abundancia de microalgas nocivas y condiciones ... · el servicio nacional de pesca y acuicultura...
TRANSCRIPT

1
INFORME FINAL
ABUNDANCIA DE MICROALGAS NOCIVAS Y CONDICIONES
OCEANOGRAFICAS EN EL ÁREA DE EMERGENCIA DE PLAGA
(SECTOR GOLFO DE PENAS, REGIÓN DE AYSÉN) Y ZONAS
ALEDAÑAS
PROYECTO EJECUTADO POR:
CENTRO INTERDISCIPLINARIO PARA LA INVESTIGACIÓN
ACUÍCOLA (INCAR)
(CONICYT-FONDAP Nº15110027)
POR ENCARGO DE:
SERVICIO NACIONAL DE PESCA Y ACUICULTURA
(SERNAPESCA)
-Julio de 2017-

2
EQUIPO DE INVESTIGADORES
Jefe de Proyecto
Dr. Renato Quiñones
Investigadores
Dra. (c) Valeria Anabalón
Dr. Renato Quiñones
Dr. Marcelo Fuentes
Dr. Rodrigo Veas
M.Sc. Luis Montecinos
Biol. Mar. Wilfredo Alarcón
Biol. Mar. Alvaro Araneda
ESTE INFORME DEBE SER CITADO DE LA SIGUIENTE MANERA:
Anabalón, V., Quiñones, R.A., Fuentes, M.E., Montecinos, L. 2017. Abundancia de
microalgas nocivas y condiciones oceanográficas en el área de emergencia de plaga (sector
Golfo de Penas, Región de Aysén) y zonas aledañas. Informe Final. Estudio encargado por
el Servicio Nacional de Pesca y Acuicultura (SERNAPESCA). Centro Interdisciplinario para
la Investigación Acuícola (INCAR; CONICYT-FONDAP Nº15110027), Universidad de
Concepción, Chile 101 páginas.
Contacto: Dr. Renato Quiñones, Centro Interdisciplinario para la Investigación Acuícola
(INCAR), Universidad de Concepción, O’Higgins 1695, Concepción. Fono: (41) 2661499.
e-mail: [email protected]

3
INDICE
Portada_______________________________________________________________1
I RESUMEN EJECUTIVO_______________________________________________4
II IINTRODUCCION____________________________________________________7
III METODOLOGIA____________________________________________________9
3.1 Muestreo durante el Crucero Golfo De Penas______________________________9
3.2 Análisis estadísticos_________________________________________________13
IV RESULTADOS_____________________________________________________16
4.1 Condiciones Meteorológicas y Oceanográficas____________________________16
4.2 Clorofila-a y Feopigmentos___________________________________________29
4.2 Nutrientes_________________________________________________________35
4.3. Composición de Especies____________________________________________39
4.4 Resultados de análisis estadísticos______________________________________57
V DISCUSION GENERAL______________________________________________76
VI CONCLUSIONES___________________________________________________87
VII AGRADECIMIENTOS______________________________________________89
VIII REFERENCIAS___________________________________________________90

4
I RESUMEN EJECUTIVO
El 2 de Febrero de 2017, SERNAPESCA tuvo conocimiento de 3 eventos significativos de
mortalidad de peces ocurridos durante su transporte marítimo entre la Región de Los Lagos
y la Región de Magallanes, asociadas al paso de estas naves en las cercanías del Golfo de
Penas, en el sur de la Región de Aysén. Ante esta situación, SERNAPESCA procedió a
declarar “Emergencia de Plaga” el 10 de febrero de 2017 y organizó un crucero científico de
prospección en el buque "Marinero Fuentealba" (Armada de Chile), con el fin de estudiar la
presencia de microalgas productoras de floraciones algales nocivas (FAN) en la zona del
Golfo de Penas y sectores aledaños. El muestreo y análisis de las FAN de este crucero de
prospección fue encargado al Centro Interdisciplinario de Investigación Acuícola (INCAR).
Los objetivos del estudio encargado al INCAR por SERNAPESCA son:
(i) Confirmar la presencia de microalgas nocivas en la zona del Golfo de Penas y
fiordos/canales adyacentes y en especial del dinoflagelado Karenia mikimotoi.
(ii) Identificar cuantitativamente la abundancia de las microalgas productoras de floraciones
algales nocivas en la zona de estudio.
(iii) Caracterizar las condiciones oceanográficas de la zona de estudio y las posibles
asociaciones entre dichas condiciones y la presencia de floraciones algales nocivas.
Durante el crucero oceanográfico, se muestrearon 21 estaciones en el Golfo de Penas y
sectores aledaños, entre los días 17 y 21 de febrero de 2017. La cobertura espacial
comprendió desde la entrada norte del Canal Moraleda (44°32'09" °S) hasta la salida sur del
Canal Wide (49°57'51" °S) y desde el punto de vista longitudinal, entre 73°28'07" y
75°44'00" °W. Las variables monitoreadas en el crucero fueron: (a) Presencia/ausencia y
cuantificación de Karenia en 3 profundidades de cada estación oceanográfica (5, 15 y 30 metros;
se tomaron muestras concentradas [20 m] y no concentradas); (b) Presencia/ausencia y
cuantificación de otras especies que generan floraciones nocivas en 3 profundidades de cada estación
oceanográfica (5, 15 y 30 metros); (c) Perfiles verticales de temperatura, salinidad y oxígeno (0-30

5
metros); (d) Mediciones de Sólidos disueltos totales (SDT), pH y REDOX en cada una de las 3
profundidades muestreadas; (e) Concentración de nutrientes (nitrato, nitrito, fosfato, silicato) en 3
profundidades de cada estación oceanográfica (5, 15 y 30 metros); (f) Concentración de clorofila-a
y feopigmentos en 3 profundidades de cada estación oceanográfica (5, 15 y 30 metros). Se
realizaron, además, mediciones de clorofila-a fraccionada por tamaño (total y <20 m); (g)
Estimación de la transparencia y de la profundidad de la zona eufótica mediante el uso de Disco
Secchi en cada estación oceanográfica.
El presente Informe Final, entrega los resultados provenientes del cumplimiento de cada
objetivo del estudio. Las principales conclusiones del estudio son:
1) Se confirma la presencia de Karenia mikimotoi en la zona de estudio, aunque presenta baja
abundancia, por lo que es posible que el muestreo se haya realizado en condiciones de post-
bloom, en especial considerando que los eventos de mortalidad de peces ocurrieron
aproximadamente 15 días antes del muestreo a bordo del Buque “Marinero Fuentealba”.
2) La presencia de varias especies de dinoflagelados y diatomeas productoras de toxinas en
la zona de estudio, entre ellas Karenia mikimotoi, permite hipotetizar que la intoxicación de
peces podría deberse a un efecto sinérgico de un conjunto de toxinas.
3) Se confirma la presencia de otras especies productoras de toxinas en la zona de estudio,
principalmente Gymnodinium spp. y Azadinium spp., las cuales presentan alta abundancia.
4) El estudio presenta el primer reporte visual del dinoflagelado nocivo Azadinium spp. en
las costas de Chile, identificado directamente por microscopía óptica.
5) La distribución del fitoplancton potencialmente nocivo estuvo fuertemente asociado con
las variables ambientales salinidad, concentración de NO3 y la proporción de Si/N. Las
especies que mostraron un mayor asociación con estas variables oceanográficas fueron los
dinoflagelados Azadinium spp., Gymnodinium aureolum, Gymnodinium spp., Karenia
mikimotoi y Dinophysis acuminata y las diatomeas Skeletonema spp., Pseudo-nitzschia
pseudodelicatissima, P. delicatissima, P. seriata, P. pungens, Cerataulina pelagica.

6
6) Los bajos niveles de nutrientes y la relaciones N/P y N/Si, principalmente dentro del Golfo
de Penas, sugieren un drástico consumo de nutrientes después de una condición postbloom.
7) La concentración de oxígeno disuelto presente en la columna de agua de la zona de estudio
sugiere que la mortandad de peces no se habría debido a un evento de hipoxia ambiental.
8) Los análisis estadísticos de las variables biológicas y oceanográficas permitieron
identificar 3 zonas diferentes: (a) Golfo de Penas y Península de Taitao, (b) Canal Moraleda
y (c) Canales sur (Messier y Wide). Las variables oceanográficas que más contribuyeron a la
diferenciación en 3 zonas fueron la salinidad, temperatura y concentración de los nutrientes
NO3 y Si(OH)4. El segundo conjunto de variables que más contribuyeron a diferenciar
dichas zonas fueron la magnitud del viento, la concentración de PO4 y la proporción Si/N.
9) Análisis de CAP más detallados dentro del la zona Golfo de Penas y Península de Taitao,
muestra que la distribución de los dinoflagelados guarda relación con la profundidad, donde
la temperatura y la salinidad son los principales parámetros que afectan la distribución en
esta zona particular.
10) Los taxones que más contribuyeron a las diferentes zonas fueron Azadinium spp.,
Gymnodinium aureolum, Karenia mikimotoi, Gymnodinium spp., representativos del Golfo
de Penas y Taitao, además de Pseudo-nitzschia pseudodelicatissima; Alexandrium ostenfeldii
fue la especie más representativa del Canal Moraleda, mientras que Skeletonema spp.,
Ceratulina pelagica y Dinophysis acuminata fueron los taxones que destacan en los Canales
Sur.
11) Las condiciones oceanográficas observadas en el Golfo de Penas pueden permitir el
desarrollo de altas abundancias de especies de dinoflagelados nocivos, en consecuencia se
recomienda evitar el recambio de agua en el transporte de smolts de salmones en dicho Golfo.

7
II INTRODUCCION
El 2 de Febrero de 2017, SERNAPESCA tuvo conocimiento de 3 eventos significativos de
mortalidad de peces ocurridos durante su transporte marítimo entre la Región de Los Lagos
y la Región de Magallanes. Los eventos de mortalidad ocurrieron durante el recambio de
agua de las naves transportadoras (well-boats) en las cercanías del Golfo de Penas (Región
de Aysén). Los eventos señalados provocaron la muerte de la totalidad de los peces
transportados, los cuales eran destinados tanto a siembra en centros de cultivo de la Región
de Magallanes como a plantas de procesamiento en la Región de Los Lagos.
Ante esta situación, SERNAPESCA procedió a declarar “Emergencia de Plaga” mediante la
Resolución Exenta N° 510, del 10 de febrero de 2017, con el objeto de aplicar medidas
inmediatas al traslado de especies hidrobiológicas y prospectar el área para identificar y
cuantificar la presencia de especies de microalgas productoras de FAN.
La identificación preliminar de microplancton reportada por el Instituto Tecnológico del
Salmón (INTESAL), mediante un muestreo de oportunidad realizado a bordo de una nave de
transporte, indicó la presencia de un dinoflagelado tipo Karenia mikimotoi en la zona del
Golfo de Penas. Esta especie es uno de los dinoflagelados más comunes causantes de FAN
en regiones del Atlántico Norte (Hoagland et al. 2002, Scatasta et al. 2003, Kim 2006). Las
floraciones de esta especie suelen cubrir extensas zonas, alcanzando valores sobre un millón
de células por litro, lo cual se asocia a grandes mortandades de fauna marina (Davidson et al.
2009, Silke et al. 2010). Si bien, las causas del efecto nocivo de K. mikimotoi no están claras,
existen mediciones de poliésteres citotóxicos que han sido extraídos de cultivos de esta
especie (Silke et al. 2005).
En este contexto, entre las medidas dispuestas por SERNAPESCA en el marco de la
mencionada Emergencia, se llevó a cabo un crucero científico de prospección en el Buque
OPV 83 "Marinero Fuentealba" (Armada de Chile), con el fin de estudiar la presencia de
microalgas productoras de floraciones algales nocivas (FAN) en la zona del Golfo de Penas

8
y sectores aledaños. El muestreo y análisis de las FAN de este crucero de prospección fue
encargado al Centro Interdisciplinario para la Investigación Acuícola (INCAR).
Los objetivos del estudio encargado al INCAR por SERNAPESCA son:
(i) Confirmar la presencia de microalgas nocivas en la zona del Golfo de Penas
y fiordos/canales adyacentes y en especial del dinoflagelado Karenia mikimotoi.
(ii) Identificar cuantitativamente la abundancia de las microalgas productoras de
floraciones algales nocivas y potencialmente nocivas en la zona de estudio.
(iii) Caracterizar las condiciones oceanográficas de la zona de estudio y las posibles
asociaciones entre dichas condiciones y la presencia de floraciones algales
nocivas.
El presente Informe Final, describe los resultados provenientes del cumplimiento de cada
objetivo del estudio.

9
III METODOLOGIA
3.1 Muestreo durante el Crucero Golfo De Penas
Se muestrearon 21 estaciones en el Golfo de Penas (Región de Aysén) y sectores aledaños
(Fig. 1) a bordo del OPV 83 "Marinero Fuentealba", perteneciente a la Armada de Chile,
entre los días 17 y 21 de febrero de 2017. La cobertura espacial comprendió desde la entrada
norte del Canal Moraleda (44°32'09" °S) hasta la salida sur del Canal Wide (49°57'51" °S) y
desde el punto de vista longitudinal, entre 73°28'07" y 75°44'00" °W.
Las variables monitoreadas durante el crucero fueron:
a) Presencia/ausencia y cuantificación de Karenia en 3 profundidades de cada estación
oceanográfica. Se colectaron muestras concentradas (20 m) y no concentradas.
b) Presencia/ausencia y cuantificación de otras especies que generan floraciones nocivas en
3 profundidades de cada estación oceanográfica.
c) Perfiles verticales de temperatura, salinidad y oxígeno (0-30 metros).
d) Mediciones de SDT, pH y REDOX, en cada una de las 3 profundidades muestreadas.
e) Concentración de nutrientes (nitrato, nitrito, fosfato, silicato) en 3 profundidades de cada
estación oceanográfica, las cuales eran similares a las del muestreo de fitoplancton.
f) Concentración de clorofila y feopigmentos en 3 profundidades de cada estación
oceanográfica (las mismas profundidades del muestreo de fitoplancton). Se realizaron,
además, estimaciones de clorofila fraccionada por tamaño (2 fracciones: fracción total [Chla-
-picoplancton]).
g) Estimación de la transparencia y de la profundidad de la zona eufótica mediante el uso de
Disco Secchi en cada estación oceanográfica

10
Figura 1. Mapa de la zona de estudio, Crucero Golfo de Penas. El recuadro rojo muestra los
canales y fiordos incluidos en la zona de emergencia de plaga por reporte de microalga
nociva según la RES. Exe. 510/17; SERNAPESCA.
Canal Moraleda
Golfo de Penas
Canal Messier
Canal Wide
L a
t i t
u d
(°S
)
Longitud (°W)

11
En cada estación oceanográfica se realizaron perfiles continuos de temperatura, salinidad y
oxígeno disuelto a través de un CTDO AML-Oceanographic (Modelo Minos X).
Para la toma de muestras discretas de agua se escogieron 3 profundidades (5, 15 y 30 m). La
toma de muestras se realizó por medio de botellas Niskin asépticas de 10 L (24 L por
estación). Se registraron medidas inmediatas de Sólidos Disueltos Totales (SDT), pH y
potencial de Oxidación-reducción (REDOX) a través de una sonda Multiparametro HANNA
(Modelo HI98191) para datos discretos.
Para el muestreo cuantitativo de microplancton vivo (reconocible por la presencia de
cloroplastos después de la fijación) del agua proveniente de las botellas Niskin, se colectaron
muestras no concentradas de agua directamente en un frasco ambar de 250 mL, las cuales se
conservaron con una solución ácida de Lugol al 0,1% (Karayanni et al. 2004). Además, para
análisis cuantitativos y cualitativos de microfitoplancton, se colectó una muestra
concentrada, para lo cual se tamizó (trama de red del tamiz: 20 m) entre 10 a 16 L de agua
en frascos estériles de polipropileno de alta densidad ambar de 250 mL. Las muestras
concentradas se conservaron inmediatamente con una solución ácida de lugol al 0,1% y una
réplica en parafolmaldehido a una concentración final de 0,1%. Todas las muestras de
fitoplacton fueron almacenadas en condiciones de frío (4 ° C) y oscuridad hasta su posterior
análisis en laboratorio. Ambas muestras con lugol fueron analizadas mediante el método de
Utermöhl (Villafañe y Reid 1995), utilizando un microscopio invertido Nikon Eclipse
TE2000S, a una resolución de 1000X y objetivos con Contraste Diferencial Interferencial
(DIC) y aumento adicional de 150X. A partir de estos análisis se obtuvieron estimaciones de
abundancia por taxones y morfotipos. La cuantificación duró hasta que se registraron al
menos 100 células (microplancton) de los 4 taxa más dominantes de en cada muestra. Para
la identificación taxonómica, se utilizaron las guías de Tomas (1997), Anderson et al. (2002),
Lynn y Small (2002), Thompson (2004), Rivera (1969) y Rivera (1985).
Para la identificación y descripción morfológica de las especies de dinoflagelados, se
consideró la forma y tamaño celular, tabulación de teca y la forma detallada de placas
individuales de acuerdo a Balech (1989, 1995). Para un estudio más exhaustivo de la
morfología de las placas, se utilizó un microscopio compuesto de epifluorescencia Nikon

12
TE2000s. Este microscopio cuenta con una cámara fría digital, más filtros UV (385–400 nm),
azul (450–480 nm) y verde (480–550 nm). Para este análisis, se usaron las muestras
concentradas preservadas en paraformaldehido (Pernthaler et al. 2001, Karayanni et al.
2004). Dentro de los 5 días posteriores a la fecha de muestreo, dos submuestras de 150 mL
(de 5 m de profundidad) fueron tratadas con Calcofluor White (Fritz y Triemer 1985) al 0,1
% y la otra con una mezcla de DAPI (4,6-diamidino-2-fenilindol) y Proflavina (3-6 diamino
acridina hemi-sulfato) a una concentración final de 5 μg mL-1 (Verity y Sieracki, 1993) sobre
filtros negros de membrana de policarbonato (10 m de poro, 25 mm de diámetro). Estos
filtros se montaron inmediatamente en portaobjetos de vidrio y se añadió una gota de aceite
de inmersión antes de cubrirlos con un cubreobjeto de vidrio. Inmediatamente, se
almacenaron a -20 °C en oscuridad hasta su posterior análisis. Además, para las especies de
cubiertas muy tenues, cuyas suturas son poco visibles y poder revelar los campos poligonales
de las cubiertas, como sucede en el caso de los Gymnodiniales, se empleó la técnica de
impregnación argentica de Chatton–Lwoff, resumida por Corliss (1952), con modificaciones
de Biecheler (1952) y Chatton (1952). Con la misma finalidad, se utilizó la tinción con yodo
(lugol) propuesta por Bursa (1958). Para la identificación a nivel de especie de algunos
géneros dominantes de diatomeas, se aplicó la técnica de acidificación utilizando parte de las
muestras concentradas para dejar sólo las valvas y analizar sus características morfológicas
(Hasle, 1978). Las proporciones de valvas en estas muestras se asignaron proporcionalmente
a las estimaciones de abundancia.
La concentración de clorofila-a se determinó a partir del filtrado de 0,3 L de agua de mar, en
duplicado, para la fracción total (Chla-a_total) y < 20 m (Chla_nano-picoplancton),
utilizando filtros Whatman® GF/F y preservados a -20 ºC hasta su posterior análisis. La
extracción fue realizada con acetona 90% y su concentración medida por fluorescencia en un
fluorómetro Turner Designs 10AU (Holm-Hansen et al. 1965). Se determinó la diferencia de
ambas fracciones para obtener la fracción > 20 m (Chl-a_micro).
Los análisis de macronutrientes (PO4, Si(OH)4, NO2, NO3), fueron realizados por el
Laboratorio de Biogeoquímica del Departamento de Oceanografía de la Universidad de
Concepción, liderado por la Dra. Laura Farías. Las muestras fueron filtradas por filtros GF/F
y almacenadas en tubos Falcon estériles de 50 mL, preservadas a -20 °C hasta el momento

13
de su análisis realizado en laboratorio. Los recipientes fueron previamente lavados con HCl
al 5% y enjuagados con agua destilada. Nitrato, nitrito y fosfato fueron medidos siguiendo
las técnicas propuestas por Strickland y Parsons (1972), con modificaciones para su análisis
en un autoanalizador Technicon® II. El silicato fue medido siguiendo el método propuesto
por Technicon (1977).
La estimación de la profundidad de compensación se realizó en base a las lecturas de turbidez
medidas con un disco Secchi, usando el coeficiente de extinción de la luz, mediante la
fórmula de Poole y Atkins (1926).
3.2 Análisis estadísticos
Para explorar la base de datos y detectar si existen diferencias entre las zonas de muestreo,
en términos de la estructura multivariada de las variables ambientales; y de la composición
y abundancia de especies de fitoplancton nocivo colectado en dichas zonas, se analizaron las
similitudes en la composición y abundancia de fitoplancton nocivo entre las estaciones de
muestreo. Estas fueron exploradas por medio de Análisis de escalamiento no-métrico
multidimensional (MDS) con datos integrados, correspondientes a los primeros 30 m de la
columna de agua. Los datos de abundancia ocupados para este análisis, se procesaron
siguiendo el método descrito por Field et al. (1982). Este procedimiento para el
pretratamiento de los datos de abundancia, consiste primeramente en estandarizar a
porcentaje del total de cada muestra y luego transformarlos, en este caso, a raíz cuarta. Con
la matriz resultante se calculó una matriz de similitud basada en el índice de Bray-Curtis, la
cual se utilizó en el análisis MDS. Además, se estimó la matriz de similitud de Jaccard, para
datos de presencia-ausencia, y posteriormente se realizó un MDS basado en esta matriz. Las
transformaciones de raíz cuarta y presencia-ausencia se realizaron con la finalidad de quitar
el mayor peso en el análisis a las especies dominantes y entregar mayor relevancia a las
especies raras (Anderson et al. 2006). Luego, para validar estadísticamente los resultados
obtenidos por los análisis MDS, se utilizaron las mismas matrices de similitud de Bray-Curtis
y Jaccard ya generadas y se llevó a cabo un PERMANOVA (“Permutational Multivariate
Analysis of Variance”) (Anderson 2001), seguido de un análisis “Pair-Wise ”, con el fin de
detectar diferencias significativas entre las estaciones de muestreo. Para todos los análisis se
utilizaron las opciones modelo no-restringido con 9999 permutaciones (“unrestricted model

14
with 9999 permutations”). Los análisis se hicieron en el programa PRIMER v6 y
PERMANOVA+ para PRIMER (Clarke y Warwick 2001, Clarke y Gorley 2006, Anderson
et al. 2008).
Para explorar que especies son las que más aportan a la similitud intra-zonas, se utilizó el
análisis SIMPER (“similarity percentages”), el cual permite identificar la contribución
porcentual de los diferentes taxa a la similitud intra-grupos, identificando los que entregan
un mayor aporte (Clarke y Warwick 2001, Clarke y Gorley 2006).
Posteriormente, para explorar las variaciones espaciales de la estructura (composición y
abundancia) de comunidades fitoplanctónicas (incluyendo solo el fitoplancton
potencialmente nocivo) y las variables ambientales asociadas (sin integrar verticalmente los
datos), se aplicaron análisis multivariados utilizando PRIMER-E (versión 6.1.13) con
PERMANOVA (versión 1.0.3) (Clarke y Gorley 2006, Anderson et al. 2008). Los datos
biológicos y ambientales fueron clasificados según la estación de muestreo. Para analizar
las matrices biológicas, se utilizó toda la base de datos (i.e. sin integración vertical) en
profundidad (5, 15, 30m) y todos los géneros o especies identificadas como potencialmente
nocivas.
La matriz ambiental incluyó las siguientes variables: Oxígeno (O2), coeficiente de extinción
de la luz (ZC), magnitud del vientos (VM), dirección del viento (VD), temperatura (TS),
salinidad (SS), densidad (DS), conductividad (CN), potencial oxido reducción (RP),
nutrientes (NO3, NO2, PO4, Si(OH)4), y las relaciones entre pares de nutrientes (N:Si y
N:PO4, con N= NO3 + NO2). Tomando en consideración que algunas de estas variables
estaban fuertemente correlacionadas (Spearman, r > 0,60), las variables incluidas en el
análisis multivariado se redujeron a las siguientes: SS, TS, VM, VD, RP, PH, PO4, NO3,
Si(OH)4, Si/N y N/P. Antes de realizar el análisis, los datos ambientales fueron
transformados mediante log10 (N + 1), excepto para TS y SS, y posteriormente fueron
normalizados. Los datos biológicos de abundancia se transformaron a raíz cuadrada. Los
taxones incluidos en las matrices multivariadas fueron todas las especies o géneros
identificados como potencialmente nocivas (i.e. 39). Como medidas de similitud, se usó la
distancia euclidiana para los datos ambientales y el coeficiente Bray-Curtis ajustado a cero
para los datos biológicos. Se aplicó escalamiento multidimensional no métrico (NMDS) a las

15
matrices biológica y ambiental para producir una ordenación sin restricciones (que maximiza
la variación total de datos) de las muestras en el espacio multivariado. Posteriormente, se
aplicó un análisis permutatorio de varianza (PERMANOVA) tanto para las matrices
ambientales, como a la matriz biológica para evaluar la importancia de la variación entre
grupos (diferencias en la ubicación de los grupos NMDS) con respecto a la variación total de
los datos. El procedimiento de PERMANOVA se basó en la suma de cuadrados (parcial) de
tipo III y en la permutación sin restricciones de los datos brutos. Dado que PERMANOVA
es sensible a las diferencias en las dispersiones entre grupos, un análisis permutatorio de
dispersiones multivariantes (PERMDISP, basado en la desviación de los centroides) probó
la homogeneidad de la dispersión intra-grupo de los grupos NMDS. Además, se obtuvo una
ordenación restringida con un análisis canónico de las coordenadas principales (CAP,
Anderson et al. 2008) para representar los ejes de ordenación en el espacio multivariado que:
a) maximizó las diferencias entre grupos (CDA: análisis discriminante canónico) y b)
maximizó las correlaciones entre la ordenación biológica y las variables ambientales
(CCorA: análisis de correlación canónica). Los análisis de PERMANOVA, PERMDISP y
CAP se realizaron con 9999 permutaciones. Para el PERMANOVA de dos factores (espacial
y profundidad) se evaluaron ambos factores juntos y separados, ya que el análisis de dos
factores produjo resultados similares, pero las diferencias dentro del grupo (PERMDISP)
resultaron ser significativas.

