a symmetry mismatch at the site of rna packaging in the polymerase complex of dsrna bacteriophage...
TRANSCRIPT

Article No. jmbi.1999.3260 available online at http://www.idealibrary.com on J. Mol. Biol. (1999) 294, 357±372
A Symmetry Mismatch at the Site of RNA Packagingin the Polymerase Complex of dsRNABacteriophage fff6
Felix de Haas1, Anja O. Paatero2, Leonard Mindich3, Dennis H. Bamford2
and Stephen D. Fuller1,4*
Introduction
Viruses are cellular parasites. Viral genomesencode a limited range of unique functions andappropriate host cell activities for the othersrequired for viral replication. The viral polymeraseplays a special role in replication, and hence it isoften carried within the virion. This need is absol-ute for double-stranded RNA (dsRNA) viruses,since cells do not have enzymes which are capableof transcribing dsRNA templates. Consequently,
E-mail address of the corresponding author:[email protected]
Abbreviations used: dsRNA, double-stranded RNA;f6, Pseudomonas phage f6; SIRT, simultaneous iterativereconstruction technique; CTF, contrast transferfunction; FSC, Fourier shell correlation; FEG, ®eldemission gun.
0022-2836/99/470357±16 $30.00/0
the dsRNA viruses, whose hosts span the rangefrom bacteria through fungi and plants to humans,all share a requirement to introduce their ownvirion-associated dsRNA polymerase into the cellalong with their dsRNA genome.
The best studied dsRNA viruses belong to thediverse Reoviridae family. All have segmentedgenomes, share a common replication strategy(Palmer & Martin, 1982; Tyler & Fields, 1990) andassemble a polymerase complex which performs(ÿ) strand and new (�) strand synthesis in a com-partment shielded from the cytoplasm. Newly syn-thesized capped (�) strands are exported from thiscomplex to serve as messages for viral proteins.
Pseudomonas phage f6 is the best characterizedmember of the Cystoviridae, the family of dsRNAviruses which infects bacteria. This virus carries athree-segment genome and shares many features
1The Structural BiologyProgramme, EuropeanMolecular Biology LaboratoryMeyerhofstrasse 169117, Heidelberg, Germany2Department of Biosciences, andInstitute of BiotechnologyUniversity of Helsinki,Biocenter 2, P.O. Box 56FIN-00014, Helsinki, Finland3Department of MicrobiologyPublic Health ResearchInstitute, New York, NY10016, USA4Division of Structural BiologyWellcome Trust Centre forHuman Genetics, University ofOxford, Roosevelt DriveOxford, OX3 7BN, UK
The polymerase complex of the enveloped double-stranded RNA(dsRNA) bacteriophage f6 ful®ls a similar function to those of otherdsRNA viruses such as Reoviridae. The f6 complex comprises proteinP1, which forms the shell, and proteins P2, P4 and P7, which areinvolved in RNA synthesis and packaging. Icosahedral reconstructionsfrom cryo-electron micrographs of recombinant polymerase particlesrevealed a clear dodecahedral shell and weaker satellites. Difference ima-ging demonstrated that these weak satellites were the sites of P4 and P2within the complex. The structure determined by icosahedral reconstruc-tion was used as an initial model in an iterative reconstruction techniqueto examine the departures from icosahedral symmetry. This approachshowed that P4 and P2 contribute to structures at the 5-fold positions ofthe icosahedral P1 shell which lack 5-fold symmetry and appear in vari-able orientations. Reconstruction of isolated recombinant P4 showed thatit was a hexamer with a size and shape matching the satellite. Symmetrymismatch between the satellites and the shell could play a role in RNApackaging akin to that of the portal vertex of dsDNA phages in DNApackaging. This is the ®rst example of dsRNA virus in which the struc-ture of the polymerase complex has been determined without theassumption of icosahedral symmetry. Our result with f6 illustrates thesymmetry mismatch which may occur at the sites of RNA packaging inother dsRNA viruses such as members of the Reoviridae.
# 1999 Academic Press
Keywords: cryo-electron microscopy; dsRNA viruses; packaging;symmetry mismatch; three-dimensional reconstruction
*Corresponding author# 1999 Academic Press

358 Symmetry Mismatch in the �6 Polymerase Complex
with the Reoviridae. The f6 polymerase complexperforms the same functions with the obviousexception of capping, which would have no role ina prokaryotic host. Further, this functional hom-ology is matched by a corresponding structuralhomology; the arrangement of the proteins of thepolymerase complex in f6 is similar to that in theReoviridae. Consequently, f6 provides an avenuefor combining the power of prokaryotic geneticswith structural analysis to examine the commonstructural solutions to the functional problemsfacing any dsRNA virus.
Bacteriophage f6 packages its dsRNA genomewithin a three-layered virion (Figure 1(a)). A spike,formed by protein P3 and its membrane anchorprotein P6, projects from the lipid envelope whichsurrounds the virus (Kenney et al., 1992; see thescheme by Li et al., 1993). P3 bears the receptor-binding activity which allows f6 to attach to thepilus of its host, Pseudomonas syringae. The boundvirions are brought into contact with the cell sur-face by retraction of the pilus (Romantschuk &Bamford, 1985). Upon contact with the outer mem-brane of the cell, P6 mediates the fusion of the bac-terial and viral membranes (Bamford et al., 1987).This fusion releases the nucleocapsid (proteins P8,P7, P4, P2 and P1 with the dsRNA genome) andprotein P5 into the periplasmic space of the hostcell. Protein P5 is a lytic enzyme that causes localdigestion of the peptidoglycan layer and opens apath for the nucleocapsid to reach the inner mem-brane (Caldentey & Bamford, 1992; Mindich &Lehman, 1979). The nucleocapsid buds through theinner membrane and enters the cytoplasm by anevent which requires both a membrane potentialand protein P8 (Ojala et al., 1990; Olkkonen et al.,1990, 1991; Romantschuk et al., 1988). P8 is releasedupon entry of the nucleocapsid into the cytoplasmactivating the four-protein polymerase complex(P1, P2, P4 and P7).
The genome-containing polymerase complexsynthesizes (�) strands which serve as messagesfor the production of viral proteins. New polymer-ase complexes are assembled from proteins P1, P2,P4 and P7. These particles package the three (�)strand RNAs (small (s), medium (m), and large (l))in an NTP hydrolysis-dependent process (Frilander& Bamford, 1995; Frilander et al., 1995; Gottliebet al., 1991) . The three (�) strands have differentbinding af®nities to the polymerase particle(s > m > l) (Juuti & Bamford, 1995), pack (Mindich,1995) and enhance each others' packaging in thesame order (Frilander et al., 1995; Qiao et al., 1995).Packaging involves the recognition of the 50 sitesand continues until all the three segments are pack-aged (reviewed by Mindich, 1999).
A model for segment packaging accounts for its®delity (Onodera et al., 1998; Qiao et al., 1997).Empty particles are hypothesized to have exteriorbinding sites for segment s which disappear whenthe full complement of s is packaged. Once thesites for s are hidden, sites for m appear. Afterpackaging of m, the sites for m disappear and the
sites for l are exposed. The demonstration that thedistribution of the genes of the three-stranded f6genome among one, two or four segments leads totheir packaging (Onodera et al., 1995, 1998) and theproduction of infectious virions demonstrates the``segment-full'' nature of the control (Mindich,1999). f6 is the only dsRNA virus in which theseideas can be critically examined, since it is the onlyone in which in vitro packaging and replicationsystems have been established.
Recombinant methods for the production of sub-sets of the proteins of the virus have allowed theassignment of functions to the four proteins of thepolymerase complex (Butcher et al., 1997; Gottliebet al., 1988; Juuti & Bamford, 1995, 1997; Olkkonen& Bamford, 1987; Paatero et al., 1995). The 85 kDaP1 protein forms the framework of the complexand is the major site of RNA binding. The 75 kDaP2 protein is the RNA-dependent RNA polymerasewhich is lacking in the cell and provides theunique synthesis of RNA from dsRNA templates.The 35 kDa P4 protein is a non-speci®c NTPasewhich provides the energy for (�) strand packa-ging. The 17 kDa P7 protein also plays a role in thestable packaging of RNA and enhances the ®delityof the subsequent (�) strand synthesis.
Cryo-electron microscopy and image reconstruc-tion have been performed on f6 subassemblieswhich have been extracted from virions or gener-ated by expression in Escherichia coli (Butcher et al.,1997). The reconstructions reveal that the P8 shellwhich surrounds the polymerase complex in thenucleocapsid is arranged with T � 13 symmetry, asis the case for the corresponding layer in the Reo-viridae for which it has been studied (Hewat et al.,1992a,b; Prasad et al., 1988, 1992; Prasad & Chiu,1994; Yeager et al., 1990). The polymerase complexitself is a dodecahedral structure which appears tobe formed of 120 copies of P1 in two sets of distinctenvironments (Butcher et al., 1997). A similar``T � 2`` organization has been reported for theorbivirus, bluetongue virus (Grimes et al., 1998).Two morphological forms of the f6 polymerasecomplex were observed in the absence of genome.The expanded form had a smooth appearance andmatched the dimensions of the ®lled polymerasecomplex (Butcher et al., 1997) while the other,unexpanded, form exhibited a slightly smallerdiameter and a more angular appearance.
This study extends the previous work (Butcheret al., 1997) by using a more comprehensive andbetter characterized array of recombinant poly-merase complex particles and more precise cor-rections of the contrast transfer function (CTF) tobetter de®ne changes in the polymerase complexstructure associated with individual proteins.Initially, all reconstructions were performedusing the full icosahedral symmetry of the par-ticle and using our standard icosahedral recon-struction package (Baker & Cheng, 1996;Crowther, 1971; Fuller et al., 1996; Mancini et al.,1997). Essentially all reconstructions of sphericalviruses described in the literature utilize icosahe-

