afferent fibres in the hypoglossal nerve of cat
TRANSCRIPT

Acta physiol. scand. 1970. 79. 24-36 From the Department of Clinical Neurophysiology, Karolinska sjukhuset, Stockholm, Sweden
Afferent Fibres in the Hypoglossal Nerve of Cat BY
JERKER HANSON and LENNART W I D ~ N
Received 1 7 July 1969
Abstract
HANSON, J. and L. W I D ~ N . Afferent fibres in thr hypoglossal n m w of cat. Acta physiol. scand. 1970. 79. 24-36.
Electrical stimulation of the hypoglossal nerve in cat elicited bilateral twitching of the vibrissae and suppression of shivering. The afferent fibres which mediate these reflexes have a higher stimulus threshold than the motor fibres. They travel to the brain stem via anastomoses be- tween the hypoglossal nerve and the vagus group in the region of the ganglion nodosum. Unitary activity was recorded in the spinal trigeminal nucleus after stimulation of the hypo- glossal nerve. The reflexes are probably nociceptive reactions since similar reflexes can be evoked from the greater part of the body surface by pain stimuli. Stimulation of a peripheral branch of the hypoglossal nerve also elicited a local reflex response in the styloglossus muscle. The reflex is polysynaptic. Any tonic background activity in the muscle is inhibited by the stimulation. I t is shown that this inhibition is probably not recurrent but mediated by afferent fibres via anastomoses to the vagus group. Afferent impulses evoked by stretching the tongue were recorded in the proximal part of the hypoglossal nerve trunk. These units behaved like muscle spindles. In cat these units are sparse and probably of secondary importance for move- ments of the tongue.
The question of whether the hypoglossal nerve consists of motor fibres only 0 1 also contains sensory fibres is disputed. Among those who ascribe the nerve sensory fibres opinions differ as to whether these travel with the nerve into the medulla oblongata or via anastomoses cross over into other nerves. Conditions appear to differ in dif- ferent animal species but the results of investigations in the same animal species are also conflicting. I n the following brief survey stress is laid mainly on investigations in cat. A more comprehensive review has been published by Blom ( 1960).
Anatomical studirs A root ganglion upon the hypoglossal nerve has been described in cat by Froriep and Beck (1895), Langworthy (1924 a, b ) and Holliger (1955) although is was not en- countered in all animals examined. Tarkhan and Abd-el-Malek (1951) found sensory nerve cells along the nerve trunk, mainly in the distal half; they were usually scattered and very few in number, in one cat, however, a ganglion with 200 cells was found.
Connections between the trunk of the hypoglossal nerve and cervical ganglia have been found in some mammals but not with certainty in the cat (Hinsey and Corbin
24

HYPOGLOSSAL N E R V E AFFERENTS 25
1934, Corbin and Harrison 1938, Downman 1939). On the other hand, peripheral anastomoses have been clearly demonstrated between the hypoglossal and lingual nerves (Fitzgerald and Law 1958, Blom 1960), lying on the surface of the styloglossus muscle. Lingual nerve fibres have not been described in the more proximal parts of the hypoglossal nerve.
According to textbooks of anatomy (e .6 . Clara 1959) there exists in man a connec- tion between the vagus groups and the hypoglossal nerve where these nerves lie in proximity to each other at the level of, and proximal to the ganglion nodosum. ‘These connections are assumed to contain autonomic fibres. It is uncertain whether these accompany the hypoglossal nerve peripherally to the tongue muscles. Boyd (1941) was unable to demonstrate this anastomosis in rabbit. O n the other hand, Tarkhan and Abou-el-Naga (1947) found in dog a small number of degenerating fibres in the hypoglossal nerve after removal of the ganglion nodosum. We have been unable to find any information regarding conditions in the cat.
Physiological studips Several attempts have been made to record afferent impulses in the proximal part of the hypoglossal nerve trunk during various types of stimulation to the cat’s tongue (Barron 1936, Corbin and Harrison 1938, Downman 1939, Blom 1960, Porter 1966) but with negative results. Cooper (1954) and Blom (1960), however, found impulse activity in the peripheral end branches of the nerve on stretching the tongue but along which path these impulses are propagated to the brain stem has not been clarified. Neither Cooper (1953), Law (1954) nor Blom (1960) were able to demon- strate muscle spindles in the intrinsic muscles of the cat’s tongue and it is therefore unknown what type of stretch receptor is activated.
