accumulation of il-12-activated antitumor effector cells into lymph nodes of tumor-bearing mice
TRANSCRIPT

Immunology Letters 61 (1998) 127–133
Accumulation of IL-12-activated antitumor effector cells into lymphnodes of tumor-bearing mice
Takashi Yahata a, Kazuhito Watanabe a, Akio Ohta a, Yasushi Ohmi a, Naoko Sato a,Kazuki Santa a, Naoko Abe a, Kenji Iwakabe a, Satoshi Kaneko a, Norihiro Suzuki a,
Sonoko Habu a, Shuhei Yasuda b, Takashi Nishimura a,*a Department of Immunology, Tokai Uni6ersity School of Medicine, Bohseidai, Isehara 259-11, Japan
b Department of Clinical Pharmacology, Yamanouchi Pharmaceutical, Tsukuba 305, Japan
Received 19 November 1997; received in revised form 11 December 1997; accepted 7 January 1998
Abstract
Simultaneous administration of high dose of IL-12 into tumor-inoculated mice resulted in a marked reduction of tumor growthin parallel with the augmented generation of cytotoxic T-cells, natural killer (NK) cells and IFN-g-producing Th cells. We foundthat these IL-12-activated antitumor effector cells preferentially accumulated in peripheral lymph nodes concomitantly withlymphadenopathy. However, IL-12 rather induced disappearance of antitumor effector cells including CD4+ T, CD8+ T and NKcells from spleen in spite of inducing splenomegaly. Lymph node cells obtained from IL-12-treated B16F0-bearing mice showeda marked IFN-g production in response to not only IL-2, IL-12, anti CD3 mAb but also B16F0 melanoma cells. Moreover, theycould lyse B16F0 melanoma cells in a long-term cytotoxicity assay. It was also confirmed that IL-12-activated IFN-g producingTh1 cells were accumulated in tumor local site. Thus, IL-12 appeared to have a capability of stimulating selective migration ofantitumor cells into lymph nodes and tumor local sites. © 1998 Elsevier Science B.V. All rights reserved.
Keywords: IL-12; Th1/Th2; Tumor; Mouse
1. Introduction
IL-12 consisting of p35 and p40 heterodimericproteins has been demonstrated to show a therapeuticactivity in various immune disease models [1,2]. Inparticular, it has been clearly demonstrated that IL-12showed a potent antitumor activity against both pri-mary and transplantable tumors [3–7]. The evidencethat IL-12 can stimulate the induction of human antitu-mor effector cells in vitro [8], indicates its rationale forthe application to cancer immunotherapy.
Many investigators have demonstrated that IL-12showed its therapeutic effect through the activation of
both antitumor killer cells and Th1 cells [9–11]. Be-cause Th1 cells, which can produce IL-2 and IFN-g, areconsidered as pivotal immunoregulatory cells in cellularimmunity [2,12,13], the selective activation of Th1 cellsby IL-12 may be a key mechanism for the induction ofantitumor immunity in vivo. In addition to the activa-tion of these antitimor effector cells, IL-12 has beendemonstrated to show other multiple functions in vivoconcerning angiogenesis, lymphocyte migration and he-matopoiesis [14–16].
In this paper, we investigated the effect of IL-12 onthe activation of antitumor effector cells and theirdistribution pattern in tumor-bearing mouse spleen,lymph nodes and tumor tissue. The present data clearlydemonstrated that IL-12-activated antitumor effectorcells selectively accumulated into lymph nodes andtumor local site, while they rather decreased in spleen.
* Corresponding author. Tel.: +81 463 931121; fax: +81 463942976.
0165-2478/98/$19.00 © 1998 Elsevier Science B.V. All rights reserved.PII S0165-2478(98)00006-6

T. Yahata et al. / Immunology Letters 61 (1998) 127–133128
2. Materials and methods
2.1. Treatment of tumor-bearing mice with IL-12
C57BL/6 mice were purchased from Charles RiverJapan (Yokohama 222, Japan). All mice were femaleand 5–8 weeks old. Generally, C57BL/6 mice weresubcutaneously (s.c.) injected with B16F0 melanomacells (106/mouse) and the tumor-bearing mice weretreated with 30 mg/kg recombinant mouse IL-12(kindly donated by Genetics Institute, MA) threetimes at intervals of 2 days. At 7–10 days after IL-12 treatment, the mice were sacrificed and examined.Their immunological parameters are described in thefollowing sections. Tumor weight was measured at 7days after tumor-inoculation.