Figure 1. (a) Life-cycle of bac-teriophage f6. The schematic rep-resentation of the stages of thebacteriophage f6 are shown,including attachment, entry,uncoating, (�) strand synthesis,assembly of proheads, expansion,(�) strand packaging, (ÿ) strandsynthesis, late transcription cycle,P8 production, envelopment andlysis. (b) The proteins of bacterio-phage f6. Coomassie blue-stainedSDS-PAGE of the particle prep-arations used in this work. Lane 1shows the whole virion and revealsthe positions of the f6 structuralproteins. The other lanes show thesubviral particles produced byvarious constructs after puri®cationas described in Materials andMethods. Lane 2 shows the product
of pLM450 expression (P1247). Lane 3 shows P124, produced by pLM574. Lane 4 shows P127, produced by PAP6.Lane 5 shows P14, produced by pLM358. The monomer molecular masses of the components derived from theirsequences are: P1, 84,986; P2, 74,791; P4, 35,032; and P7, 17,300. N-terminal analysis (A.O.P., unpublished results)demonstrates that the migration of P4 as a doublet under some electrophoresis conditions does not represent cleavageof the protein and suggests that it represents variable unfolding of the protein which alters its migration.
Symmetry Mismatch in the �6 Polymerase Complex 359
dral symmetry and an icosahedral reconstructionprocedure (Baker et al., 1999). Our initial resultsshowed that while icosahedral symmetry was anappropriate description for the P1 shell, it wasnot appropriate for the other regions of the com-plex. We overcame the requirement for sym-metry by collecting large data sets of particlesand utilizing a model-based technique whichallows the use of lower symmetries.
Our results indicate that P4 and P2 form a satel-lite at the 5-fold vertex of the P1 shell which lacksthe expected 5-fold symmetry. Reconstruction ofpuri®ed, negatively stained P4 revealed that itforms hexamers with 6-fold symmetry whichmatch the size and shape of the satellites. This6-fold, 5-fold symmetry mismatch at the site ofRNA packaging may reveal a key feature of themechanism of RNA packaging in dsRNA viruses.
Results
Particle preparation
Recombinant particles and virus were preparedas described in Materials and Methods, and the
Table 1. Comparison of the stoichiometry (%) of proteins ininfectious virion
Protein f6 P1247
P1 100 100P2 100 70P4 100 80P7 100 90
stoichiometry of the proteins in the particles wasanalyzed using Coomassie blue stained gels. Theloading of the samples was adjusted to yield equalamounts of protein P1 (Figure 1(b)) and each pro-tein species compared to its amount in the virionsample. The relative amounts of proteins are givenin Table 1 to the nearest 10 %. None of the recom-binant particles has the full complement of theother proteins. Protein P7, when present, seems tobe closest to the amount found in the infective vir-ion. P4 is always underrepresented in the recombi-nant particles (typically 70 % of the virioncomplement).
Throughout this work, P1247 will be used todenote the recombinant polymerase complex con-taining all four proteins. P124, P14 and P127will be used to denote the recombinant polymerasecomplexes lacking proteins P7, lackingboth proteins P7 and P2 and lacking protein P4,respectively.
Morphology of fff6 subassemblies
The 510 AÊ diameter, smooth surfaced RNA con-taining polymerase complex which has been iso-
the recombinant particles to those found in the
P124 P127 P14
100 100 10080 80 070 0 700 100 0

360 Symmetry Mismatch in the �6 Polymerase Complex
lated from virus is shown for comparison with thecryo-electron micrographs of other subassemblies(Figure 2(a)). Cryo-electron micrographs of therecombinant f6 particles (Figure 2(b)-(e)) showpolyhedral structures of 470 AÊ diameter. The par-ticles often orient with their 2-fold or 5-fold axes ofsymmetry near the direction of view re¯ectingtheir dodecahedral shape. Individual images ofP1247, P124, P127 and P14 are not easily dis-tinguishable. The differences between the particlesbecame more apparent upon averaging (seebelow). All of the particles show expanded andunexpanded forms (arrows in Figure 2). Only theunexpanded forms were used for processing in thiswork, since these correspond in size to the particleprior to RNA packaging (Butcher et al., 1997). Prep-arations of P1 also present a polyhedral appear-
ance (Butcher et al., 1997), however the extensiveaggregation of the particles renders them unsuita-ble for averaging and processing.
Polymerase complex structures obtained byusing full icosahedral symmetry
The icosahedral reconstruction method usedhere was identical with that used previously(Butcher et al., 1997), except for the use of a muchlarger number (between 854, P14, and 1557, P124)of particles and a more precise correction of theCTF. This CTF correction was eased by the use ofdata from a ®eld emission gun (FEG) microscopeand allows the sizes of different components in amacro-molecular structure to be compared morereliably.
Figure 2. Electron micrographsof f6 subassemblies. Electronmicrographs of frozen hydrated f6recombinant particles. (a) Bacterio-phage f6 core (polymerase com-plex) ®lled with RNA, obtained byuncoating the whole virion. (b)-(e)Typical views of recombinantparticles P1247 (procapsid), P124,P14, P127 in unexpanded (whitearrows) and expanded (blackarrows) con®gurations. (e) Viewsof the expanded particles along the2-fold axis which present a hexago-nal pro®le in contrast to the spheri-cal pro®les given by views in thesame image which are not along asymmetry axis. (f) Image of puri-®ed P4 which has been negativelystained with ammonium molyb-date. The scale bars represent1000 AÊ .

Symmetry Mismatch in the �6 Polymerase Complex 361
Icosahedral reconstructions were calculated foreach of the four recombinant particles to a resol-ution of 25 AÊ . This resolution was selected on thebasis of cross-common lines phase residual (Fulleret al., 1996) and is close to the estimated Fouriershell correlation (FSC) (van Heel, 1987) of indepen-dent reconstructions. Radially depth-cued surfacerepresentations of the reconstructions along their5-fold axes are presented at a threshold of 1s,which yields a molecular volume close to thatexpected for a P1120P212P460P760 structure(Figure 3). All of the reconstructions show thedodecahedral shell described previously (Butcheret al., 1997) which represents the ``T � 2'' frame-work of P1 in the complex. Reconstructions ofP1247, P14 and P124 all show an additional fea-ture, a satellite structure (called a turret by Butcheret al., 1997) which lies just outside the indentationsof the 5-fold facets of the dodecahedral shell. Theirinterpretation is the central theme of the work pre-sented here. These satellite structures do not
appear in the P127 particles, which lack the packa-ging NTPase, P4. No consistent feature was corre-lated with the presence of the protein P7 whichwould represent a relatively small contribution tothe mass of the complex.
The details of the satellite structures are moreapparent in a view from the center of the particle(Figure 3(b), (e), (h) and (k)). A reproducible vari-ation in the relative masses of the satellites in thedifferent particles is evident. The satellites in theP14 reconstruction (Figure 3(h)) are signi®cantlysmaller (20 %) than those seen in the P1247(Figure 3(b)) and P124 (Figure 3(e)) reconstructions.No satellite is seen in the P127 particle (Figure 3(k))which lacks P4 entirely. Hence, the satellite struc-ture requires the presence of protein P4 andappears to increase in mass (or order) when P2 ispresent.
Central sections through the reconstructions(Figure 3(c), (f) and (i)) reveal that the maximumdensity of the satellites is less than half of that
Figure 3. Views of the icosahe-dral reconstructions of recombinantparticles. The radially depth-cued(red, high radius; green, lowradius) surface representation ofthe reconstructions are viewedalong their 5-fold symmetry axes((a), (d), (g) and (j)) from outsidethe particle or from the center ofthe particle after clipping away thenear half of the density ((b), (e), (h)and (k)). Slices through the vertexregion and the satellite structures((c), (f), (i) and (l)) reveal the rela-tive density of the shell and otherfeatures. (a), (b), (c) P1247; (d), (e),(f) P124; (g), (h), (i) P14; (j), (k), (l)P127. A satellite structure ismarked with an arrow in (c), (f)and (i). The scale bars represent100 AÊ .