With intracellular recording from the hypoglossal motor neurones Green and Negishi (1963) and Porter (1965, 1966) found in addition to antidromic action potentials also postsynaptic potentials in response to stimulation of the central cut end of the nerve. Green and Negishi considered these indicated the presence of afferent fibres in the hypoglossal nerve, whereas Porter presumed that they were the result of stimulus spread to the lingual nerve.
Other indices of afferent fibres in the hypoglossal nerve of cat have been reported. Electrical stimulation of the central cut end of the nerve caused rise of blood pressure (Tarkhan 1936) and dilatation of the pupils (Downman 1939). Downman left i t an open question whether these reflexes are transmitted via the hypoglossal nerve or the vagus-accessory group but Tarkhan and Abou-el-Naga (1947) who, however, at the time were working with dogs, maintained that the afferent fibres reach the brain stem via the vagtis nerve. Recently Sauerland and Mizuno (1968) described a hypo- glosso-laryngeal reflex mediated by fibres which at the ganglion nodosum pass ovw into the vagus nerve and travel with this to the brain stem.
The present electrophysiological studies show that the hypoglossal nerve contains afferent, probably small myelinated fibres which mediate reflex responses not earlier demonstrated on stimulation of this nerve. The fibres which mediate these effects

26 JERKER HANSON A N D LENNART WIDBN have the same course as those described by Sauerland and Mizuno. Further, it is shown that afferent impulses elicited by stretching the tongue can by recorded in the proximal part of the hypoglossal nerve.
Methods Experiments were performed on 44 adult cats of either sex. Most of the animals were under light pentobarbitone (Nembutal, Abbott) anesthesia during the experiments. I n a few instances chloralosed cats were used or cats decerebrated, under pentobarbitone anesthesia, by section of the brain stem a t the midcollicular level, after which further anesthetic was not given.
Chlorided silver electrodes were used for stimulating and recording from the peripheral nerves, steel needle electrodes for the electromyogram. Microelectrodes of stainless steel, made according to the method described by Green ( 1958) were used for recording from single units in the brain stem. During these recordings the rat’s head was firmly fixed in a stereotaxic instrument (Baltimore Instrument Co.) and a large occipital decompression was made in order to decrease the pulsations. The stei eotaxic coordinates for recording from the spinal trigeminal nucleus were calculated according to Snider’s and Nienier’s atlas ( 196 1 ) ,
Electrical stimuli were delivered from a Grass Model S 8 stimulator with isolation units. Unless otherwise stated the stimuli consisted of single shocks of 0.2 msec duration. The action potentials were led to an amplifier (Grass Model P 9) connected to a loudspeaker and when necessary combined with a cathode follower. The responses were displayed on an oscilloscope (Tektronix Model 502) and photographed with a Grass camera. In some experiments a tape recorder (Epsilon Labcorder 0-5000 Hz) was employed and photographs taken during play back.
Blood pressure, respiration and shivering were recorded on a Grass polygraph, shivering in addition on the oscilloscope. The transducers used were: for blood pressure a Statham strain gauge manometer, for respiration a thermistor attached close to the opening of the tracheal cannula, and for shivering a capacitive transducer (Dickinson 1950, p. 111-116) which recorded movements of the foot. I n some animals the EMG was recorded with needle elec- trodes inserted into the m. quadriceps femoris.
The site of the microelectrode was marked with the Prussian blue method. The brains were perfused in situ with a mixture of potassium ferrocyanide and formalin and serially sectioned after removal, further fixation in formalin and imbedding in paraffin. The sections were stained with toluidine blue for microscopic investigation.
Results I. Reflexes evoked by stimulation of the hypoglossal nerve Stimulation of the hypoglossal nerve elicited various reflex effects which can be divided into the following groups:
A. Effects in reflex systems not directly related t o the function of the tonguP 1. Bilateral movement of the vibrissae During dissection of structures in the neck of the cat it was observed that pinching or section of the hypoglossal nerve produced twitching of the vibrissae. This movement could also be elicited by electrical stimulation of the hypoglossal nerve (Fig. 1 A) and was found to be a reflex discharge in m. quadratus labii superioris. At stimulus strengths well above threshold for the reflex, the latency showed 10-15 msec varia- tions in any one cat, and still larger variations between different cats. However, the latency was never shorter than 20 msec or longer than 55 msec (mean approx. 35 msec). At a stimulus frequency of ,<l/sec the duration of the response varied be- tween 10 and 60 msec depending, among other factors, on the depth of anesthesia and the excitability of the preparation. The response was usually bilateral. generally with a slightly lower stimulus threshold in the contralateral intact muscle.

