activation of basal forebrain cholinergic neurons differentially regulates brain-derived...
TRANSCRIPT

Neuroscience Letters, 136 (1992) 203-208 203 © 1992 Elsevier Scientific Publishers Ireland Ltd. All rights reserved 0304-3940/92/$ 05.00
NSL 08438
Activation of basal forebrain cholinergic neurons differentially regulates brain-derived neurotrophic factor mRNA expression in different
projection areas
L.L. Boatell a'c, N. Lindefors b, M. Ballarin a, E Ernfors a, N. Mahy c and H~ikan Persson a
Department of aMedical Chemistry, Laboratory of Molecular Neurobiology and b pharmacology, Karolinska Institutet, Stockholm (Sweden) and c Unidad de Bioquimiea, Facultad de Medicina, Universidad de Medicina, Universidad de Barcelona, Barcelona (Spain)
(Received 30 September 1991; Revised version received 29 November 1991; Accepted 2 December 1991)
Key words." NGF family of neurotrophic factors; In situ hybridization; Synaptic activation; Quisqualate; Muscimol
Afferent cholinergic pathways from the basal forebrain were activated by injections of the glutamate analog quisqualate either into the nucleus basalis or into the medial septal nucleus. Nucleus basalis injections had no effect on the expression of brain-derived neurotrophic factor (BDNF) mRNA in its neocortical projection areas as measured by in situ hybridization. In contrast, 7 h after an injection into the septum the level of BDNF mRNA increased 3- to 5-fold in the dentate gyrus, throughout CA1 to CA3 in the hippocampus and in the piriform cortex.
Brain-derived neurotrophic factor (BDNF) first iso- lated from pig brain [2] has been shown to prevent neu- ronal death in the nodose ganglion and dorsal root ganglion in vivo [10] and to support the survival of re- tinal ganglion cells in culture [12]. Molecular cloning and sequence analysis of a cDNA clone for BDNF from pig brain has shown striking amino acid similarities to that of nerve growth factor [13], a well-characterized neuro- trophic factor that supports the survival of peripheral sympathetic and neural-crest derived sensory neurons [14, 15, 20].
BDNF mRNA is present in the adult rat brain where it is expressed in specific sets of neurons with the highest levels of BDNF mRNA in the hippocampus [5, 9, 19, 21]. Outside the hippocampus, BDNF mRNA has been found in cerebral cortex, claustrum, endopyriform nucleus, amygdala and cerebellum [5, 9, 19, 21]. During development BDNF mRNA is maximally expressed in the brain at 2 weeks after birth [4]. A transient expression of BDNF mRNA during early postnatal brain develop- ment has been detected in several structures, notably in the substantia nigra and interpeduncular nucleus [7]. In contrast, the distribution of BDNF mRNA expressing neurons within the hippocampus mostly persists during postnatal brain development [6, 7] suggesting that this
Correspondence: L.L. Boatell, Department of Pharmacology, Labora- tory of Molecular Neurobiology, Karolinska Instituter, Stockholm, Sweden.
factor is not only important to maintain a proper neu- ronal innervation in the hippocampus but also plays a role in the formation of synaptic contacts and neuronal maturation during brain development.
The level of BDNF mRNAs in the dentate gyrus has been shown to increase after chronic seizures produced by lesion in the dentate gyrus [11]. Moreover, the level of BDNF mRNA increases markedly in several different brain regions, including the hippocampus, after injection of kainic acid [1, 8, 22]. In cell cultures this increase has been suggested to be due to glutamate interacting with non-NMDA receptors probably of the kainate subtype [22]. These results suggest that expression of BDNF mRNA in the brain is regulated by neuronal activity via glutamate receptors of the non-NMDA type.