Figure 4. Reproducibility of independent reconstruc-tions. Fourier shell correlation (FSC) of reconstructionsperformed from independent data sets show the repro-ducibility of the (a) icosahedral reconstructions and ofthe model-based iterative reconstructions using (b) nosymmetry or (c) D5 symmetry. The resolution at whichthe FSC curve crosses 0.5 de®nes the resolution of fea-tures which are reproducible in the reconstruction. Thisresolution (Table 2) varies for the different particles andimproves with greater averaging.
362 Symmetry Mismatch in the �6 Polymerase Complex
found in the shell. Features which obey icosahedralsymmetry and are coarser than the resolutionshould appear with the same apparent density inthe reconstruction. Structures which lack icosahe-dral symmetry will appear with lower apparentdensity because they are inappropriately averagedby the reconstruction process. We employed areconstruction method which was not dependentupon icosahedral symmetry to characterize the sat-ellite structure.
Reconstructions without icosahedral symmetry
The model-based procedure eliminates theassumption of icosahedral symmetry at theexpense of computational ef®ciency. We used thefully icosahedrally symmetric model of the struc-ture described above to generate initial orientationsand then applied an iterative approach (seeMaterials and Methods) to modify the structure tocompensate for departures from the model. Thisapproach allows us to examine perturbations tothe well-de®ned icosahedral structure. Reconstruc-tions of the recombinant particles using the modelbased iterative procedure and icosahedral sym-metry were noisier but similiar to those producedby the icosahedral reconstruction method (data notshown). The reproducibility of the reconstructionwas determined by FSC (van Heel, 1987) of recon-structions derived from independent sets of data.The FSC calculation showed that the best of themodel-based iterative reconstructions without sym-metry was reproducible to 45 AÊ (P124 and P127;Figure 4(b)). The application of D5 symmetry(using the 5-fold symmetry along z and the per-pendicular 2-fold axis rather than the full 532 sym-metry) during the reconstruction increased theaveraging and improved the resolution (Figure 4(c))of these best reconstructions to 34 AÊ (P124) and36 AÊ (P127).
The reconstruction of P124 without the use ofsymmetry is shown in Figure 5(a)-(c). The 1 sthreshold yields a surface view (Figure 5(a) and(b)) in which the indentations contain differingamounts of material. A section (Figure 5(c))through the reconstruction at a height whichpasses through the satellite structures reveals thatthe densities of these structures is not only lowerthan that in the polyhedral wall, but also differentfor each position (Figure 6). The reconstructiondemonstrates that fewer than 12 copies of the satel-lite structure are present on the particle.
The application of D5 symmetry during themodel based iterative reconstruction (Figure 5(d)-(f)) increases the averaging by tenfold, but does soat the expense of hiding the variation between thesatellites which lie off the 5-fold axis. It imposesstrict 5-fold symmetry on the two satellites whichlie along the symmetry axis and imposes no sym-metry on the other satellites. The D5 averaged off-axis satellites (Figure 5(d) and (e)) reveal that stoi-chiometry is not the only failing of the satellite

Figure 5. Reconstructions ofrecombinant particles. Views ofreconstructions of recombinant par-ticles are presented as radiallydepth-cued surface views ((a), (b),(d), (e), (g), (h), (j) and (k)) and sec-tions ((c), (f), (i) and (j)) throughthe satellite structures. Views alonga 5-fold ((a), (d), (g) and (h)) and a((b), (e), (h) and (k)) 2-fold axis ofthe shell are shown. (a) and (b) TheP124 reconstruction performedusing no symmetry. (c) and (d) TheP124 reconstruction performedwith D5 symmetry. The effect ofthe absence of P4 is shown by theSIRT reconstruction of P127 with-out (e) and (f) symmetry and using(g) and (h) D5 symmetry. Note thatall 12 satellite structures are inde-pendent in the absence of sym-metry. D5 symmetry forces twosatellites to show 5-fold symmetry(one of which is seen at (d) 6:00and (e) 12:00) and averages theother ten copies of the satellitestructure although it does notimpose any symmetry on them.The scale bars represent 100 AÊ .
Symmetry Mismatch in the �6 Polymerase Complex 363
structures. The satellites do not share the 5-foldsymmetry of the shell.
Application of the reconstruction procedure tothe P1247 and P14 recombinant particles yieldsresults similar to those obtained for the P124 par-ticle (data not shown). Occupancy was never com-plete and the satellite structures lack local 5-foldsymmetry. The satellites in the P14 recombinanthave a 20 % lower density than in the P1247 andP124 in agreement with the contribution of P2 tothe structure suggested by icosahedral reconstruc-tion. The reconstruction of the P127 recombinantparticle without symmetry (Figure 5(g)-(i)) or usingD5 symmetry (Figure 5(j)-(l)) reveals that the satel-lite structures are completely absent as they werein the icosahedral reconstructions.
Tests of the validity of the reconstruction
A mismatch of symmetry between the satelliteand its position on the P1 shell raises the questionof the validity of the reconstruction. Averagingdifferent views of the particle does not requiresymmetry but it does require that all particles inthe average be identical. A symmetry mismatchwould allow multiple distinguishable positions ofthe satellites on the shell. If N distinct positionswere possible for each satellite, the number of dis-tinguishable polymerase structures would be N11.The modest resolution in our reconstruction(�30 AÊ ) could reduce N to 2; however, this is stillfar more distinct structures than we determine inour reconstructions. Unless serendipity selects one

Figure 6. Departure from sym-metry during iterations of recon-struction. (a) The course of thereconstruction of the P1247 poly-merase structure without symmetryis shown in (a) by the change inthe density at the positions of the12 independent satellite structures.The initial model (&) used tode®ne the orientations of the par-ticle images in the data containedthe 12 identical satellite structuresfrom the icosahedral reconstructionof the P1247 particle (Figure 3(a),(b) and (c)). This was used togroup views by orientation for the®rst round of the SIRT reconstruc-tion. The densities of the satellitesafter this ®rst round are no longeridentical (*). This asymmetricstructure is used to determine theorientations and group the viewsfor the next cycle. (b) Repeatedcycles enhance the differencesbetween the densities in the inde-pendent satellites until the fourthcycle produces the distribution ofdensities (X) and the satellite struc-tures. (c) The surface views of thesatellites are arranged in pairs sothat opposing structures are atopeach other (1 is opposed to 6, 2 to7, etc.) and that the top half set ofsatellites is shown above the lowerhalf. The enhancement of the den-sity of a particular satellite isdependent upon the asymmetry inthe model as shown by rotating thethird cycle density by 180 � alongthe 2-fold before a further cycle(}). The corresponding satellite inthe rotated structure is enhancedafter the fourth cycle of re®nementshowing that the results are inde-pendent of the initial model orien-tation. (c) The positions of the 12numbered icosahedral 5-fold axeswith respect to the xyz coordinatesystem. The 5-fold axis occupied by11 and 12 is aligned with the z-axis. The icosahedral 2-fold runningbetween 5 and 7 (or 2 and 10) isaligned with the y-axis which wasthe axis used for the 180 � test.
364 Symmetry Mismatch in the �6 Polymerase Complex
set of positions in a majority of the particles in ourpopulation, our reconstruction should produce asmeared average for the satellites.
The progressive departure of our averages fromicosahedral symmetry is seen by the changes in thedensity (plotted as the volume enclosed by a 0.5scontour) at the 12 satellites during the cycles of the
reconstruction (Figure 6). The initial model (the ico-sahedral reconstruction of P1247) has equal den-sities at all 12 positions. The use of this model togroup particle images by orientation leads, bychance, to an unequal distribution of satellite den-sities in the resulting reconstruction. This asym-metric model then selects images which are