HYPOGLOSSAL NERVE AFFERENTS 27
A
zoopv I 20 m u c . T
B
T Fig. 1. A , Reflex response in m. quadratus labii superioris to electrical stimulation (ar- row) of the central cut end of the con- tralateral hypoglossal nerve. B, Same re-
close to the brain stem. C , Intracranial sec- sponse after intracranial section of the nerve C tion of the roots of the vagus group abol- ished the response; stimulus strength in- creased to ensure that no responsccould be elicited. T
Afferent reflex arc. In order to ascertain whether the impulses were transmitted to the brain stem via the hypoglossal nerve or via anastomoses between this nerve and some other nerve, the hypoglossal nerve was sectioned intracranially at its exit from the brain stem. As shown in Fig. 1 B, this had no effert on the response. Likewise ipsilateral section of the trigeminal nerve intracranially in Meckel’s cavity, section of the C1 to C4 roots or section of the spinal root of the IX nerve had no influence on the response. On the other hand, section of the I X and X nerves intracranially, thus proximal to the ganglia of these nerves, abolished the response (Fig. 1 C ) .
In order to locate the connections found between the hypoglossal nerve and the vagus and glossopharyngeal nerves, the hypoglossal nerve was dissected free from the periphery in a central direction, dissection being interrupted at intervals to stimulate the nerve immediately distal to its point of intersection with the hyoid bone. Dissec- tion of the hypoglossal nerve at its intersection with the vagus nerve and the gan- glion nodosum regularly abolished the reflex response. In other experiments it was found that section of the hypoglossal nerve distal to its intersection with the vagus nerve, i.e. at its exit from the bony canal, had no effect on the reflex response. From this it is concluded that afferent fibres travel in the peripheral part of the hypoglossal nerve from its exit from the intrinsic tongue muscles and thereafter join the vagus group at the level of the ganglion nodosum or somewhat proximal to it. At the site of intersection the nerves are surrounded by a good deal of connective tissue and it is impossible to see the anastomoses with the naked eye, neither was it possible to identify them with certainty under the dissection microscope.

28 JERKER HANSON AND LENNART W I D B N
- Fig. 2 Fig. 3
t Fig. 2. Unitary response to single shock stimulation of the central cut end of the ipsilateral hypoglossal nerve, recorded in the spinal trigeminal nucleus.
Fig. 3 . Stimulating and rerording from the hypoglossal nerve with the stimulating cathode at a distance of 25 mm from the recording electlode. The stimulus strength was in A , just liminal for the reflex response in m. quadratus labii superioris, in R, twire the threshold strength.
That spread of stimulus from the hypoglossal nerve to the vagus group played no part appears evident because, as mentioned earlier, the reflex response could be elicited by mechanical stimulation of the hypoglossal nerve. Control experiments also showed that compression of the hypoglossal nerve with forceps proximal to the stiniu- lating cathode abolished the response.
Smsory relay nucleus. The facial reflex can be elicited in decerebrated aniiiials. Accordingly it is a brain stem reflex. We have made no systematic attempts to map out the path of the impulses in the brain stem from their entrance with the vagiis group to the facial nucleus. Since the afferent fibres are probably relatively small and perhaps mediate pain impulses also (see below), it seemed natural to invrstigate whether they transmit impulses to the spinal trigeminal nucleus, which is assumed to be the terminal for pain impulses not only from the V, but also from the VII, IX and X cranial nerves (Brodal 1947).
I n a few preliniinary experiments it was found that ipsilateral section of the spinal trac,t of the trigeminal nerve abolished the reflex movement. Microelectrode recording from the area of the spinal trigeminal nucleus was performed in other experinirnts. A characteristic repetitive unitary response to single shock stimulation of the proximal cut end of the hypoglossal nerve is shown in Fig. 2. The repetitive discharge suggests that this is a postsynaptic response. The latency to the first discharge varied between 10 and 15 insec in different experiments. The latency does not of course give any definite information regarding the conduction rate in the afferent fibres since the number of synaptic relays, if any, preceding the response is unknown.
