adaptations of gracilaria pacifica (rhodophyta) to nitrogen procurement at different intertidal...
TRANSCRIPT

Marine Biology 93, 569-580 (1987) Marine ==,:==m.,BiOlOg v
�9 Springer-Verlag 1987
Adaptations of Gracilaria pacifica (Rhodophyta) to nitrogen procurement at different intertidal locations
T. E. T h o m a s *, P. J. Harr i son and D. H. Turpin **
Departments of Botany and Oceanography, University of British Columbia; Vancouver, British Columbia V6T 2B1, Canada
Abstract Introduction
Nitrogen uptake, assimilation and accumulation were studied in three populations of Gracilaria pacifica Abbott in Bamfield Inlet, British Columbia, Canada, over three summers, 1979-1981. Two of these populations were in the intertidal, one high and one low, and the third was a subtidal cultured population. Nitrate uptake rates, internal nitrate content and nitrate reductase activities were highest in the low intertidal population. Time-courses of uptake and uptake kinetics were studied. Both nitrate and ammonium were taken up simultaneously. Thalli from the high-intertidal population showed enhanced nitrate and ammonium uptake following mild desiccation, and greater tolerance to desiccation in terms of maintaining nitrogen uptake after severe desiccation. Transplants were made to determine the effect of intertidal height and geographic location on responses to desiccation, nitrogen uptake, as- similation and accumulation. Nitrate and ammonium uptake rates were dependent on intertidal height and geographic location. Transplanting up the intertidal in- creased nitrate uptake and nitrate reductase activity, but decreased the nitrate content of the thalli. There were few significant differences in ammonium uptake rates, and ammonium, amino acid, and soluble-protein content of the various populations. All high-intertidal populations, transplanted or natural, showed enhanced nitrate uptake rates following desiccation. Enhanced ammonium uptake rates following desiccation were restricted to the high- intertidal thalli in only one geographic location. Tolerance to higher levels of desiccation also appeared to be inter- tidal height-dependent, but required more than five weeks to fully develop or disappear.
* Present address: Terry Fox Laboratory, B.C. Cancer Control Centre, 601 West 10th Ave., Vancouver, British Columbia V5Z 1L3, Canada
** Present address: Department of Biology, Queen's University, Kingston, Ontario K7L 3N6, Canada
Nutrient uptake rates in marine macrophytes have been related to past nutritional history (D'Elia and DeBoer, 1978; Hanisak and Harlin, 1978; Topinka, 1978; Morgan and Simpson, 1981; Rosenberg and Ramus, 1982). A high- intertidal habitat imposes drastic changes in nutrient supply as well as creating 'other environmental stresses which may affect the physiological state of seaweeds and, in turn, their ability to take up nutrients. Thomas et al. (in press) have observed enhanced nitrogen uptake rates following mild desiccation in several species of high- intertidal macrophytes. Emergence removes seaweeds from all usable nutrient supplies except carbon. Thomas and Turpin (1980) suggest that enhanced nutrient uptake following desiccation in seaweeds is an adaptation for limiting nutrient procurement in the intertidal.
Comparisons of tolerant (high-intertidal) and non- tolerant (low-intertidal) plants are frequently made be- tween different species (Jones and Norton, 1979; Dring and Brown, 1982; Thomas etal., in press). Certain species do dominate the high-intertidal, but others are found at a variety of intertidal heights. The Gracilaria pacifica Abbott (Abott, 1985) in Bamfield Inlet, Bamfield British Columbia, Canada, is one such species. It is present in natural high and low intertidal populations as well as cultured in floating pens. The high-intertidal plants are morphologically very different.
This study investigates the effects of intertidal location and mild desiccation on nitrogen uptake, assimilation, and accumulation in these three populations of Gracilaria pacifica. The effects of intertidal height were isolated from the effects of geographic location by transplanting thalli from the high- and low-intertidal populations to the location of the other natural population and to different intertidal heights.

570 T.E. Thomas et al.: Nitrogen procurement by intertidal Gracilaria
Materials and methods
Populations
The subtidal Gracilaria pacifica Abbott was cultured by Canadian Benthic Ltd. in floating pens (Lindsay and Saunders, 1979) anchored in Wiseman's Bay, Bamfield Inlet, Bamfield, British Columbia (48~ 125~ These plants were very robust, dark red and larger than the local natural plants.
The low-intertidal (1.0 m above Canadian datum) bed of Gracilaria t)ac~ca was on a rocky beach in Wiseman's Bay. These plants were dark red and weighed 5 to 10 g. Periodic measurements (3 to 5 d) of the nitrate and ammonium concentrations in the surface water were made during August and early September. On only one occasion was there a detectable level ( 1.0~tM) recorded (18 Au- gust 1981, 8 pM nitrate) in Wiseman's Bay.
The high-intertidal Gracilaria pacifica were growing in mats on a mud and shell bottom (mudflat) at the head of Bamfield Inlet, Bamfield, British Columbia (49~ 125~ This area was very shallow and surrounded by grassland or marsh. The G. pacifica mats were under direct sunlight all day. The plants were small, twisted and yellow in coloration. Moderate concentrations of ammonium (2 to 5pM) were detected a total of six times by periodic surface-water sampling in August and early September 1980 and 1981.
These populations of Gracilaria pacifica were studied over three summers, 1979-1981. In August and early September of 1979, nitrogen uptake studies were con- ducted on all three populations. Transplants were made in June 1980 and 1981. In June and August of 1980, nitrogen uptake studies were carried out on the natural and trans- planted populations but the work with the subtidal culture was discontinued. In June and August of 1981, the nitrogen uptake rates, soluble internal nitrogen, and ni- trate reductase activity of the natural and transplanted populations were monitored.