16
IV RESULTADOS
4.1 Condiciones Meteorológicas y Oceanográficas
Las condiciones meteorológicas durante el período de muestreo indican predominancia de
vientos con dirección NW, de magnitudes promedio de 13 ± 5,6 Kt (24±10 Km/h), con cielos
principalmente nublados (Fig. 2), lo que desafortunadamente impide una buena visualización
de la zona de estudio mediante imágenes satelitales, durante los días de de muestreo. Se
observa un frente de mal tiempo durante el paso por la zona del Golfo de Penas, con
magnitudes de 20 Kt (37 Km/h) y dirección desde el NW (Tabla 1), lo que generó una gran
mezcla de la columna de agua en toda esa zona.
Figura 2. Resumen de la magnitud en nudos (Kt) y dirección del viento durante los días de
muestreo 17 al 21 de febrero 2017.
0
45
90
135
180
225
270
315
0% 5% 10% 15% 20% 25%
>0 - 10
>10 - 20
N
S
EW

17
Tabla 1. Magnitud y dirección del viento durante el período de muestreo para cada estación; Kt = nudos, Dirección de viento en grados
(°).
Variable SNP
1
SNP
3
IN SNP
4
SNP
9
SNP
10
E13 E14 E15 E16 E17 SNP
8
SNP
7
SNP
6
SNP
5
E18 E19 E20 E21 E22 E23
Viento Mag
(Kt)
14 5 9 13 20 20 20 17 20 13 8 17 17 15 10 2,2 15 15 4 14 5
Viento Dir. (°) 360 320 360 260 320 350 290 316 320 299 311 285 320 300 326 94 350 30 143 30 350
Sector N NW N W NW N W NW NW NW NW W NW NW NW NE N NE SE NE N

18
Se presentan imágenes satelitales MODIS_aqua de Temperatura Superficial de Mar (TSM)
y Clorofila-a (Chla), combinadas para 8 días, con resolución de 4 Km durante las dos semanas
previas al muestreo (2-17 de febrero) y durante la semana de ejecución del crucero (18-24 de
febrero) (Fig. 3).
En relación a la clorofila-a satelital, lamentablemente la alta cobertura de nubes impide un
buen análisis mediante imágenes de satélite en la zona monitoreada. Sin embargo es posible
ver indicios de mayores concentraciones de clorofila en la zona costera, con filamentos que
se extienden de costa a océano. Las imágenes de TSM muestran una zona con temperaturas
altas hacia la zona norte del área muestreada, con temperaturas aún más altas dentro del Canal
Moraleda, las cuales van disminuyendo gradualmente hacia el sur (Fig. 3).
Las condiciones oceanográficas observadas, en cuanto a salinidad y temperatura superficial
(Fig. 4) durante el crucero, permiten identificar tres zonas con diferencias estadísticamente
significativas: (i) desde la entrada del canal Messier al sur, (ii) la zona del Golfo de Penas y
Península de Taitao, y (iii) el Canal Moraleda.
Una capa de mezcla muy marcada hasta los 5 m de profundidad se observó en el Canal Wide
(SNP1, SNP3 e IN), registrando una marcada baja densidad superficial y alta frecuencia
Brunt Väisälä, lo que indica una alta estratificación (Fig. 5D-E). A medida que el muestreo
avanzó hacia la zona norte del área de estudio, se observó una tendencia a una menor
estratificación de la columna de agua (Fig. 5).
Angostura Inglesa (estación IN) registró las más bajas salinidades observadas (aprox. 12) en
superficie durante el crucero. La capa superficial (40 m) presentó condiciones claramente
óxicas durante el crucero (Fig. 4B).

19
Figura 3. Data satelital promedio de 8 días de Chla y TSM para las semanas previas al
muestreo (2 -17 de febrero de 2017) y durante la semana de ejecución del crucero (18-24 de
febrero de 2017).
02 -09 de febrero de 2017 10 -17 de febrero de 2017 18 -24 de febrero de 2017
longitud
longitud
L a
t i t
u d
L a
t i t
u d

20
Figura 4. Distribución superficial de parámetros Oceanográficos A) Temperatura, B)
Salinidad, C) Oxígeno y D) Densidad.

21
Figura 5. Perfiles verticales continuos de parámetros oceanográficos: A) Temperatura, B) Salinidad, C) Oxígeno, D) Densidad y E)
Frecuencia Brunt Väisälä.
Moraleda Channel
Gulf of Penas
Messier Channel
Wide Channel
A
B
C
D
E

22
En la zona de estudio, el Agua Superficial Subantártica se mezcla con agua dulce (AD) en
diferentes proporciones, según las contribuciones de los ríos, glaciares, escorrentía costera,
pluviosidad y la distancia de las fuentes de AD (Sievers y Silva, 2008). Sievers y Silva (2008)
han descrito que en la zona del Canal Messier y Canal Wide (i.e. SNP1, SNP3 e IN) se
produce un proceso de mezcla lineal entre SAAW y AD, donde el agua resultante de este
proceso (con salinidades entre 31- 33) se conoce como Agua Subantárctica Modificada
(MSAAW) y el agua de baja salinidad como Agua Estuarina (Figs. 5B y 6). Las primeras
estaciones de muestreo SNP1, SNP3 e IN muestran presencia de agua estuarina–salobre, lo
cual es consistente con lo reportado por Sievers y Silva (2008) para la zona (Figs. 4B, 5B y
6). En las estaciones ubicadas hacia el norte de Bahía Anna Pink hacia el Canal Moraleda
(E21, E22 y E23) se mantienen condiciones similares a las observadas en el Golfo de Penas
(Figs. 5B y 7), con una columna de agua muy mezclada, donde se observa principalmente un
aumento de la salinidad cerca de 30 (Figs. 4B, 5B y 8).
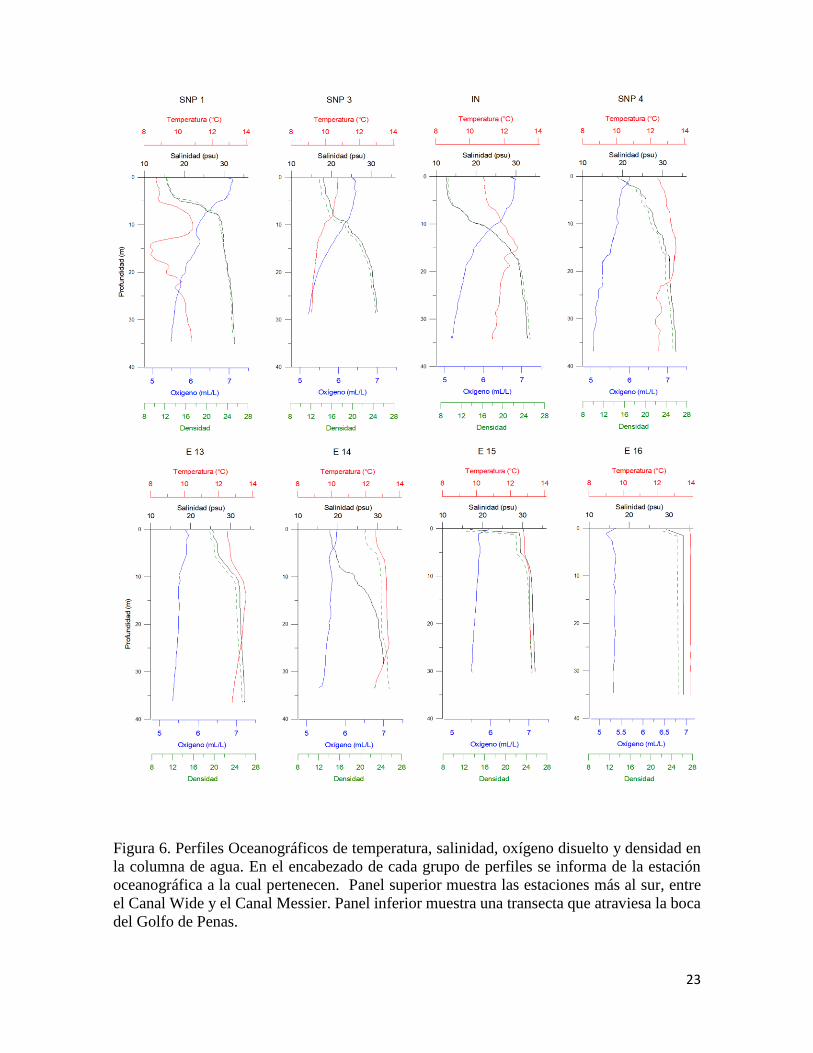
23
Figura 6. Perfiles Oceanográficos de temperatura, salinidad, oxígeno disuelto y densidad en
la columna de agua. En el encabezado de cada grupo de perfiles se informa de la estación
oceanográfica a la cual pertenecen. Panel superior muestra las estaciones más al sur, entre
el Canal Wide y el Canal Messier. Panel inferior muestra una transecta que atraviesa la boca
del Golfo de Penas.

24
Figura 7. Perfiles Oceanográficos de temperatura, salinidad, oxígeno disuelto y densidad en
la columna de agua. En el encabezado de cada grupo de perfiles se informa de la estación
oceanográfica a la cual pertenecen. Panel superior muestra las una transecta por la parte
externa a la península de Taitao y el panel inferior muestra las estaciones al interior del Golfo
de Penas.

25
Figura 8. Perfiles Oceanográficos de temperatura, salinidad, oxígeno disuelto y densidad en la columna de agua.
En el encabezado de cada grupo de perfiles se informa de la estación oceanográfica a la cual pertenecen. Panel
superior muestra las estaciones más oceánicas del Golfo de Penas y el panel inferior muestra las estaciones al
interior del Canal Moraleda.

26
Figura 9. Perfiles de (A) Sólidos disueltos totales (SDT), (B) Potencial redox y (C) pH.
A
B
C

27
Los perfiles oceanográficos de SDT, REDOX y pH, diferencian las zonas de canales de la
del Golfo de Penas y de la zona exterior a la Península de Taitao (Fig. 9). Los SDT muestran
una notoria baja concentración en los canales de la zona sur (SNP1, SNP3 e IN; Fig. 10B),
destacando además los altos valores registrados en la zona del Golfo de Penas (Fig. 7A). El
REDOX es análogo al pH, ya que el pH mide la actividad de protones y el potencial REDOX
mide la de los electrones, por lo que observamos, en términos generales, los mayores valores
de pH asociados a los valores más bajos de REDOX (Figs. 9B-C y 10). Los valores de pH se
encuentran en el rango normal esperado a la zona costera, con valores más bajos en la zona
de canales. Estos mínimos superficiales parecen estar asociados a una delgada capa de agua
menos salina, que como menciona Silva y Calvete (2002), estaría asociada a los ríos aledaños,
debido a que el agua dulce se caracteriza por tener un menor pH y una mayor solubilidad de
oxígeno que el agua de mar, ello generaría esta inversión no acompañada por una
disminución del oxígeno disuelto. Silva (2008) describe la capa superficial (30-50 m de
grosor) de los canales de la zona de estudio con alto contenido de oxígeno (5-8 mL L-1, 90-
130 % de saturación) y con altos valores de pH (8,0-8,3).
Para estimar la profundidad de la zona fótica (i.e. incidencia de luz del 1% ) se utilizó el disco
Secchi (Fig. 10A). No se aprecia un efecto directo de los SDT en profundidad de la capa
fótica (R2=0,12; p>0,05). En algunas estaciones podemos relacionar la profundidad de
compensación (Fig. 10 A) a la concentración de la clorofila-a total, mostrando una capa fótica
más somera asociada a la presencia de alta concentración de clorofila-a (Fig. 11A), como
ocurre en las estaciones E22, SNP3. Lo contrario ocurre en las zonas donde la luz alcanza
mayores profundidades (E18, 19 y 20), las cuales están asociadas a muy bajas
concentraciones de clorofila en la columna de agua (Figs. 11A y 12A).

28
Figura 10. Distribución superficial de parámetros Oceanográficos. (A) Profundidad de Compensación, (B) SDT,
(C) REDOX y (D) pH. No existen mediciones de Disco Secchi en las estaciones SNP4, IN y E23.
E23
E22
E21
E20
E19
E18
E17E16
E15E14
E13
SNP8SNP7
SNP6SNP5
SNP4
SNP1
SNP3
IN
SNP10
SNP9
E23
E22
E21
E20
E19
E18
E17E16
E15E14
E13
SNP8SNP7
SNP6SNP5
SNP4
SNP1
SNP3
IN
SNP10
SNP9
E23
E22
E21
E20
E19
E18
E17E16
E15E14
E13
SNP8SNP7
SNP6SNP5
SNP4
SNP1
SNP3
IN
SNP10
SNP9
E23
E22
E21
E20
E19
E18
E17E16
E15E14
E13
SNP8SNP7
SNP6SNP5
SNP4
SNP1
SNP3
IN
SNP10
SNP9
pHREDOX (mV)
Total Dissolved Solutes (g/L)Compensation Zone (m)
Lati
tude
Longitude Longitude
Lati
tude
A
C
B
D

29
4.2 Clorofila-a y Feopigmentos
La biomasa fitoplanctónica expresada como clorofila-a, registró valores máximos de 6,65 mg
m-3 para la fracción total, con valores más altos desde la costa hacia el océano (Fig. 8), en
donde la mayor biomasa fitoplanctónica de la zona de muestreo se encuentra dentro del Canal
Moraleda (Figs. 11, 12 y 14). Concentraciones de clorofila-a, en el rango de las observadas
en este muestreo, han sido descritas previamente para esta área (Pizarro e tal., 2000; Ramirez
y Pizarro, 2005; Iriarte etal.,2007; Silva y Palma, 2008; Paredes y Montecinos, 2011).
Ramírez y Pizarro (2005) atribuyen el crecimiento de fitoplancton en el Canal Moraleda a la
fertilización de la cuenca a través de aguas oceánicas y el gradiente térmico que se forma en
los primeros 25 m, lo que permite la permanencia del fitoplancton en la zona fótica.
La fracción de Chl-a_micro muestra el mismo patrón de distribución que la Chl-a_total,
coincidiendo en los puntos de más alta concentración (Figs. 11B, 12B), mostrando una alta
asociación (R2=0,62; <0,05) entre estas dos fracciones. La fracción < 20 m de Chl-a_nano-
pico muestra mayores concentraciones hacia la zona norte del área muestreada, con valores
máximos de 2,85 mg/m3, con una representación cercana al 68,9 % de la Chl-a_total,
mientras que en la zona sur solo llega a un 20 % de la Chl-a_total (Figs. 11C, 12C, 14). Existe
una alta correlación (R2 = 0,73; p<0,05) entre la Chl-a_total y la fracción de Chl-a_nano-
pico.
Los feopigmentos integrados (Fig. 11D) muestran solo alta representación en las estaciones
dentro del Canal Moraleda (Fig. 15), esta alta concentración se distribuye en toda la columna
de agua de las estaciones E22 y E23 (Fig. 13). Las fracciones micro_>20m y nano-
pico_<20 m muestran el mismo patrón de distribución que los feopigmentos totales, con
alta concentración en el Canal Moraleda y en las estaciones más costeras (SNP5 y SNP6)
dentro del Golfo de Penas (Figs. 13, 14).

30
Figura 11. Distribución superficial de la clorofila-a integrada (mg Chla m-2) entre 0-30m en
la zona del Golfo de Penas. A) Chla-a_total, B) Chl-a_micro C) Chl-a_nano-pico) y D)
Feopigmentos totales.
E23
E22
E21
E20
E19
E18
E17E16
E15E14
E13
SNP8SNP7
SNP6SNP5
SNP4
SNP1
SNP3
IN
SNP10
SNP9
E23
E22
E21
E20
E19
E18
E17E16
E15E14
E13
SNP8SNP7
SNP6SNP5
SNP4
SNP1
SNP3
IN
SNP10
SNP9
Total Chlorophyll-a (mg/m2)
Lati
tud
e
Chla_mycro (mg/m2)
Total Phaeopigments (mg/m2)Chla_nano-pico (mg/m2)
E23
E22
E21
E20
E19
E18
E17E16
E15E14
E13
SNP8SNP7
SNP6SNP5
SNP4
SNP1
SNP3
IN
SNP10
SNP9
E23
E22
E21
E20
E19
E16E15
E14E13
SNP8SNP7
SNP6SNP5
SNP4
SNP1
SNP3
IN
Lati
tud
e
Longitude Longitude
A B
C D

31
Figura 12. Perfiles verticales de clorofila-a en la zona del Golfo de Penas y canales aledaños. (A) Chla-a_total, (B) Chl-a_micro y (C)
Chl-a_nano-pico.
A
B
C

32
Figura 13. Perfiles verticales de feopigmentos en la zona del Golfo de Penas y canales aledaños. (A) Feopigmentos_totales, (B)
Feopigmentos_micro y (C) Feopigmentos_nano-pico.
A
B
C
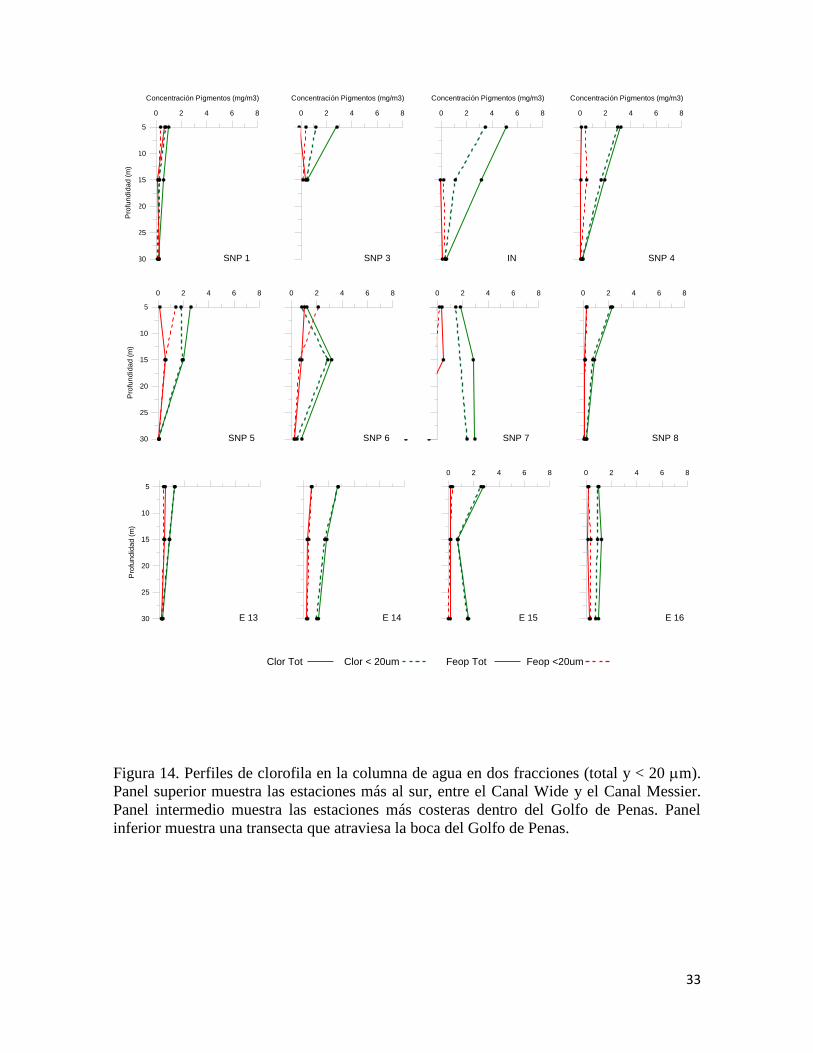
33
Figura 14. Perfiles de clorofila en la columna de agua en dos fracciones (total y < 20 m).
Panel superior muestra las estaciones más al sur, entre el Canal Wide y el Canal Messier.
Panel intermedio muestra las estaciones más costeras dentro del Golfo de Penas. Panel
inferior muestra una transecta que atraviesa la boca del Golfo de Penas.
0 2 4 6 8
Concentración Pigmentos (mg/m3)
-30
-25
-20
-15
-10
-5
Pro
fun
did
ad
(m
)
0 2 4 6 8
0 2 4 6 8
0 2 4 6 80 2 4 6 8
-30
-25
-20
-15
-10
-5
Pro
fun
did
ad
(m
)
0 2 4 6 8
0 2 4 6 8
Concentración Pigmentos (mg/m3)
0 2 4 6 8
0 2 4 6 8
Concentración Pigmentos (mg/m3)
0 2 4 6 8
Concentración Pigmentos (mg/m3)
-30
-25
-20
-15
-10
-5P
rofu
nd
ida
d (
m)
SNP 4
E 13 E 14
SNP 6
E 16
SNP 8SNP 5
E 15
IN
SNP 7
SNP 3SNP 1
Clor Tot Clor < 20um Feop <20umFeop Tot

34
Figura 15. Perfiles de clorofila en la columna de agua en dos fracciones (total y < 20 m).
Panel superior muestra las estaciones más externas del Golfo de Penas. El panel intermedio
muestra una transecta por la parte externa a la Península de Taitao y el panel inferior muestra
las estaciones al interior del Canal Moraleda.
0 2 4 6 8
Concentración Pigmentos (mg/m3)
-30
-25
-20
-15
-10
-5
Pro
fun
did
ad
(m
)
0 2 4 6 8
-30
-25
-20
-15
-10
-5
Pro
fun
did
ad
(m
)
0 2 4 6 80 2 4 6 8
-30
-25
-20
-15
-10
-5
Pro
fun
did
ad
(m
)
0 2 4 6 8
Concentración Pigmentos (mg/m3)
0 2 4 6 8
0 2 4 6 80 2 4 6 8
E 20
SNP 9
E21
E 19E 17
SNP 10
E 18
E 23E 22
Clor Tot Clor < 20um Feop <20umFeop Tot

35
4.3 Nutrientes
Las zonas de canales sur (Wide y Messier) de la zona de estudio, así como el Canal Moraleda
muestran altos niveles de macronutrientes, presentándose valores máximos de 13,6 mol L-
1 de nitrógeno inorgánico (NI; Nitrato + Nitrito) en el Canal Moraleda (E 23; Fig. 16A),
representando el nitrato en promedio el 91% del NI total (Fig. 16 A-C). Las mismas
tendencias presentan el fosfato y silicato, siendo notablemente mayores en la zona de canales,
con máximos de 1,4 y 7,8 mol L-1 respectivamente (Figs. 16, 17).
Los valores de fosfato y nitrógeno se encuentran en el rango de los valores esperados para la
zona (Silva y Calvete, 2002, Silva 2008). Silva (2008) reporta rangos para la capa superficial
(0-50m) de 0-0,8 μM de fosfato y 0-8 μM de nitrato y silicato. Las fuentes de fosfato y nitrato
provienen principalmente del mar a las zonas de canales, en donde los ríos, lluvia y el
derretimiento glacial aportan además bajas concentraciones de estos nutrientes (fosfato 0-1,2
μM, nitrato 0-2 μM) (Silva 2008). Por otro lado, el silicato es alto en estas fuentes de agua
dulce (40-150 μM) y hacen una importante contribución a la capa superficial (Silva y Calvete,
2002, Silva 2008).
El nitrógeno es usualmente descrito como un nutriente limitante en el océano (e.g. Sze 1998),
y podemos observar en el presente estudio varias estaciones dentro del Golfo de Penas
(SNP4, SNP5, SNP6, SNP7, E14, E16, E20) con valores de la relación N:P < 6 (Fig. 18),
bastante por debajo de la relación de Redfield (i.e. N:P=16:1), indicando un potencial déficit
de N (Moore et al. 2013). La Figura 17 muestra los valores integrados de macronutrientes
(0-30 m), donde se puede observar claramente la baja concentración de nitrógeno inorgánico
en la zona central del Golfo de Penas. Considerando las relaciones Si:N < 1 y Si:P ≤ 3 como
índices de una potencial limitación de Si (Harrison et al. 1977, Levasseur y Therriault 1987),
las estaciones E16 y E22 se podrían considerar como zonas de limitacón de nutrientes (Fig.
18).