In this report we show that an activation of basal fore- brain cholinergic neurons in the medial septal nucleus containing afferents to the hippocampus results in a marked increase of BDNF mRNA expression in the hip- pocampus and piriform cortex. In contrast, activation of cholinergic afferents from nucleus basalis to the cerebral cortex did not result in a change in the level of BDNF m R N A in any brain areas studied. This indicates that the neuronal activity of basal forebrain cholinergic afferents regulates BDNF mRNA expression differently in differ- ent projection areas.
Male Sprague-Dawley rats (b.wt. 250-300 g; n = 49) were anaesthetized with halothane and mounted in a stereotaxic frame. The skull was exposed and holes were
I I
i i
204
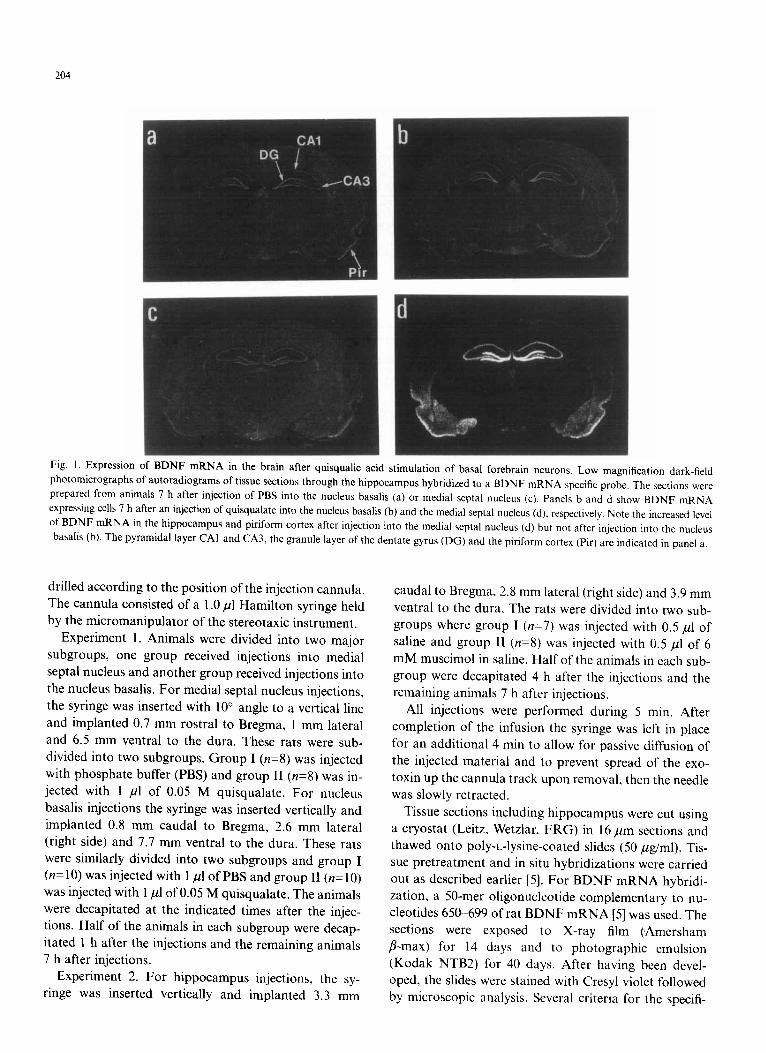
Fig. I. Expression of BDNF mRNA in the brain after quisqualic acid stimulation of basal forebrain neurons. Low magnification dark-field photomicrographs of autoradiograms of tissue sections through the hippocampus hybridized to a BDNF mRNA specific probe. The sections were prepared from animals 7 h after injection of PBS into the nucleus basalis (a) or medial septal nucleus (c). Panels b and d show BDNF mRNA expressing cells 7 h after an injection of quisqualate into the nucleus basalis (b) and the medial septal nucleus (d), respectively. Note the increased level of BDNF mRNA in the hippocampus and piriform cortex after injection into the medial septal nucleus (d) but not after injection into the nucleus basalis (b). The pyramidal layer CAI and CA3, the granule layer of the dentate gyrus (DG) and the piriform cortex (Pir) are indicated in panel a.
drilled according to the position of the injection cannula. The cannula consisted of a 1.0/11 Hamilton syringe held by the micromanipulator of the stereotaxic instrument.