Nitrogen uptake experiments
Two types of uptake experiments were conducted. Firstly, nitrate and ammonium uptake rates were continuously measured for 40 rain (time-course studies). This involved placing 4 to 5 g of plant material into 1 liter of filtered (0.45 ktm Millipore) natural seawater with 15 ktM ammo- nium or 30ktM nitrate and excess amounts of phosphate (f/20), silicate (f/2), trace metals (f/2) and vitamins (f/2) (Guillard and Ryther, 1962). Simultaneous uptake rates of nitrate and ammonium were also measured in 30ktM nitrate and 15/~M ammonium. These concentrations were used because even rapid uptake for 1 h did not decrease the nitrogen levels enough to significantly alter uptake. They also represent the magnitude and ratio of nitrate and ammonium concentrations found in the field during periods of vertical mixing and excess freshwater runoff.
The nitrate and ammonium concentrations of the medium were measured using a Technicon Autoanalyzer (Davis etal., 1973). The medium was stirred with a magnetic stirring bar rotating at 120 rpm. The incubation chamber was kept at 16 ~ under saturating light (300~tE m -~ s -1 daylight fluorescent lighting). Uptake rates were deter- mined from the decrease in nitrate or ammonium concen- tration in the medium with time. The slight volume change, due to removal of medium for analysis (3 ml rain -1) was taken into account. Time-course uptake studies were done on hydrated plants and plants which had been previously dried to 5 to 15% desiccation (see following sub-section).
In a second type of uptake experiment, thalli were incubated in a known concentration of nitrate or ammo- nium for the time period when the uptake rate was shown to be constant in the time-course studies (10 to 15 min). Plant material weighing 2 to 3 g wet wt was placed in 400 ml of filtered (0.45 ktm) seawater enriched with 30/~M nitrate, or 15 ~M ammonium and with all other nutrients, trace metals and vitamins at saturating concentrations (see above). All incubations were outdoors under saturating natural light (Hanisak and Harlin, 1978) at 16 ~ Nitrate or ammonium concentrations in the culture medium were determined at the start ( t=0) and end of the incubation period. Plants were removed and dried to a constant dry weight in a 60~ oven. Uptake rates were expressed on a per gram dry weight basis.
This second method of measuring uptake rates was used to determine the effect of previous desiccation on nitrate and ammonium uptake rates upon resubmergence. Plants were dried (see following sub-section) and then submersed in uptake medium. Nitrite was measured separately from nitrate in cases where nitrogen excretion became apparent after a period of severe desiccation. The uptake rates were plotted against the percentage desicca- tion of the thallus prior to resubmergence. There was a great deal of inter-plant variability in the uptake data. When general trends were obvious, curves were fitted by inspection. The average uptake rate of the controls (0% desiccation) was compared to the average uptake rate of plants which had been previously dried to 5 to 15% desiccation, using one-way analysis of variance, accepting 0.05 as the significance level. The effect of nitrate and ammonium concentration (0 to 60~tM) on nitrate and ammonium uptake rates (uptake kinetics) was also deter- mined using the second method of measuring uptake rates.
Desiccation procedure
The thalli were desiccated outdoors under natural condi- tions. The controls, or hydrated plants, were suspended in a glass chamber above wet paper towels at the same irra- diance and temperature and for the same period of time as the plants which were desiccating. This allowed all plants to be emerged and starved of nutrients for the same length

T. E. Thomas et al.: Nitrogen procurement by intertidal Gracilaria 571
of time, thereby assuring that the observed changes in nutrient uptake rate upon submergence could be at- tributed to desiccation and not to recent (> 2 h) nutrient starvation. The following formula was used to calculate % desiccation: % desiccation= { (wet wt-desiccated wt)/wet wt} x 100%. Desiccated weight was the weight after des- iccation prior to resubmergence (the start of the uptake rate measurement).
C:N ratios
C:N ratios of some dried pulverised plants were deter- mined using a CHN analyser (Carlo Erba).
Analysis of soluble-nitrogen content
Complete thalli were ground in boiling water to extract nitrate, ammonium, ninhydrin-positive material (assumed to be primarily amino acids) and soluble protein. Plant material (2 g wet wt) was ground with 1 g of washed, ignited sand and 25 ml of hot, distilled, deionized water. This method had previously been found to be superior to ethanol extraction (Thomas, 1983). Extracts were done in triplicate and were stored at -10 ~ and analysed later.
The amount of soluble ninhydrin-positive material in the extracts was determined by the methods of Lee and Takahashi (1966), which is a colorimetric reaction for primary amines. The absorbance at 570 nm was assumed to be primarily due to amino acids and ammonium. Standard solutions of glycine in distilled deionized water were run with each series of analyses. Standard solutions of ammonium were also run. The absorbance due to the ammonium content (measured separately using the Auto- Analyzer) was determined from the ammonium standard curve and subtracted from the total absorbance.
A Lowry method, as modified by Leggett-Bailey (1967), was used to determine the soluble protein content of the extracts. This method measures folin-positive material, which in this system was assumed to be primarily protein. Absorbance was read at 750 nm. Bovine serum albumin was used as a protein standard.
Analytical results were normalized to wet weight be- cause the need to grind tissue before extraction precluded dry weight determinations.
Half-saturation constants and the standard error of these estimates were determined by a linear regression of V/S versus V (V=uptake rate, S=substrate concentra- tion) to give an indication of relative affinity.
Transplant procedure
Thalli were transplanted in June in 1980 and 1981. Individual plants were difficult to distinguish in both intertidal Gracilaria pacifica beds (where the thalli tended to be matted in clumps), and were therefore transplanted in this state. In 1980, the procedure was as follows: ten clumps, each weighing approximately 6 g were tied with cotton string to a 3 m long galvanized chain and anchored at each end.
In 1980, plants were transplanted from the head of Barn- field Inlet (1.8 m) to two different intertidal heights at 1.0 and 1.8 m in Wiseman's Bay. The 1.0 m site was in the natural Gracilaria pac~ca bed at that site. Plants growing naturally in Wiseman's Bay (1.0 m) were transplanted to two different intertidal heights at 1.0 and 1.8 m at the head of Bamfield Inlet. The 1.8m site was in the natural G. pacifica bed.
These transplants were made between 23 and 25 June 1980. Uptake experiments were conducted approximately five weeks later.