36
Figura 16. Perfiles verticales de nutrientes en la zona del Golfo de Penas y canales aledaños.
(A) Nitrógeno Inorgánico total, (B) Nitrato, (C) Nitrito, (D) Fosfato y (E) Silicato.
D
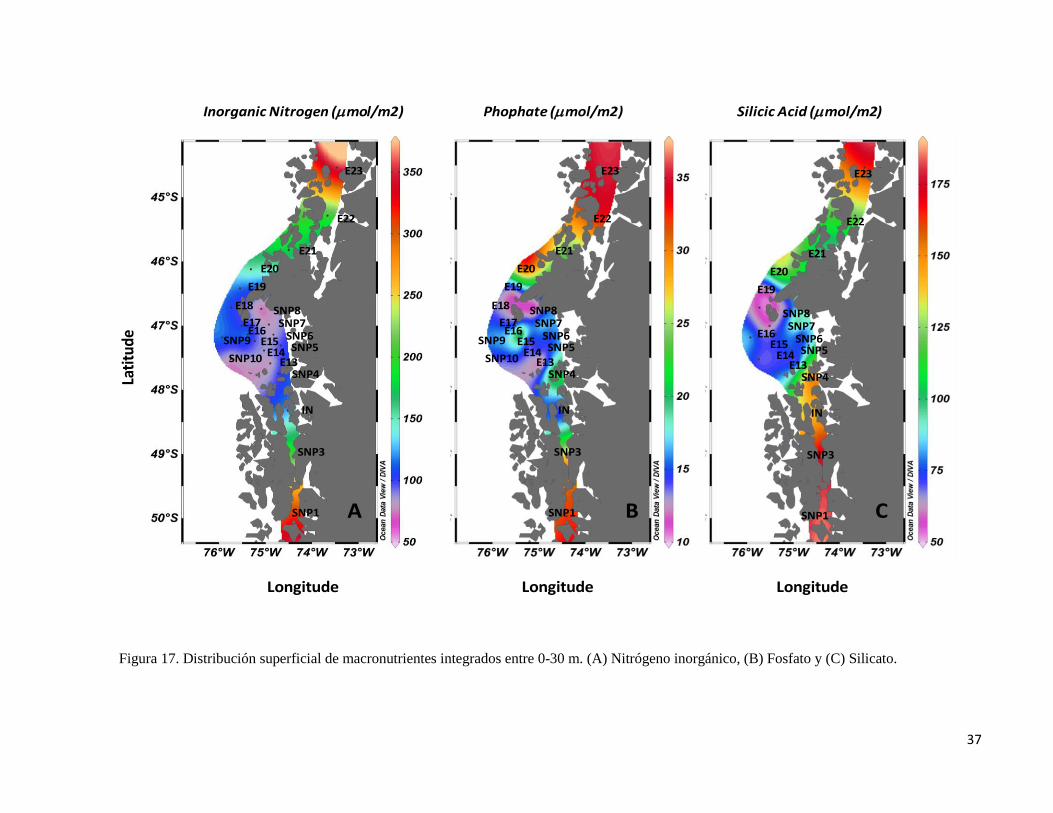
37
Figura 17. Distribución superficial de macronutrientes integrados entre 0-30 m. (A) Nitrógeno inorgánico, (B) Fosfato y (C) Silicato.
E23
E22
E21
E20
E19
E18
E17E16
E15E14
E13
SNP8SNP7
SNP6SNP5
SNP4
SNP1
SNP3
IN
SNP10
SNP9
E23
E22
E21
E20
E19
E18
E17E16
E15E14
E13
SNP8SNP7
SNP6SNP5
SNP4
SNP1
SNP3
IN
SNP10
SNP9
Inorganic Nitrogen (mol/m2)La
titu
dePhophate (mol/m2) Silicic Acid (mol/m2)
E23
E22
E21
E20
E19
E16E15
E14E13
SNP8SNP7
SNP6SNP5
SNP4
SNP1
SNP3
IN
LongitudeLongitude Longitude
A B C

38
Figura 18. Transectas de la relación entre proporciones de macronutrientes. (A) Si/N, (B) N/P y (C) Si/P. Cabe destacar que todos los datos
de la proporción N/P en la zona de estudio están por debajo de la razón de Redfield (i.e. N:P = 16:1).
A
B
C

39
4.4 Composición de Especies
El análisis de la composición de especies, muestra que las especies nocivas representan el
62% de la abundancia total de especies, mostrando alta abundancia en la mayoría de las
estaciones, alcanzando abundancias que superan el 68±8 % de la abundancia total en todas
las estaciones a excepción de las del Canal Moraleda (E21, E22 y E23), donde la abundancia
de especies nocivas es menor al 27% (ver mapa temático 1_Abundancia total de especies
nocivas).
Detalles de la morfología de los principales generos identificados en el presente informe se
muestran en la Tabla 2 y en la Figura 24, con principal énfasis en las especies objetivo del
género Karenia.
Entre las especies nocivas destacan por su abundancia las diatomeas (62%), seguidas de los
dinoflagelados (37%) (Fig. 19 A-B). Las diatomeas dominaron en 8 estaciones de muestreo,
principalmente en el borde exterior de la Península de Taitao (E18, E19, E20, E21) y en los
Canales Wide y Messier (SPN1, SPN3, IN). (Fig. 19). Las diatomeas céntricas tuvieron una
mayor contribución en 12 estaciones de muestreo (Tabla 3, ver mapa temático 2_Abundancia
de especies de diatomeas nocivas).

40
Figura 19. Abundancia de grupos principales de microfitoplancton vivo en la zona del Golfo de Penas. (A) Diatomeas, (B) Dinoflagelados,
(C) Ciliados y (D) Flagelados.
A
B
C
D

41
Los dinoflagelados fueron dominantes en 13 estaciones, principalmente en el Golfo de Penas
y en el Canal Moraleda. Los dinoflagelados atecados sobresalen en la mayoría de las
estaciones (Tabla 3, ver mapa temático 3_ Abundancia de especies de dinoflagelados
nocivos). Cabe señalar que no todos los dinoflagelados poseen clorofila, ya que estos
organismos pueden ser mixotrofos o heterótrofos, por lo que no se espera una relación directa
con las concentraciones de clorofila-a.
Destacan por su abundancia las diatomeas de los géneros Skeletonema (25,5%), Pseudo-
nitzschia (18,8 %), Leptocylindrus (16,6 %) y los dinoflagelados Gymnodinium (17,9 %),
Azadinium (10,5 %), Ceratium (3,3 %) y Karenia (2,5 %) (Figs. 20, 21 y 24; ver mapa
temático 4_ Abundancia agrupadas por género).
En relación a los máximos de clorofila observados (Fig. 11A), podemos ver zonas en donde
distintas especies de diatomeas podrían estar relacionadas a los máximos de clorofila, se
observa en casos puntuales como la estaciones E20 donde Leptocylindrus danicus (36.984
cél. L-1) es la especie más representativa, o en el caso de las estaciones IN donde Skeletonema
spp. presenta las máximas abundancias de 34.249 cél. L-1, seguidas de Pseudo-nitzschia
pseudodelicatissima y Rhizosolenia setigera con cerca de 10.000 cél. L-1 (Figs. 20, 21, 24).
La estación E22 que presenta la mayor concentración de clorofila-a, se encuentra relacionada
a una alta abundancia de las diatomeas no nocivas Thalassiosira anguste lineata (42.923 cél.
L-1) y Ditylum brightwellii (16.459 cél. L-1) (Figs. 20, 21).
Las especies Pseudo-nitzschia pseudodelicatissima, P. delicatissima, P. pungens, P. seriata,
P. australis se encuentran entre las especies productoras de ácido domoico identificadas en
esta zona, con abundancias integradas máximas de 174.273 x103 cél. m-2 (Figs. 20 y 21). La
mayor abundancia de diatomeas se observa en la zona de canales y en la zona exterior a la
Península de Taitao, mientras que las abundancias más altas de dinoflagelados se encuentran
en la zona del Golfo de Penas. En la Tabla 4 podemos observar la abundancia de las
principales especies formadoras de FAN donde se puede ver en detalle la distribución del
conjunto total de especies identificadas en este estudio (ver mapa temático 5_Abundancia
por grupos).

42
Figura 20. Abundancia de especies representativas de diatomeas (cél. vivas L-1) en la zona del Golfo
de Penas y canales aledaños.

43
Figura 21. Abundancia integrada de especies representativas de diatomeas (cél. vivas m-2) en la
zona del Golfo de Penas y canales aledaños.

44
Se identificaron 4 especies del género Karenia: K. bicuneiformis, K. brevisulcata, K.
mikimotoi y K. papilonacea, de las cuales K. mikimotoi representa el 84 % de la abundancia
del género y 2,49 % de la abundancia total de dinoflagelados, con abundancias máximas de
2.324 cél. L-1 (Figs. 22 y 23). Concentraciones consideradas bajas en relación a la formación
de FAN. Karenia presenta abundancias integradas máximas de 52.645x 103 cél m-2, con
presencia en la mayoría de las estaciones a excepción de SNP3 y E22 (Fig. 22 y 24).
Se identificaron las especies Gymnodinium aureolum y G. simplex, las cuales se deben tener
en consideración cuando se realizan conteos de FAN, ya que son morfológicamente muy
similares a Karenia mikimotoi, pudiendo llevar a sobrestimar la abundancia de K. mikimotoi.
En la Figura 24, se muestran imágenes de las especies de Karenia y Gymnodinium para
evidenciar la similitud existente (Fig. 24).
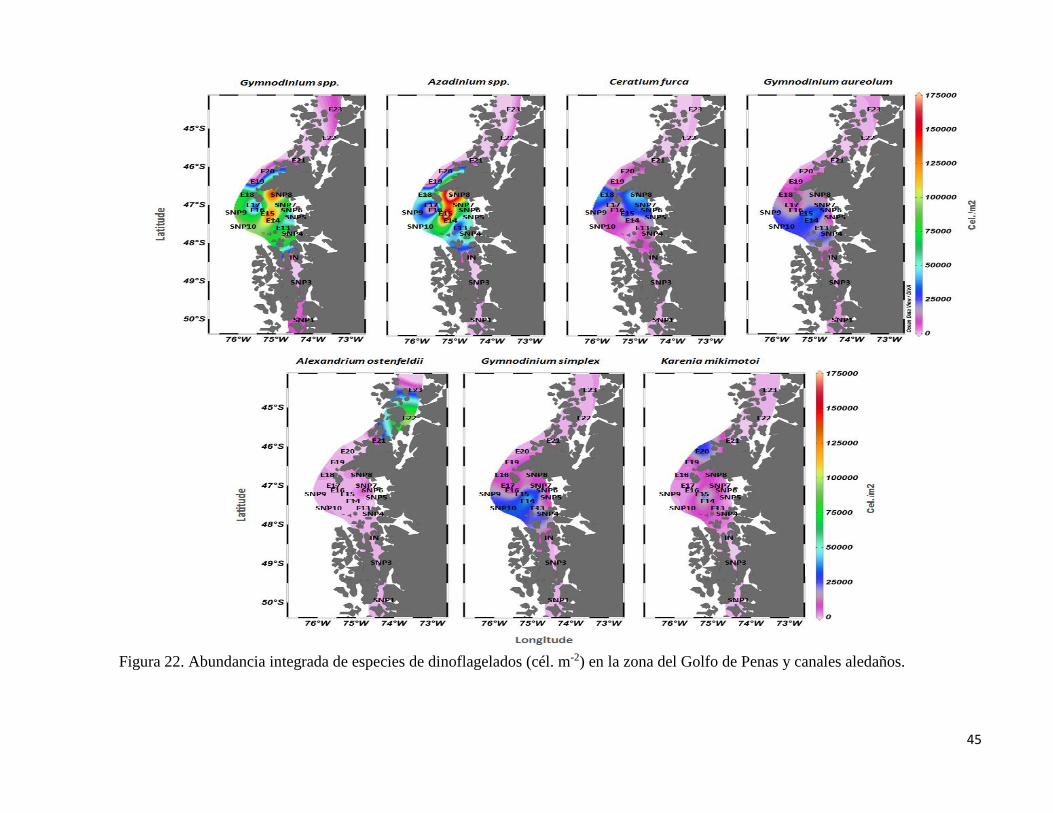
45
Figura 22. Abundancia integrada de especies de dinoflagelados (cél. m-2) en la zona del Golfo de Penas y canales aledaños.

46
Figura 23. Abundancia de especies más representativas de dinoflagelados (cél. L-1) en la zona
del Golfo de Penas.

47
Figura 24. Fotografías de los principales géneros nocivos y detalle de las especies de Karenia
donde se evidencian detalles de su morfología. Imagénes tomadas en microscopio invertido Nikon
Eclipse TE2000S a una resolución de 1000 x y objetivos en DIC.

48
Tabla 2. Fichas taxonómicas de las características morfológicas de microalgas nocivas en el Golfo de Penas y
Canales aledaños.
Taxa Morfología Rango celular
(µm)
Ref. Fig. 24
Cerataulina pelagica
Diatomea céntrica: Frústulos cilíndricos, que se unen formando
filamentos deleznables. Pared celular Hialina, sin
ornamentación visible al microscopio óptico. Valva con dos
elevaciones que sirven para unir las células y dispuestas de tal
manera que la cadena presenta rotación sobre su eje pervalvar.
Diametro = 7 - 56
µm; axis pervalvar
= 23-120 µm
A
Chaetoceros convolutus
Diatomea céntrica: Existe una cierta variabilidad en la
orientación de las setas. Aunque generalmente estas son
divergentes, a veces sus extremos distales se orientan paralelos
al eje del filamento. El tamaño de la cintura puede alcanzar
hasta la mitad del eje pervalvar. Los prehensores (espinas) en
la base de las setas permiten que estas se mantengan cruzadas.
Tiene un área silícea circular finamente perforada en el
nacimiento de las setas. Valvas subcirculares dimórficas, la
superiores convexas, las inferiores planas, setas circulares en
sus bases, luego cuadrangulares, con espinas más o menos
robustas en los márgenes.
Eje apical = 10 - 27
μm; B
Leptocylindrus danicus
Diatomea centrica: Células cilíndricas con frústulos bien
silicificados, mayor diámetro valvar y presencia de numerosos
cloroplastos de reducido tamaño en cada célula, caracteres
reconocibles con microscopía fotónica. Se diferencia también
por sus aréolas de mayor tamaño y por la presencia de un grupo
de aréolas en el área central, separadas de las restantes por un
anillo silícico. Células unidas por toda la cara valvar, forma
filamentos, pared celular hialina, sin ornamentaciones visibles
microscopia óptica, bandas intercalares difíciles de ver al
microscopio óptico.
Diámetro: 5-16
µm; eje pervalvar:
22-48 µm
C
L.mediterraneus Diatomea Céntrica: Cilíndrica- Pared celular de estructura
gruesa, célula con muchos cloroplastos- unidas en cadena.
Diametro = 7-35
µm C
L. minimus
Diatomeas Céntricas: Células cilíndricas, muy poco
silicificadas, usualmente solitarias aunque también se las
encontró unidas por la cara valvar en cadenas rectas, cortas, de
hasta tres células. Uno a dos (generalmente dos) cloroplastos
de contorno alargado en cada célula. Cingulum formado por
numerosas bandas del tipo media-banda, de contorno
trapezoidal, 1-3 µm de ancho, todas con la misma estructura
consistente en líneas verticales, algo curvadas, de pequeñas
aréolas, 11-14 en 1 µm . Cada individuo posee valvas convexas,
pero una de ellas es cóncava en su parte media, lo que facilita
la unión entre dos células contiguas; el centro de cada valva
parece estar más silicificado.
Diámetro: 1,5-5,2
µm; eje pervalvar:
22-48 µm C
Pseudo-nitzschia
australis
Diatomea penada, eje transapical más ancho que 3um, espacio
central mayor ausente, contorno de la valvas simétrica en el eje
apical, presencia de poroides en las estrías no discernibles con
el microscopio óptico, eje transapical más ancho que 5 µm,
valva apical terminada en punta, el contorno de las valvas más
pequeñas tiende a ser asimétrico en el eje apical.
Eje transapical =
6,5-8 µm; eje
apical = 75-144 µm D

49
Taxa Morfología Rango celular (µm) Ref. Fig. 24
P. delicatissima
Diatomea Penada: Nódulo central presente, costillas
transapicales más numerosas que las fibulae, difícilmente
contables o simplemente no visibles en las preparaciones,
forma valvar diferente, valvas típicamente linear-
lanceoladas.
Eje transapical = 0,7-1,5
µm; eje apical = 34-76
µm
D
P. fraudulenta
Diatomea Penada: valvas lancealada a fusiforme, nódulos
central presente, fibulae y costillas transapicales en igual
número, las ultimas fácilmente visibles si se utiliza técnicas
adecuadas de microscopia.
Eje transapical = 4,5-6,5
µm; eje apical= 64- 111
µm D
P. pseudodelicatissima
Diatomea Penada: Nódulo central presente, costillas
transapicales mas numerosas que las fibulae, difícilmente
contables o simplemente no visibles en las preparaciones,
forma valvar diferente, Eje apical mayor 34 µm, valvas
lineares o sublineares, valvas no ensanchadas en su parte
media
Eje transapical = 1,5-2,5
µm; eje apical= 59- 140
µm D
P. pungens Diatomea Penada: Nódulo central ausente, valvas lineares a
linear-lanceoladas con extremos agudos.
Eje transapical 4-6,5 μm;
eje apical= 68-140 µm D
P. seriata Diatomea Penada: Nódulo central ausente, valvas
lanceoladas a linear-lanceoladas con extremos redondeados.
Eje transapical 6-14,4 μm;
eje apical= 61-160 µm D
Pseudonitzschia sp. Diatomea Penada: Célula fusiforme- lateralmente simétrica,
con frústulos elongados
Eje transapical= 3µm; eje
apical = 40 -50µm D
P.subpacifica
Diatomea Penada: Nódulo central presente, costillas
transapicales más numerosas que las fibulae, difícilmente
contables o simplemente no visibles en las preparaciones,
valvas con un lado fuertemente convexo, el otro más o menos
derecho.
Eje transapical = 5-7 µm;
eje apical no superior a
32μm
D
Skeletonema spp
Diatomea céntrica: Células unidas, formando largas cadenas
rectas. Uno o dos cloroplastos largos por célula. Valvas
circulares. Areolas de la cara valvar dispuestas en filas
radiales. Anillo de procesos marginales que permiten la
unión de las células. Un proceso labiado por valva, con
estructura distinta según se trate de valvas intercalares o
terminales.
Diametro = 2-38 µm; eje
pervalvar 2-61 µm E
Alexadrium minimun
Dinoflagelado tecado: Especie pequeña, algo irregularmente
ovalada, a veces un poco más larga que ancha. Sin espinas ni
cuernos presentes. Cingulum profundamente excavado, su
extremo derecho desplazado posteriormente. Placa de poro
apical (Po) grande, con un agujero central en forma de coma.
Primera placa apical de romboidal en contacto directo o
indirecto con Poro apical. Sexto precingular estrecho. Sulcus
poco profundo. Placa sulcal posterior (S.p.) más ancha que
larga. Delgadas paredes tecales. Un retículo muy débil,
irregular e incompleto se ve a menudo en algunas placas.
Ancho= 18-32 µm; largo
20-48 µm F
Alexandrium ostenfeldii
Dinoflagelado tecado: Las células son grandes y casi
esféricas. Las células son simples. Epiteca e hipoteca iguales
en altura. Esta especie tiene placas delgadas y un gran poro
ventral característico en la primera placa apical. Los débiles
poros superficiales son numerosos y desigualmente
distribuidos.
Ancho= 40-50 µm; largo=
40-56 µm F
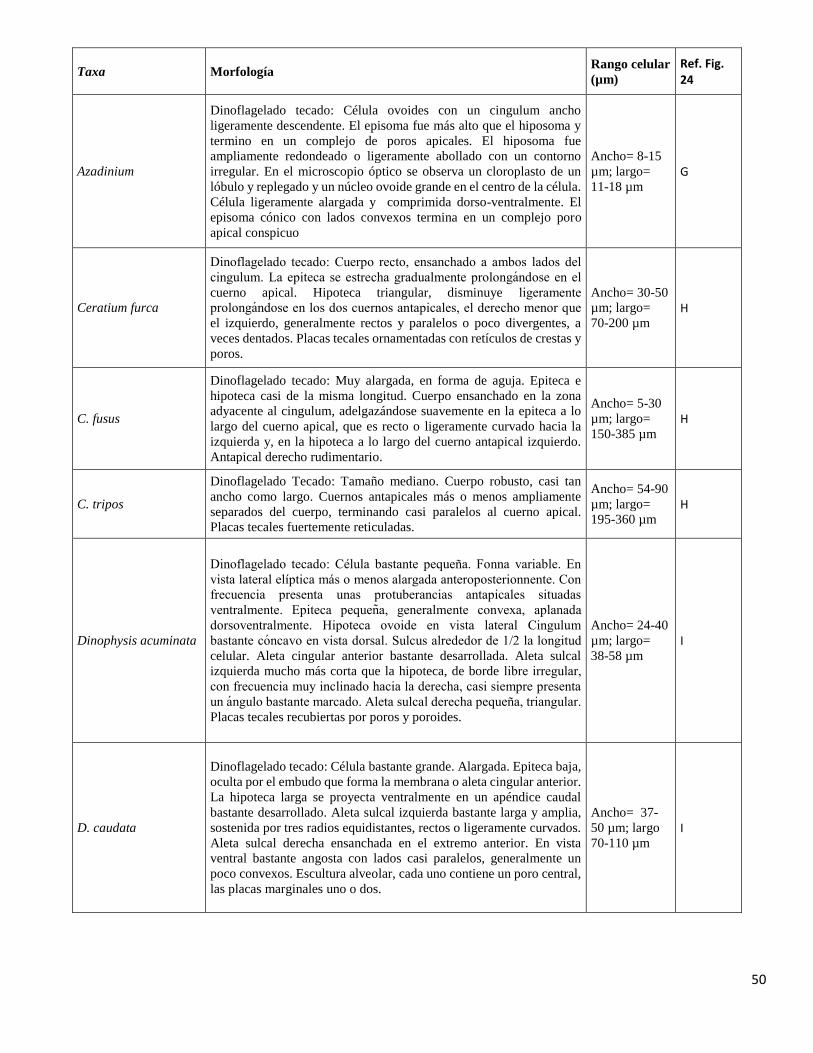
50
Taxa Morfología Rango celular
(µm)
Ref. Fig. 24
Azadinium
Dinoflagelado tecado: Célula ovoides con un cingulum ancho
ligeramente descendente. El episoma fue más alto que el hiposoma y
termino en un complejo de poros apicales. El hiposoma fue
ampliamente redondeado o ligeramente abollado con un contorno
irregular. En el microscopio óptico se observa un cloroplasto de un
lóbulo y replegado y un núcleo ovoide grande en el centro de la célula.
Célula ligeramente alargada y comprimida dorso-ventralmente. El
episoma cónico con lados convexos termina en un complejo poro
apical conspicuo
Ancho= 8-15
µm; largo=
11-18 µm
G
Ceratium furca
Dinoflagelado tecado: Cuerpo recto, ensanchado a ambos lados del
cingulum. La epiteca se estrecha gradualmente prolongandose en el
cuerno apical. Hipoteca triangular, disminuye ligeramente
prolongandose en los dos cuernos antapicales, el derecho menor que
el izquierdo, generalmente rectos y paralelos o poco divergentes, a
veces dentados. Placas tecales ornamentadas con retículos de crestas y
poros.
Ancho= 30-50
µm; largo=
70-200 µm H
C. fusus
Dinoflagelado tecado: Muy alargada, en forma de aguja. Epiteca e
hipoteca casi de la misma longitud. Cuerpo ensanchado en la zona
adyacente al cingulum, adelgazándose suavemente en la epiteca a lo
largo del cuerno apical, que es recto o ligeramente curvado hacia la
izquierda y, en la hipoteca a lo largo del cuerno antapical izquierdo.
Antapical derecho rudimentario.
Ancho= 5-30
µm; largo=
150-385 µm
H
C. tripos
Dinoflagelado Tecado: Tamaño mediano. Cuerpo robusto, casi tan
ancho como largo. Cuernos antapicales más o menos ampliamente
separados del cuerpo, terminando casi paralelos al cuerno apical.
Placas tecales fuertemente reticuladas.
Ancho= 54-90
µm; largo=
195-360 µm H
Dinophysis acuminata
Dinoflagelado tecado: Célula bastante pequeña. Fonna variable. En
vista lateral elíptica más o menos alargada anteroposterionnente. Con
frecuencia presenta unas protuberancias antapicales situadas
ventralmente. Epiteca pequena, generalmente convexa, aplanada
dorsoventralmente. Hipoteca ovoide en vista lateral Cingulum
bastante concavo en vista dorsal. Sulcus alrededor de 1/2 la longitud
celular. Aleta cingular anterior bastante desarrollada. Aleta sulcal
izquierda mucho más corta que la hipoteca, de borde libre irregular,
con frecuencia muy inclinado hacia la derecha, casi siempre presenta
un angulo bastante marcado. Aleta sulcal derecha pequeña, triangular.
Placas tecales recubiertas por poros y poroides.
Ancho= 24-40
µm; largo=
38-58 µm
I
D. caudata
Dinoflagelado tecado: Célula bastante grande. Alargada. Epiteca baja,
oculta por el embudo que forma la membrana o aleta cingular anterior.
La hipoteca larga se proyecta ventralmente en un apéndice caudal
bastante desarrollado. Aleta sulcal izquierda bastante larga y amplia,
sostenida por tres radios equidistantes, rectos o ligeramente curvados.
Aleta sulcal derecha ensanchada en el extremo anterior. En vista
ventral bastante angosta con lados casi paralelos, generalmente un
poco convexos. Escultura alveolar, cada uno contiene un poro central,
las placas marginales uno o dos.
Ancho= 37-
50 µm; largo
70-110 µm
I

51
Taxa Morfología Rango celular
(µm)
Ref. Fig. 24
D. norvegica
Dinoflagelado tecado: célula grande y robusta con una
curvatura dorsal totalmente redondeada y un perfil lateral
recto a la mitad inferior del margen ventral. La parte más
profunda de la célula está a medio camino entre la línea
cingular inferior y el antapex. La superficie de la placa, con
grandes areolas con poros. Márgenes posteriores y dorsales
a veces con protuberancias o extensiones gruesas llamadas
bultos.
Ancho= 30-50
µm; largo 40-80
µm
I
D. tripos
Dinoflagelado tecado: Célula grande, con uno o dos
apéndices caudales robustos, parecida a D. caudata, pero casi
siempre de facil diferenciacion por un segundo apendice
caudal dorsal, mucho mas corto que el ventral. Este apendice
dorsal es, empero, muy variable y, en algunos ejemplares,
apenas rudimentario, cuando no ausente. Pero aun los
individuos con atrofia casi completa del apendice dorsal se
pueden diferenciar de D. caudata por la fuerte concavidad de
la parte anterior del borde dorsal de la hipoteca y por el
extremo postero-dorsal anguloso.
Ancho= 50-60
µm; largo= 90-
125 µm
I
D. dens
Dinoflagelado tecado: especies de tamaño pequeño a
mediano con margen dorsal curvado y margen ventral
posterior angulado. Célula puede parecer a una pequeña D.
Acuta, pero no tiene la prominente profundidad postmediana
que caracteriza D. Acuta. La línea sulcal izquierda tiene
típicamente marcas superficiales como reticulaciones o
vermiculas, la profundidad de la linea es desigual y el borde
distal está parcialmente curvado.
Ancho= 28 µm;
largo= 45-60 µm I
Gonyaulax polygramma
Dinoflagelado tecado: Célula de tamaño mediano. Forma
alargada, pentagonal o biconica irregular. La epiteca mayor
que la hipoteca convexa o angulosa, terminada en un cuerno
apical bastante alto y grueso. Hipoteca mas o menos
simétrica, redondeada o truncada, con numero variable de
espinas antapicales, a veces con dos, de las que la izquierda
suele ser mayor. Cingulum descendente, desplazado
alrededor de 1.5 veces su altura. Sulcus ensanchado
posteriormente. Nucleo largo, ovoide, situado en la parte
posterior de la celula. Placas ornamentadas, predominando
las crestas verticales y alveolos mas bien pequenos.
Ancho= 26-50
µm; largo 50-70
µm
J
Gymnodinium aureolum
Célula de tamaño mediano. Forma variable redonda-ovalada,
algo aplanada dorso- ventralmente. En vista dorso-ventral, el
epicono es redondeado en su contorno superficial, de forma
tronco-conica. Hipocono de lados convexos, aplanado en su
extremo antapical, formando una pequena muesca o
hendidura donde el sulcus se une al antapex. El epicono es
normalmente mas pequeno que el hipocono. Cingulum
profundamente descendente y ancho, que se prolonga en el
epicono como un surco mas estrecho, ocupando tres cuartos
de su longitud. Cromatoforos pardo-amarillentos, elipticos o
irregulares, numerosos y distribuidos por toda la celula.
Nucleo largo, situado en la region cingular, extendiendose
hacia el epicono e hipocono.
Ancho= 16-31
µm; largo 25-40
µm K