Experiment 1. Animals were divided into two major subgroups, one group received injections into medial septal nucleus and another group received injections into the nucleus basalis. For medial septal nucleus injections, the syringe was inserted with 10 ° angle to a vertical line and implanted 0.7 mm rostral to Bregma, 1 mm lateral and 6.5 mm ventral to the dura. These rats were sub- divided into two subgroups. Group I (n=8) was injected with phosphate buffer (PBS) and group II (n=8) was in- jected with 1 /11 of 0.05 M quisqualate. For nucleus basalis injections the syringe was inserted vertically and implanted 0.8 mm caudal to Bregma, 2.6 mm lateral (right side) and 7.7 mm ventral to the dura. These rats were similarly divided into two subgroups and group I (n=10) was injected with 1 ~1 of PBS and group I! (n=10) was injected with 1 ~1 of 0.05 M quisqualate. The animals were decapitated at the indicated times after the injec- tions. Half of the animals in each subgroup were decap- itated 1 h after the injections and the remaining animals 7 h after injections.
Experiment 2. For hippocampus injections, the sy- ringe was inserted vertically and implanted 3.3 mm
caudal to Bregma, 2.8 mm lateral (right side) and 3.9 mm ventral to the dura. The rats were divided into two sub- groups where group I (n--7) was injected with 0.5 ~tl of saline and group II (n=8) was injected with 0.5/~1 of 6 mM muscimol in saline. Half of the animals in each sub- group were decapitated 4 h after the injections and the remaining animals 7 h after injections.
All injections were performed during 5 rain. After completion of the infusion the syringe was left in place for an additional 4 rain to allow for passive diffusion of the injected material and to prevent spread of the exo- toxin up the cannula track upon removal, then the needle was slowly retracted.
Tissue sections including hippocampus were cut using a cryostat (Leitz, Wetzlar, FRG) in 16 ¢tm sections and thawed onto poly-L-lysine-coated slides (50/~g/ml). Tis- sue pretreatment and in situ hybridizations were carried out as described earlier [5]. For BDNF mRNA hybridi- zation, a 50-mer oligonucleotide complementary to nu- cleotides 650-699 of rat BDNF mRNA [5] was used. The sections were exposed to X-ray film (Amersham fl-max) for 14 days and to photographic emulsion (Kodak NTB2) for 40 days. After having been devel- oped, the slides were stained with Cresyl violet followed by microscopic analysis. Several criteria for the specifi-

205
city of the hybridizations were used as have been de- scribed earlier [5]. The autoradiograms from the in situ hybridizations were analyzed with Microcomputer Im- aging Device (Imaging Research Inc., Canada). Three tissue sections from each brain were used for measure- ments and optical density in each anatomical region was determined in triplicate in each section. Optical density values were determined and converted to relative levels of mRNA using external standards exposed to the same film.
