, Prashad N, Cutler RG. Ribosomal RNA gene dosage as afunction of tissue and age for mouse and human. Biochim BiophysActa 1976;418:358–75.
Gerdes MG, Carter KC, Moen PT, Lawrence JB. Dynamic changes in thehigher-level chromatin organization of specific sequences revealedby in situ hybridization to nuclear halos. J Cell Biol 1994;126:289–304.
Gupta S. Hepatic polyploidy and liver growth control. Semin Cancer Biol2000;10:161–71.
Haaf T, Ward DC. Structural analysis of alpha-satellite DNA andcentromere proteins using extended chromatin and chromosomes.Hum Mol Genet 1994;3:697–709.
Harman D. Ageing: phenomena and theories. Ann NY Acad Sci1998;854:1–7.
Heng HH, Goetze S, Ye CJ, Liu G, Stevens JB, Bremer SW, Wykes SM,Bode J, Krawetz SA. Chromatin loops are selectively anchoredusing scaffold/matrix-attachment regions. J Cell Sci2004;117:999–1008.
Kominami R, Hamada H, Fujii-Kuriyama Y, Muramatsu M. 59-Terminalprocessing of ribosomal 28S RNA. Biochemistry 1978;17:3965–70.
Long EO, Dawid IB. Repeated genes in eukaryotes. Annu Rev Biochem1980;49:727–64.
Ma H, Nagata T. Study on RNA synthesis in the liver of aging mice bymeans of electron microscopic radioautography. Cell Mol Biol1990;36:589–600.
Ma HJ, Gao FL, Olea MT, Nagata T. Protein synthesis in the livers ofaging mice studied by electron microscopic radioautography.Cell Mol Biol 1991;37:607–15.
Meunier-Rotival M, Cortadas J, Macaya G, Bernardi G. Isolation andorganization of calf ribosomal DNA. Nucleic Acids Res1979;6:2109–23.
Mondin M, Santos-Serejo JA, Aguiar-Perecin MLR. Karyotypecharacterization of Crotalaria juncea L. (Leguminosae–Papilionoideae) by chromosome banding and in situ hybridizationof rDNA 45S and 5S. Genet Mol Biol 2007;30:65–72.
Moraes AS, de Campos Vidal B, Guaraldo AM, Mello ML. Chromatinsupraorganization and extensibility in mouse hepatocytes followingstarvation and refeeding. Cytometry A 2005;63:94–107.
Moraes AS, Guaraldo AM, Mello ML. Chromatin supraorganization andextensibility in mouse hepatocytes with development and aging.Cytometry A 2007;71:28–37.
Nickerson J. Experimental observations of a nuclear matrix. J Cell Sci2001;114:463–74.
Oberdoerffer P, Sinclair DA. The role of nuclear architecture in genomicinstability and ageing. Nat Rev Mol Cel Biol 2007;8:692–702.
Pan D, Zhang L. Tandemly arrayed genes in vertebrate genomes.Comp Funct Genomics 2008;545269.
Pandita TK, Hunt CR, Sharma GG, Yang Q. Regulation of telomeremovement by telomere chromatin structure. Cell Mol Life Sci2007;64:131–8.
Pardoll DM, Vogelstein B. Sequence analysis of nuclear matrixassociated DNA from rat liver. Exp Cell Res 1980;128:466–70.
Perry RP. Processing of RNA. Annu Rev Biochem 1976;45:605–29.
Tsutsui KM, Sano K, Tsutsui K. Dynamic view of the nuclear matrix.Acta Med Okayama 2005;59:113–20.
Vidal BC. Extended chromatin fibres: crystallinity, molecular order andreactivity to concanavalin-A. Cell Biol Int 2000;24:723–8.
Wellauer PK, Reeder RH, Carroll D, Brown DD, Deutch A,Higashinakagawa T, Dawid IB. Amplified ribosomal DNA fromXenopus laevis has heterogeneous spacer lengths. Proc Natl AcadSci USA 1974;71:2823–7.
Whitelaw CBA, Grolli S, Accornero P, Donofrio G, Farini E, Webster J.Matrix attachment region regulates basal beta-lactoglobulintransgene expression. Gene 2000;244:73–80.
Received 8 December 2009/ 6 May 2010; accepted 2 June 2010
Published as Immediate Publication 2 June 2010, doi 10.1042/CBI20090457
Cell Biol. Int. (2010) 34, 925–931
E The Author(s) Journal compilation E 2010 Portland Press Limited Volume 34 (9) N pages 925–931 N www.cellbiolint.org 931