In 1981, this transplant procedure was expanded. Quadrate frames (0.5 m x 1.0 m) were constructed of PVC piping. Nylon fish-netting was stretched across the qua- drats. Clumps of plants were tied to each square of the net with cotton string and the quadrats were anchored at each corner with stakes. The use of quadrats allowed more biomass to be transplanted. In 1981, plants were trans- planted from the head of Bamfield Inlet at 1.8 m to 1.0 m in the same location, as well as two sites in Wiseman's Bay, one at 1.0 m and the other at 1.8 m. Plants from the natural bed (1.0 m) in Wiseman's Bay were transplanted higher up in the intertidal to 1.8 m as well as to 1.8 and 1.0m at the head of Bamfield Inlet. These transplants were made between 6 and 9 June. Six to eight weeks later, uptake experiments were run, internal soluble-nitrogen content measured, and nitrate reductase activities deter- mined.
Nitrate reductase assay
In vitro nitrate reductase activity was assayed as outlined in Thomas (1983) and Thomas and Harrison (in prepara- tion). Two grams of plant material were ground with 1 g of sand and 25 ml of phosphate buffer. Optimum pH, opti- mum polyvinyl pyrrolidone (PVP) concentration, linearity with time and extract volume, and the effect of N A D H , NO;- and Mg +4 concentrations were tested for both inter- tidal populations.
Results
Time-courses of nitrogen uptake
There were differences in the nitrate and ammonium uptake rates from June to August, but the short-term (< 1 h) characteristics of uptake such as duration of constant uptake rates and whether the rates increased or decreased, did not change. Both intertidal populations of Gracilaria pacifica took up nitrate and ammonium simul- taneously (Fig. 1).

572
A 30"
c-
O
"~ 25-
Z 0 o
I 10 20 30
T i m e
B
D
lb 2'0 3o
min)
Fig. 1. Gracilaria pacifica. Decrease in nitrate (A, B) or ammo- nium (C, D) concentration in incubation medium monitored con- tinuously over time (min) when either 30 #M nitrate or 15 pM am- monium was added (continuous lines) or when both 30 ktM nitrate and 15 gM ammonium were added (dashed lines), for low (A, C) and high (B, D) intertidal plants
The high-intertidal plants showed a constant rate of nitrate uptake for longer than 30 rain (Fig. 1 B). Nitrate uptake by the low-intertidal plants was stimulated after 15 rain of exposure to nitrate-enriched seawater (Fig. 1 A). The presence of ammonium inhibited nitrate uptake by 50% in the high-intertidal plants, but had no significant effect on nitrate uptake in the low-intertidal population.
The initial ammonium uptake rate of the low-intertidal Gracilaria pacifica was maintained for more than 30 min, whereas ammonium uptake by the high-intertidal plants decreased slowly after 10 rain (Fig. 1 C, D). The presence of nitrate increased ammonium uptake rates by 40% in the low-intertidal G. pacifica, but had no significant effect on ammonium uptake by the high-intertidal plants.
Nitrogen uptake kinetics
Nitrate uptake in the low-intertidal Gracilaria pacifica appeared to have a hyperbolic component at low nitrate concentrations and a linear component at nitrate concen- trations between 15 and 50/~M (Fig. 2A). If the uptake due to this linear (possibly diffusion) component is sub- tracted from the uptake rates (D'Elia and DeBoer, 1978) the resulting uptake curve gives an estimation of the active uptake component which had an approximate Ks of 1 pM. The uptake rates of the high-intertidal G. pacifica followed saturable kinetics when plotted as a function of nitrate concentration (Fig. 2B). The Vmax was approximately 4 #mol g-~ dry wt h -~ and the Ks was approximately 6 #M.
There were also differences in the ammonium uptake kinetics between the two intertidal populations. The
T. E. Thomas et al.: Nitrogen procurement by intertidal Gracilaria
ammonium uptake rates for 'the low-intertidal Gracilaria pacifica were approximately proportional to the ambient ammonium concentration up to 40/~M (Fig. 3 A). Ammo- nium uptake kinetics for the high-intertidal G. pac~'ca were saturable but did not clearly follow a rectangular hyperbola (Fig. 3 B). The Vmax was approximately 30#tool g-~ dry wt h -I and the Ks was approximately 10 #M.
Nitrate uptake
Effect of desiccation - natural populations
The natural high-intertidal Gracilaria pacifica showed a response to desiccation not apparent in the natural low- intertidal and subtidal plants. Mild desiccation caused a later increase in nitrate uptake when the plants were sub- merged (Fig. 4, Table 1). These enhanced uptake rates were 2.5 (1980) and 5.0 (1981) times the uptake rates of the fully hydrated plants (controls) and only occurred in late summer and not in June. Such relative increases in uptake rates will be referred to as the relative degree of enhancement of uptake.
Nitrate uptake in the subtidal population dropped to zero after only 10% desiccation, and nitrate, instead of the expected nitrite, was excreted after greater desiccation (Fig. 4). The response of the natural low-intertidal popula- tion was intermediate between the other two populations. There was no enhancement of uptake following desicca- tion, but positive nitrate uptake rates were maintained after up to 30% desiccation (Fig. 4).
Effect of desiccation - transplants
The plants transplanted from Wiseman's Bay (1.0 m) to the head of Bamfield Inlet (1.8 m) responded to mild desiccation in a manner similar to the natural high-inter- tidal plants at the head of Bamfield Inlet. The relative enhancement of nitrate uptake for these plants was 1.3 and 3.0 in 1980 and 1981, respectively (Fig. 5). Their tolerance to higher desiccation (20 to 60%) in terms of nitrate uptake was less than that of the resident plants at the head of Bamfield Inlet. A complete set of data is presented elsewhere (Thomas, 1983).
Moving the high-intertidal plants from the head of Bamfield Inlet to a high-intertidal site in Wiseman's Bay (1.8 m) did not alter their response to desiccation: they showed enhanced nitrate uptake rates following desicca- tion (Fig. 5). These plants also retained their tolerance to higher degrees of desiccation. Uptake rates after 30% desiccation were equal to those of the control (hydrated) plants (Thomas, 1983). When these high-intertidal plants were moved to a low-intertidal site in Wiseman's Bay they showed no increase in nitrate uptake following mild desiccation (Fig. 5). However, these plants retained their tolerance to higher desiccation; plants dried to 30% desic- cation showed only slightly lower nitrate uptake rates

T. E. Thomas et al.: Nitrogen procurement by intertidal Gracilaria
>,, 24
rob
E
16 cr
0,.