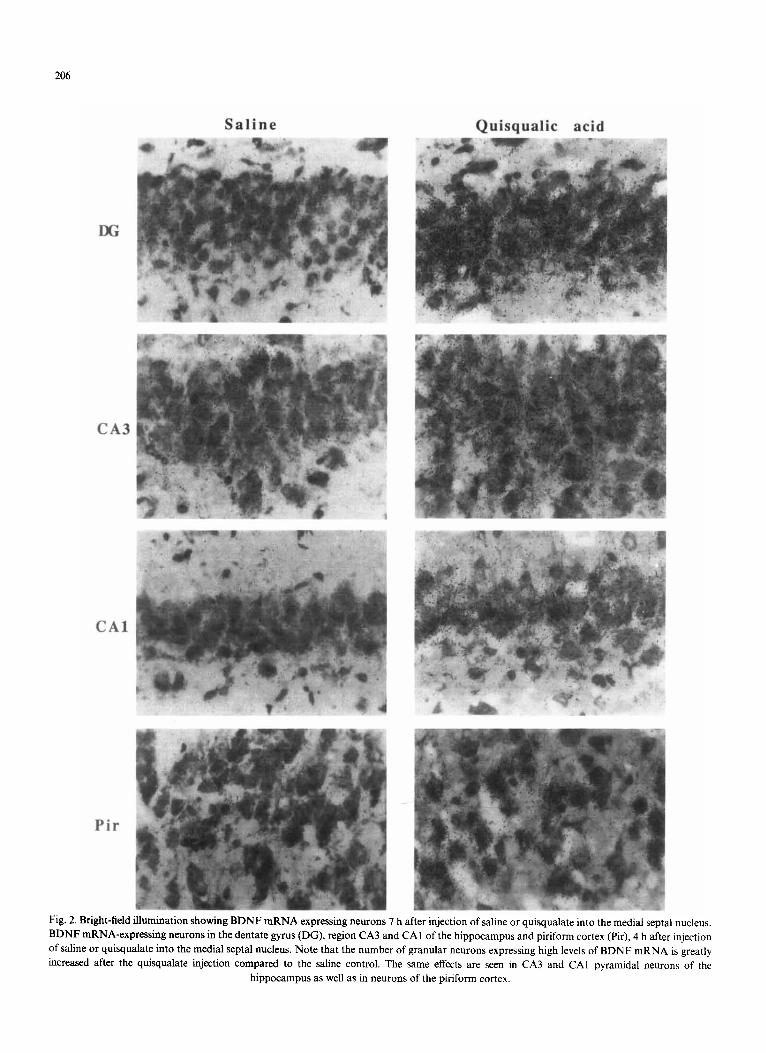
In agreement with earlier reports [5, 9, 19], in situ hy- bridization of coronal sections through the rat brain at the level of the hippocampus revealed BDNF mRNA expressing cells in the dentate gyrus and CA3 regions of the hippocampus with lower levels in CA 1 and CA2 (Fig. I a). BDNF mRNA expressing cells were also seen in the external and internal pyramidal layer of cerebral cortex and in cells of the piriform cortex. The level of BDNF mRNA in these regions did not change 1 or 7 h after an injection of the glutamate analog quisqualate into the nucleus basalis (Fig. lb, showing the result 7 h after the injection). In contrast, injection of quisqualate into the medial septal nucleus caused a marked bilateral increase in the level of BDNF mRNA in the hippocampus 7 h after the injection (Fig. ld). A less but clear increase was also seen 1 h after the injection (data not shown). The increase of BDNF mRNA was seen in both dentate gyrus and throughout the pyramidal layer (CA1 to CA3) of the hippocampus. Computerized image analysis re- vealed a 5-fold increase in dentate gyrus and CA1 and a 3-fold increase in CA3. The injection of quisqualate into the medial septal nucleus also resulted in a 3-fold eleva- tion in the level of BDNF mRNA in piriform cortex (Fig. l d). Analysis of emulsion autoradiographs of the sec- tions showed that the increase of BDNF mRNA was due to both a higher number of neurons expressing de- tectable levels of BDNF mRNA as well as a marked in- crease in the intensity of labeling over individual neurons (Fig. 2). The increase in BDNF mRNA was seen already at 1 h after the injection, reached a maximum at 4 h and remained at this elevated level until 7 h after the injec- tion.
The results presented above indicate that an activation of neurons in the medial septal nucleus causes an in- crease in the expression of BDNF mRNA in both the hippocampus and piriform cortex. In order to test if a stimulation of inhibitory neurotransmission within the hippocampus effects the expression of BDNF mRNA, the GABAA receptor agonist muscimol was injected into the right dorsal hippocampus. In situ hybridization of sections prepared from muscimol injected animals sa- crificed 4 and 7 h after the injection showed levels of BDNF mRNA in the hippocampus, cerebral cortex and
piriform cortex that were comparable to the levels seen in PBS injected control animals (Fig. 3).