2.2. Cytotoxicity assay
The cytotoxic activity of NK cells and T-cells weredetermined by 4 h 51Cr-release assay. NK activitywas determined using YAC-1 cells as target-cells.The specific CTL activity of B16F0-bearing mice wasdetermined using 51Cr-labeled B16F0 cells and non-specific CTL activity was measured by reverse anti-body-dependent cellular cytotoxicity (RADCC)assay using 51Cr-labeled P815 cells in the presence ofanti-mouse CD3 (2C11, 10 mg/ml) mAb. The % cy-totoxicity was calculated as described previously[17].
2.3. Phenotypic analysis and separation of lymphocytesby flow cytometry
The analysis of cell surface markers was carriedout by FACScan using the Consort 30 program. Insome experiments, CD4+ T-cells and CD8+ T-cellswere isolated by FACStar after staining with FITC-conjugated anti-CD4 mAb and PE-conjugated anti-CD8 mAb. The detailed procedure for staining andsorting was described in a previous paper [18].All FITC or PE-conjugated mAbs used in thispaper was purchased from PharMingen (San Diego,CA).
2.4. Cytokine production
Spleen cells and lymph node cells (5×105/well)were stimulated with soluble anti-CD3 mAb (10 mg/ml), IL-2, IL-12, or B16F0 tumor cells (104 cells)for 36 h in Falcon 96 well plate. After incubation,culture supernatants were harvested and theirIFN-g activity was determined by RPN2717mouse IFN-g ELISA kits (Amersham Tokyo, Japan)[19].
3. Results
3.1. Induction of splenomegaly and lymphadenopathyby administration of IL-12
C57BL/6 mice bearing B16F0 melanoma cells weretreated with i.p. injection of IL-12 (30 mg/kg) threetimes, at intervals of 2 days. Like previous results,IL-12 showed a marked antitumor effect in vivo andinhibited tumor growth (Fig. 1). A total of 8 days afterIL-12 administration, the tumor-bearing mice were sac-rificed to examine the effect of IL-12 on variouslymphoid tissues (thymus, axillary and inguinal lymphnodes and spleen). As control, the tissues from saline-treated mice were used. A typical changes of thymus,lymph nodes and spleen obtained from saline-treatedmice and IL-12-treated mice are shown in the photo-graphs (Fig. 2AB). IL-12 administration induced amarked splenomegaly and lymphadenopathy, though itrather induced thymic atrophy. As shown in Fig. 2C,the tissue weight and the number of cells in both lymphnodes and spleen also greatly increased in IL-12-treatedmice compared with those in saline-treated mice (3–4-fold increase in lymph node; 5–6-fold increase inspleen). However, the tissue weight and the number ofcells rather decreased in the thymus from IL-12-treatedmice.
Fig. 1. Inhibition of tumor-growth in vivo by IL-12 administration.B16F0 melanoma cells (106 cells) were i.d. injected into abdominalskin of C57BL/6 mice. A total of 3 days after tumor inoculation,various doses of IL-12 (0, 0.3, 1, 3, 10 and 30 mg/kg) were i.p. injectedinto the mice three times at 2 day intervals. Then, 7 days after tumorinoculation, tumor masses were removed and their weight was mea-sured. The bars represent mean9SE of five mice.