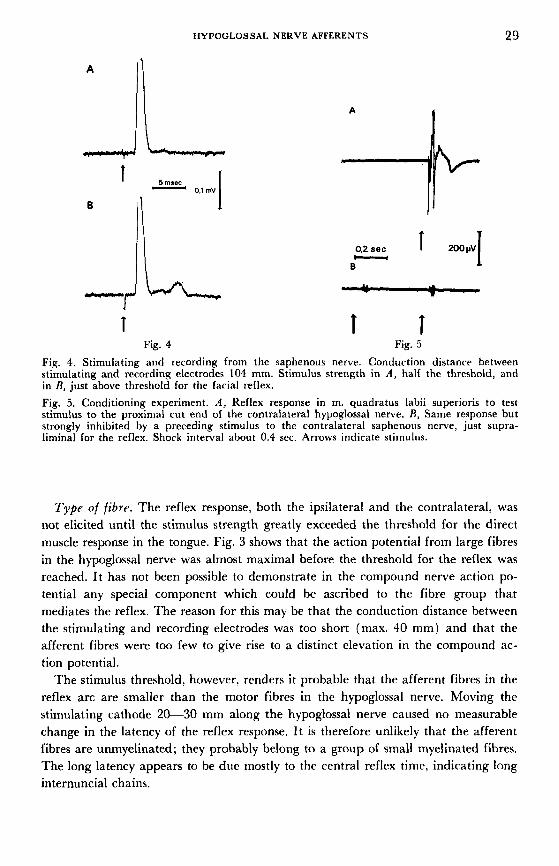
HYPOGLOSSAL NERVE AFFERENTS 29
A
-r 1 L, Fig. 4
A
T T Fig. 5
Fig. 4. Stimulating and recording from the saphenous nerve. Conduction distance between stimulating and recording electrodes 104 mm. Stimulus strength in A, half the threshold, and in R, just above threshold for the facial reflex. Fig. 5. Conditioning experiment. A, Reflex response in m. quadratus labii superioris to test stimulus to the proximal cut end of the contralateral hypoglossal nerve. R, Same response but strongly inhibited by a preceding stimulus to the contralateral saphenous nerve, just supra- liminal for the reflex. Shock interval about 0.4 sec. Arrows indicate stimulus.
Type of fibre. The reflex response, both the ipsilateral and the contralateral, was not elicited until the stimulus strength greatly exceeded the threshold for the direct muscle response in the tongue. Fig. 3 shows that the action potential from large fibres in the hypoglossal nerve was almost maximal before the threshold for the reflex was reached. It has not been possible to demonstrate in the compound nerve action po- tential any special component which could be ascribed to the fibre group that mediates the reflex. The reason for this may be that the conduction distance between the stimulating and recording electrodes was too short (max. 40 mm) and that the afferent fibres were too few to give rise to a distinct elevation in the compound ac- tion potential.
The stimulus threshold, however, renders it probable that the afferent fibres in the reflex arc are smaller than the motor fibres in the hypoglossal nerve. Moving the stimulating cathode 20-30 mm along the hypoglossal nerve caused no measurable change in the latency of the reflex response. It is therefore unlikely that the afferent fibres are unmyelinated; they probably belong to a group of small myelinated fibres. The long latency appears to be due mostly to the central reflex time, indicating long internuncial chains.

30 JERKER HANSON AND LENNART W I D ~ N
1 1 sec
Fig. 6. Inhibition of shivering in the ipsilateral hindlimb induced by rcpetitive stimulation (15 stimuli) of the proximal cut end of the hypoglossal nerve. Arrows indicate beginning and end of stimulation.
It was found that a similar bilateral reflex movement of the cat’s vibrissae, duc. to contraction of mm. quadrat; labii superioris, could be elicited with pain stimuli, such as pinching the skin with forceps or pricking with a needle, from a large receptive area comprising in excitable animals practically the cntire body surface, in less excit- able animals restricted to the head. Touch or pressure stimulation of the skin or passive movements of the limbs never evoked the reflex response.
Electrical stimulation of the saphenous nerve in the hind leg evoked a bilateral reflex response in m. quadratus labii superioris with a latency about 15 msec longer than that evoked by stimulation of the hypoglossal nerve in the same animal. Simul- taneous recording of the compound action potential from the saphenous nerve in these experiments (Fig. 4 ) shows that the threshold for the reflex response was not reached until the stimulus strength produced an almost maximal AP-spike and a well- developed deflection corresponding to an impulse wave in a fibre group whose fastest fibres had a conduction rate of about 15 m/sec, i.e. belonged to the delta group.