The rapid and transient bilateral increase in the level of BDNF mRNA in the dentate gyrus, regions CA1 and CA3 of the hippocampus as well as in the piriform cortex following an injection of quisqualate into medial septum may possibly be due to the increased release of en- dogenous acetylcholine and glutamate that activates BDNF mRNA expressing neurons in the hippocampus. A glutamate-mediated mechanism for increasing BDNF mRNA expression in the hippocampus has also been proposed from experiments that show a marked and transient increase of BDNF mRNA after systemic [22], intraventricular [8] or intrahippocampal [1] injections of kainic acid. Furthermore, stimulation of the glutama- tergic perforant pathway afferents to the hippocampus by a quisqualic acid injection into the entorhinal cortex increases the expression of BDNF mRNA within the hippocampus [16]. In addition, kindling, evoked by elec- trical stimulations in the hippocampus, causing a release of endogenous glutamate has been shown to increase BDNF mRNA expression in the hippocampus, parietal cortex, amygdala and piriform cortex [3]. The involve- ment ofcholinergic afferents in the quisqualate mediated increase of BDNF mRNA after activation of neurons in the medial septum is suggested by the fact that this in- crease is completely blocked in vivo by pretreating the animals with the muscarinic receptor antagonist scopo- lamine [16]. Taken together the studies mentioned above as well as the results presented in this study, suggest that expression of BDNF mRNA in the brain is regulated by neuronal activity possibly via endogenous release of ace- tylcholine and glutamate.
The present study was performed in order to answer two questions in relation to these findings. (i) Will an activation of different afferents in the brain influence the expression of BDNF mRNA in their target areas in dif- ferent ways? (ii) Is the expression of BDNF mRNA within the hippocampus influenced by an enhanced in- hibitory neurotransmission? The basal forebrain con- tains two major cholinergic pathways [17]. The most ro- stromedial cells (medial septum and vertical limb of the diagonal band) project primarily to both sides of the hip- pocampus while more caudal cells (horizontal limb of the diagonal band and nucleus basalis) divide into a separate cell group of each hemisphere and project mostly to the ipsilateral amygdala and to ipsilateral regions of the neo- cortex. Thus, a neuronal activation by injection of quisqualate into either the medial septum or the nucleus basalis, supposedly lead to an increased release of acetyl- choline from afferents in the hippocampus and the neo- cortex, respectively. This system was chosen in order to test if BDNF mRNA expression is differentially regu-
206
S a l i n e Quisqualic acid
DG
CA3
CA1
Pir
Fig. 2. Bright-field illumination showing BDNF mRNA expressing neurons 7 h after injection of saline or quisqualate into the medial septal nucleus. BDNF mRNA-expressing neurons in the dentate gyrus (DG), region CA3 and CAI of the hippocampus and piriform cortex (Pir), 4 h after injection of saline or quisqualate into the medial septal nucleus. Note that the number of granular neurons expressing high levels of BDNF mRNA is greatly increased after the quisqualate injection compared to the saline control. The same effects are seen in CA3 and CA1 pyramidal neurons of the
hippocampus as well as in neurons of the piriform cortex.