Table 2. Reproducibility of independent reconstructions
Symmetry appliedComplex No. particles Icosahedral (AÊ ) D5 (AÊ ) None (AÊ )
P1247 779 22 40 56P124 1557 25 34 45P14 854 28 43 48P127 1179 25 36 45
The resolution to which the reconstructions are reproducible are shown by the point at which the Fourier shell correlation(Figure 4) of independent reconstuctions reaches the value of 0.5. The precise value of the cutoff is relatively unimportant due to thesteepness of the FSC curve (van Heel, 1987).
Symmetry Mismatch in the �6 Polymerase Complex 365
consistent with its projections in subsequent cycles.The result is an enhancement of the differencesbetween the satellites in subsequent cycles, sinceparticle images which agree in satellite orientationwith the model are averaged constructively whileimages from other orientations interfere to blur theaverage. The values after four cycles break intothree groups (Figure 6(a)). Two of the satellites (4and 11) have relatively high values, three satellites(7, 1, and 2) have relatively low values, and theremaining seven have similar, intermediate values.The fact that the seven intermediate values matchthose in the icosahedral reconstruction is consistentwith their being inappropriately averaged. The twosatellites which are enhanced by averaging showan increase in density which exceeds the inputmodel as would occur if these features were nowbeing appropriately averaged. Hence, our recon-struction overcomes the problem of multiple orien-tations for the satellites by aligning only a subsetof the sites. FSC analysis of aligned reconstructionsderived from independent data sets indicates thatthe departures from icosahedral symmetry are afeature of the population and not an artifact of thereconstruction method.
Controls using combinations of models andmodel projections were used to de®ne the limits ofthis procedure. Simulated projections of modelslacking an opposed pair of satellites give rise toaverages which lack opposed satellites even whena fully occupied structure is used for orientation.The importance of the model in selecting a particu-lar group of orientations can be seen by rotatingthe model by 180 � after three cycles and observingthat the fourth cycle produces the satellite densitiesin rotated positions (Figure 6(a), open diamonds).These tests indicate that the reconstruction sup-ports variable orientations of the satellite on the P1shell. Our reconstruction without symmetry wouldhave generated a clear, non-icosahedral structure ifthe majority of the P1247 particles had identical,non-icosahedral structures. The results (Figure 6)also support occupancy of three-quarters of thesites and dimensions of the averaged satellite struc-ture similar to those seen in Figure 5.
Reconstruction of purified protein P4
Reconstructions of the complete recombinantpolymerase complex suggested that the satellitestructure did not have local 5-fold symmetry. We
took advantage of the availability of puri®ed, bio-logically active recombinant P4 (Juuti et al., 1998)to examine protein P4 in isolation and determineits structure and symmetry.
An electron micrograph (Figure 2(f)) showing P4complexes in an unsupported layer of stain over ahole, reveals both rounded and rectangular views.Two-dimensional averages of these characteristicviews are seen in Figure 7(b) and (d). Analysis oftilted pairs of images allowed the assignment ofparticles to either a one-layered or a two-layeredstructure and showed that 20 % of the particles inthe preparation contain two layers. A reconstruc-tion using the tilt pair data for the 80 % of the par-ticles which are single-layered is shown inFigure 7(c), (e), (g) and (h) with the averages corre-sponding to views (c) and (e) shown in (d) and (f),respectively. The single layer comprises six 40 AÊ
diameter domains with ®ne projections extendingfrom each domain. Tilt pair data were necessary toreveal the heterogeneity, since the single-layeredparticle lies ¯at on the grid and hence presents noside view. As a result, image processing and three-dimensional reconstruction using 1250 untiltedparticle images and applying 3-fold, 6-fold or noaxial symmetry produced the two-layeredstructure seen in Figure 7(a). Each layer appears toconsist of six 40 AÊ diameter domains skewed withrespect to each other and joined to the opposinglayer by six ®ne connections at the edges. FSCanalysis con®rmed the reproducibility of bothreconstructions to 20 AÊ resolution. The agreementbetween the reconstructions using single anddouble-layered particles con®rms that the orien-tation of the particle in the stain layer does notaffect the structure.
Analytical ultracentrifuge data reveal that P4 isabout 4.4 nm thick with a hydrodynamic diameterof about 6.6 nm (Juuti et al., 1998), demonstratingthat the single-layered structure corresponds to thespecies found in solution and that the six-memberring is a hexamer of P4. A minority of the hexam-ers in the preparation tend to associate under ourconditions to form dodecamers.
Analysis of P4 reconstructions with imposed 5-fold symmetry and rotational analysis of averagedimages provided no evidence for 5-fold symmetricparticles. Further, we were never able to identify a5-fold axial view in our population of images oftilted and untilted particles.
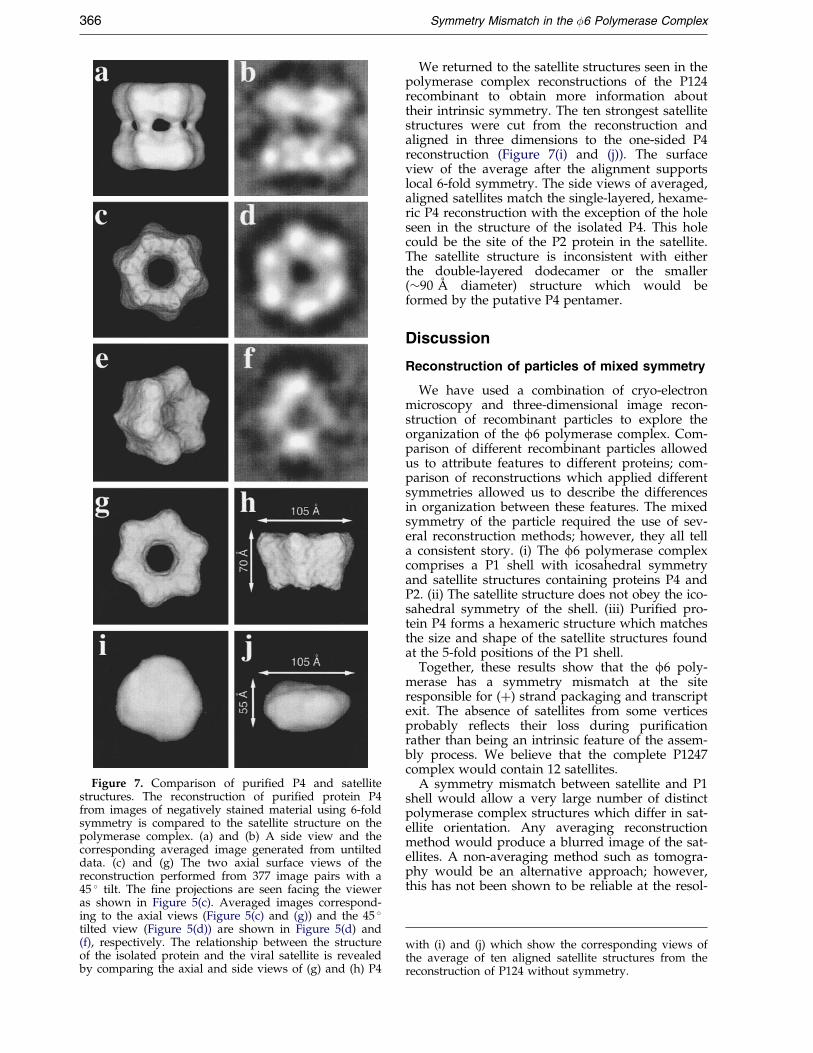
Figure 7. Comparison of puri®ed P4 and satellitestructures. The reconstruction of puri®ed protein P4from images of negatively stained material using 6-foldsymmetry is compared to the satellite structure on thepolymerase complex. (a) and (b) A side view and thecorresponding averaged image generated from untilteddata. (c) and (g) The two axial surface views of thereconstruction performed from 377 image pairs with a45 � tilt. The ®ne projections are seen facing the vieweras shown in Figure 5(c). Averaged images correspond-ing to the axial views (Figure 5(c) and (g)) and the 45 �tilted view (Figure 5(d)) are shown in Figure 5(d) and(f), respectively. The relationship between the structureof the isolated protein and the viral satellite is revealedby comparing the axial and side views of (g) and (h) P4
366 Symmetry Mismatch in the �6 Polymerase Complex
We returned to the satellite structures seen in thepolymerase complex reconstructions of the P124recombinant to obtain more information abouttheir intrinsic symmetry. The ten strongest satellitestructures were cut from the reconstruction andaligned in three dimensions to the one-sided P4reconstruction (Figure 7(i) and (j)). The surfaceview of the average after the alignment supportslocal 6-fold symmetry. The side views of averaged,aligned satellites match the single-layered, hexame-ric P4 reconstruction with the exception of the holeseen in the structure of the isolated P4. This holecould be the site of the P2 protein in the satellite.The satellite structure is inconsistent with eitherthe double-layered dodecamer or the smaller(�90 AÊ diameter) structure which would beformed by the putative P4 pentamer.
Discussion
Reconstruction of particles of mixed symmetry
We have used a combination of cryo-electronmicroscopy and three-dimensional image recon-struction of recombinant particles to explore theorganization of the f6 polymerase complex. Com-parison of different recombinant particles allowedus to attribute features to different proteins; com-parison of reconstructions which applied differentsymmetries allowed us to describe the differencesin organization between these features. The mixedsymmetry of the particle required the use of sev-eral reconstruction methods; however, they all tella consistent story. (i) The f6 polymerase complexcomprises a P1 shell with icosahedral symmetryand satellite structures containing proteins P4 andP2. (ii) The satellite structure does not obey the ico-sahedral symmetry of the shell. (iii) Puri®ed pro-tein P4 forms a hexameric structure which matchesthe size and shape of the satellite structures foundat the 5-fold positions of the P1 shell.
Together, these results show that the f6 poly-merase has a symmetry mismatch at the siteresponsible for (�) strand packaging and transcriptexit. The absence of satellites from some verticesprobably re¯ects their loss during puri®cationrather than being an intrinsic feature of the assem-bly process. We believe that the complete P1247complex would contain 12 satellites.
A symmetry mismatch between satellite and P1shell would allow a very large number of distinctpolymerase complex structures which differ in sat-ellite orientation. Any averaging reconstructionmethod would produce a blurred image of the sat-ellites. A non-averaging method such as tomogra-phy would be an alternative approach; however,this has not been shown to be reliable at the resol-
with (i) and (j) which show the corresponding views ofthe average of ten aligned satellite structures from thereconstruction of P124 without symmetry.