52
Taxa Morfología Rango celular (µm) Ref. Fig. 24
G. simplex
Pequena, claramente elipsoidal. Epicono estrecho y corto e
hipocono circular en sección transversal, un poco aplanado en
sentido horizontal. Cingulum situado un poco por encima del
centro de la celula. SuIcus muy superficial no siempre
perceptible, no se extiende hacia el epicono. Extremo anterior
de la celula redondeado. Hipocono de lados redondeados,
aplanado en su extremo antapical, formando una pequena
muesca donde el sulcus se une al antapex. Nucleo esferico,
central o ligeramente situado en posicion posterior.
Cloroplastos verde-amarillentos normalmente presentes en
numero de 2 a 4 situados en la perisferia de la celula,
ocasionalmente algunos de ellos pueden estar ausentes.
Tricocistos presentes.
Ancho= 6-10 µm;
largo 8-14 µm K
Gymnodinium sp.
Dinoflagelados atecados: Celulas generalmente globulares u
ovoides, eventualmente comprimidas dorso- ventralmente, de
fomas y tamanos muy variables. El cingulum cerrado o abierto
puede dar una vuelta o mas alrededor de la celula, situado en
posicion central o desplazada no mas de 1/5 de la longitud total
del cuerpo, hacia el extremo anterior o posterior de la celula.
El su1cus puede extenderse del apex al antapex o puede ser
incompleto, restringido al hipocono, al epicono (se emplean
estos terminos al no poseer una verdadera teca) o a ambos,
normalmente sin torsión. Hay especies con cloroplastos y otras
sin ellos. Nucleo normalmente en posicion central, siendo su
posicion uno de los caracteres utilizados en las
detenninaciones especificas.
Ancho : 20 -50 µm;
largo 22-70 µm K
Heterocapsa
Dinoflagelado tecado: Celula muy pequena. Biconica,
achatada dorsoventralmente. Epiteca de lados mas o menos
rectos, o algo sinuosos, asimetricos, terminada en punta roma.
Hipoteca conica terminada en punta diferenciada, como un
pequeno cuerno. Cingulum excavado, descendente,
desplazado. Sulcus ancho y corto, no llega al antapex. Nucleo
situado en la epiteca.
Ancho= 6-22 µm;
largo 10-33 µm L
Karenia bicuneiformis
Dinoflagelado atecado: Presenta el epicono cónico, algunas
veces redondeado no presenta carena, el núcleo es ovalado y
ocupa la parte de la izquierda del hipocono, el cual está
dividido en 2 extensiones con extremidades acentuadas en
forma de `W' que excede la longitud del epicono, muestra
numerosos cloroplastos distribuidos a través de toda la célula.
Ancho= 30-34µm;
largo= 32-38 µm M
K. brevisulcata
Dinoflagelado atecado: Célula con desplazamiento del
cíngulum (proporción de la longitud de la célula) 11-27%,
pequeña extensión del sulcus, triangular. Ranura apical lineal,
muy corta; extendiéndose 1/3 a 1/2 en el lado ventral, 1/3 o
menos hacia abajo del lado dorsal de la epicono. El núcleo
esférico u ovoide y localizado en el lado izquierdo del
hipocono a horizontalmente alargado de izquierda a derecha
del hipocono. Posee dos a numeroso cloroplastos de forma
alargada
Ancho= 10-22 µm
largo= 18-37µm N
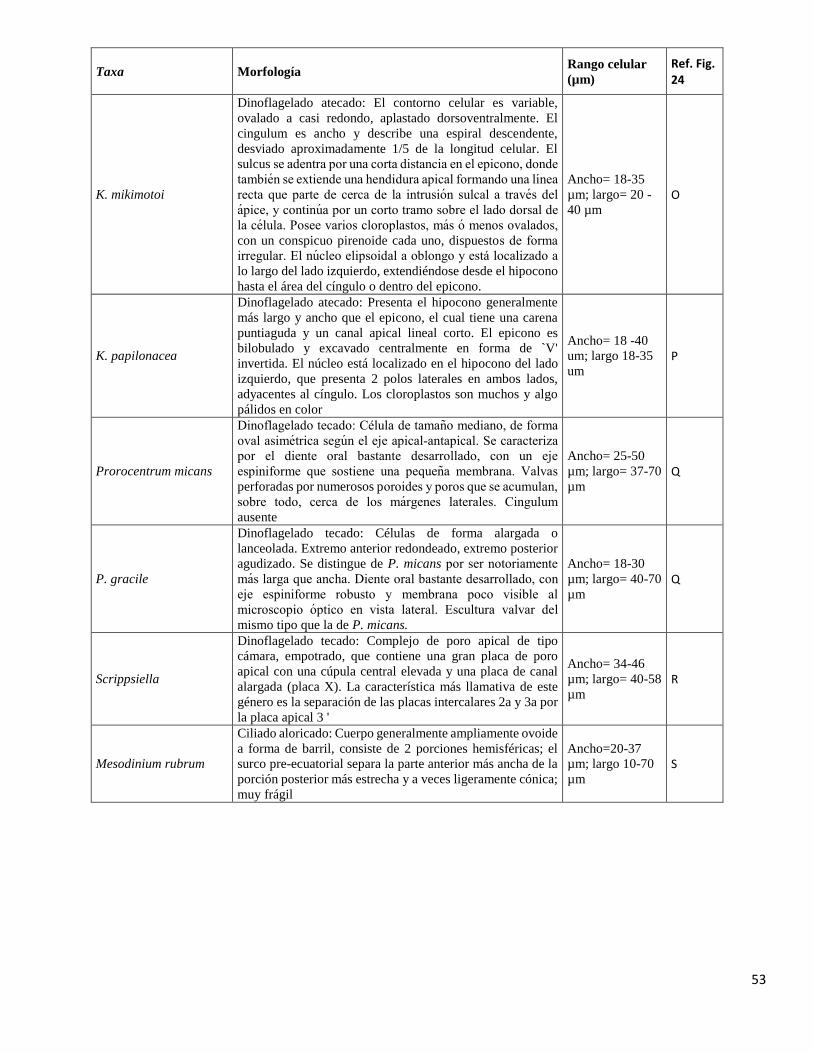
53
Taxa Morfología Rango celular
(µm)
Ref. Fig. 24
K. mikimotoi
Dinoflagelado atecado: El contorno celular es variable,
ovalado a casi redondo, aplastado dorsoventralmente. El
cingulum es ancho y describe una espiral descendente,
desviado aproximadamente 1/5 de la longitud celular. El
sulcus se adentra por una corta distancia en el epicono, donde
tambien se extiende una hendidura apical formando una linea
recta que parte de cerca de la intrusion sulcal a traves del
apice, y continua por un corto tramo sobre el lado dorsal de
la celula. Posee varios cloroplastos, mas o menos ovalados,
con un conspicuo pirenoide cada uno, dispuestos de forma
irregular. El nucleo elipsoidal a oblongo y está localizado a
lo largo del lado izquierdo, extendiéndose desde el hipocono
hasta el área del cíngulo o dentro del epicono.
Ancho= 18-35
µm; largo= 20 -
40 µm
O
K. papilonacea
Dinoflagelado atecado: Presenta el hipocono generalmente
más largo y ancho que el epicono, el cual tiene una carena
puntiaguda y un canal apical lineal corto. El epicono es
bilobulado y excavado centralmente en forma de `V'
invertida. El núcleo está localizado en el hipocono del lado
izquierdo, que presenta 2 polos laterales en ambos lados,
adyacentes al cíngulo. Los cloroplastos son muchos y algo
pálidos en color
Ancho= 18 -40
um; largo 18-35
um
P
Prorocentrum micans
Dinoflagelado tecado: Célula de tamano mediano, de forma
oval asimetrica segun el eje apical-antapical. Se caracteriza
por el diente oral bastante desarrollado, con un eje
espiniforme que sostiene una pequena membrana. Valvas
perforadas por numerosos poroides y poros que se acumulan,
sobre todo, cerca de los margenes laterales. Cingulum
ausente
Ancho= 25-50
µm; largo= 37-70
µm
Q
P. gracile
Dinoflagelado tecado: Células de forma alargada o
lanceolada. Extremo anterior redondeado, extremo posterior
agudizado. Se distingue de P. micans por ser notoriamente
mas larga que ancha. Diente oral bastante desarrollado, con
eje espiniforme robusto y membrana poco visible al
microscopio optico en vista lateral. Escultura valvar del
mismo tipo que la de P. micans.
Ancho= 18-30
µm; largo= 40-70
µm
Q
Scrippsiella
Dinoflagelado tecado: Complejo de poro apical de tipo
cámara, empotrado, que contiene una gran placa de poro
apical con una cúpula central elevada y una placa de canal
alargada (placa X). La característica más llamativa de este
género es la separación de las placas intercalares 2a y 3a por
la placa apical 3 '
Ancho= 34-46
µm; largo= 40-58
µm
R
Mesodinium rubrum
Ciliado aloricado: Cuerpo generalmente ampliamente ovoide
a forma de barril, consiste de 2 porciones hemisféricas; el
surco pre-ecuatorial separa la parte anterior más ancha de la
porción posterior más estrecha y a veces ligeramente cónica;
muy frágil
Ancho=20-37
µm; largo 10-70
µm
S

54
Tabla 3. Abundancia integrada (0-30 m) de grupos funcionales (células vivas x106m-2), en 21 estaciones del Golfo de Penas y canales
aledaños. Cada grupo ha sido separado en sus principales componentes: Diatomeas penadas y céntricas, Dinoflagelados tecados y
atecados y Ciliados loricados y aloricados.
sil icoflagelados Aundancia Total
Estacion Centricas Penadas Totales Atecados Tecados Totales Loricado Aloricado Totales Totales Desnudos Total (106 cel m-2)
E13 147 30 177 107 41 149 3 1 3 2 0 2 331
E14 201 113 315 240 114 353 15 16 31 8 1 9 707
E15 157 74 231 354 316 670 20 22 42 5 3 8 950
E16 73 67 139 97 50 147 16 1 17 0 0 0 303
E17 58 102 160 85 42 127 13 2 15 1 1 2 304
E18 588 224 813 166 145 312 14 1 14 0 2 2 1141
E19 730 352 1082 48 22 70 20 3 23 1 0 1 1176
E20 634 200 834 49 29 79 9 0 10 0 2 2 924
E21 66 18 85 4 14 18 3 0 4 0 0 0 106
E22 1204 67 1271 5 105 110 2 1 3 0 0 0 1384
E23 29 47 75 18 24 42 2 1 3 0 0 0 120
IN 1285 565 1849 21 36 57 5 2 7 6 0 6 1919
SNP 3 623 115 738 6 14 20 2 1 2 1 2 3 762
SNP 8 24 20 44 101 95 196 8 2 10 2 0 2 253
SNP 9 81 107 189 207 125 332 13 4 17 4 0 5 542
SNP4 212 15 228 230 121 351 8 5 12 5 0 6 597
SNP5 5 14 19 298 300 598 15 5 21 1 1 2 639
SNP6 15 63 78 174 138 312 3 1 4 2 1 2 396
SNP7 145 293 439 293 170 463 33 3 36 3 1 4 942
SPN 10 53 41 94 195 95 291 16 3 19 5 3 8 412
SNP1 1064 75 1139 13 15 28 3 3 6 0 0 0 1173
flageladosCiliados Dinoflagelados Diatomeas

55
Tabla 4. Abundancia (cél. /L) de los principales géneros y especies de Diatomeas y Dinoflagelados nocivos de las estaciones SNP1 a la E17 en la
zona del Golfo de Penas y Canales aledaños.
SNP1 SNP3 IN SNP4 SNP10 SNP9 E13 E14 E15 E16 E17
Taxa 5 15 30 5 15 30 5 15 30 5 15 30 5 15 30 5 15 30 5 15 30 5 15 30 5 15 30 5 15 30 5 15 30
Cerataulina pelagica 2824 0 0 2017 81 7261 2541 145 242 0 0 0 0 0 0 0 0 605 48 0 403 0 121 121 0 0 0 0 0 0 0 0
Chaetoceros convolutus 0 0 0 0 0 0 0 0 0 40 0 0 0 40 0 0 81 121 0 30 0 104 0 242 109 0 0 0 0 0 0 38
Leptocylindrus danicus 807 0 0 605 0 0 242 0 1452 0 0 242 242 0 1452 1380 0 61 0 0 0 104 0 0 0 0 1714 605 0 202 605 0
Leptocylindrus mediterraneus 0 0 0 0 0 0 0 218 0 40 0 0 0 1130 0 0 40 0 0 30 0 104 161 0 0 0 0 0 945 0 81 719
Leptocylindrus minimus 0 0 0 0 0 0 0 0 0 0 0 0 323 0 0 1017 0 0 0 0 0 0 0 0 0 0 0 35 227 0 0 0
Pseudo-nitzschia Austral 0 0 0 0 0 0 0 0 0 0 0 0 0 40 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Pseudo-nitzschia delicatissima 2219 0 0 1210 121 6777 242 0 81 121 0 121 161 0 1210 2324 0 545 0 0 7766 207 0 1089 218 0 656 588 38 444 565 0
Pseudo-nitzschia fraudulenta 0 0 43 1210 0 0 242 1307 81 0 40 61 121 726 81 73 323 545 0 545 0 0 524 0 327 81 151 0 303 0 0 681
P. pseudodelicatissima 2622 48 0 4236 484 8230 10650 0 403 40 0 666 766 0 2017 2469 0 847 242 0 2723 622 645 3873 218 0 1714 242 113 2017 1493 151
Pseudonitzschia pungens 807 97 0 807 40 968 242 581 403 0 81 61 20 766 565 145 484 121 242 121 1614 0 1654 1694 436 242 555 0 605 928 242 643
Pseudonitzschia seriata. 101 0 0 403 444 1694 6414 436 242 0 0 0 0 81 0 0 0 545 0 0 1210 0 645 726 0 40 0 0 454 0 0 416
Pseudonitzschia sp. 0 0 0 0 0 0 0 218 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Pseudonitzschia subpacifica 0 0 0 0 0 0 242 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 81 81 38
Skeletonema spp 75034 1210 0 62327 6898 70919 34250 0 29288 2501 0 424 403 0 0 0 81 10953 2517 0 16136 0 0 15007 327 0 605 0 0 81 81 0
Alexadrium minimun 0 0 0 0 0 0 0 0 888 171 0 424 40 0 242 0 0 61 0 0 202 385 0 0 297 0 454 0 0 0 0 0
Alexadrium ostenfeldii 0 0 0 0 0 0 0 0 0 30 0 0 0 0 0 0 202 0 0 61 0 30 0 0 30 40 0 0 0 0 0 38
Azadinium 0 0 0 0 0 0 0 0 5164 3389 0 2541 1977 0 2743 2905 121 787 1404 0 1009 4461 0 12949 12853 40 4790 311 0 484 686 0
Ceratium furca 0 0 0 0 0 121 81 218 0 121 1735 0 0 2461 81 101 1372 0 0 514 50 0 323 0 0 4518 50 17 983 40 81 1135
Ceratium fusus 0 0 0 0 0 121 0 0 81 40 40 121 0 121 121 0 0 61 0 0 50 0 0 0 0 0 50 0 38 81 0 38
Ceratium tripos 0 0 0 0 0 0 0 0 81 0 40 0 0 81 0 0 0 0 0 0 50 0 0 0 0 40 0 0 76 0 0 0
Dinophysis acuminata 202 48 43 0 81 726 121 0 242 0 0 0 40 0 0 73 0 0 0 0 0 0 0 61 0 0 0 0 0 40 0 0
Dinophysis caudata 101 0 0 0 0 0 0 0 0 0 0 0 0 40 0 0 0 0 0 0 0 0 40 0 0 0 0 0 38 0 0 0
Dinophysis norvegica 0 0 0 202 0 968 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Dinophysis tripos 0 0 0 0 0 0 0 0 40 81 0 61 0 0 282 0 0 121 0 0 50 104 0 61 109 0 50 0 0 0 121 0
Dynophysis dens 0 0 0 101 0 484 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Gonyaulax polygramma 0 0 0 0 0 0 0 0 0 0 0 0 0 282 0 0 40 0 0 0 0 0 0 0 0 81 0 0 38 0 0 38
Gymnodinium aureolum 0 48 86 0 48 0 169 0 0 1759 1049 0 1194 1049 0 2004 282 0 891 363 0 2614 202 242 2244 1089 101 304 719 0 581 416
Gymnodinium simplex 0 61 0 0 61 0 212 0 0 2199 40 0 1493 121 0 2505 0 0 1113 30 0 3268 0 0 2805 0 0 380 0 0 726 38
Gymnodinium sp. 403 133 0 0 133 0 466 0 9278 4837 0 7503 3284 0 4881 5511 0 3207 2450 0 3328 7189 0 12465 6170 0 5950 837 0 2299 1598 0
Heterocapsa 0 0 0 0 40 0 0 0 0 161 0 0 323 0 40 290 40 0 0 0 50 207 121 61 109 0 50 0 0 20 0 0
Karenia bicuneiformis 0 0 0 0 0 0 0 290 161 40 0 666 81 0 202 73 81 0 0 0 50 0 0 1694 109 81 353 0 0 0 0 0
Karenia brevisulcata 0 0 0 0 0 0 0 0 0 40 0 0 20 0 0 218 121 0 0 0 0 0 0 61 0 282 151 0 76 0 0 0
Karenia mikimotoi 0 48 0 0 0 0 0 0 403 282 0 182 81 40 121 145 0 121 339 0 101 934 0 242 1961 161 50 69 113 40 202 0
Karenia Papilonacea 0 0 0 0 0 0 0 0 0 0 40 30 40 161 40 73 161 0 0 0 0 104 0 0 327 1170 0 17 227 0 0 38
Prorocentrium micans 0 0 0 0 40 242 0 0 0 0 81 0 14 242 0 24 121 0 0 30 0 101 0 0 0 161 0 0 0 0 0 38
Prorocentrum gracile 202 0 0 202 0 0 0 0 0 0 0 0 0 0 40 0 0 0 0 0 0 0 0 121 0 0 0 0 0 0 0 0
Scrippsiella 0 0 0 0 0 0 121 0 0 0 0 0 0 0 242 218 0 0 0 0 0 207 0 0 218 0 0 35 0 81 121 0
Mesodinium rubrum 101 48 0 0 40 0 121 0 0 0 0 61 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0

56
Tabla 4 (continuación). Abundancia (cél. /L) de los principales géneros y especies de Diatomeas y Dinoflagelados nocivos de las estaciones SNP5
a la E23 en la zona del Golfo de Penas y Canales aledaños.
SNP5 SNP6 SNP7 SNP8 E18 E19 E20 E21 E22 E23
Taxa 5 15 30 5 15 30 5 15 30 5 15 30 5 15 30 5 15 30 5 15 30 5 15 30 5 15 30 5 15 30
Cerataulina pelagica 0 0 0 0 0 0 0 0 0 0 0 0 0 0 151 0 121 168 24 387 0 65 0 0 194 48 0 0 0 0
Chaetoceros convolutus 0 0 0 0 0 0 0 0 0 0 0 0 0 0 303 45 121 0 0 0 48 0 0 0 0 0 0 0 0 0
Leptocylindrus danicus 0 0 0 0 0 0 605 1037 0 0 145 0 1045 61 605 2935 12042 168 1404 36985 0 452 0 0 0 0 0 32 69 0
Leptocylindrus mediterraneus 0 0 0 0 0 562 0 0 3026 0 0 242 0 0 57335 0 0 36979 0 0 3485 0 0 323 0 0 0 0 0 73
Leptocylindrus minimus 0 0 0 0 0 0 0 0 0 0 0 161 0 0 303 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Pseudo-nitzschia Austral 0 0 0 0 0 0 0 0 0 0 0 0 0 0 151 0 0 336 0 0 145 0 0 65 0 0 0 65 0 24
Pseudo-nitzschia delicatissima 605 0 0 2420 48 0 605 363 0 888 48 0 275 0 3782 271 1271 9245 145 387 339 0 41 65 97 0 0 0 0 0
Pseudo-nitzschia fraudulenta 0 0 692 0 48 346 0 311 12506 0 0 403 0 0 4841 1039 1573 2521 0 3582 581 0 0 32 0 242 0 0 0 0
P. pseudodelicatissima 0 97 0 0 629 43 4841 4979 0 242 145 0 110 30 1967 768 1089 1849 145 5131 0 0 28 0 775 0 0 129 14 0
Pseudonitzschia pungens 202 0 0 202 145 2593 908 674 2824 40 0 323 0 0 7413 271 424 9077 242 1646 484 0 0 290 0 145 129 0 0 48
Pseudonitzschia seriata. 0 0 0 0 0 303 0 0 807 0 0 0 0 0 3933 0 0 4370 24 194 97 0 0 65 0 0 0 0 0 24
Pseudonitzschia sp. 0 0 0 0 0 0 0 0 0 0 0 0 0 0 303 0 0 336 0 0 0 0 0 0 0 0 0 0 0 0
Pseudonitzschia subpacifica 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 387 0 0 0 0 0 0 0 0 28 0
Skeletonema spp 0 0 0 0 0 0 0 0 202 1654 194 0 0 242 0 90 303 0 436 3873 48 0 235 161 775 0 0 323 0 0
Alexadrium minimun 0 0 0 0 0 0 303 0 0 403 0 0 825 30 151 0 0 0 0 0 0 0 0 0 387 0 0 0 0 0
Alexadrium ostenfeldii 202 0 216 0 0 0 0 0 1009 0 0 0 0 0 303 0 0 0 0 0 145 0 0 710 0 0 12909 0 0 2154
Azadinium 27230 6148 0 10690 2590 43 3177 4046 605 3671 2711 0 12927 121 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Ceratium furca 0 0 6094 0 0 4409 151 156 4639 40 0 2743 55 30 5597 316 61 0 48 194 145 65 14 0 0 0 0 0 0 0
Ceratium fusus 0 0 0 0 0 0 0 52 101 0 0 0 110 0 0 45 0 84 24 48 0 161 28 0 0 0 0 32 28 0
Ceratium tripos 0 0 0 0 0 86 0 0 101 0 0 40 0 0 0 45 0 0 0 0 0 129 0 16 97 0 0 32 0 0
Dinophysis acuminata 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 45 61 0 0 48 0 194 0 0 97 0 0 0 0 0
Dinophysis caudata 101 0 0 0 48 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 14 0 0 0 0 0 0 0
Dinophysis norvegica 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 61 0 0 0 0 0 0 0 0 0 0 0 0 0
Dinophysis tripos 0 48 0 0 24 0 151 259 0 0 24 0 55 61 76 45 61 0 24 0 0 0 0 97 0 0 0 0 0 0
Dynophysis dens 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Gonyaulax polygramma 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 84 0 0 48 0 0 0 0 0 0 0 0 0
Gymnodinium aureolum 807 1075 475 0 460 821 0 1909 2017 0 484 524 0 375 605 0 254 168 0 58 145 0 0 32 0 10 0 0 83 73
Gymnodinium simplex 0 1343 0 0 575 0 0 2386 0 0 605 0 0 469 0 0 318 0 0 73 0 0 0 0 97 12 0 0 104 0
Gymnodinium sp. 22994 2955 0 12506 1265 0 8018 5249 0 4720 1331 0 12817 1032 0 406 699 0 920 160 0 0 0 0 0 27 0 742 228 0
Heterocapsa 403 290 0 0 0 0 151 0 0 0 411 0 110 272 76 45 0 84 0 0 0 0 0 0 0 0 0 0 0 0
Karenia bicuneiformis 2219 48 562 1009 0 173 0 0 202 403 0 40 440 30 303 226 0 0 97 0 0 0 0 0 0 0 0 0 0 0
Karenia brevisulcata 0 194 0 0 0 0 0 104 101 40 24 40 165 0 303 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0
Karenia mikimotoi 706 533 0 403 169 86 151 674 0 161 97 40 330 182 76 0 303 0 0 2324 0 0 0 0 0 0 0 0 28 0
Karenia Papilonacea 0 0 43 0 0 173 0 0 605 0 0 202 0 0 756 0 0 168 0 48 145 0 0 65 0 0 0 0 0 0
Prorocentrium micans 202 0 43 0 0 43 0 0 403 0 0 40 0 0 151 0 0 84 0 67 97 0 0 32 0 0 0 0 0 0
Prorocentrum gracile 202 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 0 24 0 0 0 0 0 0 0 0 0 0 0
Scrippsiella 202 97 0 202 24 0 0 207 0 40 48 40 0 30 0 0 61 0 0 0 0 0 0 0 0 0 0 0 14 0
Mesodinium rubrum 0 48 0 0 0 0 151 52 0 0 0 0 0 0 0 0 0 84 0 0 0 65 0 0 0 0 0 0 0 0

57
4.5 Resultados de análisis estadísticos
En base a las características espaciales y oceanográficas, se decidió agrupar las estaciones de
muestreos en cuatro zonas: Canal Moraleda, Canal Sur (i.e. canales Messier y Wide), Golfo
de Penas y Península de Taitao (Tabla 5).
Tabla 5. Estaciones de muestreo por zona geográfica.
Estación Zona Latitud (°S) Longitud (°W) Fecha
SNP1 Canal Sur 49,96 74,41 17-02-2017
IN Canal Sur 48,32 74,52 17-02-2017
SNP3 Canal Sur 49,05 74,40 17-02-2017
SNP4 Golfo 47,75 74,72 18-02-2017
SNP10 Golfo 47,52 75,75 18-02-2017
SNP9 Golfo 47,18 75,93 18-02-2017
E13 Golfo 47,59 74,90 19-02-2017
E14 Golfo 47,38 75,04 19-02-2017
E15 Golfo 47,23 75,24 19-02-2017
E16 Golfo 47,13 75,39 19-02-2017
E17 Golfo 47,00 75,52 19-02-2017
SNP5 Golfo 47,31 74,68 19-02-2017
SNP6 Golfo 47,14 74,83 19-02-2017
SNP7 Golfo 46,98 74,93 19-02-2017
SNP8 Golfo 46,73 75,09 19-02-2017
E18 Taitao 46,72 75,73 21-02-2017
E19 Taitao 46,42 75,55 21-02-2017
E20 Taitao 46,12 75,32 21-02-2017
E21 Moraleda 45,82 74,50 21-02-2017
E22 Moraleda 45,28 73,67 21-02-2017
E23 Moraleda 44,54 73,47 21-02-2017
Los valores promedio de las variables ambientales registrados por zona durante el crucero,
se entregan en la Tabla 6. Se puede observar que el Canal Sur presenta salinidad alrededor
de 26, la que es notoriamente más baja que las otras zonas, aunque el Canal Moraleda también
presenta salinidad más baja (~31) que la del Golfo de Penas y Taitao. En ambos canales (i.e.