In order to determine whether the impulses from the saphenous nerve arc mediated to the facial nucleus via the same pathway as the impulses from the hypo- glossal nerve, a conditioning experiment of the type illustrated in Fig. 5 was donr. When a stimulus to the saphenous nerve, just supraliminal for the reflex response. preceded stimulation of the hypoglossal nerve, the reflex response to the latter was reduced by more than 95 % during a stimulus interval of up to about half a second. From this and similar experiments it is concluded that the impulses from the saphenous and hypoglossal nerves before reaching the facial nucleus converge on the same postsynaptic structures somewhere in the brain stem and that the reflex arc is partly the same in both cases.
2. Suppression of shivering In cats with spontaneous “shivering” repetitive shock stimuli to the hypoglossal nerve induced suppression of shivering after a latency of almost one second (Fig. 6 ) . The cffect disappears after intracranial section of the vagus group.
Similar suppression of shivering can be obtained with pain stimuli (pinching the skin with forceps) to the limbs or trunk but not by touch, pressure or passive move- ments of the limbs.

HYPOQLOSSAL NERVE AFFERENTS 31
Fig. 7. Reflex response in the sty- loglossus muscle evoked by a stimulus to the proximal cut end of a branch of the ipsilateral hypoglossal nerve. T lOmsec
3. Effect on blood pressure and pupils In cats under pentobarbitone anesthesia and in cats decerebrated under anesthesia stimulation of the central cut end of the hypoglossal nerve produced no unequivocal effects on the blood pressure. The pupils were unaffected. In cats under chloralose anesthesia, however, a rise in blood pressure (about 20 mm Hg) and pupillary dilata- tion were regularly obtained, which is in accordance with the findings of Downman (1939).
B. Local reflexes related to the function of the tongue 1. A hypoglosso-hypoglossal reflex in the m . styloglossus Electrical stimulation of the central cut end of a peripheral branch of the hypoglossal nerve elicited a reflex response in the styloglossus muscle after a latency of about 15 msec (Fig. 7 ) . The threshold for this reflex response, which is abolished by intra- cranial section of the vagus group, is about half the threshold for the reflex response from m. quadratus labii superioris.
The styloglossus muscle is supplied by the hypoglossal nerve ( Barnard 1940, Clara 1959, p. 344), a finding we have confirmed by demonstrating that all activity in the muscle is abolished after section of the hypoglossal nerve at its point of exit from the brain stem. Accordingly, the reflex represents a hypoglosso-hypoglossal reflex.
2. Inhibition of tonic activity in extrinsic muscles of the tongue The stimulation of the hypoglossal nerve described in the foregoing induced, in addi- tion, suppression of the tonic activity readily elicited in the extrinsic muscles of the tongue by holding the cat’s mouth wide open (Fig. 8 A ) . The suppression occurs after a latency of 10-15 msec and lasts about half a second. Blom (1960), who observed this suppression, ascribed it to recurrent inhibition. The suppression was abolished by section of the vagus group of nerves at their point of entrance into the brain stem (Fig. 8 B), which shows that it is mediated by fibres which cross over from the hypo- glossal nerve to the vagus group.
The threshold for the suppression is about the same as the threshold for the reflex response in m. styloglossus but the suppression can, however, sometimes be induced without the reflex response and accordingly is not secondary to it.

32 A
JERKER HANSON A N D LENNAKT W I D B N
Fig. 8. Electromyographic recording of tonic activity in the styloglossus muscle evoked by holding the rat’s mouth open. In A , electrical stimulation of the proximal cut end of a branch of the ipsilateral hypoglossal nerve inhibits this ,activity. E , After intracranial section of the roots of the vagus group no inhibition is ob- tained.
11. Impulse actiuity in the hypoglossal afferent n e r w fibre3 Several attempts were made to record afferent impulses in filaments of the hypo- glossal nerve during application of various types of stimuli to the tongue, e.g. touch. pressure, stretching, twisting, pinching, heat and cold. The filaments werr dissected free at the junction of the hypoglossal nerve and the carotid artery, or more proximal, in order to avoid involvement of lingual nerve anastomoses. I t was difficult to ob- tain satisfactory recording conditions since stretching of the tongue readily caused movements in the floor of the mouth which cannot be immobilized. Movements be- tween filaments and recording electrodes with the risk of damaging vulnerable slender fibres could not always be avoided. The fact that the majority of the fibres in this nerve are efferents rendered, a priori, the chances of finding a functioning filament small. Moreover, the larger fibres could be assumed to shunt and thereby conceal afferent activity in isolated small fibres even in slender filaments.