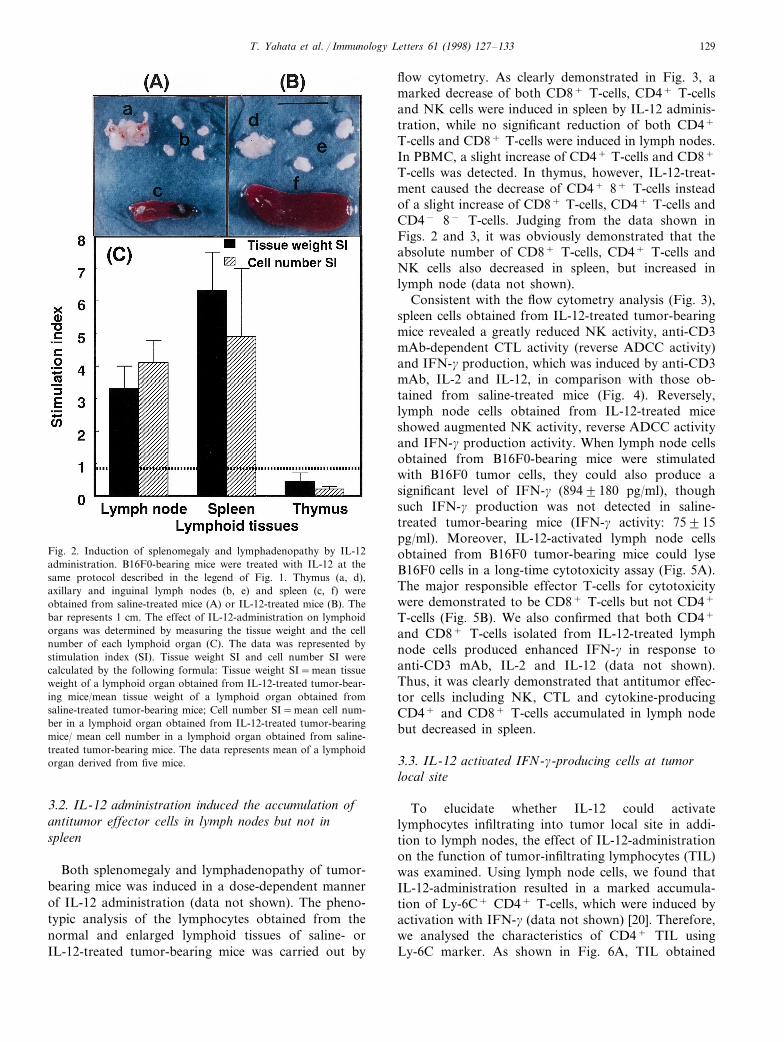
T. Yahata et al. / Immunology Letters 61 (1998) 127–133 129
Fig. 2. Induction of splenomegaly and lymphadenopathy by IL-12administration. B16F0-bearing mice were treated with IL-12 at thesame protocol described in the legend of Fig. 1. Thymus (a, d),axillary and inguinal lymph nodes (b, e) and spleen (c, f) wereobtained from saline-treated mice (A) or IL-12-treated mice (B). Thebar represents 1 cm. The effect of IL-12-administration on lymphoidorgans was determined by measuring the tissue weight and the cellnumber of each lymphoid organ (C). The data was represented bystimulation index (SI). Tissue weight SI and cell number SI werecalculated by the following formula: Tissue weight SI=mean tissueweight of a lymphoid organ obtained from IL-12-treated tumor-bear-ing mice/mean tissue weight of a lymphoid organ obtained fromsaline-treated tumor-bearing mice; Cell number SI=mean cell num-ber in a lymphoid organ obtained from IL-12-treated tumor-bearingmice/ mean cell number in a lymphoid organ obtained from saline-treated tumor-bearing mice. The data represents mean of a lymphoidorgan derived from five mice.
flow cytometry. As clearly demonstrated in Fig. 3, amarked decrease of both CD8+ T-cells, CD4+ T-cellsand NK cells were induced in spleen by IL-12 adminis-tration, while no significant reduction of both CD4+
T-cells and CD8+ T-cells were induced in lymph nodes.In PBMC, a slight increase of CD4+ T-cells and CD8+
T-cells was detected. In thymus, however, IL-12-treat-ment caused the decrease of CD4+ 8+ T-cells insteadof a slight increase of CD8+ T-cells, CD4+ T-cells andCD4− 8− T-cells. Judging from the data shown inFigs. 2 and 3, it was obviously demonstrated that theabsolute number of CD8+ T-cells, CD4+ T-cells andNK cells also decreased in spleen, but increased inlymph node (data not shown).
Consistent with the flow cytometry analysis (Fig. 3),spleen cells obtained from IL-12-treated tumor-bearingmice revealed a greatly reduced NK activity, anti-CD3mAb-dependent CTL activity (reverse ADCC activity)and IFN-g production, which was induced by anti-CD3mAb, IL-2 and IL-12, in comparison with those ob-tained from saline-treated mice (Fig. 4). Reversely,lymph node cells obtained from IL-12-treated miceshowed augmented NK activity, reverse ADCC activityand IFN-g production activity. When lymph node cellsobtained from B16F0-bearing mice were stimulatedwith B16F0 tumor cells, they could also produce asignificant level of IFN-g (8949180 pg/ml), thoughsuch IFN-g production was not detected in saline-treated tumor-bearing mice (IFN-g activity: 75915pg/ml). Moreover, IL-12-activated lymph node cellsobtained from B16F0 tumor-bearing mice could lyseB16F0 cells in a long-time cytotoxicity assay (Fig. 5A).The major responsible effector T-cells for cytotoxicitywere demonstrated to be CD8+ T-cells but not CD4+
T-cells (Fig. 5B). We also confirmed that both CD4+
and CD8+ T-cells isolated from IL-12-treated lymphnode cells produced enhanced IFN-g in response toanti-CD3 mAb, IL-2 and IL-12 (data not shown).Thus, it was clearly demonstrated that antitumor effec-tor cells including NK, CTL and cytokine-producingCD4+ and CD8+ T-cells accumulated in lymph nodebut decreased in spleen.