58
Moraleda, Messier y Wide), además, se registraron aguas de menor temperatura que en el
Golfo de Penas y Taitao, en especial en el Canal Sur (~10 °C), y pH más bajo (en ambos
canales <8). En cuanto a la concentración de nutrientes, en la zona Golfo de Penas y Taitao,
la concentración de nitrato, nitrito, fosfato y ácido silícico fue cerca de la mitad de las
concentraciones determinadas en el Canal Sur y Moraleda (Tabla 6).
Tabla 6. Valores promedio (± D.E.) de las variables ambientales medidas durante el crucero,
agrupadas por zonas. STD = Sólidos Totales Disueltos, NO3- = Nitrato, NO2
- = Nitrito, NI
total = Nitrógeno Inorgánico Total (suma de nitrito y nitrato), PO43- = Fosfato.
Variable Ambiental Canal Sur Golfo de Penas Moraleda Taitao
Salinidad 26.09 ± 1.21 32.52 ± 1.28 31.18 ± 0.95 33.19 ± 0.15
Temperatura (°C) 10.27 ± 1.35 13.39 ± 0.41 12.51 ± 0.27 13.74 ± 0.11
Oxígeno Disuelto (mL L-1) 5.97 ± 0.22 5.53 ± 0.10 5.49 ± 0.23 5.56 ± 0.06
STD (g L-1) 24.02 ± 2.2 31.38 ± 1.21 30.38 ± 0.82 32.06 ± 0.16
Conductividad (mS cm-1) 28.26 ± 2.22 38.34 ± 1.69 37.54 ± 0.82 39.97 ± 0.74
Potencial Ox-Red (mV) 202.99 ± 38.88 147.26 ± 47.42 188.00 ± 63.66 145.22 ± 23.37
pH 7.79 ± 0.27 8.08 ± 0.09 7.92 ± 0.01 8.02 ± 0.02
NO3- (µmol m-2) 0.200 ± 0.91 0.082 ± 0.015 0.229 ± 0.091 0.113 ± 0.023
NO2- (µmol m-2) 0.014 ± 0.005 0.011 ± 0.001 0.015 ± 0.004 0.012 ± 0.001
NI total (µmol m-2) 0.22 ± 96.17 0.093 ± 0.015 0.244 ± 0.094 0.125 ± 0.024
PO43- (µmol m-2) 0.025 ± 0.009 0.015 ± 0.004 0.031 ± 0.006 0.019 ± 0.011
Ácido Silícico (µmol m-2) 0.167 ± 0.022 0.082 ± 0.022 0.130 ± 0.033 0.082 ± 0.040
El análisis de ordenación MDS basada en las variables ambientales con datos promedios e
integrados, muestra que en términos del conjunto total de variables medidas, las estaciones
ubicadas en el canal Sur y en el canal Moraleda tienden a diferenciarse de las estaciones
ubicadas en el Golfo de Penas y Taitao (Fig. 25). El análisis PERMANOVA para estos datos
indica que hay diferencias significativas en la estructura multivariada de las zonas en función
de las variables ambientales (Tabla 7). Adicionalmente, los test Pair Wise indican que no hay
diferencias significativas entre ambos Canales (i.e. Moraleda y Canal sur), ni entre Golfo de
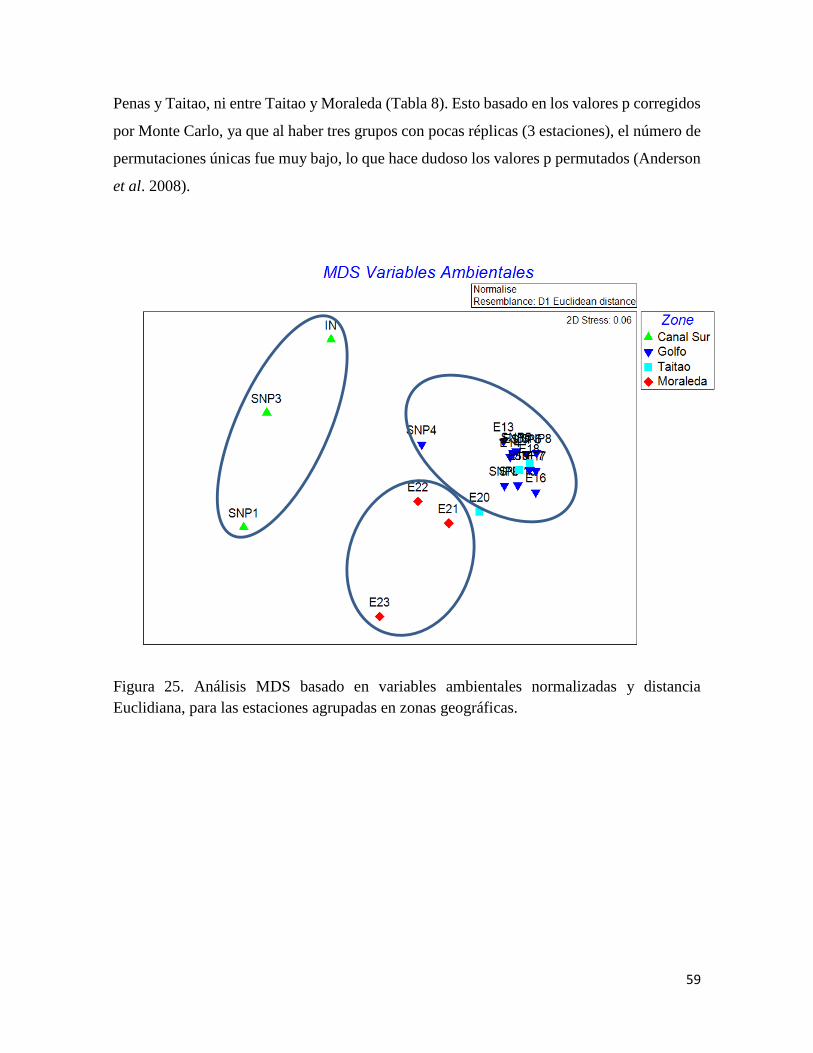
59
Penas y Taitao, ni entre Taitao y Moraleda (Tabla 8). Esto basado en los valores p corregidos
por Monte Carlo, ya que al haber tres grupos con pocas réplicas (3 estaciones), el número de
permutaciones únicas fue muy bajo, lo que hace dudoso los valores p permutados (Anderson
et al. 2008).
Figura 25. Análisis MDS basado en variables ambientales normalizadas y distancia
Euclidiana, para las estaciones agrupadas en zonas geográficas.

60
Tabla 7. Resultados del análisis PERMANOVA, para matriz de distancia Euclidiana basada
en datos ambientales normalizados registrados durante el crucero en las 4 zonas estudiadas.
Valores-p significativos en negrita. P(perm) = valor-p permutacional, P(MC) = valor-p
corregido por Monte Carlo.
Fuente de
Variación
g.l.
SC
CM
Pseudo-F
P(perm)
Permutaciones
únicas
P(MC)
Zona 3 166,9 55,6 10,16 0,0001 9923 0,0001
Residuos 17 93,1 5,5
Total 20 260
Tabla 8. Resultados del test Pair-Wise evaluando diferencias entre cada una de las cuatro
zonas estudiadas en términos de las variables ambientales. Valores-p significativos en
negrita. P(perm) = valor-p permutacional, P(MC) = valor-p corregido por Monte Carlo.
Grupos
T
P(perm)
Permutaciones
únicas
P(MC)
Canal Sur, Golfo 4.93 0.003 455 <0.001
Canal Sur, Taitao 2.93 0.097 10 0.009
Canal Sur, Moraleda 1.89 0.102 10 0.058
Golfo, Taitao 1.01 0.358 455 0.385
Golfo, Moraleda 3.26 0.002 455 <0.001
Taitao, Moraleda 1.84 0.091 10 0.083
Los resultados de los análisis estadísticos (Tabla 5), y considerando solo los datos de
presencia-ausencia de las especies de fitoplancton nocivo en el área de estudio, muestran que
en la zona Golfo de Penas y Península de Taitao se encuentran la mayor cantidad de especies
(35 y 36 especies respectivamente), mientras que en los canales Sur y Moraleda se registraron
27 especies en cada uno. La ordenación por medio del MDS basado en Jaccard, nos sugiere
en base a los parámetros físico-químicos, que el Canal Sur y Moraleda son distintos a las
zonas Golfo de Penas y Península de Taitao (Fig. 26). Concordante con lo anterior, el análisis
PERMANOVA encuentra diferencias significativas en la composición de especies de las

61
zonas (Tabla 9). Además los análisis Pair-Wise encuentran que ambos canales no presentan
diferencias significativas entre sí, y además Taitao tampoco presenta diferencias
significativas con el canal Moraleda, ni con el Golfo de Penas (Tabla 10).
Figura 26. Análisis MDS en base a matriz de similitud de Jaccard, a partir de datos de
presencia-ausencia de fitoplancton nocivo. Las estaciones se muestran agrupadas en zonas
geográficas.

62
Tabla 9. Resultados del análisis PERMANOVA, en base a matriz de similitud de Jaccard, a
partir de datos de presencia-ausencia de fitoplancton nocivo. Factor Zona se refiere a las
zonas geográficas definidas y registradas durante el crucero. Valores-p significativos en
negrita. P(perm) = valor-p permutacional, P(MC) = valor-p corregido por Monte Carlo.
Source
g.l.
SC
CM
Pseudo-F
P(perm)
Permutaciones
únicas
P(MC)
Zona 3 9348.6 3116.2 4.4 0.0001 9908 0.0001
Residuos 17 11925 701.45
Total 20 21273
Tabla 10. Resultados del test Pair-Wise evaluando diferencias entre cada una de las cuatro
zonas estudiadas en términos de la presencia-ausencia del fitoplancton nocivo. Valores-p
significativos en negrita. P(perm) = valor-p permutacional, P(MC) = valor-p corregido por
Monte Carlo.
Grupos
t
P(perm)
Permutaciones
únicas
P(MC)
Canal Sur, Golfo 3.07 0.002 455 <0.001
Canal Sur, Taitao 2.08 0.098 10 0.038
Canal Sur, Moraleda 1.32 0.203 10 0.202
Golfo, Taitao 1.51 0.039 455 0.065
Golfo, Moraleda 2.25 0.002 455 0.002
Taitao, Moraleda 1.37 0.107 10 0.169
Cuando se considera, tanto la composición de especies como su abundancia, el MDS da un
resultado levemente diferente al anterior, observándose Taitao con un poco menos similitud
con el Golfo de Penas (Fig. 27). En este caso, el análisis PERMANOVA también encuentra
diferencias significativas entre zonas (Tabla 11), y los test Pair-Wise no encuentran
diferencias significativas solamente entre el canal Sur y canal Moraleda, y entre Taitao y el
Canal Moraleda (Tabla 12).

63
Figura 27. Análisis MDS en base a matriz de similitud de Bray-Curtis, a partir de datos de
abundancia de fitoplancton nocivo, estandarizada y luego transformada a raíz cuarta. Las
estaciones se muestran agrupadas en las 4 zonas geográficas.
Tabla 11. Resultados del análisis PERMANOVA, en base a matriz de similitud de Bray-
Curtis, a partir de datos de abundancia de fitoplancton nocivo, estandarizada y luego
transformada a raíz cuarta. Factor Zona se refiere a las zonas geográficas definidas y
registradas durante el crucero. Valores-p significativos en negrita. P(perm) = valor-p
permutacional, P(MC) = valor-p corregido por Monte Carlo.
Fuente de
Variación
g.l.
SC
CM
Pseudo-F
P(perm)
Permutaciones
únicas
P(MC)
Zona 3 9385.1 3128.4 7.71 0.0001 9912 0.0001
Residuos 17 6902 406
Total 20 16287
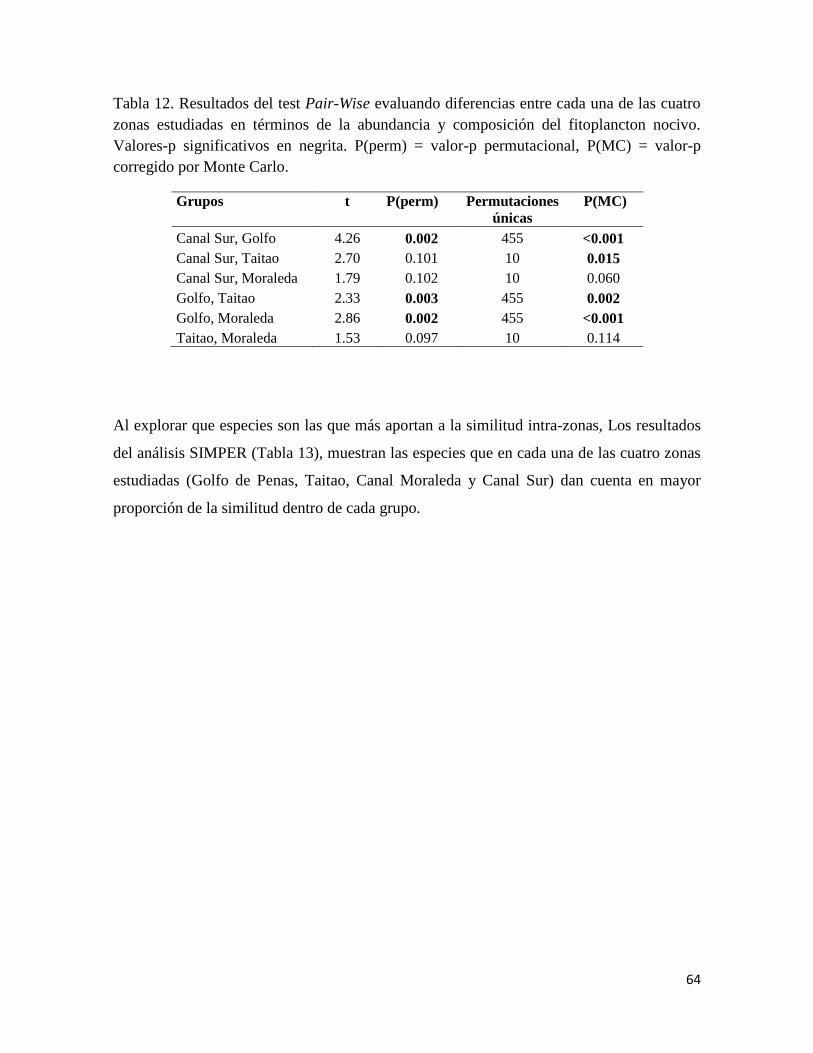
64
Tabla 12. Resultados del test Pair-Wise evaluando diferencias entre cada una de las cuatro
zonas estudiadas en términos de la abundancia y composición del fitoplancton nocivo.
Valores-p significativos en negrita. P(perm) = valor-p permutacional, P(MC) = valor-p
corregido por Monte Carlo.
Grupos
t
P(perm)
Permutaciones
únicas
P(MC)
Canal Sur, Golfo 4.26 0.002 455 <0.001
Canal Sur, Taitao 2.70 0.101 10 0.015
Canal Sur, Moraleda 1.79 0.102 10 0.060
Golfo, Taitao 2.33 0.003 455 0.002
Golfo, Moraleda 2.86 0.002 455 <0.001
Taitao, Moraleda 1.53 0.097 10 0.114
Al explorar que especies son las que más aportan a la similitud intra-zonas, Los resultados
del análisis SIMPER (Tabla 13), muestran las especies que en cada una de las cuatro zonas
estudiadas (Golfo de Penas, Taitao, Canal Moraleda y Canal Sur) dan cuenta en mayor
proporción de la similitud dentro de cada grupo.

65
Tabla 13. Resultados del análisis SIMPER, para cada zona estudiada. Se entregan en orden
decreciente las especies que más aportan a la similitud intra zona. a) Canales Sur, b) Golfo
de Penas, c) Península de Taitao, y d) Canal Moraleda.
a) Canales Sur (Messier y Wide)
Similitud Promedio: 77.07%
Especies Abund.Prom. Sim.Prom. Sim/DE Contrib% Acum.%
Skeletonema spp. 2,97 15,96 7,48 20,7 20,7
Pseudo-nitzschia pseudodelicatissima 1,6 7,65 14,37 9,93 30,63
Cerataulina pelágica 1,37 6,97 11,32 9,05 39,68
Pseudo-nitzschia delicatissima 1,23 6,42 10,7 8,32 48,01
Pseudo-nitzschia pungens 1,01 5,46 10,98 7,08 55.09
b) Golfo de Penas
Similitud Promedio: 76.18%
Especies Abund.Prom. Sim.Prom. Sim/DE Contrib% Acum.%
Gymnodinium sp. 2,2 7,73 11,11 10,15 10,15
Azadinium sp. 2,05 6,8 6,76 8,93 19,08
Gymnodinium aureolum 1,57 5,47 18,17 7,18 26,26
Gymnodinium simplex 1,51 5,1 11,36 6,7 32,95
Ceratium furca 1,49 4,77 4,58 6,26 39,22
c) Península de Taitao
Similitud Promedio: 73.18%
Especies Abund.Prom. Sim.Prom. Sim/DE Contrib% Acum.%
Leptocylindrus mediterraneus 2,16 6,28 3,17 8,59 8,59
Pseudo-nitzschia fraudulenta 1,53 5,42 11,26 7,41 16
Leptocylindrus danicus 2,01 5,29 2,02 7,23 23,23
Pseudo-nitzschia pungens 1,57 5,26 18,33 7,19 30,42
Pseudo-nitzschia pseudodelicatissima 1,43 4,61 6,1 6,3 36,72
d) Moraleda
Similitud Promedio: 51.97%
Especies Abund.Prom. Sim.Prom. Sim/DE Contrib% Acum.%
Alexadrium ostenfeldii 2,61 11,21 3,48 21,56 21,56
Skeletonema spp. 1,62 6,63 8,41 12,77 34,33
Pseudo-nitzschia pseudodelicatissima 1,23 5,51 4,02 10,6 44,92
Pseudo-nitzschia pungens 1,33 5,24 5,86 10,08 55,01
Ceratium tripos 0,99 3,88 7,14 7,46 62,47
En el Canal Sur dominan las diatomeas. Skeletonema spp. por si solas dan cuenta del 20% de
la similitud, siendo seguida por Pseudo-nitzschia pseudodelicatissima con un 9.9%, y por
Cerataulina pelagica con un 9% (Tabla 13a). En el caso del Golfo de Penas, las especies que
más aportan a la similitud intra-zona son los dinoflagelados Gymnodinium spp. (10%),

66
Azadinium spp. (9%) y Gymnodinium aureolum (7%) (Tabla 13b). En la zona de Taitao
también dominaron las diatomeas, siendo Leptocylindrus mediterraneus (8.6%) la más
importante, seguida de Pseudo-nitzschia fraudulenta, Leptocylindrus danicus y Pseudo-
nitzschia pungens, aportando con un 7% cada una (Tabla 13c). Finalmente, las especies más
importantes en el canal Moraleda fueron diatomeas y dinofagelados: Alexadrium ostenfeldii
(22%), Skeletonema spp. (13%), Pseudo-nitzschia pseudodelicatissima (10%) y P. pungens
(10%) (Tabla 13d).
Los análisis de NMDS exploratorios de matrices ambientales y biológicas, identificaron tres
grandes zonas: (a) Golfo de Penas y Península de Taitao, (b) Canal Moraleda, y (c) Canales
Sur. Un análisis PERMANOVA basado en estos grupos fue altamente significativo (p
<0,001) para la abundancia y la profundidad de fitoplancton potencialmente nocivo (pseudo-
F = 10,55 y 6,2 respectivamente), así como para los datos ambientales (pseudo-F = 14,6 y
3,2). La dispersión dentro del grupo en todos los casos no fue significativa (PERMDISP p>
0,1). El análisis de CAP-CDA con las tres zonas NMDS se representaron, por razones de
simplicidad, en gráficos de dos dimensiones. Para la matriz ambiental, la diferencia entre
grupos fue altamente significativa (p <0,001) y el primer eje canónico se separó claramente
en tres zonas, las cuales coinciden con el NMDS inicial (correlación canónica d1 = 0,91). La
clasificación total de CAP para cada estación muestreada en las diferentes zonas fue alta
(95%). Las variables que más contribuyeron a la separación en la ordenación a lo largo del
primer eje fueron la salinidad (r = 0,7), temperatura (r = 0,9), NO3 (r = 0,6) y Si(OH)4 (r=0,7),
mientras que en el segundo eje fueron dirección del viento (r = 0,5) y PO4 (r = 0,7) (Fig.
28A). Las variables ambientales que caracterizaron la zona del Golfo de Penas y Península
de Taitao fueron salinidad y temperatura, dirección del viento y menor concentración de
nutrientes (NO3, (SiOH)4 y PO4), en comparación con los Canales Moraleda y Canales Sur.
El Canal Moraleda se caracterizó por presentar una mayor temperatura, salinidad, PO4 y
menores relaciones de N/P que los Canales Messier y Wide (Tabla 6).
Para los datos de abundancia de fitoplancton potencialmente nocivo, el análisis CAP-CDA
indicó que las diferencias taxonomicas entre las tres zonas (i.e. Golfo de Penas y Península
de Taitao, Canal Moraleda, y Canales Sur) eran muy altas (d1 = 0,94). La clasificación total
de CAP para las estaciones muestreadas en cada zona fue alta (95%). Los taxones que más

67
contribuyeron a la separación en la ordenación a lo largo del primer eje fueron Azadinium
spp., Gymnodinium aureolum, Karenia mikimotoi, Gymnodinium spp., representativos del
Golfo de Penas y Península de Taitao (r ≥ 0,42 en los 5 casos) y en el segundo eje Pseudo-
nitzschia pseudodelicatissima, mientras que Alexandrium ostenfeldii (r= 0,43), a lo largo del
primer eje, fue representativo del Canal Moraleda (Fig. 28B). Por otra parte las especies de
Skeletonema spp., Cerataulina pelagica y Dinophysis acuminata, ubicada a lo largo del
segundo eje, fueron representativos de los Canales sur (r ≥ 0,53 en los tres casos).
La abundancia promedio de fitoplancton nocivo (5 a 30 m de profundidad) fue un orden de
magnitud más bajo en la zona del Canal Moraleda en comparación con las zonas de Canales
sur y el Golfo de Penas y Taitao. Un patrón diferente se observó en la fracción de clorofila
de microplancton (Chla_micro), donde las máximas concentraciones se encontraron en los
Canales Moraleda y Sur. Las concentraciones más bajas de Chla_micro en la zona del Golfo
de Penas y Taitao son atribuibles a una menor contribución de diatomeas y una mayor
concentración de dinoflagelados en la zona, como lo reflejan los análisis CAP. Los géneros
de diatomeas que más contribuyeron a la abundancia de fitoplancton nocivo fueron
Skeletonema, Pseudo-nitzschia y Leptocylindrus. Mientras que los géneros de dinoflagelados
que más contribuyeron a la abundancia de fitoplancton nocivo fueron Azadinium y
Gymnodinium como se refleja en el análisis CAP-CDA (Figura 28B).
El análisis CAP-CCorA combinando la abundancia de fitoplancton potencialmente nocivo
con las variables ambientales (Fig. 28C), indicó la existencia de fuertes correlaciones y
significativas entre ambos conjuntos (p <0,001; d1 = 0,65 y d2 = 0,5). El desplazamiento
espacial en la estructura del fitoplancton potencialmente nocivo estuvo fuertemente asociado
(autovector canónico) con salinidad y NO3 (r= 0,5) y la relación Si/N (r = 0,5) en el segundo
eje, principalmente dado por Skeletonema spp., Azadinium spp., Pseudo-nitzschia
pseudodelicatissima, P. delicatissima, Cerataulina pelagica (Fig.28D).
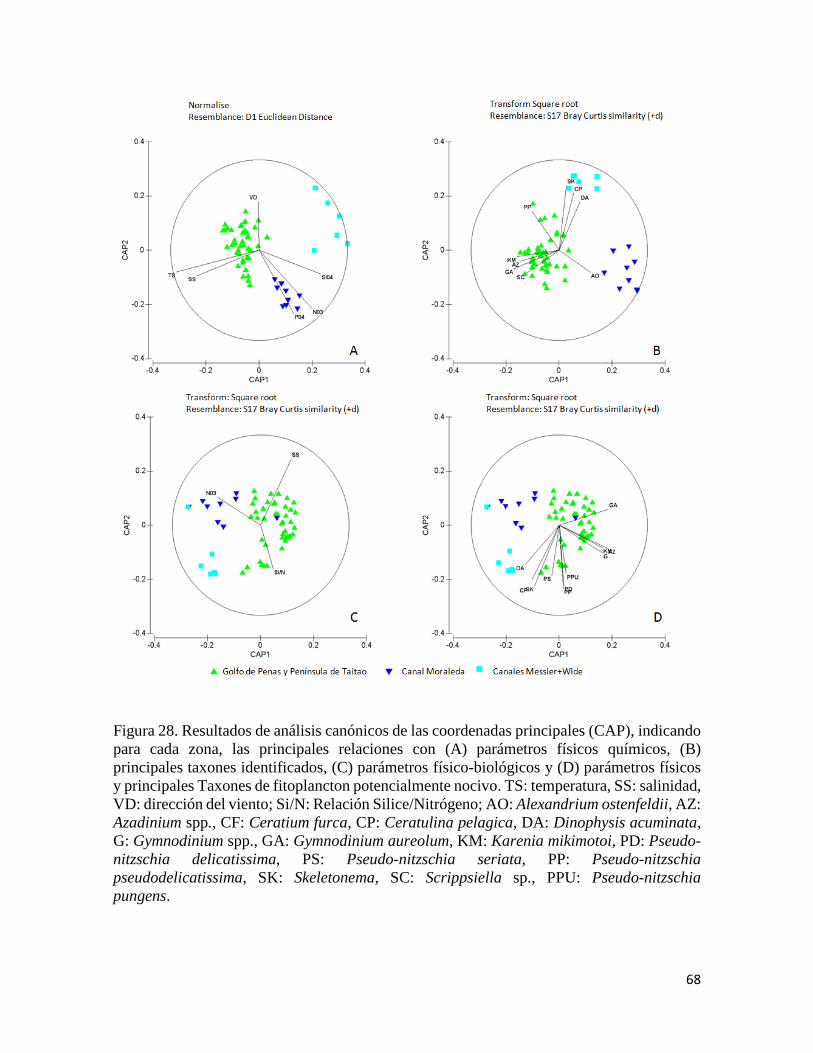
68
Figura 28. Resultados de análisis canónicos de las coordenadas principales (CAP), indicando
para cada zona, las principales relaciones con (A) parámetros físicos químicos, (B)
principales taxones identificados, (C) parámetros físico-biológicos y (D) parámetros físicos
y principales Taxones de fitoplancton potencialmente nocivo. TS: temperatura, SS: salinidad,
VD: dirección del viento; Si/N: Relación Silice/Nitrógeno; AO: Alexandrium ostenfeldii, AZ:
Azadinium spp., CF: Ceratium furca, CP: Ceratulina pelagica, DA: Dinophysis acuminata,
G: Gymnodinium spp., GA: Gymnodinium aureolum, KM: Karenia mikimotoi, PD: Pseudo-
nitzschia delicatissima, PS: Pseudo-nitzschia seriata, PP: Pseudo-nitzschia
pseudodelicatissima, SK: Skeletonema, SC: Scrippsiella sp., PPU: Pseudo-nitzschia
pungens.