8
4,--'
z
A
o |
/./ L. ', ' u ' �9 ' , '
10 20 3 0 40
Nitrate Concentrat ion (pM)
573
I c"
-s To~
0 E
rr
G)
Z
.
4-
2 " / /
i I
I
g 8
o �9 o 4
/
o , "
8 ,# o
o
o
o . . . . . . . . . . .
I
o
l b " 2o 3'0 4o B Nitrate Concentrat ion (pM)
Fig. 2. Gracilaria pacifica. Relationship between nitrate uptake rates and nitrate concentration for natural populations of low (A) and high (B) intertidal plants in August 1981. Uptake rates minus non-saturable component (dot-dashed line in "A") were determined to assess active up- take component. Non-saturable component was determined from slope of the line at higher ni- trate concentrations
upon resubmergence than the hydrated plants (Thomas, 1983).
The plants transplanted from 1.0 m in Wiseman's Bay to 1.0m at the head of Bamfield Inlet and the plants moved down the intertidal zone at the same site showed no enhancement of nitrate uptake after desiccation (Fig. 5 B). Transplanting up the intertidal zone in Wise- man's Bay resulted in a 1.5-fold enhancement in nitrate uptake followig desiccation.
Uptake rates of hydrated plants
The nitrate uptake rates of either transplanted or resident plants growing in Wiseman's were 3 to 5 times greater
than the plants growing at the head of Bamfield Inlet (Fig. 5). All but one of the transplants between these two geographical locations resulted in nitrate uptake rates which were intermediate between the two natural popula- tions (Fig. 5).
Moving up the intertidal zone in Wiseman's Bay increased the nitrate uptake rates of control plants from 10.2 to 14.5 j~mol g-1 dry wt h -1 (Fig. 5 B). Moving down the intertidal zone at the head of Bamfield Inlet had no significant effect on nitrate uptake rates.
Nitrate uptake rates for the natural low-intertidal population were much lower in 1979 than in the other two years (Table 1). Such large year-to-year differences did not occur in June. The high-intertidal plants took up nitrate more rapidly in June than in August.

574 T.E. Thomas et al.: Nitrogen procurement by intertidal Gracilaria
-~ 60.
-5 E :3_
4--'
cO 4 0 rr �9
E "' 20, E O E E ,<
A
$ * | �9
2 '0 4'0 6'0 Ammonium Concentrat ion (~M)
8b
T e-
3 0 "0 T
O~
-5 E
20 cO r r
ID
Q_
IO- E
t -
O
E E <
B
o~ /
o /
l o $
/o O / ~
* /
/
/
g /o
l � 9 /
/ g
0 0 0 0
o o o
o / o / " o
o / o / o /
/ o g
2'O 4'0 6'O Ammonium C o n c e n t r a t i o n (tJM)
Ammonium uptake
Effect o f desiccation - natural populations
The high-intertidal Gracilaria pacifica showed significant enhancemen t of ammonium uptake following mild desiccation in August 1980 and 1981 (Table 1). This enhancement was not as pronounced as it was with nitrate. The low-intertidal G. pacifica showed no enhancement of ammonium uptake following desiccation.
Effect o f desiccation - transplants
The loss of transplanted material in 1980 made an in- vestigation of effect of desiccation on the ammonium
80
Fig. 3. Gracilaria paeifica. Relationship between ammo- nium uptake rates and ammonium concentration for natu- ral populations of low (A), and high (B) intertidal plants in August 1981
uptake rates of the plants transplanted to 1.0 m at the head of Bamfield Inlet and to 1.8 m in Wiseman's Bay impossible. The resident plants at the head of Bamfield Inlet as well as the plants transplanted to the natural bed (1.8 m) at the head of Bamfield Inlet showed increased ammonium uptake rates following mild desiccation in 1980 and 1981 (Fig. 5). The two transplants at 1.8 m in Wiseman's Bay showed enhanced nitrate uptake after desiccation, but no increased ammonium uptake (Fig. 5 B). Plants transplanted from the head of Bamfield Inlet to 1.8 m in Wiseman's Bay showed a greater tolerance to desiccation than the low-intertidal Gracilaria pacifica," ammonium uptake rates only decreased by approximately one-fifth after plants had been dried to 30% desiccation (Thomas, 1983).

T. E. Thomas et al.: Nitrogen procurement by intertidal Gracilaria 575
Table 1. Gracilaria pacifica. Nitrate and ammonium uptake rates ~umol g-1 dry wt h -1) for three pop- ulations. Values are an average rate +95% confidence limits. * Indicates significant enhancement of uptake rate following desiccation (desicc.) (one-way ANOVA, accepting 0.05 as significance level). High-intertidal plants were from head of Bamfield Inlet, low-intertidal plants from Wiseman's Bay
Intertidal Nitrate uptake Ammonium uptake location
Control (n) 5-15% desicc. (n) Control (n) 5-15% desicc, (n)
Aug.-Sep. 1979 high 5.9• (3) 7.4• (4) 20.6• (3) 22.9• (4) low 5.2• 1.4 (6) 3.2 • 1.1 (6) 7.6 _+ 1.4 (4) 5.7 • 0.4 (4) subtidal 3.8• (3) -1.6• (5) ND ND
Aug.-Sep. 1980 high 4.2• 1.9 (3) 11.4• (3) 8.8• (3) 11.1• (4) low 27.2• (3) 26.6• (3) 12.2• (3) 11,6• (3)
Aug.-Sep. 1981 high 2.2• 1.5 (3) 10.8• 1.5" (8) 13.8_+ 1.0 (4) 17,7• (4) low 10.2• 6.3• 26.6• 27,1_+1.0(6)
June 1980 high 10.2• 1.6 (7) 9.6• 1.7 (7) 13.6• 1.1 (3) 11,8• (4) low 12.2• (4) 10.7• (4) 11.1• (3) 11.2• (3)
June 1981 high 6.5• (3) 14.1• (4) 18.2_+4.2 (4) 19.9_+4.2 (8) low 13.9• (3) 18.4• (9) 18.1• (4) 18.7_+ 1.9 (8)
T
"O 7
-4'
c~
O.