Symmetry Mismatch in the �6 Polymerase Complex 367
ution required to identify satellite orientations. Thedistribution of satellite densities seen in our aver-age structures supports its variable orientation andthe occupancy of a majority of the sites. The mod-est resolution of the non-symmetrized reconstruc-tions re¯ects this variability and limits theprecision with which we can describe the structure.Nevertheless, the argument for a symmetry mis-match does not rely upon a single reconstruction,but rather the consistent results of independentreconstructions of several particles. The icosahedralreconstructions indicate that the satellite does notobey the icosahedral symmetry of the shell. Themodel-based iterative reconstruction of the poly-merase complex con®rmed this lack of symmetry.The reconstruction of isolated P4 identi®ed itsbasis as the lack of 5-fold symmetry in the isolatedprotein. The fact that dimensions and shape of theaveraged satellite match those of the P4 hexamerseen in isolation strongly suggest that the samehexamer is present at the 5-fold site. The only sig-ni®cant difference (at the level of resolutionde®ned by FSC) from the structure of the isolatedP4 is the ®lled center of the satellite. This mayre¯ect the position of P2.
Departures from icosahedral symmetry havebeen noted previously in icosahedral reconstruc-tions. There are two fundamentally distinct waysin which the satellite structure could disobey icosa-hedral symmetry: low occupancy and inappropri-ate local symmetry. If the satellite structure wereonly present at half of the 12 5-fold positions buthad local 5-fold symmetry, it would be representedat roughly half of the expected density as was seenfor the Psu protein in bacteriophage P4 (Doklandet al., 1992, 1993). Inappropriate local symmetrywould describe the case in which 12 copies of thesatellite structure are present but lack local 5-foldsymmetry. The representation of the adenovirus II®ber at 20 % of its expected density is an exampleof this case (Stewart et al., 1991).
These cases are not exclusive as seen by the pre-sence of both in the polymerase complex. Eitherinvalidates the use of icosahedral reconstructionalone for determining the structure of the complex.Our model based approach builds on the icosahe-dral structure and uses iteration to de®ne thedeparture from symmetry. The results of thisapproach must be carefully examined for modelbias by FSC, since it reinforces departures from thewell de®ned icosahedral structure. With this cau-tion, the model based approach may be a usefulone for other structures of mixed symmetry.
Functional implications of symmetry mismatch
Symmetry is such a familiar concept in structuralvirology that one is drawn immediately to try to®nd a functional explanation for its violation. Ourunderstanding of the principles of virus assembly(Caspar, 1964, 1979, 1980; Caspar & Klug, 1962)motivates such a reaction. Virus assembly is robustbecause it utilizes regularity to reduce the number
of different types of interactions necessary. A lackof regularity should have a cost for the ef®ciencyof the assembly process which must be compen-sated by a functional role. Whether or not one isconvinced by this logic, the observation is thatsymmetry mismatches often occur at sites ofspecial functional signi®cance.
Symmetry mismatch has been described for bac-teriophages (Moody, 1965). The portal vertex ofdsDNA phages is one of the best characterizedexamples of a functionally important symmetrymismatch (Bazinet & King, 1985). These virusesassemble an empty procapsid and then proceed topack their genome into it through a specializedportal vertex in an energy dependent process. Theenergy to accomplish the packaging of the genomeis generated by ATP hydrolysis by a terminase thatremains associated with the cut end of the genomeas it attaches to the portal. A 10 AÊ resolution struc-ture of one of the f29 DNA tranlocating machineshas recently been presented (Valpuesta et al., 1999),demonstrating its unambiguous 12-fold symmetry.Although a variation of symmetry has beensuggested for different dsDNA phages (van Heelet al., 1996), the symmetry of the portal nevermatches its location on a 5-fold symmetry axis ofthe procapsid. The actual mechanism by whichDNA is packaged through this structure remainsunclear; however, the mismatch was taken as anindication that the portal may rotate with respectto the procapsid during the DNA translocation(Hendrix, 1978). This story has received anadditional twist recently with the discovery of ahexameric pRNA in dsDNA bacteriophage f29(Guo et al., 1998; Hendrix, 1998; Zhang et al., 1998).One model proposes that the pRNA interactssequentially with the genome to convert the energyof ATP hydrolysis to a rotation and translation ofthe DNA (Guo et al., 1998). While pRNA has onlybeen identi®ed in f29, the general features of thetranslocating portal structure are seen in the otherdsDNA phages and may be present in dsDNAviruses of mammals (Hendrix, 1998). It is signi®-cant that the role of the pRNA was established inf29 because the system is so well de®ned in vitro.Similarly, the work presented here has exploitedthe relatively advanced state of the in vitro f6system in comparison to other dsRNA viruses inour structural and functional analyses.
The features of the satellite structure in thedsRNA virus f6 parallel those of the DNA translo-cating portal. Protein P4, the major component ofthis structure, is the NTPase which powers thepackaging of the (�) strand. The symmetry mis-match of the P4 hexamer and the location of thesatellite at the 5-fold position of the P1 shell mayallow it to rotate, and hence ease the translocationof the (�) strands. Our results support the associ-ation of P2, the RNA-dependent polymerase, withthe satellite structure. P7, which plays a role in thestability of the packaged RNA, could also be pre-sent here, although it is too small to be detected inour reconstructions.

368 Symmetry Mismatch in the �6 Polymerase Complex
This organization of components suggests amodel of the polymerase complex in which theestablished enzymatic functions are associatedwith the satellite structures. This may ease the dif-®culties of continually and simultaneously replicat-ing the several segments of the viral genome toproduce new (�) strands which are exported foruse as message. The RNA polymerization site inprotein P2 should lie towards the interior of theparticle where it would be accessible to the pack-aged RNA so that movement of the template to thedistal portion of the satellite can be avoided. Theremust also be communication between the P1 shelland the external density at the 5-fold axes. Thiscould be accomplished by the major conformation-al changes which occur during maturation includ-ing expansion of the particle (Figure 2) andmovement of the 5-fold outwards (Butcher et al.,1997). Expansion may provide the signal to thesatellite that the P1 shell is full and that RNAsynthesis can begin.
Conclusions
Our work has combined icosahedral and model-based iterative reconstruction of the f6 polymerasecomplex with the structure determination of theisolated P4 protein to demonstrate the presenceof a symmetry mismatch in the portion of thepolymerase complex which is involved in RNApackaging. The use of a reconstruction with (inap-propriate) icosahedral symmetry as a startingmodel for de®ning the nature of non-icosahedralcomponents may be a useful one in other systemswhere a departure from symmetry is suspected.
The f6 system is a particularly convenient onefor studying the phenomena of packaging and theregulation of replication. A simple, well-de®nedsystem of four recombinant proteins can accom-plish both packaging and replication (Frilander &Bamford, 1995; Gottlieb et al., 1990; Juuti &Bamford, 1995; Ojala & Bamford, 1995). The factthat other dsRNA viruses such as the Reoviridaeremain less tractable motivates an extension of thelessons of the f6 system to them. f6 differs fromthe dsDNA phages which use a single portal forboth entry and exit of the dsDNA in its simul-taneous use of multiple sites. The mechanismwhich we describe above accommodates the con-trol of multiple entry sites in a way which is inde-pendent of the number of segments in the genome.Indeed, f6 manages perfectly well when its gen-ome is split into four strands rather than its accus-tomed three (Onodera et al., 1995). Adaptation tothe Reoviridae which utilize ten to 12-strand gen-omes would present no conceptual problem, andknowledge of the f6 model may guide the identi®-cation of corresponding components in otherdsRNA viruses although one must be careful indrawing conclusions across such a broad biologicalspectrum (Mindich, 1999). Comparison of themechanism by which analogous functions are
accomplished in different systems is a promisingpath to a deeper understanding of the importantfeatures of each.
Materials and Methods
Recombinant particle preparation
Complete and incomplete procapsids were producedin E. coli JM109 cells from plasmids pLM450, pLM358,and pLM574 (Gottlieb et al., 1990) , and in HMS174(DE3)cells from plasmid pAP6 (Paatero et al., 1998). Particleswere puri®ed in principle as described (Frilander &Bamford, 1995). The overnight culture was induced by1 mM IPTG. Protein expression was carried out over-night at 23 �C, except for pLM450 for 4.5 hours at 28 �C.The collected cells were lysed by passing twice througha French pressure cell and extracted with 5 % (v/v) andthen 3 % (v/v) TritonX-114. The particles were sedimen-ted through a sucrose gradient (Beckman SW40 rotor,27,000 rpm, two hours and 15 minutes, 10 �C; 5 %-20 %sucrose, 20 mM Tris (pH 8), 100 mM NaCl, 1 mM ATP,0.25 mM CaCl2). The light-scattering bands were col-lected and concentrated either by pelleting (BeckmanTSL-55 rotor, 55,000 rpm, 25 minutes, 4 �C) and resus-pending into 20 mM Tris (pH 8), 100 mM NaCl, 1 mMATP, 0.25 mM CaCl2 (pLM450, 358, 574) or by dialyzingunder vacuum against 20 mM Tris (pH 8), 100 mM NaClin a Sartorius collodion bag (cut off of 12 kDa) for4.5 hours at 4 �C (pAP6). The puri®ed particles were ana-lyzed by SDS-PAGE (Olkkonen & Bamford, 1989) andthe protein bands in the gel were quanti®ed by imageanalysis of Commassie blue-stained gels (BioImage,Whole Band analysis software).
Electron microscopy
f6 recombinant particles were resuspended to a con-centration of 0.3 mg/ml in 50 mM Tris, 100 mM NaClafter pelleting for cryo-electron microscopy. Coppergrids covered with a thin ®lm of carbon with 0.5 to 1 mmholes (Fukami & Adachi, 1965) were rendered morehydrophilic by glow discharge (Dubochet & Groom,1982) for one second immediately before use. A 5 ml dro-plet containing the sample was applied to the grid andthe excess solution was removed by blotting with a pieceof ®lter paper, after which the grid was plunged intofreshly prepared liquid ethane slush held in a bath ofliquid nitrogen (Adrian et al., 1984). The grid was thentransferred to a GATAN cryo-holder and observed in aPhilips CM200FEG at ÿ168 �C. Pairs of images (nominaldefocus values of ÿ1 and ÿ5 mm) were taken at a nom-inal magni®cation of 38,000� under low-dose conditions.
Puri®ed protein P4 (Juuti et al., 1998) was prepared forelectron microscopy using ammonium molybdate as anegative stain. Copper grids covered with a holey carbon®lm were prepared as above. A 5 ml droplet of a 0.1 mg/ml solution of P4 was applied to the grid, the excessblotted away gently with ®lter paper and a droplet ofstain (2 % (w/v) ammonium molybdate in water, pH 7.4)was added. The stain was left on the grid for 15 secondsbefore it was blotted dry. Pairs of images (nominaldefocus ÿ1 and ÿ3 mm ) were recorded at a nominalmagni®cation of 66,000� using the Philips CM200FEGunder conventional dose conditions.
Electron micrographs were recorded on Kodak SO163®lm. The low-dose cryo-electron micrographs weredeveloped in full strength D19 for 12 minutes. Images of