69
Debido a la alta abundancia de dinoflagelados en las estaciones del Golfo de Penas y
Península de Taitao, se realizaron análisis estadísticos con la data de estas estaciones por
separado. Los análisis de NMDS exploratorios de matrices ambientales y biológicas,
identificaron diferencias entre las estaciones del Golfo de Penas y las estaciones de la
Península de Taitao. Un análisis PERMANOVA basado en estos grupos fue significativo (p
≤ 0,031) para la abundancia y datos ambientales (pseudo-F = 2,55; pseudo-F = 6,88
respectivamente), dada principalmente por las estaciones 18 y 20 de Taitao. La dispersión
dentro del grupo en todos los casos no fue significativa (PERMDISP p > 0,5). El análisis de
CAP-CDA con las estaciones NMDS se representaron, por razones de simplicidad, en
gráficos de dos dimensiones (Fig. 29). Para la matriz ambiental, la diferencia entre grupos
fue altamente significativa (p <0,0001) y el primer eje canónico separó principalmente las
estaciones 18 y 20 de la Península de Taitao del resto de las estaciones, las cuales coinciden
con el NMDS inicial (correlación canónica d1 = 0,99; Fig. 29B). La clasificación total de
CAP para cada estación muestreada fue media (64%), pero alta entre zonas (90%). Las
variables que más contribuyeron a la separación en la ordenación a lo largo del primer eje
fueron la dirección del viento (r = 0,7) y la extinción de la luz (r = 0,5), mientras que en el
segundo eje fue la magnitud del viento (r = 0,9) (Fig. 29B).
Para los datos de abundancia de fitoplancton potencialmente nocivo, el análisis CAP-CDA
indicó que hay diferencias taxonómicas significativas (p < 0,0002) entre el Golfo de Penas y
Península de Taitao (d1 = 0,7). La clasificación total de CAP para las estaciones muestreadas
fue baja por estaciones de muestreo (7%), pero alta entre zonas (89 %). Los taxones más
representativos en la ordenación a lo largo del primer eje fueron Azadinium, Karenia
mikimotoi, Karenia bicuneiformis, Pseudo-nitzschia subpacifica, Pseudo-nitzschia
pseudodelicatissima y Leptocylindrus danicus, representativos del Golfo de Penas y
Península de Taitao (r ≥ 0,5) y en el segundo eje Skeletonema spp. y Cerataulina pelágica (r
≥ 0,5 en ambos casos), (Fig. 29A).
El análisis CAP-CCorA combinando la abundancia de fitoplancton potencialmente nocivo
con las variables ambientales (Fig. D), indicó la existencia de correlaciones fuertes entre
ambos conjuntos (d1 = 0,61 y d2 = 0,53), los cuales no fueron significativas (0,09). El
desplazamiento espacial en la estructura del fitoplancton potencialmente nocivo estuvo

70
fuertemente asociado (autovector canónico) a las variables que más contribuyeron a la
separación en la ordenación a lo largo del primer eje, las cuales fueron: zona de compensación
(zc) y la proporción Si/N (r ≥ 0,5 en ambos casos), mientras que en el segundo eje las
variables que estuvieron más asociadas al desplazamiento espacial fueron la temperatura (TS)
y el pH (r ≥ 0,5 en ambos casos). El desplazamiento espacial en la estructura del fitoplancton
potencialmente nocivo estuvo fuertemente asociado (autovector canónico) con la relación
Si/N, temperatura, extinción de la luz y pH (r = 0,5). Los taxones que más contribuyeron a la
separación en la ordenación a lo largo del primer eje fueron Azadinium spp., Gymnodinium
spp., Pseudo-nitzschia fraudulenta, Karenia mikimotoi y Karenia bicuneiformis (r ≥ 0,60),
mientras que en el segundo eje Procentrum micans, Leptocylindrus danicus, Pseudo-
nitzschia pungens, Karenia papilonacea, Alexandrium ostenfeldii, Gymnodinium aureolum,
Leptocylindrus mediterraneus y Ceratium Furca (r ≥ 0,60) (Fig. 29C).
Los análisis de PERMANOVA en términos de abundancia para las especies o géneros
Azadinium, Karenia y Gymnodinium entre las estaciones del Golfo de Penas y las estaciones
de la Península de Taitao, mostró bajas diferencias entre ambas zonas (p < 0,034). La
dispersión dentro del grupo en todos los casos no fue significativa (PERMDISP p > 0,2). Para
los datos de abundancia de fitoplancton potencialmente nocivo, el análisis CAP-CDA indicó
que las diferencias significativas entre zonas (p < 0,001) taxonómicas entre el Golfo de Penas
y Península de Taitao eran medias (d1 = 0,4). La clasificación total de CAP para las
estaciones muestreadas fue baja por estaciones de muestreo (7%), pero alta entre zonas (93
%). Los taxones más representativos en la ordenación a lo largo del primer eje fueron
Azadinium, Karenia mikimotoi, Gymnodinium spp. y Karenia brevisulcata representativos
del Golfo de Penas y Península de Taitao (r ≥ 0,5) y en el segundo eje Karenia bicuneiformis
y Gymnodinium simplex (r ≥ 0,5), (Fig. 29 E). Los análisis CAP-CDA y CAP-CCorA para
las variables ambientales siguen el mismo patrón anteriormente descrito.

71
Figura 29. Análisis canónicos de coordenadas principales (CAP), indicando para el Golfo de Penas y
Península de Taitao, las principales relaciones con (A) parámetros físicos químicos, (B) principales

72
taxones identificados, (C) parámetros físico-biológicos y (D) parámetros físicos y principales Taxones
de fitoplancton potencialmente nocivo. TS: temperatura, SS: salinidad, VD: dirección del viento;
Si/N: relación Silicio/Nitrógeno; ZC: zona de compensación; AO: A. ostenfeldii, AZ: Azadinium spp.,
CFR: C. furca, CP: C. pelagica, G: Gymnodinium spp., GA: G. aureolum, KM: K. mikimotoi, KP: K.
papilonacea, KBI K. bicuneiformis, PSP: P. subpacifica, SK: Skeletonema spp., SC: Scrippsiella spp.,
PPU: P. pungens; LD: L. danicus, SK: Skeletonema spp., PP: P. pseudodelicatissima.
Finalmente, para mayor claridad se realizó un análisis por profundidad en las estaciones del
Golfo de Penas y Península de Taitao. Los análisis de NMDS exploratorios de matrices
ambientales no muestran diferencias significativas entre las 3 profundidades de muestreo, a
diferencia de la matriz biológica, que si muestra diferencias significativas entre las 3
profundidades (5, 15 y 30 m) de las diferentes estaciones del Golfo de Penas y Península de
Taitao. Un análisis PERMANOVA basado en estos 3 estratos para las 15 estaciones del Golfo
de Penas y Taitao no mostró diferencias significativas en los datos ambientales (pseudo-F =
1,04; p = 0,39), sin embargo, la abundancia fue altamente significativa (pseudo-F = 15,989;
p =0,001). La dispersión dentro del grupo en todos los casos no fue significativa (PERMDISP
p > 0,5). El análisis de CAP-CDA con las estaciones NMDS se representaron, por razones
de simplicidad, en gráficos de dos dimensiones (Fig. 30). Para la matriz ambiental, la
diferencia entre grupos no fue significativa (p <0,1) entre los diferentes estratos para las
diferentes estaciones, las cuales coinciden con el NMDS inicial (correlación canónica d1 =
0,7). La clasificación total de CAP para cada estación muestreada fue media (50%). Las
variables que más contribuyeron a la separación en la ordenación a lo largo del primer eje
fueron la salinidad y la proporción Si/N (r = 0,5), mientras que en el segundo eje fue la
temperatura y la salinidad (r = 0,5) (Fig. 30B).
Para los datos de abundancia de fitoplancton potencialmente nocivo, el análisis CAP-CDA
indicó que existen diferencias taxonómicas muy altas entre los tres estratos (p<0,0001) las
cuales coinciden con el NMDS inicial (correlación canónica d1 = 0,98) La clasificación total
de CAP para las estaciones muestreadas fue alta (100 %). Los taxones más representativos
en la ordenación a lo largo del primer eje fueron Azadinium spp., Leptocylindrus danicus,
Leptocylindrus mediterraneus, Karenia mikimotoi, Karenia papilonacea, Gymnodinium
spp., Ceratium Furca, Alexadrium ostenfeldii, Pseudo-nitzschia seriata, Pseudo-nitzschia

73
fraudulenta, Prorocentrum micans y Gonyaulax polygramma (r ≥ 0,5 en todos los casos)
representativos del Golfo de Penas y Península de Taitao y en el segundo eje Gymnodinium
aureolum, Gymnodinium simplex, Karenia bicuneiformis (r ≥ 0,5 en los tres casos).
El análisis CAP-CCorA combinando la abundancia de fitoplancton potencialmente nocivo
con las variables ambientales (Fig.28C), indicó la existencia de correlaciones fuertes entre
ambos conjuntos (d1 = 0,71 y d2 = 0,59). El desplazamiento espacial en la estructura del
fitoplancton potencialmente nocivo estuvo fuertemente asociado (autovector canónico) a la
variable que más contribuyó a la separación en la ordenación a lo largo del primer eje fue la
Salinidad (r> 0,5), mientras que el segundo eje la variable que estuvo más asociada al
desplazamiento espacial fue la temperatura (r > 0,5). Los taxones que más contribuyeron a la
separación en la ordenación a lo largo del primer eje fueron Ceratium Furca, Dinophysis
tripos, Gonyaulax polygramma, Gymnodinium spp., Pseudo-nitzschia pseudodelicatissima,
Karenia mikimotoi, Karenia papilonacea y Prorocentrum micans (r ≥ 0,50 en todos los
casos), mientras que en el segundo eje Azadinium spp., Pseudo-nitzschia subpacifica,
Leptocylindrus danicus, Karenia bicuneiformis (r ≥ 0,50) (Fig. 30C).
El análisis de PERMANOVA, en términos de abundancia para las especies o géneros
Azadinium, Karenia y Gymnodinium basado en estos 3 estratos para las 15 estaciones del
Golfo de Penas y Taitao no mostró diferencias significativas en los datos ambientales
(pseudo-F = 1,04; p = 0,39), a diferencia de la abundancia quefue altamente significativa
(pseudo-F = 27,9; p =0,0001). La dispersión dentro del grupo en todos los casos no fue
significativa (PERMDISP p > 0,4). Para los datos de abundancia de especies o géneros
Azadinium, Karenia y Gymnodinium, el análisis CAP-CDA indicó que existen diferencias
taxonómicas muy altas entre los tres estratos (p < 0,0001) las cuales coinciden con el NMDS
inicial (correlación canónica d1 = 0,98). La clasificación total de CAP para las estaciones
muestreadas fue alta (100 %). Los taxones más representativos en la ordenación a lo largo
del primer eje fueron Azadinium spp., Gymnodinium aureolum, Gymnodinium spp., y karenia
papilonacea (r ≥ 0,60 en los 4 casos), mientras que en el segundo eje fueron Karenia
bicuneiformis, Gymnodinium simplex (r ≥ 0,60 en ambos casos) (Fig. E). El análisis CAP-
CDA para las variables ambientales sigue el mismo patrón descrito anteriormente al igual el
análisis CAP-CCorA anteriormente descrito.

74
Figura 30. Relación entre análisis canónicos de las coordenadas principales (CAP), del Golfo
de Penas y Península de Taitao, y las principales relaciones con (A) parámetros físicos químicos,
(B) principales taxones identificados, (C) parámetros físico-biológicos y (D) parámetros físicos
y principales taxa de fitoplancton potencialmente nocivo. TS: temperatura, SS: salinidad, VD:

75
dirección del viento; Si/N: Relación Silicio/Nitrógeno; ZC: Zona de compensación; AO:
A.ostenfeldii, AZ: Azadinium spp., CFR: C. furca, CP: C. pelagica, G: Gymnodinium spp., GA:
G. aureolum, KM: K.mikimotoi, Kp: K.papilonacea, KBI K.bicuneiformis, PSP: Pseudo-
nitzschia subpacifica, SK: Skeletonema, SC: Scrippsiella sp., PPU: Pseudo-nitzschia pungens;
LD: L. danicus, SK: Skeletonema spp., PP: P. pseudodelicatissima.

76
V DISCUSIÓN GENERAL
Las floraciones de algas nocivas están ocurriendo cada vez con mayor frecuencia,
comprometiendo áreas costeras mucho más extensas del planeta (Anderson 1989, Smayda
1990, Hallegraeff 1993, Wells et al. 2015). En la actualidad, cerca de 100 especies de
microalgas son consideradas nocivas debido a los efectos perjudiciales de grandes
floraciones en el ecosistema marino y por consecuencia, en humanos (Sanseverino et al.
2016). Los efectos nocivos pueden incluir toxicidad (biotoxinas), ictiotoxicidad (ej. vía
ácidos grasos poli-insaturados, especies reactivas de oxígeno y gimnocina), daño físico,
anoxia y reducción drástica de la irradiación (Barnes et al. 2015).
Entre las especies nocivas descritas, se enumeran muchos miembros de la clase Dinophyceae,
incluyendo la familia Gymnodiniaceae con los géneros Gymnodinium y Karenia que han sido
responsables de la muerte de peces y crustáceos en todo el mundo (Evans 1975, Mahoney et
al. 1990, Dahl y Tangen 1993, Uribe y Ruiz 2001, Faust y Gulledge 2002 y Davidson et al.
2009). Nuestros resultados confirman la presencia de ambos géneros en la zona de estudio,
principalmente en la zona del Golfo de Penas y Península de Taitao, siendo las especies
Karenia mikimotoi, Gymnodinium aureolum, Azadinium spp. y Scrippsiella spp. quienes
contribuyen a marcar las diferencias entre la zona del Golfo de Penas - Taitao, y los Canales
Moraleda y Sur, mientras que en términos de diatomeas el género Pseudo-Niztschia fue el
que contribuyó de mayor manera a la separación entre zonas y fue característica de la zona
del Golfo de Penas y Taitao. Estos resultados se explicarían por la diferencia observada en
los parámetros ambientales entre estas zonas, especialmente en términos de salinidad,
temperatura, concentración de nutrientes y proporción de N/P.
La ausencia de conocimiento fundamental acerca de los mecanismos que conducen a las
proliferaciones de algas nocivas complica los pronósticos de su prevalencia futura,
especialmente considerando un escenario de cambio climático global, donde cambios en la
temperatura, estratificación, luz, acidificación de los océanos, ingreso de nutrientes, y el
pastoreo, afectarán directamente la variabilidad de las floraciones de algas nocivas (Wells et
al. 2015).

77
La composición y abundancia del fitoplancton nocivo (como indican los análisis estadísticos)
se correlacionan principalmente con las variables temperatura, salinidad, nitrato y ácido
silícico (Figura 28A). Los parámetros físicos como la temperatura y la salinidad pueden
regular los patrones de distribución regional de una especie y contribuir a la aparición de los
máximos estacionales, aunque generalmente se piensa que la disponibilidad de luz y/o los
nutrientes determinan el crecimiento, la biomasa y la duración de las floraciones en escala
local (Barnes et al. 2010).
La distribución de nutrientes en la zona (Golfo de Penas y Canales Australes) sugiere que las
fuentes de nitrato y fosfato serían principalmente de origen marino, ya que el agua dulce
desde ríos, lluvia y derretimiento glaciar en la zona de estudios son pobres en estos nutrientes,
pero contribuyen significativamente en ácido silícico a la capa superficial (Silva 2008). Silva
y Palma (2008) describen varias relaciones de nitrato-fosfato en la columna de agua,
planteando que bajas concentraciones superficiales pueden deberse al consumo por
fitoplancton. Esto se ve reforzado por el bajo ingreso de nitrógeno y fosforo mediante agua
dulce, situación que se ve bien representada en el Golfo de Penas y Península de Taitao, con
alta abundancia de dinoflagelados, pero bajas concentraciones de clorofila (total y fracción
> 20 m), a diferencia de lo reportado por Silva y Palma (2008) para dicha zona. Los bajos
niveles de clorofila están asociados al hecho que el componte principal del microfitoplancton
en esta zona corresponde a dinoflagelados, muchos de los cuales presentan un
comportamiento trófico de mixotrofia y heterotrofia (Anabalón et al. 2014).
En el presente estudio se identificaron estaciones dentro del Golfo de Penas y Península de
Taitao (SNP4, SNP10, SNP7, SNP6) con proporciones de N:P <6, lo que sugiere una
potencial limitación por N, el cual es un nutriente fundamental para el crecimiento del
fitoplancton. Sin embargo, Gymnodinium, Azadinium y Karenia presentan ventajas
fisiológicas, debido a su capacidad de usar diversas fuentes de nitrógeno para su crecimiento
(Yamaguchi y Itakura, 1999, Jauffrais et al., 2013, Li et al., 2016). Por ejemplo, K. mikimotoi
puede crecer utilizando nitrato, nitrito o amonio como fuente de nitrógeno (Yamaguchi e
Itakura, 1999). Además se detectaron estaciones (SNP4, SNP9, E14, E16, SNP6, E20) con
proporciones muy bajas de N:P <6, bordeando el límite de ser consideradas potencialmente
deficientes en N (excluyendo NH4), lo cual podría parcialmente explicar la prevalencia de

78
conjuntos de grupos taxonómicos mixtos, autotróficos / mixotróficos encontrados en la zona
del Golfo de Penas y Taitao.
Una floración de fitoplancton nocivo, puede influir fuertemente en la disponibilidad de los
nutrientes en la columna de agua, considerando además que cada taxón posee requerimientos
de nutrientes específicos para su desarrollo (Quigg et al. 2003). Por ejemplo, varios géneros
de diatomeas céntricas (e.g. Thalassiosira, Cocinodiscus), son sensibles a bajas
concentraciones de N, pero son capaces de almacenarlo en vacuolas; sin embargo la
abundancia de células disminuye bajo esta condición (Van Ruth et al. 2012). Esto podría
explicar, a modo de hipótesis, la menor presencia de diatomeas céntricas en la zona del Golfo
de Penas y Península de Taitao.
Las zonas de Canales Sur (Messier y Wide), muestran alta estratificación, debido a la
presencia de una capa superficial de agua dulce, además de presentar mayor contenido de
nutrientes en relación al resto de la zona muestreada, lo que podría favorecer la aparición de
diatomeas céntricas. Altas concentraciones de clorofila-a han sido descritas en el rango de las
observadas en este muestreo en el área. Estudios en la zona atribuyen el crecimiento de
fitoplancton a la fertilización de la cuenca a través de aguas oceánicas y la formación del
gradiente térmico en la capa superficial, lo que permite la permanencia del fitoplancton en la
zona fótica (Ramírez y Pizarro, 2005)
En la zona superficial de los canales Moraleda Messier y Wide, especialmente en estos dos
últimos, se aprecian valores de pH menores a los reportados en el resto de la zona muestreada.
Silva (2008) explica que situaciones particulares, donde el pH y el oxígeno difieren de esta
forma, se atribuyen al ingreso de agua dulce más ácida y oxigenada, como sucede en las
zonas de descarga del río Baker.
El subconjunto de especies de fitoplancton que proliferan para cualquier evento FAN es a
menudo al azar, pero típicamente se potencia algún tipo particular de organismo dependiendo
de sus adaptaciones a la turbulencia, concentración de nutrientes, estratificación, adaptación
a la luz reducida, etc. (Smayda y Reynolds 2001, Estrada et al. 2003). Especies del género
Skeletonema presentan altas tasas de crecimiento asociadas a mayor disponibilidad de
nutrientes (Sommer 1984, Collos 1986, Yamamoto y Hatta 2004). Fawcett et al. (2007),

79
informaron una alternancia de eventos FAN de diatomeas (Pseudo-nitzschia) y
dinoflagelados (P. reticulatum, Dinophysis) en el sur de Benguela, impulsados por cambios
en la intensidad y estratificación de la surgencia.
Estudios sobre distribución cualitativa y cuantitativa del microplancton marino realizados
entre Puerto Montt y la Laguna San Rafael, indican que las áreas con mayores
concentraciones de microfitoplancton se sitúan por el norte en Seno Reloncaví y Golfo de
Ancud, mientras que por el sur se encuentran en los Canales Moraleda, Jacaf y Puyuguapi,
con densidades entre 1 a 5 millones de cél. L-1 (Avaria et al.1997).
El conjunto de especies nocivas (diatomeas y dinoflagelados) detalladas en el presente
estudio, mostró abundancias altamente variables. Las mayores abundancias de
dinoflagelados corresponden a especies de los géneros Gymnodinium, Azadinium., Karenia
(principalmente K. mikimotoi), siendo esta última la que enciende la alarma acerca de la
producción de FAN en la zona sur de nuestro país en febrero de 2017. Sin embargo, las
abundancias reportadas para K. mikomitoi en este estudio, por sí solas no podrían causar
efectos tóxicos agudos. Considerando las mortalidades de peces reportadas a inicios de
febrero en la zona de estudio, es altamente probable que el crucero de prospección se haya
realizado en una etapa post-bloom, y las abundancias registradas correspondan a su etapa de
decaimiento. Es importante considerar que las FAN se encuentran típicamente asociadas a
escalas de variabilidad temporales de días a semanas (incluso meses) y escalas espaciales de
cientos de metros a decenas de kilómetros (Gentien et al. 2005, Durham y Stocker 2012). En
las aguas costeras de América del Norte, diferentes especies de dinoflagelados desarrollan
con frecuencia floraciones que duran varios meses, tales como las observadas en el este del
Golfo de México, el Golfo de Maine y a lo largo de la costa del Pacífico (Horner et al. 1997,
Anderson et al. 2005). Estas floraciones se extienden por distancias de decenas a cientos de
kilómetros en la superficie del océano, mientras que otras especies como Karenia mikimotoi
y Dinophysis acuminata, pueden existir como “capas” en la columna de agua, con un grosor
que oscila entre decenas de centímetros a metros (Gentien et al. 2005).
Por otra parte, es necesario destacar la presencia de varias especies algales nocivas
(potencialmente productoras de toxinas) en la zona de estudio durante el crucero

80
oceanográfico. Si bien, la exposición de la biota a mezclas de diferentes grupos de biotoxinas,
es un tema que ha sido escasamente estudiado, se ha reportado la co-ocurrencia de biotoxinas
en mariscos que fueron contaminados por diferentes especies de microalgas
simultáneamente, por lo que la co-ocurrencia de varias especies productoras de toxinas debe
ser considerada en los estudios de los efectos de las FAN (Hess 2002, 2010, Amzil et al.
2008, Twiner et al. 2008, Reguera et al. 2012, Suikkanen et al. 2013). El estudio de la
dinámica poblacional de las floraciones nocivas es imperativo a la hora de definir la toxicidad
(Zingone y Enevoldsen, 2000), ya que la sinergia entre compuestos generadas por diversas
especies podría incrementar la toxicidad (Mayali y Doucette 2002, Haywood et al. 2004).
Especies del género Gymnodinium son altamente abundantes y fácilmente confundibles con
Karenia. Gymnodinium es un género altamente complejo, el cual se compone de más de 200
especies con alta variabilidad morfológica, lo que hace muy difícil su identificación a nivel
de especie (Taylor et al. 2003). Gymnodinium spp. pueden presentar veneno paralítico de
mariscos (VPM) y producto de la acumulación de sus saxitoxinas en mariscos, estos pueden
resultar tóxicos para consumo humano (Van Dolah 2000). Existen varios reportes que
destacan la presencia de mareas café de Gymnodinium spp. en la zona de Chiloé y fiordos
Magallánicos (Lembeye et al. 1993, Clément 1999, Guzmán et al. 1999, Uribe et al. 2001,
Uribe y Ruiz 2001). Estas investigaciones informan concentraciones moderadas a altas de
Gymnodinium (15.600–43.000 cél. L-1), con duraciones de hasta tres semanas en los fiordos,
causando la muerte de variados invertebrados (erizos, estrellas de mar, lapas, caracoles y
pulpos) y peces (Uribe et al. 2001). En Chiloé y fiordos de Magallanes, se ha reportado la
muerte de salmónidos en piscifactorías, abalones y fauna nativa, asociada a la presencia de
dinoflagelados del género Gymnodinium. Uribe y Ruiz (2001) reportan mortalidades de
invertebrados y peces asociados a abundancias similares a las encontradas en el presente
estudio de dos morfos no-identificados que se asemejarían a Gymnodinium mikimotoi
(Karenia mikimotoi).
Especies del género Azadinium han sido altamente abundantes en el ensamble de
dinoflagelados observados en este estudio. Estos organismos producen veneno azaspiracido
(AZA) causante de síndromes diarreicos en humanos, por consumo de moluscos
contaminados. Esta biotoxina provoca síntomas similares a los mostrados por Intoxicación