3.3. IL-12 acti6ated IFN-g-producing cells at tumorlocal site
To elucidate whether IL-12 could activatelymphocytes infiltrating into tumor local site in addi-tion to lymph nodes, the effect of IL-12-administrationon the function of tumor-infiltrating lymphocytes (TIL)was examined. Using lymph node cells, we found thatIL-12-administration resulted in a marked accumula-tion of Ly-6C+ CD4+ T-cells, which were induced byactivation with IFN-g (data not shown) [20]. Therefore,we analysed the characteristics of CD4+ TIL usingLy-6C marker. As shown in Fig. 6A, TIL obtained
3.2. IL-12 administration induced the accumulation ofantitumor effector cells in lymph nodes but not inspleen
Both splenomegaly and lymphadenopathy of tumor-bearing mice was induced in a dose-dependent mannerof IL-12 administration (data not shown). The pheno-typic analysis of the lymphocytes obtained from thenormal and enlarged lymphoid tissues of saline- orIL-12-treated tumor-bearing mice was carried out by

T. Yahata et al. / Immunology Letters 61 (1998) 127–133130
from IL-12-treated tumor-bearing mice containedhigher frequency (50.4%) of activated CD4+ T-cellsexpressing Ly-6C antigen compared with TIL fromsaline-treated tumor-bearing mice (20.8%). Moreover,TIL obtained from tumor-bearing mice produced highlevels of IFN-g, while IL-4 production was markedlyreduced compared with those in TIL obtained fromcontrol mice (Fig. 6B). Thus, IL-12 appeared to acti-vate both lymph node cells and TIL, which facilitatedTh1-dominant immunity.
4. Discussion
It is now well accepted that IL-12 is a useful cytokinefor the application to therapy of immune diseases in-cluding tumor, infectious diseases, allergy and autoim-mune diseases [3–8,21]. This multifunction of IL-12 ismediated by the activation of Th1-dominant immunitywhich is essential for the induction of cell-mediatedimmunity [2,12,13]. The present paper further demon-strates an interesting effect of IL-12 administration invivo on the distribution of antitumor effector cells intumor bearing mice.
Consistent with previous data [7], a single injection ofIL-12 into the mice caused enhancement of NK, CTLand IFN-g producing T-cells in the spleen (data notshown), but simultaneous administration of IL-12 at ahigh dose (\10 mg/kg) rather induced suppression ofantitumor effector T-cell generation in the spleen. This
is because of the disappearance of CD4+ T-cells,CD8+ T-cells and NK cells from the spleen (Fig. 3).However, interestingly, the antitumor effector cells suchas NK cells, CTL and IFN-g producing Th cells accu-mulate into peripheral lymph nodes (Figs. 4 and 5). Asshown in Fig. 6, IL-12-activated antitumor effector cellsalso accumulated in tumor-local sites. The precisemechanisms of IL-12-induced accumulation of acti-vated cells into peripheral lymph nodes remains un-clear. However, recently, it is reported that Th1 cellsare prone to migrate into peripheral lymph nodes whichpreferentially express addressin for Th1 cells [22]. Wealso demonstrated that lymph node cells obtained fromIL-12-administered mice show a markedly increasedLFA-1-dependent-cell aggregation [23] compared withlymph node cells obtained from saline-treated mice(data not shown). Therefore, it is suggested that theupregulation of adhesion molecules including LFA-1,ICAM-1 and other tissue specific putative homing re-ceptors on IL-12-activated cell populations may be themajor cause of the augmented migration of IL-12-acti-vated cells into peripheral lymph nodes via vascularendothelium. Such accelerated migration by IL-12 ofperipherally circulated T-cells into lymph nodes mayinduce putative leukopenia as described previously [15].However, this may not be serious for IL-12-treatedmice, because the decrease of lymphocyte number is notdue to the deletion of lymphocytes but because of theaccumulation of lymphocytes into lymph nodes. In-deed, here, we confirm that the treatment of tumor-
Fig. 3. Disappearance of CD4+ T-cells, CD8+ T-cells and NK cells from spleen of tumor-bearing mice by IL-12 administration in vivo. Spleencells (A, E, F, J), lymph node cells (B, G), thymocytes (C, H) and peripheral blood mononuclear cells (D, I) were prepared from saline- (A–E)or IL-12-treated B16F0-bearing mice (F–J). The cells were stained with PE-conjugated anti-CD8 mAb plus FITC-conjugated anti-CD4 mAb todetermine the effect of IL-12 on CD4+ T-cells and CD8+ T-cells. The existence of NK cells was determined by staining with PE-conjugatedanti-NK1.1 mAb plus FITC-conjugated anti-TCRgd mAb. The staining profile was analyzed by FACScan. The figures in the FACScan profilesrepresent mean9SE of positive cell percentage in three separate experiments. Three mice per group were used.