Symmetry Mismatch in the �6 Polymerase Complex 369
negatively stained samples were developed for ®ve min-utes in full strength D19.
Image processing
Micrographs were scanned on a Zeiss SCAI scanner ata step size of 14 mm and converted to a stamped MRCformat (Fuller et al., 1996) and then to SPIDER (http://www.wadsworth.org) (Frank et al., 1981, 1996) formatfor particle selection.
Isolated P4 complexes were selected from areas in themicrograph where they were embedded in an unsup-ported layer of stain over a hole in the support ®lm.These particles were placed into boxes of 72 by 72 pixels,the contrast reversed and the density values within �4SD of the mean were normalized.
Images of the recombinant polymerase complexeswere selected in boxes of 191 by 191 pixels, contrastreversed, normalized and output from SPIDER in MRCformat for icosahedral reconstruction (Baker & Cheng,1996; Crowther, 1971; Fuller, 1987; Fuller et al., 1996;Mancini et al., 1997). These images were interpolated to128 by 128 pixels for the model-based iterative recon-structions.
CTF determination
The CTF parameters of the images were determinedfrom the power spectrum. The power spectra from con-tiguous 256 by 256 pixel areas covering the image weresummed and rotationally averaged. A ®rst estimate forthe defocus was derived from the rotationally averagedpower spectrum using the position of ®rst zero and atable of ®rst CTF zeros calculated using the Cs (2 mm forthe Philips CM200FEG) (Erickson & Klug, 1971) and thefraction of amplitude contrast (0.1) typical for vitri®edsamples (Toyoshima & Unwin, 1988). This estimate ofthe defocus value was used as the starting value to opti-mize agreement of the predicted power spectrum (Fit) tothe observed power spectrum. The two curves wereplotted and the defocus (�f ) was adjusted until the bestagreement between the Fit function and the observedpower spectrum was achieved. The predicted powerspectrum was a function of the scattering vector (n), theelectron wavelength (l), the defocus (�f ), fraction ofamplitude contrast (Famp), and parameters describing thebaseline (c1,c2) and an exponential decay (c3,c4):
Fit�n;�f ;Famp; c1; c2; c3; c4� � c1eÿc2n
� �c3CTF�n;�f ;Famp�eÿc4n2 �2
where:
CTF�n� � �1ÿ Famp� sin�w�n�� � Famp cos�w�n��and:
x�n� � pln2��f ÿ 0:5Csl2n2�
The baseline was determined from the values of therotationally averaged power spectrum at the ®rst andthird zero of the CTF, based on the CTF for a given defo-cus. The decay represents the effects of limited temporalcoherence of the electron beam, ice thickness, movementand other factors which degrade the high-resolutioninformation in the image.
CTF correction
Once the parameters for the imaging conditions weredetermined, they were used to compensate for the CTFof each micrograph. A modi®ed Wiener ®lter:
W�n�i �CTF�n;�fi; Famp�
CTF�n;�fi; Famp�2 � d
was used for the correction. The parameter d in theWiener ®lter re¯ects the signal to noise and determinesthe weight of the low-amplitude information near theCTF nodes in the corrected image. Typically, the valuesof d were between 0.1 and 0.3 . We applied the correc-tion to the original images (Conway et al., 1997) ratherthan to the ®nal reconstruction (Ferlenghi et al., 1998;Mancini et al., 1997).
Three-dimensional reconstruction procedures
The icosahedral reconstruction procedures have beendescribed (Baker et al., 1999; Butcher et al., 1997; Fulleret al., 1996; Mancini et al., 1997). Model-based orientationsearches (Baker & Cheng, 1996) were used with a start-ing model based on the previous reconstructions(Butcher et al., 1997) and images which were CTF cor-rected as described above. Programs and documentationare available at http://www.EMBL-Heidelberg.DE/ExternalInfo/fuller/.
The program package SPIDER (Frank, 1973; Franket al., 1981, 1996) was used to calculate the three-dimen-sional reconstructions which did not employ icosahedralsymmetry. We developed a SPIDER batch procedurewhich uses an initial model to obtain preliminary orien-tations for the individual particle images. These orien-tations are subsequently re®ned in cycles whichconverge to a ®nal reconstruction. This procedure doesnot require symmetry, although symmetry can be incor-porated. Initial orientations are obtained using theSPIDER verb AP MD which requires a set of projectionsof the initial model along directions that are equallyspaced in orientation space, and the set of images. To ®llorientation space in the absence of symmetry, we usedan increment of 6 � resulting in a total set of 598 orien-tations. This set of 598 orientations represents one half oforientation space; the other half is mirror related andgenerated by AP MD. When symmetry is employed, thefraction of orientation space which needs to be ®lledwith projections is decreased. For example, only 96 pro-jections (separated by 6 �) were needed when 6-fold sym-metry was used for the P4 reconstruction, since onlyone-sixth of orientation space was unique. Prior to use inthe reconstruction procedure, images corresponding tothe same orientation were averaged and normalized toovercome the in¯uence of unevenly populated orien-tations (Boisset et al., 1998; de Haas et al., 1996a,b). Thisalso increases signal to noise and improves compu-tational ef®ciency. The Simultaneous Iterative Recon-struction Technique (SIRT) originally developed byGilbert (1972a,b,c) was used to calculate the reconstruc-tion. It allows the application of constraints such as sym-metry at each cycle of the reconstruction process. Theprogress of the SIRT reconstruction of the P1247 was fol-lowed (Figure 6) by calculating the volume in the pos-ition of the satellites enclosed by a contour at 0.5s. Theeffect of inhomogeneity in the sample was examined byusing different models (for example, the one-layered hex-amer and two-layered dodecamer of P4) and separating