81
Diarreica por Mariscos (Twiner et al. 2008), la cual ha sido identificada en moluscos,
incluyendo gastrópodos, en la zona sur de Chile (Zamorano et al. 2013).
Por su parte, Karenia mikimotoi produce biotoxinas (brevitoxinas) con efectos neurotóxicos,
hemolíticos o citotóxicos en peces. Algunas especies de Karenia productoras de Veneno
Neurotóxico de moluscos (VNM) (Morris et al. 1991, Magaña et al. 2003, Kirkpatrick et al.
2004, Watkins et al. 2008, Fleming et al. 2011) han sido relacionadas directamente con la
mortandad masiva de peces, a menudo en sitios donde se realiza acuicultura en Europa
(Scatasta et al. 2003), Estados Unidos (Hoagland et al. 2002), Japón (Kim 2006), y México
(Maciel-Baltazar y Hernández-Becerril 2013), entre otros, generando importantes pérdidas
económicas (Trainer y Yoshida, 2014).
El nivel de toxicidad de cada especie es muy variable en el tiempo y depende de sus
condiciones ecológicas y fisiológicas. Algunos dinoflagelados tóxicos del género Dinophysis
tienen un alto contenido de toxina por célula, pudiendo causar intoxicaciones en humanos, a
través de la ingesta de moluscos contaminados, incluso cuando la abundancia de la especie
es inferior a 1000 cél. L-1, en comparación a otras especies que deben superar el millón de
cél. L-1 para producir el mismo efecto (Reguera et al. 2012). La intoxicación diarreica de
moluscos es causada por el ácido okadaico o sus derivados dinofisistoxinas, producidos por
10 especies del género Dinophysis (Reguera et al. 2012).
Entre las 14 especies de diatomeas con efectos potencialmente nocivos que han sido descritas
en este informe, destacan por su alta abundancia en la zona de estudio los géneros
Skeletonema spp., Leptocylindrus spp. (L. mediterraneus, L. danicus) y Pseudo-nitzschia spp
(P. pseudodelicatissima, P. delicatissima, P. pungens, P. fraudulenta, P. seriata).
Skeletonema spp. al igual que Chaetoceros spp. producen daño a las branquias de los peces,
por sus resaltadas estructuras silíceas (Freer y Vargas-Montero 2003).
Entre las especies más estudiadas de Skeletonema se encuentra S. costatum (no se identificó
a nivel de especie en este estudio), de la cual se señala que su crecimiento se ve fuertemente
influenciado por los nutrientes (Liu et al. 2002), al igual que por la luz, temperatura, salinidad
y materia en suspensión (Li et al. 2005, Sun et al. 2008). Esta especie tiene una ventaja

82
competitiva en ambientes ricos en nutrientes, en los cuales, prolifera rápidamente y presenta
un florecimiento en un período más corto (Zhu y Wang 2015).
Leptocylindrus mediterraneus y L. danicus son especies no tóxicas, que en condiciones de
floración provocan hemorragias en las branquias de peces en cultivo o irritaciones, con
producción de mucus y reducción del intercambio gaseoso (Avaria et al. 1999). Se detectó la
mayor abundancia en la zona exterior de la Península de Taitao, presentando relaciones con
la concentración de ácido silícico, temperatura y salinidad.
Alta abundancia de especies del género Pseudo-nitzchia fueron detectadas en esta
investigación, principalmente en el Golfo de Penas-Taitao. Pseudo-nitzschia causante de
Veneno Amnésico de Mariscos (VAM), es muy común en el plancton marino de Chile,
registrando floraciones de esta especie, por lo que debe considerarse como un organismo
importante en los programas de vigilancia del plancton, ya que potencialmente podría afectar
a la salud humana, la maricultura y los recursos pesqueros del país. Otros reportes, indican
que Pseudo-nitzchia multiseries, P. pseudodelicatissima y P. australis son diatomeas
productoras de ácido domoico, produciendo trastornos gastrointestinales y neurológicos,
tales como calambres, diarreas, vómitos, desorientación y pérdida de memoria (Bates et al.
1989, Avaria et al. 1999, Amzil et al. 2001). En sistemas de surgencia costera de bordes
orientales se han descrito altas abundancias de Pseudo-nitzschia asociadas a la fase de
relajación de la surgencia o con surgencia débil (Kudela et al. 2004, Díaz et al. 2014, Louw
et al. 2016). También han sido reportadas como un componente dominante en las zonas de
transición costera de Concepción (Menschel et al. 2016), así como en aguas oligotróficas del
Pacífico sur y Tropical (Gómez et al. 2007), por lo que aparentemente este género resiste
muy bien las bajas concentraciones de nutrientes (Staats et al. 2000, Anderson et al. 2010) y
explicaría en parte su mayor abundancia en la zona de Golfo de Penas y Península de Taitao.
En el norte de Chile, se ha identificado la presencia de ácido domoico en ostiones
(Argopecten purpuratus) en sitios de acuicultura como Bahía Guanaqueros, Bahía Tongoy y
Bahía Inglesa (Suárez-Isla et al. 2002; Uribe et al. 2010). Además, piures han sido también
identificados como vector para la toxina en Bahía Inglesa (López-Rivera et al. 2006). Las
concentraciones de ácido domoico en el norte de Chile estuvieron en algunos casos sobre el

83
límite reglamentario (20 mg kg-1), lo cual condujó a la prohibición de la cosecha de mariscos
de poblaciones naturales y en sitios de acuicultura (Álvares et al. 2009; Uribe et al., 2010).
La presencia de variadas especies de dinoflagelados de gran tamaño (>20 m) productoras
de toxinas, entre ellas Karenia mikimotoi, sugieren que podría haber ocurrido un efecto
nocivo combinado de especies de los géneros Gymnodinium, Azadinium, Dinophysis y
Karenia, y posiblemente con contribuciones de las especies de diatomeas Pseudo-nitzchia
spp., las cuales pueden interactuar en distintos grados sobre moluscos, peces y como
consecuencia en humanos. La alta abundancia de especies de Gymnodinium y la similitud
con especies de Karenia, permiten hipotetizar que la intoxicación de salmones ocurrida en
febrero de 2017 a bordo de las naves transportadoras (well-boats), en la zona de studio, habría
sido producto principalmente de estas especies y del efecto combinado de sus toxinas.
En el presente estudio se registraron 39 taxas (16 géneros) de fitoplancton nocivo (Fig. 24;
Tablas 2 y 4). Es importante destacar que en este muestreo se realizó la primera identificación
del género Azadinium a través de microscopía. En Chile, se han descrito varias especies de
dinoflagelados formadores de FAN, entre los cuales destacan: Alexandrium catenella
productor del Veneno Paralizante de los Mariscos (Aguilera-Belmonte et al. 2011, Varela et
al. 2012), Dinophysis acuta productor de toxinas lipofílicas diarreicas, tales como el ácido
okadaico, Dinofisitoxina-1 (García et al. 2004, 2005), D. Acuminata productor de
Pectenotoxinas (PTX-2) (Blanco et al. 2007, Fux et al. 2011) y Protoceratium reticulatum
productor de Yesotoxinas (YTXs) (Álvarez et al. 2011). Destaca también el flagelado
Heterosigma akashiwo por su efecto tóxico para la salmonicultura (Avaria et al. 1999,
Twiner et al. 2001). Además, existen reportes de varios géneros formadores de FAN, los
cuales se presentan a continuación: Dinofíceas: Alexandrium, Dinophysis, Gymnodinium,
Gambierdiscus y Prorocentrum, Rafidofíceas: Chatonella y Heterosigma, Crisofíceas:
Aureococcus, Primnesiofíceas: Chrysochromulina, Phaeocystis y Prymnesium,
Silicoflagelados: Dictyocha y Bacillarofíceas: Chaetoceros, Leptocilyndrus, Pseudo-
nitzschia y Rhizosolenia (Wright y Cembella, 1998 Avaria et al. 1999, Guzmán et al. 2002,
Suárez et al. 2002, Anderson et al. 2001, Lembeye 2008). Los trabajos de Muñoz (1985) y
Muñoz et al. (1992) describen un listado de especies de fitoplancton marino potencialmente
nocivas presentes en Chile, como por ejemplo Chaetoceros convolutus, Cerataulina

84
pelagica, Leptocylindrus minimus, Skeletonema costatum, Alexandrium ostenfeldii,
Dinophysis acuminata, Dinophysis fortii, Dinophysis rotundata, Dinophysis tripos,
Gonyaulax polyhedra, Gymnodinium catenatum, Gymnodinium splendens, Prorocentrum
micans, Prorocentrum gracile, Ceratium tripos, Ceratium furca, Scrippsiella trochoidea,
Noctiluca scintillans. Los taxa remarcados en negro en este párrafo corresponden a aquellos
también encontrados en el presente estudio.
Nuestros resultados permiten diferenciar varias zonas en base a la abundancia de especies.
Las zonas del Canal Sur (Wide + Messier) y la Península de Taitao, destacan por el dominio
de diatomeas, siendo Skeletonema spp. las más abundante en el Canal Sur y Leptocylindrus
spp. en la zona Taitao. La zona del Golfo de Penas, muestra alta presencia de dinoflagelados,
con las mayores abundancias de Gymnodinium spp. y Azadinium spp. La zona del Canal
Moraleda que presenta una mezcla de especies de diatomeas y dinoflagelados, se ve
representada por una alta abundancia Alexandrium ostenfeldii (Tabla 13). El análisis general
muestra que las diatomeas presentan asociaciones significativas con la temperatura y la
salinidad, y una estrecha relación con el fosfato (Fig. 28). Por su parte los dinoflagelados
presentan asociaciones significativas con la temperatura y los nutrientes (Fig. 28).
Los cruceros CIMAR Fiordos en las zonas de Aysen y Magallanes describen la dominancia
de especies de diatomeas r-estrategas en el microfitoplancton, los cuales muestran notables
cambios longitudinales en composición y biomasa, lo que permite diferenciar tres zonas:
fiordos y canales interiores, fiordos y canales externos, y zona oceánica (Avaria et al. 1997,
1999). El fitoplancton muestra grandes variaciones cualitativas y cuantitativas entre invierno
y primavera, en donde las mayores concentraciones celulares se registran durante la
primavera. La variedad de diatomeas y dinoflagelados, descritas en los canales externos es
mayor e incluye las especies tóxicas Alexandrium catenella, A. ostenfeldii y Dinophysis
acuta.. (Avaria et al. 1997, 1999, 2004).
La biomasa de fitoplancton depende de las variaciones estacionales en la irradiación, así
como también con otras variables oceanográficas como la temperatura, salinidad,
estratificación y concentración de nutrientes (Pizarro et al. 2000). De hecho se sabe que el
aumento de la estabilidad vertical y la estratificación de la superficie de la columna de agua,

85
debido al ingreso de agua dulce producto del derretimiento de hielo en los fiordos más
australes, juega un papel importante en el crecimiento y acumulación de fitoplancton (Pizarro
et al. 2000).
Las FAN responden a numerosos y complejos factores climáticos, hidrológicos y
oceanográficos que interactúan con procesos biológicos y que actualmente no es posible
predecir con certeza. La distribución de las FAN a gran escala están potencialmente
influenciados por procesos físicos de mesoescala, como corrientes, giros, zonas de
convergencia y surgencia (Trainer et al. 2002, Anderson et al. 2005). Además, existe escasa
o nula información cuantitativa en la zona de estudio, acerca de las entradas de nutrientes
desde ríos, escorrentía, derretimiento glaciar, y de la cantidad de materia orgánica de origen
antropogénico derivadas de centros urbanos (en expansión), actividad agrícola, ganadera, y
acuícola (ej. contribución por alimentos sin comer, fecas, excreción de peces). Numerosos
estudios, han evidenciado como el efecto de fertilizantes, aguas residuales, desechos
animales, inputs atmosféricos y la acuicultura costera contribuyen a elevar las
concentraciones de nutrientes en las aguas costeras, con fuertes evidencias de que altas
concentraciones de nutrientes han lproducido un aumento de la biomasa del fitoplancton y
de la producción primaria en algunos lugares (Trainer et al. 2002, Anderson et al. 2008,
Heysler et al. 2008, Gowen et al 2012).
A pesar del consenso general de que los cambios en la cantidad y composición del suministro
de nutrientes (crónicos y episódicos) derivados de las actividades antropogénicas afectan el
crecimiento del fitoplancton, así como su abundancia, frecuencia y toxicidad de las FAN
(Officer y Ryther 1980, Heisler et al. 2008, Anderson 2009), la evidencia científica que
sostiene este vínculo causal es escasa y depende en gran medida de las condiciones de
nutrientes a nivel local (Davidson et al. 2012). Por ejemplo, la composición del fitoplancton
no se ve afectada en lugares donde los nutrientes se encuentran en concentraciones no
limitantes para el crecimiento del fitoplancton. La evidencia de que la alteración en las
proporciones de nitrógeno: fósforo (N: P) puede promover FAN ha sido detectada solo para
localidades específicas, pero no se ha demostrado un vínculo inequívoco entre los cambios
en los índices de nutrientes y FAN (Davidson et al. 2012). Las proporciones de nutrientes se
han asociado con la cantidad y la tasa de producción de toxinas solo en condiciones de

86
laboratorio (Fehling et al. 2004, Graneli y Flynn 2006), pero es difícil realizar una
extrapolación directa de esta relación con el ambiente (Marchetti et al. 2004, Davidson et al.
2012). Por lo tanto, teniendo en cuenta el muy limitado conocimiento existente sobre los
factores que promueven el desarrollo y el decaimineto de las proliferaciones de algas nocivas
en Chile, es necesario asignar mayores esfuerzos al estudio de la relación entre los índices de
nutrientes y las FAN en la Patagonia chilena.
Por último, se evidencia la necesidad de fortalecer el sistema de monitoreo de FAN en la
zona sur-austral de Chile, de tal forma de potenciar su contribución en la mitigación de los
impactos ecológicos, económicos y sociales de estos eventos.

87
VI CONCLUSIONES
1) Se confirma la presencia de Karenia mikimotoi en la zona de estudio, aunque presenta baja
abundancia, por lo que es posible que el muestreo se haya realizado en condiciones de post-
bloom, en especial considerando que los eventos de mortalidad de peces ocurrieron
aproximadamente 15 días antes del muestreo a bordo del Buque “Marinero Fuentealba”.
2) La presencia de varias especies de dinoflagelados y diatomeas productoras de toxinas en
la zona de estudio, entre ellas Karenia mikimotoi, permite hipotetizar que la intoxicación de
peces podría deberse a un efecto sinérgico de un conjunto de toxinas.
3) Se confirma la presencia de otras especies productoras de toxinas en la zona de estudio,
principalmente Gymnodinium spp. y Azadinium spp., las cuales presentan alta abundancia.
4) El estudio presenta el primer reporte visual del dinoflagelado nocivo Azadinium spp. en
las costas de Chile, identificado directamente por microscopía óptica.
5) La distribución del fitoplancton potencialmente nocivo estuvo fuertemente asociado con
las variables ambientales salinidad, concentración de NO3 y la proporción de Si/N. Las
especies que mostraron un mayor asociación con estas variables oceanográficas fueron los
dinoflagelados Azadinium spp., Gymnodinium aureolum, Gymnodinium spp., Karenia
mikimotoi y Dinophysis acuminata y las diatomeas Skeletonema spp., Pseudo-nitzschia
pseudodelicatissima, P. delicatissima, P. seriata, P. pungens, Cerataulina pelagica.
6) Los bajos niveles de nutrientes y la relaciones N/P y N/Si, principalmente dentro del Golfo
de Penas, sugieren un drástico consumo de nutrientes después de una condición postbloom.
7) La concentración de oxígeno disuelto presente en la columna de agua de la zona de estudio
sugiere que la mortandad de peces no se habría debido a un evento de hipoxia ambiental.
8) Los análisis estadísticos de las variables biológicas y oceanográficas permitieron
identificar 3 zonas diferentes: (a) Golfo de Penas y Península de Taitao, (b) Canal Moraleda

88
y (c) Canales sur (Messier y Wide). Las variables oceanográficas que más contribuyeron a
la diferenciación en 3 zonas fueron la salinidad, temperatura y concentración de los nutrientes
NO3 y Si(OH)4. El segundo conjunto de variables que más contribuyeron a diferenciar dichas
zonas fueron la magnitud del viento, la concentración de PO4 y la proporción Si/N.
9) Análisis de CAP más detallados dentro del la zona Golfo de Penas y Península de Taitao,
muestra que la distribución de los dinoflagelados guarda relación con la profundidad, donde
la temperatura y la salinidad son los principales parámetros que afectan la distribución en
esta zona particular.
10) Los taxones que más contribuyeron a las diferentes zonas fueron Azadinium spp.,
Gymnodinium aureolum, Karenia mikimotoi, Gymnodinium spp., representativos del Golfo
de Penas y Taitao, además de Pseudo-nitzschia pseudodelicatissima; Alexandrium ostenfeldii
fue la especie más representativa del Canal Moraleda, mientras que Skeletonema spp.,
Ceratulina pelagica y Dinophysis acuminata fueron los taxones que destacan en los Canales
Sur.
11) Las condiciones oceanográficas observadas en el Golfo de Penas pueden permitir el
desarrollo de altas abundancias de especies de dinoflagelados nocivos, en consecuencia se
recomienda evitar el recambio de agua en el transporte de smolts de salmones en dicho Golfo.

89
VII AGRADECIMIENTOS
Esta investigación ha sido financiada por el Servicio Nacional de Pesca y Acuicultura
(SERNAPESCA), mediante el “Contrato de prestación de servicio especializado de
monitoreo y análisis de fitoplancton, perfil oceanográfico y nutrientes en área de emergencia
de plaga (sector Golfo de Penas, Región de Aysén) y zonas aledañas”. Se agradece, además,
el apoyo logístico recibido de las siguientes instituciones: SERNAPESCA, Instituto
Tecnológico del Salmón (INTESAL), Armada de Chile, e Instituto de Fomento pesquero
(IFOP).

90
VIII REFERENCIAS
Aguilera-Belmonte A, Inostroza I, Franco JM, Riobo P, Gómez PI. 2011. The growth,
toxicity and genetic characterization of seven strains of Alexandrium catenella (Whedon and
Kofoid) Balech 1985 (Dinophyceae) isolated during the 2009 summer outbreak in southern
Chile. Harmful Algae, 12: 105–112.
Alvarez G, Uribe E, Díaz R, Braun M, Mariño C, Blanco J. 2011. Bloom of the yessotoxin
producing dinoflagellate Protoceratium reticulatum (Dinophyceae) in northern Chile. J Sea
Res, 65: 427-434.
Alvarez G, Uribe E, Quijano-Scheggia S, López-Rivera A, Mariño C, Blanco J. 2009.
Domoic acid production by Pseudo-nitzschia australis and Pseudo-nitzschiacalliantha
isolated from North Chile. Harmful Algae 8: 938–945
Amzil Z, Fresnel J, Le Gal D, Billard C.2001. Domoic acid accumulation in french sehllfish
in relation to toxic species of Pseudo-nitzschia multiseries and P.pseudodelicatissima.
toxicon, 39: 245–251.
Amzil Z, Sibat M, Royer F, Savar V. 2008. First report on azaspiracid and yessotoxin groups
detection in French shellfish. Toxicon, 52(1): 39–48.
Anabalón V, Arístegui J, Morales CE, Andrade I, Benavides M, Correa-Ramirez MA, Espino
M, Ettahiri O, Hormazabal S, Makaoui A, Montero MF, Orbi A. 2014. The structure of
planktonic communities under variable coastal upwelling conditions off Cape Ghir (31N) in
the Canary Current System (NW Africa). Prog. Oceanogr, 120, 320–339.
Anderson DM. 1989. Toxic algal bloom and red tides: a global perspective. En: Okaichi T,
Anderson DM, Nemoto T (Eds.), Red Tides: Biology, Environmental Science and
Technology. Elsevier, 11–16.
Anderson MJ. 2001. A new method for non-parametric multivariate analysis of variance.
Austral Ecol, 26: 32–46.
Anderson DM, Andersen P, Bricel VM, Cullen JJ, Rensel JE. 2001. Monitoring and
management strategies for harmful algal blooms in coastal waters. PEC#201-ME-01.1,
APEC Program, Singapore, and Intergovernmental Oceanographic Commission. Technical
Series, 59: 268 pp.
Anderson DM, Glibert PM,Burkholder JM. 2002. Harmful algal blooms and eutrophication:
nutrient sources, composition, and consequences. Estuaries, 25:704–726.
Anderson DM, Stock C, Keafer BA, Bronzino A, Thompson B, McGillicuddy D, Keller M,
Matrai PA, Martin J. 2005. Alexandrium fundyense cyst dynamics in the Gulf of Maine.
Deep-Sea Research II, 52:2522–2542.

91
Anderson MJ, Ellingsen K, McArdle B. 2006. Multivariate dispersion as a measure of beta
diversity. Ecology Letters, 9: 683–693.
Anderson MJ, Gorley RN, Clarke KR. 2008. PERMANOVA+ for PRIMER: guide to
software and statistical methods. PRIMER-e, Plymouth, UK.
Anderson DM. 2009. Approaches to monitoring, control and management of harmful algal
blooms (HABs). Ocean & Coast Manag 52: 342-347.
Anderson TR. 2010: Progress in marine ecosystem modelling and the “unreasonable
effectiveness of mathematics”. J Mar Syst, 81: 44–11.
Aoki K, Kameda T, Yamatogi T, Ishida N, Hirae S, Kawaguchi M, Syutou T. 2017. Spatio-
temporal variations in bloom of the red-tide dinoflagellate Karenia mikimotoi in Imari Bay,
Japan, in 2014: Factors controlling horizontal and vertical distribution. Marine Pollution
Bulletin, 124: 130-138.
Avaria S, Cassis D, Muñoz P, Vera P. 1997. Distribución del microfitoplancton marino en
aguas interiores del sur de Chile en Octubre de 1995 (Crucero Cimar-Fiordo I). Cienc Tec
Mar, 20: 107–123.
Avaria S, L Jorquera, Muñoz P, Vera P. 1999. Distribución del microfitoplancton marino en
la zona de aguas interiores comprendida entre el golfo de Penas y el Estrecho de Magallanes,
Chile, en la primavera de 1996 (Crucero CIMAR Fiordo 2). Cienc Tecnol Mar, 22: 81–110.
Avaria S, Caceres C, Castillo P, Muñoz P. 2003. Distribución del microfitoplancton marino
en la zona Estrecho de Magallanes – Cabo de Hornos, Chile, en la primavera de 1998.
Crucero CIMAR 3 Fiordos. Cienc Tecnol Mar, 26 (29): 79–96.
Balech E. 1989. Redescription of Alexandrium minimum Halim (Dinophyceae, type species
of genus Alexandrium. Phycologia, 28: 737–745.
Balech E. 1995. The genus Alexandrium Halim. Sherkin Island Marine Station, Sherkin
Island. 151 pp.
Barnes C, Maxwell D, Reuman DC, Jennings S. 2010. Global patterns in predator-prey size
relationships reveal size-dependency of trophic transfer efficiency. Ecology, 91:222-232.
Barnes MK, Tilstone GH, Suggett DJ, Widdicombe CE, Bruun J, Martinez-Vicente V, Smyth
TJ. 2015. Temporal variability in total, micro- and nano-phytoplankton primary production
at a coastal site in the western English Channel. Progress in Oceanography, 137: 470–483.
Bates SS, Bird CJ, de Freitas ASW, Foxall R, Gilgan M, Hanic LA, Johnson GR, Mcculloch
AW, Odense P, Pocklington R, Quilliam MA, Sim PG, Smith JC, Subba, Rao DV, Tood
ECD, Walter JA, Wright JLC. 1989. Pennate diatom Nitzschia pungensas the primary source

92
of domoic acid, a toxin in shellfish from eastern Prince Edward Island, Canada. Can J Fish
Aquat Sci, 46: 1203–1215.
Biecheler B. 1952. Recherches sur les Péridiniens. Bull Biol de la France et de la Belgique,
36: 1–147.
Blanco J, Álvarez G, Uribe E. 2007. Identification of pectenotoxins in plankton, filter feeders,
and isolated cells of a Dinophysis acuminate with an atypical toxin profile, from Chile.
Toxicon, 49: 710–716.
Bursa A. 1958. The freshwater dinoflagellateWoloszynnskia limnetica n. sp. Membrane
and protoplasmic structures. J Protozool, 5: 299–304.
Carrascal LM, Galvan I, Gordo O. 2009. Partial least squares regression as an alternative to
current regression methods used in ecology. Oikos, 118: 681–690.
Chatton E. 1952. Classe des Dinoflagelles ou Peridiniens. En: Grasse PP, (eds), Traitede
Zoologie. Masson and Cie, Paris, 309–390.
Clarke KR, Warwick RM. 2001. Change in marine communities: an approach to statistical
analysis and interpretation, 2nd edn. PRIMER-e, Plymouth, UK.
Clarke KR, Gorley RN. 2006. PRIMER v6: User manual/tutorial. PRIMER-e, Plymouth.
Clément A. 1999. Floraciones nocivas de Gymnodinium sp. y sus efectos en la
salmonicultura. Informe Técnico. Programa Monitoreo Fitoplancton INTESAL, 29 pp.
Collos Y. 1986. Time-lag algal growth dynamics: biological constraints on primary
production in aquatic ecosystems. Marine Ecology Progress Series, 33: 193–206.
Corliss JO. 1952. Comparative studies on holotrichous ciliates in the Colpidium-Glaucoma-
Leucophrys-Tetrahymena group. I. General considerations and history of strains in pure
culture. Trans Amer Micr Soc, 71: 159–84.
Dahl E, Tangen K. 1993. 25 years experience with Gyrodinium aureolum in Norwegian
waters. En: Smayda, T.J., Shimizu, Y. (Eds.), Toxic Phytoplankton Blooms in the Sea.
Developments in Marine Biology. Elsevier Science Publ BV, 15–21.
Davidson K, Miller P, Wilding TA, Shutler J, Bresnan E, Kennington K, Swan S. 2009. A
large and prolonged bloom of Karenia mikimotoi in Scottish waters in 2006. Harmful Algae,
8:349–361.
Davidson K, Gowen RJ, Tett P, Bresnan E, Harrison PJ, McKinney A, Milligan S, Mills DK,
Silke J, Crooks AM. 2012. Harmful algal blooms: How strong is the evidence that nutrient
ratios and forms influence their occurrence? Est. Coast. Shelf. Sci. 115: 399-413.