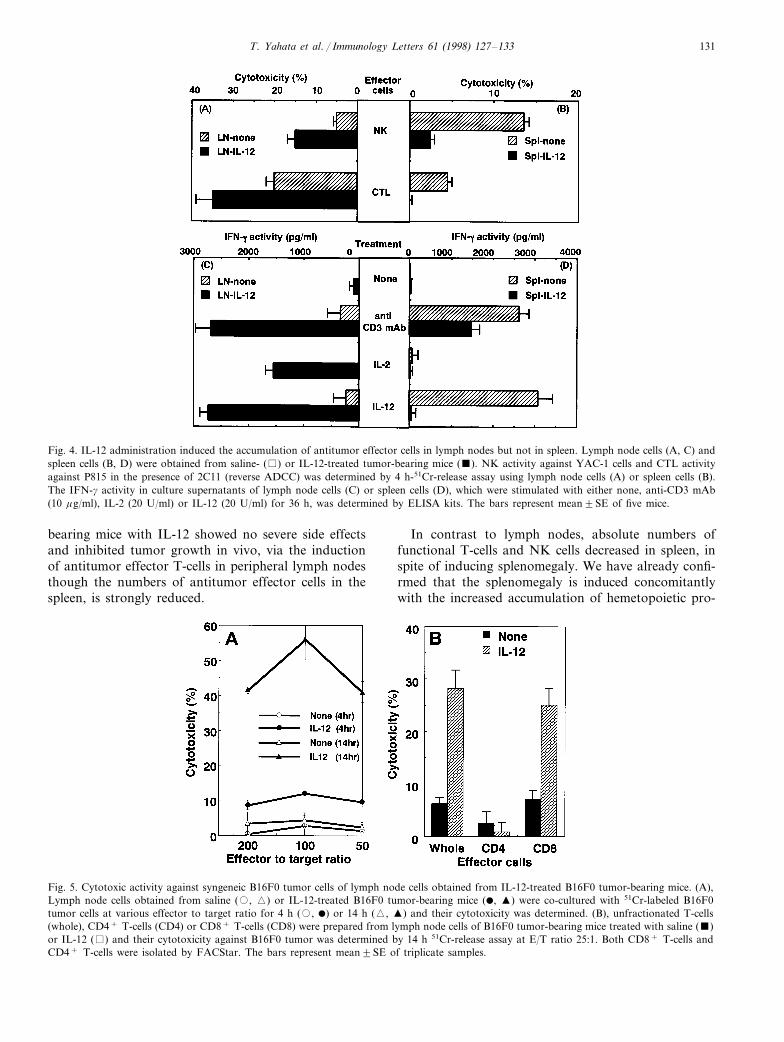
T. Yahata et al. / Immunology Letters 61 (1998) 127–133 131
Fig. 4. IL-12 administration induced the accumulation of antitumor effector cells in lymph nodes but not in spleen. Lymph node cells (A, C) andspleen cells (B, D) were obtained from saline- ( ) or IL-12-treated tumor-bearing mice (). NK activity against YAC-1 cells and CTL activityagainst P815 in the presence of 2C11 (reverse ADCC) was determined by 4 h-51Cr-release assay using lymph node cells (A) or spleen cells (B).The IFN-g activity in culture supernatants of lymph node cells (C) or spleen cells (D), which were stimulated with either none, anti-CD3 mAb(10 mg/ml), IL-2 (20 U/ml) or IL-12 (20 U/ml) for 36 h, was determined by ELISA kits. The bars represent mean9SE of five mice.
bearing mice with IL-12 showed no severe side effectsand inhibited tumor growth in vivo, via the inductionof antitumor effector T-cells in peripheral lymph nodesthough the numbers of antitumor effector cells in thespleen, is strongly reduced.