h
207
Fig. 3. Effects of an intrahippocampal injection of saline or muscimol on the expression of BDNF mRNA in the hippocampus 7 h after the injections. Adult Sprague-Dawley rats were injected into the right dorsal hippocampus with either saline (a) or muscimol (b). The brains were sectioned 7 h after the injections, and the sections were hybridized to an oligonucleotide probe specific for BDNF mRNA. The injected side is on the right hand side of
the hippocampus.
lated in different projection areas following activation of
basal forebrain cholinergic neurons. In the control, non- stimulated rat brain, B D N F m R N A expressing neurons are found in the dentate gyrus and throughout the pyra- midal layer of the hippocampus notably in region CA3 as well as in the external and internal pyramidal layer of the neocortex and neurons in the amygdaloid complex [5, 9,
11]. Thus, B D N F m R N A expressing neurons are widely distributed in the target areas of basal forebrain choli- nergic neurons. The injection of quisqualate into the
medial septum caused a marked increase of B D N F m R N A in all areas of the hippocampus known to express B D N F m R N A but no effect was seen on B D N F m R N A in the neocortex or amygdala following an injection into the nucleus basalis. A difference in the expression of B D N F m R N A was therefore seen depending on which cholinergic afferents were stimulated suggesting that dif- ferent mechanisms may regulate B D N F m R N A expres- sion in different brain areas. However, it can not be ex- cluded that the lack of stimulation seen following nucleus basalis injections is due to that an insufficient number of cholinergic neurons were activated by the injected quisqualate, since nucleus basalis has a long rostro- caudal extension.
The influence of inhibitory neurotransmission on the expression of B D N F m R N A was tested by an intrahip- pocampal injection of muscimol, a GABAA receptor agonist which binds to most parts of the hippocampal formation [18]. The muscimoi injections had no effect on B D N F m R N A expression 7 h after the injection but in- creased the level of B D N F m R N A to some extent 4 h after the injection. However, as previously reported [1] an increase of B D N F m R N A was also seen at these time points following a similar injection of saline poibibly as a response to the neuronal damage obtained after the needle insertion [1]. The lack of response 7 h after the muscimol injection is in sharp contrast to the marked increase of B D N F m R N A seen 7 h after an intrahippo-
campal injection of kainic acid [1]. This suggests that in contrast to a stimulation of excitatory amino acid re- ceptors, an activation of inhibitory GABAA receptors within the hippocampus does not change the expression of B D N F mRNA.
In conclusion, the present data show that B D N F m R N A expression in the hippocampus is increased after activation of cholinergic afferents from the medial septum but no change is seen in the cholinergic projec- tion areas of the neocortex following an activation in the nucleus basalis. In contrast, to an excitatory stimulus in the hippocampus, an activation of inhibitory GABAA re- ceptors in the hippocampus did not influence B D N F m R N A expression in this brain region. These results in- dicate that B D N F m R N A expression is differentially regulated in different projection areas of basal forebrain cholinergic neurons.
This work was supported by the Swedish National Science Research Council, Petrus and Augusta Hedlunds Stiftelse, US Grants (University of Colorado) A G 04418, the Swedish Medical Research Council (8653) and funds from the Karolinska Institute. M.B. was supported by a C.I.R.I.T. fellowship from the Generalitat de Catalunya (Spain) and P.E. by the Swedish Medical Research Council.
1 Ballarin, M., Ernfors, P., Lindefors, N. and Persson, H., Rapid induction of mRNA for brain derived neurotrophic factor after in- trahippocampal injection of kainic acid, Exp. Neurol., 114 (1991) 35~,3.
2 Barde, Y.-A., Edgar, D. and Thoenen, H., Purification of a new neurotrophic factor from mammalian brain, EMBO J., 1 (1982) 549-553.
3 Ernfors, E, Bengzon, J., Persson, H. and Lindvall, O., Increased levels of messenger RNA for neurotrophic factors in the brain dur- ing kindling epileptogenesis, Neuron, 7 (1991) 165-176.
4 Ernfors, P., Ibanez, C.F., Ebendal, T., Olson, L. and Persson, H., Molecular cloning and neurotrophic activities of a protein with structural similarities to nerve growth factor: developmental and

208
topographical expression in the brain, Proc. Natl. Acad. Sci. USA, 87 (1990) 5454-5458,
5 Ernfors, P., Wetmore, C., Olson, L. and Persson, H., Identification of cells in rat brain and peripheral tissues expressing mRNA for members of the nerve growth factor family, Neuron, 5 (1990) 165 176.