~-8.
Z
j1 -~" o~
�9 \' '~ k
\. ~ z~ . \ a \ . a
', . " \ . , o \
~ �9 \
A
i
a - - I
2b 4b % Desiccation
6'0
Fig. 4. Gracilaria pacifica. Relationship between nitrate uptake rates and % desiccation prior to submergence in late summer 1979 for natural high (�9 and low (o) intertidal and subtidal (a) plants
The natural populat ion in Wiseman's Bay as well as all populat ions transplanted to the low-intertidal site in Wise- man ' s Bay or the head of Bamfield Inlet, showed no en- hancement of a m m o n i u m uptake following mild desicca- tion (5 to 15%) and had a low tolerance to higher desiccation. After 30% desiccation, a m m o n i u m uptake rates were less than ha l f the rates after 0% desiccation (Thomas, 1983).
Uptake rates of hydrated plants
The a m m o n i u m uptake rates in June for the control (0% desiccation) high- and low-intertidal plants were very similar and did not differ significantly from year to year, but the former was not true later in the summer (Table 1). In both 1980 and 1981 the Wiseman's Bay populat ions had higher uptake rates than those at the head of Bamfield Inlet, but the reverse was true in 1979 (Table 1). The ammon ium uptake rates of the transplanted and resident plants were similar, with the exception of the plants transplanted down the intertidal zone at the head of Bamfield Inlet, which showed an increase in a m m o n i u m uptake rate (Fig. 5 B).
Soluble-nitrogen content
In June, both intertidal populat ions of Gracilaria pacifica had very low nitrate content, approximate ly 0.04 #mol g-1

576 T.E. Thomas et al.: Nitrogen procurement by intertidal Gracilaria
1980
WISEMAN'S BAY
( d a r k p l a n t s )
HEAD OF BAMFIELD INLET
( l i g h t p l a n t s )
1 .8m- T r a n s p l a n t
NO 3 22 .5 • '.Oi~xl. 2)
1.0va- N~t~lve
NO~ 2 7 . 2 • +
Nit 4 1 2 . 0 •
A
T r a n s p l ~ ~ / / ] - 1 . 8 m S
�9 / / NO 3 7 . 6 + 1 . 0 ( x l . 3 ) 4 . 2 •
,, . o + ? ; 8 ( x l . 7) 8 . 8 •
'1 . .
T r a n s p l a n t s T r a n s p l a n t s -1 .0m
35.0+5.6" ' . id no t s u r v i v e
- / \ / ' - J " - / ~ / : 7 �9 -:\ _ . . . ; / .
1981
WISEMAN'S BAY
( d a r k p l a n t s )
HEAD OF BAMFIELD INLET
( l i g h t p l a n t s )
T r
2 3 . 0 + 1 . 1 " f . l ,NS ) . .
/ / / '
1 Ova- T r a n s p l a n t �9 *xat l ye ~ r a n s p l a n t s / '~
NO 3 - 10.2+_0.7 7 . 5 + 3 . fi 4.5_~1.3 2.2_+_0.4
B NH 4 2 6 . 6 • 22 5_-t_5 6.:. % ~ , ~ 15.5_+_3.8 28.5_+_.3.8
1.8m- ' T r a n s p ) a n t s a ' - l . 8 m _ - - ...~,,
NO 3 NO 3 2 . 3 + 1 . 3 ( x 3 . 0 ) 2 . 2 + 1 . 5 ( x 5 . 0 )
,< ~ . o . . ;:::\-- ~ ; lo.~il."- ~(~.~ ) 1~.8~1.o(~1 .~ ) "-'~..." 2,?
- 1. Ova
Fig. 5. Gracilaria pacifica. Summary of nitrate and ammonium uptake rates (umol g-1 dry wt h -1 _+ 95% confidence limits) of hydrated plants in August 1980 (A), and 1981 (B). Values in parentheses show relative degree of enhancement of uptake following mild desiccation (5 to 15%). Arrows indicate direction of the transplants

T. E. Thomas et al.: Nitrogen procurement by intertidal Gracilaria 577
Table 2. Gracilaria pacifica. Total C:N (by atoms) and soluble nitrate, ammonium, amino acids and protein content per gram wet weight in natural populations of high- and low-intertidal plants in June and August 1981, _+ 95% confidence limits, n = 3
Population Nitrate Ammonium Amino acids Protein C:N (pmol) (/zmol) (pmol) (mg)
High intertidal June 0.04+_0.04 0.8_+0.6 ND ND ND Aug. 0.57_+0.50 2.2-+0.3 6.9-+2.1 7.7• 9.0-+ 1.2
Low intertidal June 0.03-+0.05 2.3_+0.8 Aug. 4.40_+1.30 4.9_+1.4 5.5_+3.3 9.5_+1.6 13.0_+0.9
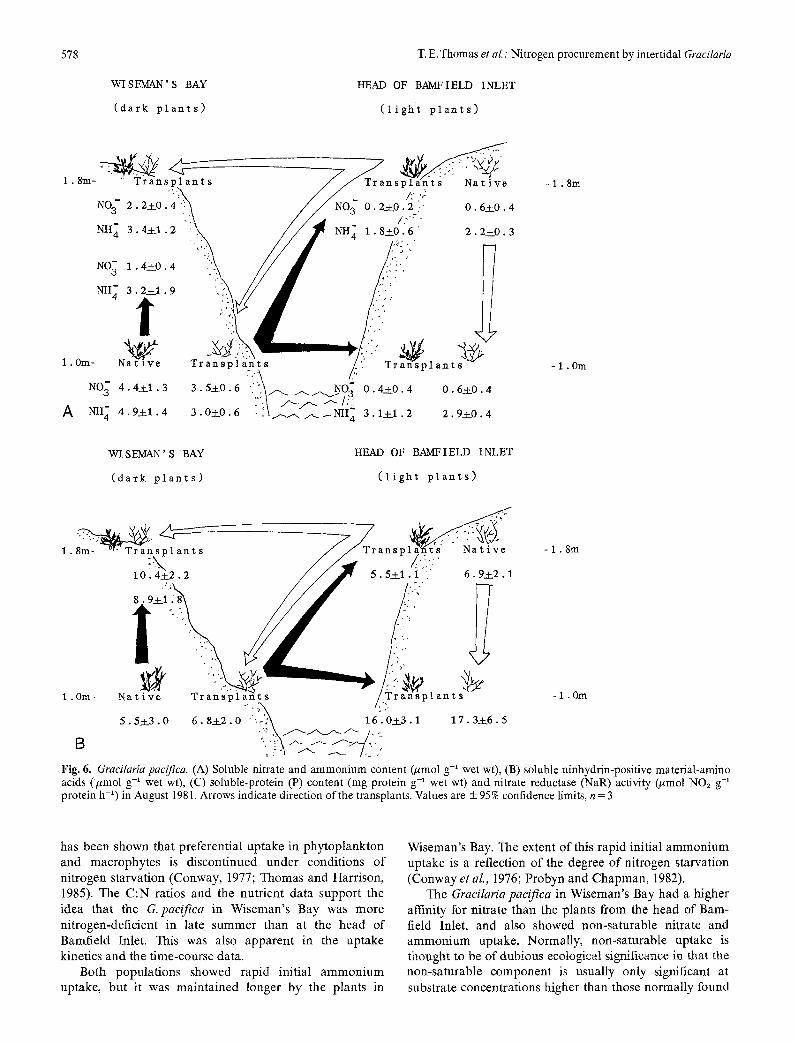
wet wt or 0.02% of the total nitrogen in the plant (Table 2). The nitrate content of the plants in Wiseman's Bay in late summer were much greater than at the head of Bamfield Inlet and either natural populations in June (Table 2, Fig. 6 A). Transplanting up the intertidal in Wiseman's Bay resulted in a decrease in the nitrate content.
The precise amounts of ammonium, amino acids, and protein extracted were not determined due to the crude nature of the extracts which may contain compounds that interfere with the colorimetric analysis. Nevertheless, general trends rather than absolute amounts can be discussed. The ammonium content of the high-intertidal plants increased three-fold from June to August; this trend was not as drastic in the low-intertidal population (Ta- ble 2). Transplanting had little or no effect on the ammoni- um content (Fig. 6 A).