93
Del Amo Y, Queguiner B, Treguer P, Breton H, Lampert L. 1997. Impacts of high-nitrate
freshwater inputs on macrotidal ecosystems. II. Specific role of the silicic acid pump in the
year-round dominance of diatom in the Bay of Brest (France). Mar Ecol Prog Ser, 161:213–
224.
Díaz PA, Ruiz-Villarreal M, Velo-Suárez L, Ramilo I, Gentien P, Lunven M, Fernand L,
Raine R, Reguera B. 2014. Tidal and wind-event variability and the distribution of two
groups of Pseudo-nitzschia species in an upwelling-influenced Ría. Deep Sea Res II, 101:
163–179
Durham WM, Stocker R. 2012. Thin Phytoplankton Layers: Characteristics, Mechanisms,
and Consequences. Annu Rev Mar Sci, 4:177–207
Estrada M, Berdalet E, Vila M, Marrasé C. 2003. Effects of pulsed nutrient enrichment on
enclosed phytoplankton ecophysiological and successional responses. Aquatic Microbial
Ecology, 32: 61–71.
Evans D. 1975. The occurrence of Gyrodinium aureolum in the eastern Irish Sea, 1975.
Lancashire and Western Irish Sea Fisheries Joint Committee Scientific Report. pp. 85–89.
Faust MA, Gulledge RA. 2002. Identifying harmful marine dinoflagellates. Contributions
from the United States National Herbarium. 42, 1–144.
Fawcett A, Pitcher GC, Bernard S, Cembella AD, Kudela RM. 2007. Contrasting wind
patterns and toxigenic phytoplankton in the southern Benguela upwelling system. Marine
Ecology Progress Series, 348: 19–31.
Fehling J, Davidson K, Bolch CJ, Bates SS. 2004. Growth and domoic acid production of
Pseudo-nitzschia seriata (P.T. Cleve) H. Peragallo (Bacillariophyceae) under phosphate and
silicate limitation. Journal of Phycology 40: 674e683.
Field JG, Clarke KR, Warwick RM. 1982. A practical strategy for analyzing multispecies
distribution patterns. Mar Ecol Progr Ser, 8: 37–52.
Fleming LE, Kirkpatrick B, Backer LC, Walsh CJ, Nierenberg K, Clark J, Reich A,
Hollenbeck J, Benson J, Cheng YS, Naar J, Pierce R, Bourdelais AJ, Abraham WM,
Kirkpatrick G, Zaias J, Wanner A, Mendes E, Shalat S, Hoagland P, Stephan W, Bean J,
Watkins S, Clarke T, Byrne M, Baden DG. 2011. Review of Florida red tide and human
health effects. Harmful Algae 10(2): 224-233Fux E, Smith JR, Tong M, Guzman L,
Anderson DM. 2011. Toxin profiles of five geographical isolates of Dinophysis spp. from
North and South America. Toxicon, 57: 275–287.
Freer E, Vargas-Montero M. 2003. Floraciones algales nocivas en la costa pacífica de Costa
Rica: toxicología y sus efectos en el ecosistema y salud pública. Acta Médica Costarricense,
45(4): 158-164.

94
Fritz L, Triemer RE. 1985. A rapid simple technique utilizing Calcofluor white M2R for the
visualization of dinoflagellate thecal plates. J Phycol, 21: 662–664.
García C, González V, Cornejo C, Palma-Fleming H, Lagos N. 2004. First evidence of
Dinophysistoxin-1and carcinogenic polycyclic aromatic hydrocarbons in smoked bivalves
collected in the Patagonia fjords. Toxicon, 43: 121–131.
García C, Truan D, Lagos M, Santelices JP, Díaz JC, Lagos N. 2005. Metabolic
transformation of dinophysistox-in-3 into dinophysistoxin-1 causes human intoxication by
consumption of O-acyl-derivatives dinophysistoxins contaminated shellfish. J Toxicol Sci,
30: 287–296.
García-Soto C, Pingree RD. 2009. Spring and summer blooms of phytoplankton
(SeaWiFS/MODIS) along a ferry line in the Bay of Biscay and western English Channel.
Continental Shelf Research, 29: 111–1122.
Gentien P. 1998. Bloom dynamics and ecophysiology of the Gymnodinium mikimotoi
species complex. En: Anderson, D.L., Cembella, A.D., Hallegraeff, G.M. (Eds.),
Physiological Ecology of Harmful Algal Blooms. NATO ASI Series. Springer-Verlag, pp.
155–173.
Gentien P, Donaghay P, Yamazaki H, Raine R, Reguera B, Osborn T. 2005. Harmful algal
blooms in stratified environments. Oceanography, 18(2): 172–183.
Gómez F. 2007. Nuevos registros del dinoflagelado Craspedotella Kofoid, 1905
(Dinophyceae: Noctilucales) en el Océano Pacífico. Rev Biol Mar y Oceanogr, 42: 83–87.
Gowen RJ, Mckinney A, Tett P, Bresnan E, Davidson K, Harrison PJ, Milligan S, Mills DK,
Silke J, Crooks AM. 2012. Anthropogenic nutrient enrichment and blooms of harmful
phytoplankton. Oceanogr Mar Biol, 50: 65–126.
Granéli E, Flynn K. 2006. Chemical and physical factors influencing toxin content. In:
Granéli E, Turner TT (Eds.) Ecology of Harmful Algae, Springer, Berlin, pp. 229-241.
Guzmán L, Pizarro G, Alarcón C, Chacón V, Igor R, Barticevic E, Oyarzo O, Atalah A,
Foppiano J. 1999. Difusión Programa Marea Roja en la Región de Magallanes y Antártica
Chilena, Tercera Etapa. Primer Informe de Avance proyecto FNDR - XII Región / 20092815
- 0. Tomo I, 74 p. Instituto de Fomento Pesquero, Punta Arenas.
Guzmán L, Pacheco H, Pizarro G, Alarcón C. 2002.Alexandrium catenella y veneno
paralizante de los mariscos en Chile. En: E. A. Sar, M. E. Ferrario & B. Reguera (eds.).
Floraciones algales nocivas en el cono sur americano. Instituto Español de Oceanografía,
Madrid, pp. 237–256.
Hallegraeff GM. 1993. A review of harmful algal blooms and their apparent global increase.
Phycologia, 32: 79–99.

95
Harrison PJ, Conway HL, Holmes RW, Davis CO. 1977. Marine diatoms grown in
chemostats under silicate or ammonium limitation. 3. Cellular chemical composition and
morphology of Chaetoceros debilis, Skeletonema costatum and Thalassiosira gruvidu. Mar
Biol, 43: 19– 32.
Haywood AJ, Steidinger KA, Truby EW. 2004. Comparative morphology and molecular
phylogenetic analysis of three new species of the genus Karenia (Dinophyceae) from New
Zealand. J Phycol, 40: 165–179.
Hess P. 2002. Use of LC-MS testing to identify lipophilic toxins, to establish local trends and
interspecies differences and to test the comparability of LC-MS testing with the mouse
bioassay: an example from the Irish biotoxin monitoring programme 2001. En: Villalba A,
Reguera B, Romalde JL, Beiras R, editores. Fourth International Conference on Molluscan
Shellfish Safety. IOC of UNESCO; Santiago de Compostela: pp. 57–66.
Hess P. 2010. Requirements for screening and confirmatory methods for the detection and
quantification of marine biotoxins in end-product and official control. Anal Bio Chem,
397(5): 1683–94.
Heisler J, Glibert PM, Burkholder JM, Anderson DM, Cochlan W, Dennison WC, Dortch Q,
Gobler CJ, Heil CA, Humphries E, Lewitus A, Magnien R, Marshall HG, Sellner K,
Stockwell DA, Stoecker DK, Suddleson M. 2008. Eutrophication and harmful algal blooms:
a scientific consensus. Harmful Algae 8: 3-13.
Hoagland P, Anderson DM, Kaoru Y, White AW. 2002. The economic effects of harmful
algal blooms in the United States: estimates, assessment issues, and information needs.
Estuaries, 25:677–695.
Holm-Hansen O, Lorenzen CJ, Holmes RW, Strickland JDH. 1965. Fluorometric
determination of chlorophyll. J Cons Perm Int Explor Mer, 30: 3–15.
Horner RA, Garrison DL, Plumley FG. 1997. Harmful algal blooms and red tide problems
on the U.S. West Coast. Limnol and Ocean, 42:1076–1088.
Iriarte JL,González HE, Liu KK, Rivas C, Valenzuela C. 2007. Spatialand temporal
variability of chlorophyll and primary productivity in surface waters of southern Chile (41.5–
431S). Estuar Coast Shelf Sci, 74: 471–480.
Karayanni H, Christaki U, Van Wambeke F, Dalby AP. 2004. Evaluation of double formalin–
Lugol’s fixation in assessing number and biomass of ciliates: An example of estimations at
mesoscale in NE Atlantic, J Microbiol Methods, 56: 349–358.
Jauffrais T, Séchet V, Herrenknecht C, Truquet P, Véronique S, Tillmann U, Hess P. 2013.
Effect of environmental and nutritional factors on growth and azaspiracid production of the
dinoflagellate Azadinium spinosum. Harmful Algae, 27: 138–148.

96
Kudela RM, Cochlan WP, Roberts A. 2004. Spatial and temporal patterns of Pseudo-
nitzschia spp. in central California related to regional oceanography. Harmful Algal Blooms
2002. En: Steidinger KA, Landsberg JH, Tomas CR, Vargo GA, editors. Harmful Algae
2002; Proceedings of the Xth International Conference on Harmful Algae. Florida Fish and
Wildlife Conservation Commission and Intergovernmental Oceanographic Commission of
UNESCO, pp. 347–349.
Kim HG. 2006. Mitigation and controls of HABs. En: Granéli E, Turner JT, editores. Ecology
of harmful algae. Ecological Studies. Vol. 189. Springer-Verlag; Berlin, 327–338.
Kim G, Lee YW, Joung DJ, Kim KR, Kim K. 2006. Real-time monitoring of nutrient
concentrations and red-tide outbreaks in the southern sea of Korea. Geophys Res Lett, 33:
L13607.
Kirkpatrick B, Fleming LE, Squicciarini D, Backer LC, Clark R, Abraham W, Benson J,
Cheng YS, Johnson D, Pierce R, Zaias J, Bossart GD, Baden DG. 2004. Literature Review
of Florida Red Tide: Implications for Human Health Effects. Harmful Algae, 3(2): 99–115.
Lembeye G, Yasumoto T, Zhao J y Fernandez R. 1993. DSP outbreaks in chilean fiords. En:
Smayda TJ, Shimizu Y, eds. Toxic Phytoplankton Blooms in the Sea. Elsevier Science
Publishers, 525 – 529.
Lembeye G. 2008. Harmful algal blooms in the austral Chilean channels and fjords. Progress
in the oceanographic knowledge of Chilean interior waters, from Puerto Montt to Cape Horn.
En: Silva N y Palma S (eds.). Comité Oceanográfico Nacional - Pontificia Universidad
Católica de Valparaíso, Valparaíso, 99–103.
Levasseur ME, Therriault JC. 1987. Phytoplankton biomass and nutrient dynamics in a
tidally induced upwelling: the role of the N03:Si0, ratio. Mar Ecol Prog Ser, 39: 87–97
Li JT, Zhao WH, Yang DF, Wang JT. 2005. Effect of turbid water in Changjiang (Yangtze)
estuary on the growth of Skeletonema costatum. Marine Sciences, 29 (1): 34–37.
Li A, Jiang B, Chen H, Gu H. 2016. Growth and toxin production of Azadinium poporum
strains in batch cultures under different nutrient conditions. Ecotoxicol Environ Saf. 127:117-
26.
Liu YD, Sun J, Chen TZ, Wei TD. 2002. “Effect of N/P ratio on the growth of a red tide
diatom Skeletonema costatum,” Transaction of Oceanology and Limnology, 2: 39–44.
López-Rivera A, Pinto M, Uribe E, Álvarez G, Fernández Amandi M, Lehane M,Suárez-Isla
BA, James KJ. 2006. A new potential vector of domoic acid: The tunicate Pyura chilensis
(Molina). AOAC Task Force. AOAC, Baiona, Spain, 76 pp.

97
Louw,DC,vanderPlas A, Mohrholz V, Wasmund N, Junker T, Eggert A. 2016. Seasonal and
inter-annual phytoplankton dynamics and forcing mechanisms in the northern Benguela
upwelling system. J Mar Syst, 157: 124–134.
Lynn DH, Small EB. 2002. Phylum Ciliophora. En Lee JJ, Leedale GF, Bradbury P (eds),
An Illustrated Guide to the Protozoa. Society of Protozoologists, Lawrence, Kansas, 371–
656.
Maciel-Baltazar E, Hernandez-Becerril DU. 2017. Especies de dinoflagelados atecados
(Dinophyta) de la costa de Chiapas, sur del Pacífico mexicano. Rev Biol Mar Oceanogr,
48(2): 245–259.
Magaña HA, Contreras C, Villareal TA. 2003. A historical assessment of Karenia brevis in
the western Gulf of Mexico. Harmful Algae, 2:163–171.
Mahoney JB, Olsen P, Cohn M. 1990. Blooms of a dinoflagellate Gyrodinium cf aureolum
in New Jersey coastal waters and their occurrence and effects worldwide. Journal of Coastal
Research 6, 121–135.
Marchetti A, Trainer VL, Harrison PJ. 2004. Environmental conditions and phytoplankton
dynamics associated with Pseudo-nitzschia abundance and domoic acid in the Juan de Fuca
eddy. Marine Ecology Progress Series 281: 1-12.
Mayali X, Doucette G. 2002. Microbial community interactions and population dynamics
of an algicidal bacterium active against Karenia brevis. Harmful Algae, 1: 277–293.
Menschel E, González HE, Giesecke R. 2016. Coastal-oceanic distribution gradient of
coccolithophores and their role in thecarbonate flux of the upwelling systemoff Concepción,
Chile (36°S). Plankton Res 00(00): 1–20.
McArdle BH, Anderson MJ. 2001. Fitting multivariate models to community data: a
comment on distance-based redundancy analysis. Ecology, 82(1): 290-297.
Mevik BH, Wehrens R. 2007. The PLS package: principal components and partial least
squares regression in R. Journal of Statistical Software, 18: 1–24.
Moore C M, Mills MM, Arrigo KR, Berman-Frank I, Bopp L, Boyd PW, et al. 2013.
Processes and patterns of oceanic nutrient limitation. Nat. Geosci, 6: 701–710.
Morris PD, Campbell DS, Taylor TJ, Freeman JI. 1991. Clinical and epidemiological features
of neurotoxic shellfish poisoning in North Carolina. Am J Public Health, 81(4): 471–4.
Muñoz P, Savaria H, Sievers, Prado R. 1992. Presencia de dinoflagelados tóxicos del género
Dinophysis en el Seno Aysén, Chile. Revista de Biología Marina (Chile), 27: 187-212.

98
Muñoz P. 1985. Revisión taxonómica de los dinoflagelados de Chile. Rev Biol Mar, 21(1):
31-60.
Officer CB, Ryther JH. 1980. The possible importance of silicon in marine eutrophication.
Marine Ecology Progress Series. 3: 83-91.
Paredes MA, Montecinos V. 2011. Size diversity as an expression of phytoplankton
community structure and the identification of its patterns on the scale of fjords and channels.
Continental ShelfResearch, 31: 272–281
Pernthaler J, Posch T, Simek K, Vrba J, Pernthaler A, Glockner FO, et al. 2001. Predator-
specific enrichment of actinobacteria from a cosmopolitan freshwater clade inmixed
continuous culture. Appl Environ Microbiol, 67:2145–2155.
Pizarro G, Iriarte JL, Montecino V, Blanco JL, Guzmán L. 2000. Distribución de la biomasa
fitoplanctónica y productividad primaria máxima de fiordos y canales australes (47°S- 50°S)
en octubre de 1996. Cienc Tecnol. Mar, 23: 25-48.
Poole HH, Atkins WRG. 1929. Photoelectric measures of submarine illumination throughout
the year. J Mar Biol Ass UK, 16:297–324.
Quigg A, Finkel ZV, Irwin AJ, Rosenthal Y, Ho TY, Reinfelder JR, et al. 2003. The
evolutionary inheritance of elemental stoichiometry in marine phytoplankton. Nature, 425
291–294.
Ramírez B, Pizarro E. 2005. Distribución de clorofila a y feopigmentos en los canales
australes comprendidos entre Puer to Montt y la laguna San Rafael, Chile. Cienc Tecnol Mar,
28(1): 45-62.
Reguera B, Velo-Suárez L, Raine R, Park MG. 2012. Harmful Dinophysis species: a review.
Harmful Algae, 14: 87–106.
Rivera P. 1969. Sinopsis de las diatomeas de Bahía Concepción, Chile. Gayana Botánica,
18:1-112.
Rivera P. 1985. Las especies del género Nitzchia Hassal, sección Peudonitzschia
(Bacillariophyceae) en las aguas marinas chilenas. Gayana Botánica, 42:9-39.
Sanseverino I, Conduto D, Pozzoli L, Dobricic S, Lettieri T. 2016. Algal bloom and its
economic impact. Joint Research Centre, the European Commission. EUR27905EN
Scatasta S, Stolte W, Graneeli E, van Ierland E. 2003. The socio-economic impact of harmful
algal blooms in European marine waters and description of future risks. Fifth Deliverable for
the EU-funded project ECOHARM.

99
Sievers HA, Silva N. 2006. Masas de agua y circulación en los canales y fiordos australes.
En: Silva, N., S. Palma (eds.) Avances en el conocimiento oceanográfico der las aguas
interiores chilenas, Puerto Montt a cabo de Hornos. Comité Oceanográfico Nacional-
Pontificia Universidad Católica de Valparaiso, 53–58.
Silke J, Beirn FO, Cronin M. 2005. Karenia mikimotoi: an exceptional dinoflagellate bloom
in western irish waters, summer 2005. Marine Environment and Health Series, 21, 1649–
0053.
Silva N, Calvete C. 2002. Características oceanográficas físicas y químicas de canales
australes chilenos entre el golfo de Penas y el estrecho de Magallanes (Crucero CIMAR
Fiordo 2). Ciencia Tecnol Mar, 22(1): 23–88.
Silva N. 2008. Dissolved oxygen, pH, and nutrients in the austral Chilean channels and fjords.
En: Silva N, Palma S, editors. Progress in the oceanographic knowledge of Chilean interior
waters, from Puerto Montt to Cape Horn. Comité Oceanográphico Nacional-Pontificia
Universidad Católica de Valparaíso, Chile, 37–43.
Silva N, Palma S. 2008. The CIMAR Program in the austral Chilean channels and fjords. En:
Silva, N. & S. Palma. (eds.) Progress in the oceanographic knowledge of Chilean interior
waters, from Puerto Montt to Cape Horn. Comité Oceanográfico Nacional - Pontificia
Universidad Católica de Valparaíso, 11–15.
Smayda TJ. 1990. Novel and nuisance phytoplankton blooms in the sea: evidence for a global
epidemic. En: Graneli E, Sundstrom B, Edler L, Anderson DM, Eds.Toxic Marine
Phytoplankton. Elsevier, pp. 29–40.
Smayda TJ, Reynolds CS. 2001. Community assembly in marine phytoplankton: application
of recent models to harmful dinoflagellate blooms. Journal of Plankton Research, 23, 447–
461.
Sommer U. 1984. The paradox of the plankton fluctuations of phosphorus availability
maintain diversity of phytoplankton in flow-through cultures. Limnol and Ocean, 29: 633–
636.
Staats N, Stal LJ, Mur LR. 2000. Exopolysaccharide production by the epipelic diatom
Cylindrotheca closterium: effects of nutrient conditions. Journal of Experimental Marine
Biology and Ecology, 249:13–27.
Strickland JDH, Parsons TR. 1972. A practical handbook of seawater analysis. Fisheries
Researches Board of Canada, Bulletin, 167: 1–310.
Suárez-Isla BA, López-Rivera A, Hernández C, Clement A, Guzmán L. 2002. Impacto
económico de las floraciones de microalgas nocivas en Chile y datos recientes sobre la
ocurrencia de veneno amnésico de los mariscos. En: Sar E, Ferrario ME, Reguera B. (Eds.),

100
Floraciones Algales Nocivas en el Cono Sur Americano. Instituto Español de Oceanografía,
pp. 257–268.
Suikkanen S, Kremp A, Hautala H, Krock B. 2013.Paralytic shellfish toxins or spirolides?
The role of environmental and genetic factors in toxin production of the Alexandrium
ostenfeldii complex. Harmful Algae, 26: 52–59.
Sun BY, Liang SK, Wang CY, et al. 2008. Role of irradiance on the seasonality of
Skeletonema costatum Cleve blooms in the Coastal Area in East China Sea. Environ Sci,
29:1849–1854.
Sze, P. 1998. A Biology of the Algae. The McGraw Hill Companies Inc. 3rd edition.
WCB/McGraw-Hill, Boston. 277 pp
Taylor FJR, Fukuyo Y, Larsen J, Hallegraeff GM. 2003. Taxonomy of Harmful
Dinoflagellates. En Manual on Harmful Marine Microalgae; Hallegraeff GM, Anderson DM,
Cembella AD, eds. UNESCO Intergovernmental Oceanographic Commission: Paris, France,
389–432.
Technicon. 1977. Technicon Ind. Method 186-72/W: Silicate in water and seawater,
Technicon Ind. Systems, Tarrytown, 10591 pp.
Thompson PA, Guo MX, Harrison PJ. 1992. Effects of variation in temperature. i. on the
biochemical composition of eight species of marine phytoplankton. Journal of Phycology,
28: 481–488.
Tomas CR. 1997. Identifying marine phytoplankton. Academic press, Harcourt. Brace and
Company, Toronto, 858 pp.
Trainer VL, Hickey BM, Horner RA. 2002. Biological and physical dynamics of domoic acid
production off the Washington coast. Limnol and Oceanogr, 47:1438–1446.
Trainer VL, Yoshida T. 2014 Proceedings of the Workshop on Economic Impacts of Harmful
Algal Blooms on Fisheries and Aquaculture. PICES Scientific Reports, 47:85.
Twiner MJ, Dixon SJ, Trick CG. 2001. Toxic effects of Heterosigma akashiwo do not appear
to be mediated by hydrogen peroxide. Limnol. Oceanogr, 46, 1400–1405.
Twiner MJ, Rehmann N, Hess P, Doucette GJ. 2008. Azaspiracid shellfish poisoning: a
review on the chemistry, ecology, and toxicology with an emphasis on human health impacts.
Mar Drugs. 6(2): 39–72.
Uribe E, Álvarez G, Blanco J, González F, Rosales S, Ramírez P, Díaz R, Castillo C, Gajardo
T. 2010. Prospección y seguimiento de microalgas nocivas para la actividad de cultivo de
pectinidos en las regiones de Antofagasta, Atacama y Coquimbo. Informe Final FIP Nº 2007
20. 254 pp.

101
Uribe J, C Garcia, M Rivas, Lagos N. 2001. First report of diarrhetic shellfish toxins in the
Southern Magellanic fjords, Chile. Journal of Shellfish Research, 20 (1): 69–74.
Uribe JC, Ruiz M. 2001. Gymnodinium Brown Tide in the Magellanic Fjords, Southern
Chile. Revista de Biología Marina y Oceanografía, 36 (2): 155 – 164.
Van Dolah FM. 2000. Marine algal toxins: origins, health effects, and their increased
occurrence. Environ Health Perspect, 108 (1): 133–41.
Van Ruth PD, Qin JG, Branford AJ. 2012. Size dependent competition in centric diatoms as
a function of nitrogen and silicon availability. Open J Marine Sci, 2:33–42.
Varela D, Paredes J, Alves de Souza C, Seguel M, Sfeir M, Frangópulos M. 2012.
Intraregional variation among Alexandrium catenella (Dinophyceae) strains from southern
Chile: Morphological, toxicological and genetic diversity. Harmful Algae, 15: 8–18.
Verity PG, Sieracki ME. 1993. Use of color image analysis and epifluorescence microscopy
to measure plankton biomass. En: Kemp PF,Sherr BF, Sherr EB, Cole JJ (eds) Hand book of
methods in aquatic microbial ecology. Lewis Publ, London, 327 pp.
Villafañe VE, Reid FMH. 1995. Métodos de microscopía para la cuantificación del
fitoplancton. En Alveal K, Ferrario ME, Oliveira EC, Sar E, eds. Manual de Métodos
Ficológicos. Concepción: Edit. Universitaria, 169–185.
Watkins SM, Reich A, Fleming LE, Hammond R. 2008. Neurotoxic shellfish poisoning. Mar
Drugs, 6: 431–455.
Wells ML, Trainer VL, Smayda TJ, et al. 2015. Harmful algal blooms and climate change:
Learning from the past and present to forecast the future. Harmful algae, 49:68–93.
Yamamoto T, Hatta G. 2004. Pulsed nutrient supply as a factor inducing phytoplankton
diversity. Ecological Modelling, 171: 247–270.
Zamorano R, Marn M, Cabrera F, Figueroa D, Contreras C, Barriga A, Lagos N, Garca C.
2013. Determination of the variability of both hydrophilic and lipophilic toxins in endemic
wild bivalves and carnivorous gastropods from the Southern part of Chile, Food Additives &
Contaminants: Part A, 30(9): 1660–1677.
Zhu L, Wang Q. 2015. Numerical Mesocosm Experimental Study on Harmful Algal Blooms
of Two Algal Species in the East China Sea.Mathematical Problems in Engineering, Article
ID 169860, 8 pp
Zingone A, Enevoldsen HO. 2000. The diversity of harmful algal blooms: a challenge for
science and management. Ocean Coast Manag, 43: 725–748.