In contrast to lymph nodes, absolute numbers offunctional T-cells and NK cells decreased in spleen, inspite of inducing splenomegaly. We have already confi-rmed that the splenomegaly is induced concomitantlywith the increased accumulation of hemetopoietic pro-
Fig. 5. Cytotoxic activity against syngeneic B16F0 tumor cells of lymph node cells obtained from IL-12-treated B16F0 tumor-bearing mice. (A),Lymph node cells obtained from saline (�, �) or IL-12-treated B16F0 tumor-bearing mice (�, �) were co-cultured with 51Cr-labeled B16F0tumor cells at various effector to target ratio for 4 h (�, �) or 14 h (�, �) and their cytotoxicity was determined. (B), unfractionated T-cells(whole), CD4+ T-cells (CD4) or CD8+ T-cells (CD8) were prepared from lymph node cells of B16F0 tumor-bearing mice treated with saline ()or IL-12 ( ) and their cytotoxicity against B16F0 tumor was determined by 14 h 51Cr-release assay at E/T ratio 25:1. Both CD8+ T-cells andCD4+ T-cells were isolated by FACStar. The bars represent mean9SE of triplicate samples.

T. Yahata et al. / Immunology Letters 61 (1998) 127–133132
Fig. 6. Accumulation of IL-12-activated IFN-g producing CD4+ T-cells into the tumor tissues. Lymph node cells (A, B) or TIL (C, D, E) wereobtained from saline- (A, C) or IL-12-treated tumor-bearing mice (B, D) using five mice per group. The existence of activated CD4+ T-cells wasdetermined by staining with PE-conjugated anti-CD4 mAb plus FITC-conjugated anti-Ly-6C mAb using pooled lymph node cells or TIL. The% of activated CD4+ Ly-6C+ T-cells in total CD4+ T-cells was 38.7, 59.3, 20.8 and 50.4% in A, B, C and D, respectively. The IFN-g ( ) andIL-4 () producing ability of TIL obtained from saline- (none) or IL-12-treated tumor-bearing mice (IL-12) was determined by stimulating withimmobilized anti-CD3 mAb for 36 h (E). The IFN-g activity in culture supernatants was measured by ELISA kit. The data represent mean9SEof five mice.
genitor cells expressing c-Kit and Sca-1 antigens (datanot shown). This is consistent with the recent findingthat IL-12 induces the proliferation of hematopoieticprogenitor cells both in vitro and in vivo [15,16,24].Therefore, IL-12 administration in vivo appears to in-duce ‘habitat segregation’ of antitumor effector cellsand hematopoietic progenitor cells in lymph nodes andspleen. Here, we reported the results obtained fromtumor-bearing mice, but the same results were demon-strated in IL-12-injected tumor-free normal mice.Therefore, IL-12 can induce the augmented accumula-tion of activated lymphocyte in lymph nodes and thatof hematopoietic progenitor cells in spleen without theinfluence of tumor burden.
Thus, the present finding that IL-12-activated antitu-mor effector cells accumulate in lymph nodes and TIL,strongly suggests a useful role for lymph node cells andTIL after IL-12 administration in vivo. Moreover, itprovides some warning that too much surgical resectionof lymph nodes near the tumor local site may not bepreferable to induce IL-12-induced tumor immuno-therapy.
Acknowledgements
The authors would like to thank Drs MichikoKobayashi and Steven H. Herrmann for their kind giftof IL-12. This work was supported in part by a Grant-in-Aid for The Science Frontier Program of MESC (the
Ministry of Education Science and Culture, Japan), aGrant-in-Aid for Scientific Research on Priority Areasfrom MESC, and a Grant-in-Aid from the Ministry ofHealth and Welfare for Cancer Control, and by aGrant-in-Aid for the IL-12 project of Tokai UniversitySchool of Medicine.