Sporadic discharges or brief bursts of single action potentials were recorded in these experiments. They appeared only during complicated mechanical stimulations with severe deformation of the tongue and therefore could not be related to any special type of stimulus, or else they were evoked only by such strong compression of the tongue with forceps that injury discharges from intraglossal nerve fibres could he suspected.
I n a total of over 300 filaments examined, only 10-15 tonically active units were encountered whose discharge frequency increased on stretching the tongue straight forward or drawing the tip of the tongue out of the mouth and to one side. I n the experiment illustrated in Fig. 9, stretching of the tongue doubled the discharge fre- quency of such a unit. Release of the tongue induced at first complete silence of thr

HYPOGLOSSAL NERVE AFFERENTS 33
Fig. 9. Unit activity in a tion of the nerve and the of the lower line.
filament from the hypoglossal nerve, 3 mm proximal to the intersec- carotid artery. Stretching of the tongue is indicated by upward shift
receptor for about one-third of a second, followed by tonic activity of relatively low frequency. Injection of 15-20 ,ug/kg of succinylcholine iodide into the external carotid artery in this and another experiment produced a strong increase (>200 ”/o) of the impulse activity, without stretching of the tongue.
Active contraction of the intrinsic tongue muscles, produced by electrical stimula- tion through needle electrodes inserted into the tongue, induced a “silent period” (Fig. 10 A). On the other hand, contraction of the muscles in the floor of the mouth caused no receptor silence (Fig. 10 B). If the receptor was a stretch receptor, which its behaviour indicates, it seems highly probable that it was situated in some intrinsic muscle and was un!oaded on its shortening.
Discussion
The present investigation has shown that the hypoglossal nerve in cat contains af- ferent fibres which, in the vicinity of the ganglion nodosum, unite with the nerves of the vagus group. In addition, impulse activity from stretch sensitive receptors has been demonstrated in the proximal part of the hypoglossal nerve.
The factors which establish the presence of afferent fibres in the hypoglossal nerve and which have been demonstrated in this study can be divided into three groups, which we consider can most suitably be discussed separately.
1. The facial reflex, suppression of shivering and effects on blood pressure were all induced a t approximately the same stimulus strength and were mediated via anasto-
3-703003. A614 p h p i o l . rcand. Vol. 79: 1

34 JERKER HANSON AND LENNART W I D ~ N
0.1 ... T . B
I
Fig. 10. Same experiment as in Fig. 9. A, The receptor becomes silent during contraction of
I I I I I I I l l I I I I I11 I I. I I I I I I I I I I I I I intrinsic muscles, produced -by a stimulus through needle electrodes inserted in the tongue. R, A stimulus applied to the floor of the mouth and rausing contraction of extrinsic
9 t tongue muscles has no effect on receptor ac-
tivity.
moses between the hypoglossal and vagus group nerves. The facial reflex did not appear until the A-spike of the hypoglossal nerve was almost maximal (Fig. 3 ) and thus was evoked from small fibres, probably not C fibres however, but apparently fibres of the same size as those which mediated the reflex response elicited by stimu- lation of the saphenous nerve, i.e. delta fibres. A similar facial reflex could be elicited by pain stimuli to different parts of the head, trunk and limbs. The reflex is pre- sumably identical with the “movement of the vibrissae” described by Woodworth and Sherrington (1904) and which they considered one of the most discrete and easily elicited components of a nociceptive reaction, “the pseudoaffective reflex”, in decerebrate cat. As Reis (1961) points out, the facial reflex has similarities with the palmomental reflex in man, which he considers represents “a low threshold plateau of a generalized somatic response to nociceptive stimuli”. Of the natural stimuli, only pain induced suppression of shivering. I t is therefore highly possible that the afferent fibres in the hypoglossal nerve which mediate these reactions are “pain fibres”.
Rise of blood pressure and dilatation of the pupil are generally taken as conven- tional indices of pain reactions in experimental animals (Bishop 1946). In chloralosed animals these reactions were elicited by stimulation of the hypoglossal nerve. Accord- ing to Tarkhan (1936) and Downman (1939) this also applies in urethanized cats and cats decerebrated under ether anesthesia. We were unable to evoke these reac- tions in cats anesthetized with pentobarbitone. Thus, the reactions are not particular- ly strong, which may be due to the fact that the afferent fibres mediating the reac- tions are few in number.