370 Symmetry Mismatch in the �6 Polymerase Complex
the population of input images on the basis of relativecross-correlation with the models.
The use of the SIRT reconstruction method in thiswork is simply a convenient approach to generatingasymmetric models for orientation (F.d.H. et al., unpub-lished results). Tests revealed that actual reconstructionis very similar whether the SIRT algorithm is used or ageneral weighted back projection algorithm (Frank et al.,1996) is applied using the same models. Fourier shellcorrelation (FSC) between reconstructions of indepen-dent data sets (van Heel, 1987) was always used toestablish the reproducibility of the reconstructions and tocontrol for the effects of model bias on the reconstruc-tions.
Averaging
Averages in two dimensions were calculated using areference-free alignment procedure as implemented inSPIDER and calculating the average with the SPIDERverb AD. Alignment in three dimensions utilized theSPIDER verb OR 3Q.
Visualization
Some reconstructions are presented as radially depth-cued surface representations (Spencer et al., 1997) gener-ated using AVS modules written by S. Fuller (Sheehanet al., 1996). The modules are available upon request. Thestereo views are generated by rotations of �6 � aroundthe vertical. Surface representations are presented ata contour level which corresponds to one standarddeviation above background. This is a relatively conserv-ative threshold which corresponds to a mass ofP1120P212P460P760 and is unlikely to miss any signi®cantfeature of the structure (Conway et al., 1996). The bulk ofthe polymerase complex is P1; loss of a fraction of pro-tein from the complex would lower the effective contourlevel and hence reveal lower density features.
Acknowledgments
Dr Jarmo Juuti (University of Helsinki) is acknowl-edged for kindly providing the puri®ed protein P4 andMs Marja-Leena PeraÈ laÈ (University of Helsinki) for herskilful technical assistance. The authors also gratefullyacknowledge the contributions of Ralph Heinkel (EMBL)for computational support, Brent Gowen (presently atImperial College, London) for some of the electronmicroscopy and Mairi Clarke (EMBL) for help with thebiochemical preparations. We thank Dr Sarah Butcher(University of Helsinki) and Dr Dennis Thomas (EMBL)and Doryen Bubeck (EMBL) for insightful discussionsand critical readings of the manuscript. We are gratefulto Dr Roman Tuma (University of Alabama at Birming-ham) for discussions of his ultracentrifugation results.This work was supported by grants from the FinnishAcademy of Sciences (nos 37725 and 30526 to D.H.B.)and European Union (BIO4-CT97-2364 to D.H.B. andS.D.F). A.O.P. was supported by a fellowship from Hel-sinki Graduate School in Biotechnology and MolecularBiology (University of Helsinki) and FEMS travel fellow-ship for visiting EMBL.
References
Adrian, M., Dubochet, J., Lepault, J. & McDowall, A. W.(1984). Cryo-electron microscopy of viruses. Nature,308, 32-36.
Baker, T. S. & Cheng, R. H. (1996). A model-basedapproach for determining orientations of biologicalmacromolecules imaged by cryoelectronmicroscopy. J. Struct. Biol. 116, 120-130.
Baker, T. S., Olson, N. H. & Fuller, S. D. (1999). Addingthe third dimension to virus life cycles: three-dimensional reconstruction of icosahedral virusesfrom cryo-electron micrographs. Microbiol. Mol. Biol.Rev. 63, In the press.
Bamford, D. H., Romantschuk, M. & Somerharju, P. J.(1987). Membrane fusion in prokaryotes: bacterio-phage phi6 membrane fuses with the Pseudomonassyringae outer membrane. EMBO J. 6, 1467-1473.
Bazinet, C. & King, J. (1985). The DNA translocatingvertex of dsDNA bacteriophage. Annu. Rev. Micro-biol. 39, 109-129.
Boisset, N., Penczek, P. A., Taveau, J.-C., You, V., deHaas, F. & Lamy, J. (1998). Overabundant single-particle electron microscope views induce a three-dimensional reconstruction artefact. Ultramicroscopy,74, 201-207.
Butcher, S., Dokland, T., Ojala, P., Bamford, D. & Fuller,S. (1997). Intermediates in the assembly pathway ofthe double stranded RNA virus f-6. EMBO J. 16,4477-4487.
Caldentey, J. & Bamford, D. H. (1992). The lytic enzymeof the Pseudomonas phage phi 6. Puri®cation andbiochemical characterization. Biochim. Biophys. Acta,1159, 44-50.
Caspar, D. L. D. (1964). Design and assembly of orga-nized biological structures. In Molecular Architecturein Cell Physiology (Hayashi, T. & Szent-Gyorgyi,A. G., eds), pp. 191-207, Prentice Hall, New Jersey.
Caspar, D. L. D. (1979). Dynamics of virus assemblies.J. Supramol. Struct. S3, 81.
Caspar, D. L. D. (1980). Movement and self-control inprotein assemblies. Biophys. J. 32, 103-138.
Caspar, D. L. D. & Klug, A. (1962). Physical principlesin the construction of regular viruses. Cold SpringHarbor Symp. Quant. Biol. 27, 1-24.
Conway, J. F., Trus, B. L., Booy, F. P., Newcomb, W. W.,Brown, J. C. & Steven, A. C. (1996). Visualization ofthree-dimensional density maps reconstructed fromcryoelectron micrographs of viral capsids. J. Struct.Biol. 116, 200-208.
Conway, J. F., Cheng, N., Zlotnick, A., Wing®eld, P. T.,Stahl, S. J. & Steven, A. C. (1997). Visualization of a4-helix bundle in the hepatitis B virus capsid bycryo-electron microscopy. Nature, 386, 91-94.
Crowther, R. A. (1971). Procedures for three-dimensionalreconstruction of spherical viruses by Fourier syn-thesis from electron micrographs. Phil. Trans. Roy.Soc. ser. B, 261, 221-230.
de Haas, F., Zal, F., Lallier, F. H., Toulmond, A. &Lamy, J. N. (1996a). Three-dimensional reconstruc-tion of the hexagonal bilayer hemoglobin of hydro-thermal vent tube worm Riftia pachyptila by cryo-electron microscopy. Proteins: Struct. Funct. Genet.26, 241-256.
de Haas, F., Zal, F., You, V., Lallier, F., Toulmond, A. &Lamy, J. N. (1996b). Three-dimensional reconstruc-tion by cryo-electron microscopy of the giant hemo-globin of the polychaete worm Alvenella pompejana.J. Mol. Biol. 264, 111-120.

Symmetry Mismatch in the �6 Polymerase Complex 371
Dokland, T., Lindqvist, B. H. & Fuller, S. D. (1992).Image reconstruction from cryo-electron micro-graphs reveals the morphopoetic mechanism in theP2-P4 bacteriophage system. EMBO J. 11, 839-846.
Dokland, T., Isaksen, M. L., Fuller, S. D. & Lindqvist,B. H. (1993). Capsid localization of the bacterio-phage-P4 Psu protein. Virology, 194, 682-687.
Dubochet, J. & Groom, M. (1982). The mounting ofmacromolecules for electron microscopy with par-ticular reference to surface phenomena and thetreatment of support ®lms by glow discharge.Advan. Optical Electron Microsc. 8, 107-135.
Erickson, H. P. & Klug, A. (1971). Measurement andcompensation of defocusing and aberrations byfourier processing of electron micrographs. Proc.Trans. Roy. Soc. London, 261, 105-118.
Ferlenghi, I., Gowen, B., Haas, F. D., Mancini, E. J.,Garoff, H., SjoÈberg, M. & Fuller, S. D. (1998). The®rst step: maturation of the Semliki Forest virusspike occurs through a dramatic localized confor-mational change. J. Mol. Biol. 283, 71-81.
Frank, J. (1973). Computer processing of electron micro-graphs. In Advanced Techniques in Biological ElectronMicroscopy (Koehler, J. K., ed.), vol. 1, pp. 215-274,Springer-Verlag, Berlin.
Frank, J., Shimkin, B. & Dowse, H. (1981). SPIDER - amodular software system for electron image proces-sing. Ultramicroscopy, 6, 343-358.
Frank, J., Radermacher, M., Penczek, P., Zhu, J., Li, Y.,Ladjadj, M. & Leith, A. (1996). SPIDER and WEB:processing and visualization of images in 3D elec-tron microscopy and related ®elds. J. Struct. Biol.116, 190-199.
Frilander, M. & Bamford, D. H. (1995). In vitro packa-ging of the single-stranded RNA genomic precur-sors of the segmented double-stranded RNAbacteriophage phi 6: the three segments modulateeach other's packaging ef®ciency. J. Mol. Biol. 246,418-428.
Frilander, M., Poranen, M. & Bamford, D. H. (1995). Thelarge genome segment of dsRNA bacteriophage phi6 is the key regulator in the in vitro minus and plusstrand synthesis. RNA, 1, 510-518.
Fukami, A. & Adachi, K. (1965). A new method of prep-aration of a self-perforated micro plastic grid andits application. J. Electron Microsc. 14, 112-118.
Fuller, S. D. (1987). The T � 4 envelope of Sindbis virusis organized by complementary interactions with aT � 3 icosahedral capsid. Cell, 48, 923-934.
Fuller, S. D., Butcher, S. J., Cheng, R. H. & Baker, T. S.(1996). Three-dimensional reconstruction of icosahe-dral particles: The uncommon line. J. Struct. Biol.116, 48-55.
Gilbert, P. (1972a). Iterative methods for the three-dimensional reconstruction of an object from projec-tions. J. Theoret. Biol. 36, 105-117.
Gilbert, P. F. C. (1972b). An iterative method for three-dimensional reconstruction from electron micro-graphs. Fifth European Regional Conference of ElectronMicroscopy, 602-603.
Gilbert, P. F. C. (1972c). The reconstruction of a three-dimensional structure from projections and itsapplication to electron microscopy. II. Directmethods. Proc. Roy. Soc. ser. B, 182, 89-102.
Gottlieb, P., Strassman, J., Bamford, D. & Mindich, L.(1988). Production of a polyhedral particle in Escher-ichia coli from a cDNA copy of the large genomesegment of f6. J. Virol. 62, 181-187.
Gottlieb, P., Strassman, J., Quao, X., Frucht, A. &Mindich, L. (1990). In vitro replication, packaging,and transciption of the segmented, double-strandedRNA genome of bacteriophage f6: studies withprocapsids assembled from plasmid-encoded pro-teins. J. Bacteriol. 172, 5774-5782.
Gottlieb, P., Strassman, J., Frucht, A., Quao, X. &Mindich, L. (1991). In vitro packaging of the bac-teriophage f6 ssRNA genomic precusors. Virology,181, 589-594.
Grimes, J. M., Bourroughs, J. N., Gouet, P., Diprose,J. M., Malby, R., ZieÂntara, S., Mertens, P. P. C. &Stuart, D. I. (1998). The atomic structure of the blue-tongue virus core. Nature, 395, 470-478.
Guo, P., Zhang, C., Chen, C., Garver, K. & Trottier, M.(1998). Inter-RNA interaction of phage f29 pRNAto form a hexameric complex for viral DNA tran-scription. Mol. Cell, 2, 149-155.
Hendrix, R. W. (1978). Symmetry mismatch and DNApackaging in large bacteriophages. Proc. Natl Acad.Sci. USA, 75, 4779-4783.
Hendrix, R. W. (1998). Bacteriophage DNA packaging:RNA gears in a DNA transport machine. Cell, 94,147-150.
Hewat, E. A., Booth, T. F., Loudon, P. T. & Roy, P.(1992a). Three-dimensional reconstruction of baculo-virus expressed bluetongue virus core-like particlesby cryo-electron microscopy. Virology, 189, 10-20.
Hewat, E. A., Booth, T. F. & Roy, P. (1992b). Structureof bluetongue virus particles by cryoelectronmicroscopy. J. Struct. Biol. 109, 61-69.
Juuti, J. T. & Bamford, D. H. (1995). RNA binding,packaging and polymerase activities of the differentincomplete polymerase complex particles of dsRNAbacteriophage ù6. J. Mol. Biol. 249, 545-554.
Juuti, J. T. & Bamford, D. H. (1997). Protein P7 of phagephi6 RNA polymerase complex, acquiring of RNApackaging activity by in vitro assembly of the puri-®ed protein onto de®cient particles. J. Mol. Biol. 266,891-900.
Juuti, J., Bamford, D. H., Tuma, R. & Thomas, G. J. J.(1998). Structure and NTPase activity of the RNAtranslocating protein (P4) of bacteriophage phi-6.J. Mol. Biol. 279, 347-359.
Kenney, J. M., Hantula, J., Fuller, S. D., Mindich, L.,Ojala, P. M. & Bamford, D. H. (1992). Bacteriophagef-6 envelope elucidated by chemical crosslinking,immunodetection and cryo-electron microscopy.Virology, 190, 635-644.
Li, T., Bamford, D. H., Bamford, J. K. H. & Thomas,G. J. J. (1993). Structural studies of the envelopeddsRNA bacteriophage phi 6 of Pseudomonas syringaeby Raman spectroscopy. I. The virion and its mem-brane envelope. J. Mol. Biol. 230, 461-472.
Mancini, E. J., de Haas, F. & Fuller, S. D. (1997). Highresolution icosahedral reconstruction: ful®lling thepromise of cryo-electron microscopy. Structure, 5,741-750.
Mindich, L. (1995). Heterologous recombination in thesegmented dsRNA genome of bacteriophage phi6.Semin. Virol. 6, 75-83.
Mindich, L. (1999). Precise packaging of the three geno-mic segments of the double-stranded-RNA bacterio-phage f-6. Microbiol. Mol. Biol. Rev. 63, 149-160.
Mindich, L. & Lehman, J. F. (1979). Cell wall lysin as acomponent of the bacteriophage ù6 virion. J. Virol.30, 489-496.
Moody, M. F. (1965). The shape of the T-even bacterio-phage head. Virology, 26, 567-576.