References
[1] M. Kobayashi, L. Fitz, M. Ryan, R.M. Hewick, S.C. Clark, S.Chan, R. Loudon, F. Sherman, B. Perussia, G. Trinchieri, J. Exp.Med. 170 (1989) 827.
[2] G. Trinchieri, Blood 84 (1994) 4008.[3] M.J. Brunda, L. Luistro, R.R. Warrier, R.B. Wright, B.R.
Hubbard, M. Murphy, S.F. Wolf, M.K. Gately, J. Exp. Med. 178(1993) 1223.
[4] H. Tahara, H.J. Zer III, W.J. Storkus, I. Pappo, S.C. Watkins,U. Gubler, S.F. Walf, P.D. Robbins, M.T. Lotze, Cancer Res. 54(1994) 182.
[5] J.-P. Zou, T. Yamamoto, H. Fujii, M. Takenaka, M. Kobayashi,S.H. Herrmann, S.F. Wolf, H. Fujiwara, T. Hamaoka, Int.Immunol. 7 (1995) 1135.
[6] T. Nishimura, K. Watanabe, U. Lee, T. Yahata, M. Kobayashi,S.H. Herrmann, S. Habu, Immunol. Lett. 48 (1995) 149.
[7] T. Nishimura, K. Watanabe, T. Yahata, U. Lee, K. Ando, M.Kimura, I. Saiki, T. Uede, S. Habu, Cancer Chemother. Pharma-col. 38 (1996) 27.
[8] S. Kuge, K. Watanabe, K. Makino, Y. Tokuda, T. Mitomi, N.Kawamura, S. Habu, T. Nishimura, Jpn. J. Cancer Res. 86 (1995)135.
[9] C.L. Nastla, H.D. Edington, T.G. Mckinney, H. Tahara, M.A.Nalensnik, M.J. Brunda, M.K. Gately, S.F. Wolf, R.D. Schreiber,W.J. Strkus, M.T. Lotze, J. Immunol. 153 (1994) 1697..

T. Yahata et al. / Immunology Letters 61 (1998) 127–133 133
[10] Y. Noguchi, E.C. Richards, Y.-T. Chen, L.J. Old, Proc. Natl.Acad. Sci. 92 (1995) 2219.
[11] M.K. Gately, R.R. Warrier, S. Honasoge, D.M. Carvajal, D.A.Faherty, Int. Immunol. 6 (1994) 157.
[12] T.R. Mosmann, S. Sad, Immunol. Today 17 (1996) 138.[13] C.S. Hsieh, S.E. Macatonica, C.S. Tripp, S.F. Wolf, A. O’Garra,
K.M. Murphy, Science 260 (1993) 547.[14] A.L. Angiollilo, C. Sgadari, T. Giocanna, Ann. New York Acad.
Sci. 795 (1996) 158.[15] N.S. Tare, S. Bowen, R.R. Warrier, D.M. Carvajal, W.R. Ben-
jamin, J.H. Riley, T.D. Anderson, M. Gately, J. InterferonCytokine Res. 15 (1995) 377.
[16] J. Jackson, Y. Yan, M.J. Brunda, L.S. Kelsey, E. Talmadge,Blood 85 (1995) 2371.
[17] T. Nishimura, S.J. Burakoff, S.H. Herrmann, J. Immunol. 139(1987) 2888.
[18] T. Nishimura, Y. Takeuchi, Y. Ichimura, X. Gao, A. Akatsuka,N. Tamaoki, H. Yagita, K. Okumura, S. Habu, J. Immunol. 145(1990) 4012.
[19] Y. Tanaka, A. Takahashi, K. Watanabe, K. Takayama, T.Yahata, S. Habu, T. Nishimura, Int. Immunol. 8 (1996) 569.
[20] F.J. Dumont, R.G.E. Palfree, L.Z. Coker, J. Immunol. 137(1986) 201.
[21] M.T. Lotze, G. Trinchieri, M. Gately, S. Wolf (Eds.), Interleukin12: Cellular and Molecular Immunology of Important Regula-tory Cytokine. Ann. New York Acad. Sci. 795 (1996) 1.
[22] E.N.T. Meeusen, R.R. Premier, M.R. Brandon, Immunol. To-day 17 (1996) 421.
[23] T. Nishimura, Y. Takeuchi, K. Urano, S. Habu, J. Immunol.147 (1991) 2094.
[24] I. Dybedal, S. Larsen, E.W. Jacobsen, J. Immunol. 154 (1995)4950.
.
.