2. The reflex responses in the styloglossus muscle and inhibition of tonic activity in this muscle. The stimulus threshold for these effects was slightly lower than that re- quired to elicit movement of the vibrissae. It is difficult to say whether the reason for this was that the afferent fibres are larger or that it was not necessary to activate an equally large number of fibres to obtain the reflex effects in the styloglossus muscle.
Tarkhan ( 1936) recorded contralateral reflex movements in the tongue elicited by stimulation of the central cut end of the hypoglossal nerve, an observation not confirmed by other workers (see Biom 1960). The hypoglosso-hypoglossal reflex

HYPOGLOSSAL NERVE AFFERENTS 35
demonstrated in this study is obviously not a monosynaptic reflex; the afferent fibres of the reflex arc have a higher stimulus threshold than the efferent. Nothing indicates that it is a myotatic reflex of the kind studied by electrical nerve stimulation in other cat muscles.
The inhibition of tonic activity in the styloglossus muscle, induced by stimulation of a hypoglossal nerve filament from this muscle, was interpreted by Blom (1960), who first described this phenomenon, as a recurrent inhibition. Accordingly, this would indicate the existence of recurrent collaterals in the hypoglossal nucleus, al- though Cajal (1909) was unable to find any. If the inhibition were recurrent it should have the same threshold as the motor fibres, but its threshold is in fact con- siderably higher (c f . also Porter 1965). The fact that intracranial section of the vagus group abolished the inhibition makes i t unlikely that it is recurrent.
3. That pain stimuli to the tongue evoked no definitely identifiable impulse activity in proximal hypoglossal filaments was probably due to the technical reasons men- tioned earlier. In a few instances, however, we succeeded in recording activity elicited by stretching of the tongue (Fig. 9, 10). Cooper (1954) and Blom (1960) recorded activity in distal hypoglossal filaments close to their exit from the intrinsic muscles on stretching of the tongue. O n the other hand, such activity has not earlier been shown in the proximal parts of the nerve and consequently it has not been possible to determiAe whether the impulses reach the brain stem via the hypoglossal or the lingual nerve. Our results show that such impulses travel in the hypoglossal nerve beyond its connection with the lingual nerve but we do not know by which cranial nerve root they enter the brain stem.
The receptors we studied behaved in every way like muscle spindles but can, of course, have been some other type of stretch receptors. The fact that despite long- lasting experiments on several cats with recording from several hundred filaments we were only able to record activity in a total of 10--15 such receptors probably indicates that the afferent fibres from them in the hypoglossal nerve are few in number. This is also in agreement with the anatomical observations of Blom. It has not been pos- sible to demonstrate activity from stretch receptors of this type in the lingual nerve (Blom 1960, cf. also Porter 1966). Therefore Blom’s conclusion that other types of receptors, situated in the submucosa, rapidly adapting and sensitive, among other things, to phasic movements in the tongue muscles, are of greater importance for movements of the cat’s tongue appears to be well founded.
Cooper (1953) found muscle spindles in the intrinsic muscles of the tongue in man and rhesus monkey. To judge from the investigations of Bowman and Combs (1968) in the rhesus monkey, fibres from stretch receptors of the muscle spindle type in the distal half of the hypoglossal nerve are considerably more numerous in these animals than in the cat. The authors assume that in the rhesus monkey the muscle spindles are of vital importance for the proprioception of the tongue and mean that there is no reason to ascribe the touch receptors a special role. Accordingly there appears to be fundamental differences in the anatomical and functional substrata for the proprioception of the tongue in different species of higher mammals.

36 JERKER HANSON AND LENNART W I D ~ N
References BARNARD, J. W., The hypoglossal complex of vertebrates. J. cornp. Neurol. 1940. 72. 489-524. BARRON, D. H., A note on the course of the proprioceptor fibres from the tongue. Anat. Rec .
BISHOP, G. H., Neural mechanisms of cutaneous sense. Physiol. Rev. 1946. 26. 77-102. BLOM, S., Afferent influences on tongue muscle activity. Acta physiol. scand. 1960. 49. Suppl.
BOWMAN, J. P. and C. M. COMBS, Discharge patterns of lingual spindle afferent fibers in the
BOYD, J. D., The sensory component of the hypoglossal nerve in the rabbit. J. Anat . (Lond.)