372 Symmetry Mismatch in the �6 Polymerase Complex
Ojala, P. M. & Bamford, D. H. (1995). In vitro transcrip-tion of the double-stranded RNA bacteriophage phi6 is in¯uenced by purine NTPs and calcium. Virol-ogy, 207, 400-408.
Ojala, P. M., Romantschuk, M. & Bamford, D. H. (1990).Puri®ed phi6 nucleocapsids are capable of pro-ductive infection of host cells with partially dis-rupted outer membranes. Virology, 178, 364-372.
Olkkonen, V. M. & Bamford, D. H. (1987). The nucleo-capsid of the lipid containing double stranded bac-teriophage f6 contains a single protein species.J. Virol. 61, 2362-2367.
Olkkonen, V. M. & Bamford, D. H. (1989). Quantitationof the adsorption and penetration stages of bacterio-phage phi6 infection. Virology, 171, 229-238.
Olkkonen, V. M., Gottlieb, P., Strassman, J., Qiao, X. Y.,Bamford, D. H. & Mindich, L. (1990). In vitro assem-bly of infectious nucleocapsids of bacteriophage phi6: formation of a recombinant double-strandedRNA virus. Proc. Natl Acad. Sci. USA, 87, 9173-9177.
Olkkonen, V. M., Ojala, P. M. & Bamford, D. H. (1991).Generation of infectious nucleocapsids by in vitroassembly of the shell protein on to the polymerasecomplex of the dsRNA bacteriophage phi 6. J. Mol.Biol. 218, 569-581.
Onodera, S., Qiao, X., Qiao, J. & Mindich, L. (1995).Acquisition of a fourth genomic segment in bac-teriophage phi6, a a bacteriophage with a genomeof three segments of dsRNA. Virology, 212, 204-212.
Onodera, S., Qiao, X., Qiao, J. & Mindich, L. (1998).Directed changes in the number of double-strandedRNA segments in bacteriophage phi6. Proc. NatlAcad. Sci. USA, 95, 3920-3924.
Paatero, A. O., Syvaoja, J. E. & Bamford, D. H. (1995).Double-stranded RNA bacteriophage f6 protein P4is an unspeci®c nucleoside triphosphatase activatedby calcium ions. J. Virol. 69, 6729-6734.
Paatero, A. O., Mindich, L. & Bamford, D. H. (1998).Mutational analysis of the role of the NTPase P4 inthe assembly of the RNA polymerase complex ofbacteriophage f6. J. Virol. 72, 10058-10065.
Palmer, E. L. & Martin, M. L. (1982). Reoviridae. InAtlas of Mammalian Viruses, pp. 23-33, CRC Press,Inc., Boca Raton.
Prasad, B. V. V. & Chiu, W. (1994). Structure of rota-virus. In Rotaviruses (Ramig, R. F., ed.), pp. 9-29,Springer-Verlag, Berlin, Heidelberg.
Prasad, B. V. V., Wang, G. J., Clerx, J. P. M. & Chiu, W.(1988). Three-dimensional structure of rotavirus.J. Mol. Biol. 199, 269-275.
Prasad, B. V., Yamaguchi, S. & Roy, S. (1992). Three-dimensional structure of single-shelled bluetonguevirus. J. Virol. 66, 2135-2142.
Qiao, X., Qiao, J. & Mindich, L. (1997). Stoichiometricpackaging of the three genome segments of double-stranded RNA bacteriophage phi6. Proc. Natl Acad.Sci. USA, 94, 4074-4079.
Romantschuk, M. & Bamford, D. H. (1985). Function ofpili in bacteriophage phi6 penetration. J. Gen. Virol.66, 2461-2469.
Romantschuk, M., Olkkonen, V. M. & Bamford, D. H.(1988). The nucleocapsid of bacteriophage phi6penetrates the host cytoplasmic membrane. EMBOJ. 7, 1821-1829.
Sheehan, B., Fuller, S. D., Pique, M. E. & Yeager, M.(1996). AVS software for visualization in molecularmicroscopy. J. Struct. Biol. 116, 99-105.
Spencer, S. M., Sgro, J.-Y., Dryden, K. A., Baker, T. S. &Nibert, M. L. (1997). IRIS explorer software forradial-depth cueing reovirus particles and othermacromolecular structures determined by cryoelec-tron microscopy and image reconstruction. J. Struct.Biol. 120, 11-21.
Stewart, P. L., Burnett, R. M., Cyrklaff, M. & Fuller, S. D.(1991). Image reconstruction reveals the complexmolecular organization of adenovirus. Cell, 67, 145-154.
Toyoshima, C. & Unwin, N. (1988). Contrast transfer forfrozen-hydrated specimens: determination frompairs of defocused images. Ultramicroscopy, 25, 279-292.
Tyler, K. L. & Fields, B. N. (1990). Reovirus. In Virology.(Fields, B. N., ed.), pp. 1307-2989, Raven Press,New York.
Valpuesta, J. M., Fernandez, J. J., Carazo, J. M. &Carrascosa, J. L. (1999). The three-dimensionalstructure of a DNA translocating machine at 10 AÊ
resolution. Struct. Fold. Des. 7, 289-296.van Heel, M. (1987). Similarity measures between
images. Ultramicroscopy, 21, 95-100.van Heel, M., Orlova, B. V., Dube, P. & Tavares, P.
(1996). Intrinsic versus imposed curvature in cycli-cal oligomers: the portal protein of bacteriophageSPP1. EMBO J. 15, 4785-4788.
Yeager, M., Dryden, K. A., Olson, N. H., Greenberg,H. B. & Baker, T. S. (1990). Three-dimensional struc-ture of rhesus rotavirus by cryoelectron microscopyand image reconstruction. J. Cell Biol. 110, 2133-2144.
Zhang, F., Lemieus, S., Wu, X., St.-Armaud, D., Mc.,Murray C. T., Major, F. & Anderson, D. (1998).Function of the hexameric RNA in the packaging ofbacteriophage f29 DNA in vitro. Mol. Cell, 2, 141-147.
Edited by M. F. Moody
(Received 1 July 1999; received in revised form 1 October 1999; accepted 5 October 1999)