There was no significant difference between the two natural populations in the total amount of soluble nin- hydrin-positive material per gram (Table 2), but the thalli transplanted to 1.0 m at the head of Bamfield Inlet had a uniquely high average soluble-amino acid content (Fig. 6 B). The plants in Wiseman's Bay which had origi- nated at that site had a higher soluble-protein content than the other Gracilaria #acifica populations tested (Fig. 6C). Moving these plants to the head of Bamfield produced plants with a very low soluble-protein content (Table 2, Fig. 6 C).
Nitrate reductase activity
Nitrite production was linear with time, and activity was proportional to the volume of extract used (Thomas, 1983): The nitrate reductase enzyme was insensitive to Mg ++ concentration, pH change, and PVP concentration. Nitrate and NADH supply had a similar effect on the nitrate reductase activity for both intertidal populations. The half- saturation constants (Ks_+ 1SE) for both nitrate and NADH were 70_+ 7.0 and 75_+ 46, respectively (Thomas, 1983).
In June, the Vma x for the nitrate reductase activity of both natural populations was about 150#mol NO2 g-1 protein h-L In August, the natural population of Gracila- ria pacifica in Wiseman's Bay had twice the nitrate
reductase activity of the natural population at the head of Bamfield Inlet (Fig. 6 C). Transplanting from Wiseman's Bay to the head of Bamfield Inlet (at both the high- and low-intertidal site) resulted in a decrease in nitrate reductase activity (Fig. 6 C). Moving up the intertidal zone in Wiseman's Bay increased the already high nitrate reductase activity. Transplanting from the head of Barn- field Inlet to 1.0 m in Wiseman's Bay increased nitrate reductase activity. When plants were transplanted to 1.8 m in Wiseman's Bay from the head of Bamfield Inlet, the nitrate reductase activity remained constant at about 45 tool NO2 g-1 protein h -1.
C:N ratios
The high-intertidal plants had a significantly lower C:N than the low-intertidal plants (Table 2).
Discussion
Recent studies have revealed the morphological plasticity of Gracilaria pacifica (Bird et al., 1982; Abbott, 1985). The present study has shown that there is great flexibility in the nitrogen physiology of G. pacifica. These differences were demonstrated in field plants and were not the potentials of laboratory cultures. The natural G. pacifica populations differed in geographical location and inter- tidal height. The first caused differences in substrate, nutrient regime, irradiance and wave action," and the second meant changes in time emerged and subsequent desiccation, exposure to direct sunlight and removal from nutrient supplies.
Nitrate appeared to be a more important source of nitrogen in Wiseman's Bay than at the head of Bamfield Inlet. This was not surprising considering the greater chance of influx of nitrate-rich deep water in Wiseman's Bay and the occasional presence of ammonium at the head of Bamfield Inlet. These differences were reflected in the nitrogen physiology of the plants. The Gracilaria pacifica in Wiseman's Bay had higher nitrate uptake and assimilation rates and larger nitrate reserves, and the presence of ammonium did not inhibit nitrate uptake. It
578
WI SEMAN' S BAY
(da rk p l a n t s )
T. E. Thomas et aL: Nitrogen procurement by intertidal Gracilaria
HEAD OF BAMFIELD INLET
( l i g h t p l a n t s )
. < T -...:,...7;;. 1 .8m- " T r a n s p l a n t s r a n s p l a n t s N a t i v e - 1 . S m
" ' - I : 'J . 4 NO 3- 2 . 2 + 0 . 4 ~ / / ~ N O 3 : ;;.:b_O. 2 '.'//H., 0.6_+_0
NH; 3.4_+_1.2 "!:~ / / # NH4 1.8+0.6' 2.2+0.3
�9 m- N a t i v e T r a n s p l a n t s " r - 1 . 0 m
NO 3 4.4_+_1.3 3 .5+0 .6 " ~ . / . . . .A . jN03 0 .4+0 .4 O. 6+0.4 '-":/ 1 - - / - , / " - / : ;
A NH; 4 . 9 • 3 . 0 + 0 . 6 - . ' ~ / - ~ x / . . _ N I - I 4 3 . 1 + 1 2 2 . 9 + 0 . 4
WI SEMAN ' S BAY
( d a r k p l a n t s )
HEAD OF BAMFIELD INLET
( l i g h t p l a n t s )
1 . v_, . , _ _ s p l a n t s a n s p / . : , ' . ' - 1 . S m
1 0 . 4 + 2 . . 5 + 1 . 1 " 6 . 9 + 2 . 1 ;.7."