BRODAL, A., Central course of afferent fibers for pain in facial, glossopharyngeal and vagus
CAJAL, S. R. Y., Histologie d u systdrne nerveux de l‘homme et des vertkbre‘s. I. A. Maloine,
CLARA, M., Das Nervensystem des Menschen. 3rd edition. J. A. Barth Verlag, Leipzig. 1959. COOPER, S., Muscle spindles in the intrinsic muscles of the human tongue. J . Physiol. (Lond.)
COOPER, S., Afferent impulses in the hypoglossal nerve on stretching the cat’s tongue. J . Physiol.
CORBIN, K. B. and F. HARRISON, Further studies on tongue innervation. Proc. SOC. exp. Riol.
DICKINSON, C. J., Electrophysiological Technique. Electronic Engineering, (Lond.) 1950. 141 p. DOWNMAN, C. B. B., Afferent fibres of the hypoglossal nerve. J. Anat. (Lond.) 1939. 73. 387-
FITZGERALD, M. J. T. and M. E. LAW, The peripheral connexions between the lingual and
FRORIEP, A. and W. BECK, Uber das Vorkommen dorsaler Hypoglossuswuneln mit Ganglion, in
1936. 66. 11-15.
170.
hypoglossal nerve of the rhesus monkey. Exp. Neurol. 1968. 21. 105-119.
1941. 75. 330-345.
nerves. Arch. Neurol. Psychiat. (Chic.) 1947. 57. 292-306.
Paris. 1909.
1953. 122. 193-202.
(Lond.) 1954. 126. 32 P.
(N.Y.) 1938. 38. 308-310.
395.
hypoglossal nerves. J. Anat . (Lond.) 1958. 92. 178-187.
der Reihe der Saugetiere. Anat. Anz. 1895. 10. 668-696. GREEN, J. D., A simple microelectrode for recording from the central nervous system. Nature
GREEN, J. D. and K. NEGISHI, Membrane potentials in hypoglossal motoneurons. J . Neuro- (Lond.) 1958. 182. 962.
physiol. 1963. 26. 835-856. HINSEY, J. C. and K. G. CORBIN, Observations on the peripheral course of the sensory fibers in
the first four cervical nerves of the cat. J. cornp. Neurol. 1934. 60. 37-44 . HOLLIOER, A., Contribution B 1’6tude de la constitution de l’anse du nerf hypoglosse; Ctude
d’anatomie comparie. Arch. Anat . Hist. Embr. 1955. 38. 4 - 4 6 . LANOWORTHY, 0. R., A study of the innervation of the tongue musculature with particular
reference to the proprioceptive mechanism. J. cornp. Neurol. 1924a. 36. 273-297. LANOWORTHY, 0. R., Problems of tongue innervation: Course of proprioceptive nerve fibers,
autonomic innervation of skeletal musculature. Bull. Johns Hopkins Hosp. 1924b. 35. 239- 246.
LAW, M. E., Lingual proprioception in pig, dog and cat. Nature (Lond.) 1954. 174. 1107- 1108.
PORTER, R., Synaptic potentials in hypoglossal motoneurones. J . Physiol. (Lond. ) 1965. 180.
PORTER, R., Lingual mechanoreceptors activated by muscle twitch. J. Physiol. (Lond.) 1966.
REIS, D. J., The palmomental reflex. A fragment of a general nociceptive skin reflex: A
SAUERLAND, E. K. and N. MIZUNO, Hypoglossal nerve afferents: elicitation of a polysynaptic
SNIDER, R. S. and W. T. NIEMER, A Stereotaxic Atlas of the Cat Brain. University of Chicago
TARKHAN, A. A., Uber die Vorhandensein afferenter Fasern im Nervus hypoglossus. Arch.
TARKHAN, A. A. and I. ABOU-EL-NAGA, Sensory fibres in the hypoglossal nerve. J. Anat.
TARKHAN, A. A. and S. ABD-EL-MALEK, On the presence of sensory nerve cells on the hypo-
WOODWORTII, R. S. and C. S. SHERRINGTON, A pseudaffective reflex and its spinal path. J.
209-224.
183. 101-1 1 1 .
physiological study in normal man. Arch. Ncurol. 1961. 4. 4 8 6 4 9 8 .
hypoglosso-laryngeal reflex. Brain Res. 1968. 10. 256-258.
Press, Chicago. 1961.
Psychiat. Nervenkr. 1936. 105. 4 7 5 4 8 3 .
(Lond.) 1947. 81. 23-32.
glossal nerve. J. cornp. Neurol. 1951. 93. 219-228.
Physiol. (Lond.) 1904. 31. 234--243.