6 Friedman, W.J., Ernfors, P. and Persson, H., In situ hybridization reveals both transient and persistent expression of HDNF/NT-3 mRNA in the rat brain during postnatal development, J. Neurosci., 11(1991)1577 1584.
7 Friedman, W.J., Olson, L. and Persson, H., Brain-derived neuro- trophic factor mRNA expressing cells in the developing postnatal brain, Eur. J. Neurosci., 3 (1991) 688 697.
8 Gall, C., Murray, K. and J, I.P., Kainic acid-induced seizures stimu- late increased expression of nerve growth factor mRNA in rat hip- pocampus, Mol, Brain Res., 9 (1991) 113 123.
9 Hofer, M., Pagliusi, S.R., Hohn, A,, Leibrock, J. and Barde, Y.A., Regional distribution of brain-derived neurotrophic factor mRNA in the adult mouse brain, EMBO J., 9 (1990) 2459 -2464.
10 Hofer, M.M. and Barde, Y.-A., Brain-derived neurotrophic factor prevents neuronal death in vivo, Nature, 331 (1988) 261 262.
11 lsackson, EJ., Huntsman, M.M., Murray, K.D. and M, G.C., BDNF mRNA expression is increased in adult forebrain after limbic seizures: Temporal patterns of induction distinct from NGF, Neuron, (1991) 937-948.
12 Johnson, J.E., Barde, Y.-A., Schwab, M. and Thoenen, H., Brain- derived neurotrophic factor supports the survival of cultured rat retinal ganglion cells, J, Neurosci., 6 (1986) 3031 3038.
13 Leibrock, J., kottspeich, F., Hohn, A., Hofer, M., Hengerer, B., Masiakowski, P., Thoenen, H. and Barde, Y.A., Molecular cloning
and expression of brain-derived neurotrophic factor, Nature, 341 (1989).
14 Levi-Montalcini, R., The nerve growth factor 35 years later, Science, 237 (1987) 1154-1162.
15 Levi-Montalcini, R. and Angeletti, EU., Nerve growth factor, Physiol. Rev., 48 (1968) 534-569.
16 Lindefors, N., Ernfors, P., Falkenberg, T. and Persson, H., Septal cholinergic afferents regulate expression of brain-derived neuro- trophic factor and B-nerve growth factor mRNA in rat hip- pocampus, Exp. Brain Res., in press.
17 McGeer, EL., McGeer, E.G. and Peng, J.H., Choline acetyl- transferase: Purification and immunohistochemical localization, Life Sci., 34 (1984) 2319-2338.
18 Penney, J,B., Pan, H.S., Young, A.B., Frey, K.A. and Dauth, G.W., Quantitative autoradiography of (3H)-muscimol binding in the rat brain, Science, 214 (1981) 1036 1038.
19 Phillips, H.S., Hains, J.M., Laramee, G.R., Rosenthal, A. and Winslow, J.W., Widespread expression of BDNF but not NT-3 by target areas of basal forebrain cholinergic neurons, Science, 250 (1990) 290-294.
20 Thoenen, H. and Barde, Y.A., Physiology of nerve growth factor, Physiol. Rev., 60 (1980) 1284 1335.
21 Wetmore, C., Ernfors, P,, Persson, H. and Olson, L., Localization of brain-derived neurotrophic factor mRNA to neurons in the brain by in situ hybridization, Exp. Neurol., 109 (1990) 141 152.
22 Zafra, F., Hengerer, B., Leibrock, J., Thoenen, H. and Lindholm, D., Activity dependent regulation of BDNF and NGF mRNAs in the rat hippocampus is mediated by non-NMDA glutamate re- ceptors, EMBO J., 9 (1990) 3545 3550.