8 �9 9-t-1,
1 . 0 m - N a t i v e T r a n ' s p l ~ t t s l a n t s - 1 . 0 m
�9 < . ' ~ 1 6 ". 0+3 . 1 1 7 . 3 + 6 . 5 5.5_+_3 �9 0 6 �9 8..+_2 0 -'=' .;." , .X, /~- . . -x_ .~ .~ /. =.
B ". \! .~/._ 1,_ ~- - , , - / . . �9 r q ~ ~ t ". ,"
Fig. 6. Gracilaria pacifica. (A) Soluble nitrate and ammonium content Cumol g-1 wet wt), (B) soluble ninhydrin-positive material-amino acids (/xmol g-1 wet w0, (C) soluble-protein (P) content (rag protein g-1 wet wt) and nitrate reductase (NaR) activity (umol NO~ g-~ protein h -~) in August 1981. Arrows indicate direction of the transplants. Values are _+ 95 % confidence limits, n = 3
has been shown that preferential uptake in phytoplankton and macrophytes is discontinued under conditions of nitrogen starvation (Conway, 1977; Thomas and Harrison, 1985). The C:N ratios and the nutrient data support the idea that the G. pacifica in Wiseman's Bay was more nitrogen-deficient in late summer than at the head of
Bamfield Inlet. This was also apparent in the uptake kinetics and the time-course data.
Both populations showed rapid initial ammonium uptake, but it was maintained longer by the plants in
Wiseman's Bay. The extent of this rapid initial ammonium uptake is a reflection of the degree of nitrogen starvation (Conway et al., 1976; Probyn and Chapman, 1982).
The Gracilaria pacifica in Wiseman's Bay had a higher affinity for nitrate than the plants from the head of Barn- field Inlet, and also showed non-saturable nitrate and ammonium uptake�9 Normally, non-saturable uptake is thought to be of dubious ecological significance in that the non-saturable component is usually only significant at substrate concentrations higher than those normally found

T. E. Thomas et al.: Nitrogen procurement by intertidal Gracilaria
WISEMAN'S BAY HEAD OF BAMFIELD INLET
(dark p l a n t s ) ( l i g h t p l a n t s )
579
NaR 43[ffL N / / ~ I ~ N a R 60.i~':
P 8 .3+1.2 / / / P 6:4+2.1
N 1.0m- l'~a t i r e Transpl"ant s [.-':
C
~la "..: . . . .
44.2
7.7.+_2.0
T r a n s p l a n t s
NaR 85.0 50.9- ' i NaR 14.2 26.9
P 9 .5+1.6 8.0_.+_1.9\~'-'~" ~ i,__ ~-/:p 6.7_+._2.2 7 .3• -i/~ l-c. ~ / / .
$ -
- 1 . 8 m
- 1 . 0 m
Fig. 6C
in the field. The nitrate uptake versus nitrate concentration plot for the resident plants from Wiseman's Bay showed that the linear component makes a considerable contribu- tion to uptake at > 5/~M nitrate. It is conceivable that these G. pacifica obtained a large portion of their nitrogen from nitrate pulses of such magnitude.
The nature of the uptake experiments in this study is such that the resulting uptake rates were an estimate of short-term uptake which is often rapid, has low substrate affinity, and appears to be uncoupled from growth (Probyn and Chapman, 1982). Short-term uptake can be important to long-term nitrogen procurement if the nitro- gen supply is periodic or in "patches". Ryther et al. (1982) have shown that Gracilaria tikvahiae can maintain maxi- mum growth rates if only exposed to nitrogen-enriched seawater for 6 h every 2 wk. There is some evidence that there were "patches" of higher nitrate and ammonium in the Bamfield area.
Periodic emergence also imposes an intermittent nitro- gen regime. Transplanting up the intertidal zone led to increased nitrate uptake rates and nitrate reductase ac- tivity but decreased internal nitrate levels, suggesting accelerated nitrate utilization. The most likely explanation is that the limited and interrupted nitrogen supply caused by periodic emergence produces a requirement for ac- celerated nitrogen procurement when the alga is sub- merged. Enhancement of uptake after desiccation would be an additional means of accomplishing this.
Previous studies have shown that high-intertidal macrophytes have enhanced nitrogen uptake rates fol-
lowing mild desiccation (Thomas and Turpin, 1980; Thomas etal., (in press). The present study with Gracilaria pac~ca reveals that this is an intraspecific as well as an interspecific adaptation, which is dependent on intertidal height and not on geographical location or plant origin. Plants transplanted to high-intertidal sites devel- oped this physiological adaptation for nitrogen procure- ment, while those transplanted to the low-intertidal sites did not. The only exceptions were the transplants at 1.8 m in Wiseman's Bay, which showed enhanced nitrate uptake rates but no enhancement of ammonium uptake. The ammonium uptake rates of hydrated plants were very high, and similar to the Vmax found during the measure- ment of ammonium uptake kinetics. It is likely that further enhancement of uptake was physiologically impossible.
Another possible adaptation to a high-intertidal loca- tion is the maintenance of normal nitrogen uptake rates following a greater degree of desiccation (20 to 30%). In the transplant experiment, this tolerance to desiccation appeared to depend on the intertidal location; but the transplanted plants often showed uptake rates, following 30% desiccation, which were between those of the resident plants and the population from which they came (Thomas, 1983). This phenomenon may simply depend upon inter- tidal level and not on the origin of the thallus, but require more than five weeks to develop or disappear.
The transplant experiments in this study confirmed the hypothesis that enhanced nutrient uptake following desic- cation is dependent primarily on attachment height in the intertidal zone. The nitrogen uptake and assimilation rates

580 T. E. Thomas et al.: Nitrogen procurement by intertidal Graeilarh7
are controlled by complex interactions between certain physical factors and the physiological state of the thallus. Previous nutrient supply, the nutritional status of the thallus, a history of repeated periodic desiccation, the previous degree o f desiccation, and the type and amount of nutrient present are some of the variables involved. These factors can be controlled in laboratory cultures, but observed responses are of questionable ecological signifi- cance. Field studies must be conducted and some factors can be defined and manipulated using transplants.
Acknowledgements . This research was conducted with funds from the National Sciences and Engineering Research Council of Canada (NSERC) and with aid of a NSERC Postgraduate Fellowship to one o f us (TET). We acknowledge the use of facilities at Bamfield Marine Sta- tion, British Columbia, and thank Mr. P. Thompson for his assistance with the AutoAnalyzer.
Literature cited
Abbott, I. A.: New species of Graeilaria Grev. (Gracilariaceae, Rhodophyta) from California and Hawaii. In: Taxonomy of economic seaweeds, pp 115-121. Ed. by I. A. Abbott. La Jolla, California: Sea Grant Publishers 1985
Bird, C. J., J. P. Van der Meer and J. McLachlan: A comment on Gracilaria verrucosa (Huds.) Papenf. (Rhodophyta: Gigartina- les). J. mar. biol. Ass. U.K. 62, 453-459 (1982)
Conway, H. L.: Interactions of inorganic nitrogen in the uptake and assimilation by marine phytoplankton. Mar. Biol. 39, 221-232 (1977)
Conway, H. L., P. J. Harrison and C. O. Davis: Marine diatoms grown in chemostats under silicate or ammonium limitation. II. Transient response of Skeletonema eostatum to a single addition of the limiting nutrient. Mar. Biol. 35, 187-199 (1976)
Davis, C. O., P. J. Harrison and R. C. Dugdale: Continuous cul- ture of marine diatoms under silicate limitation. I. Syn- chronized life cycle of Skeletonema costatum. J. Phycol. 9, 175-180 (1973)
D'Elia, C. F. and J. A. DeBoer: Nutritional studies of two red algae. II. Kinetics of ammonium and nitrate uptake. J. Phycol. 14, 266-272 (1978)
Dring, M. J. and F. A. Brown: 13hotosynthesis of intertidal brown algae during and after periods of emersion. A renewed search for physiological causes of zonation. Mar. Ecol. Prog. Set. 8, 301-308 (1982)
Guillard, R. R. L. and J. H. Ryther: Studies on marine planktonic diatoms. I. Cyclotella nana (Hustedt) and Detonula confer- vaceae (Cleve) Gran. Can. J. Microbiol. 8, 229-239 (1962)
Hanisak, M. D. and M. M. Harlin: Uptake of inorganic nitrogen by Codium fragile subsp, tomentosoides (Chlorophyta). J Phycol. 14, 450-454 (1978)
Jones, H. G. and T. A. Norton: Internal factors controlling the rate of evaporation from fronds of some intertidal algae. New Phytol. 83, 771-781 (1979)
Lee, Y. P. and T. Takahashi: An improved colorimetric deter- mination of amino acids with the use of ninhydrin. Analyt. Biochem. 14, 17-77 (1966)
Leggett-Bailey, J.: Miscellaneous analytical methods: estimation of protein Folin-Ciocalteu reagent. In: Techniques in protein chemistry, 2nd. ed. pp 340-342. New York: Elsevier 1967
Lindsay, J. G. and 1L G. Saunders: Experiments with Gracilaria in a floating algal culture system, 40pp. Victoria, British Columbia: Ministry of Environment 1979 (Fisheries Develop- ment Report No. 17)
Morgan, K. C. and F. J. Simpson: The cultivation of Palmaria palmata. Effect of light intensity and nitrate supply on growth and chemical composition. Botanica mar. 24, 273-277 (1981)
Probyn, T. A. and A. R. O. Chapman: Nitrogen uptake charac- teristics of Chordaria JTagelliformis (Phaeophyta) in batch mode and continuous mode experiments. Mar. Biol. 71, 129-133 (1982)
Rosenberg, G. and J. Ramus: Ecological growth strategies in the seaweeds Graeilaria foliifera (Rhodophyceae) and Ulva sp. (Chlorophyceae): soluble nitrogen and reserve carbohydrates. Mar. Biol, 66, 251-259 (1982)
Ryther, J. H., N. Corwin, T. A. DeBusk and L. D. Williams: Nitrogen uptake and storage by the red alga Gracilaria tikvahiae (McLachlan, 1979). Aquaculture, Amsterdam 26, 107-115 (1982)
Thomas, T. E.: Ecological aspects of nitrogen uptake in intertidal macrophytes, 207pp. Ph.D. thesis, University of British Columbia, Vancouver, Canada 1983
Thomas, T. E. and P. J. Harrison: Effect of nitrogen supply on nitrogen uptake, accumulation and assimilation in Porphyra perforata (Rhodophyta). Mar. Biol. 85, 269-278 (1985)
Thomas, T. E. and P. J. Harrison: fn vivo and in vitro nitrate reductase activity in three intertidal seaweeds. (In preparation)
Thomas, T. E. and D. H. Turpin: Desiccation enhanced nutrient uptake rates in the intertidal alga Fucus distichus. Botanica mar. 23, 479-481 (1980)
Thomas, T. E., D. H. Turpin and P. J. Harrison: Desiccation en- hanced nitrogen uptake rates in intertidal seaweeds. Mar. Biol. (In press).
Topinka, J. A.: Nitrogen uptake by Fucus spiralis (Phaeophy- ceae). J. Phycol. 14, 241-247 (1978)
Date of final manuscript acceptance: September 12, 1986. Communicated by R. S. Carney, Baton Rouge