a molecular perspective on ecological differentiation and biogeography of cyclotrichiid ciliates
TRANSCRIPT

A Molecular Perspective on Ecological Differentiation and Biogeography ofCyclotrichiid Ciliates
DAVID BASS,a,b NICK BROWN,c JACKIE MACKENZIE-DODDS,b PATRICIA DYAL,b SANDRA A. NIERZWICKI-BAUER,d
ALEXEY A. VEPRITSKIYd and THOMAS A. RICHARDSa,b,1
aDepartment of Zoology, The University of Oxford, South Parks Road, Oxford OX1 3PS, United Kingdom, andbDepartment of Zoology, The Natural History Museum, Cromwell Road, London SW7 5BD, United Kingdom, and
cDepartment of Plant Sciences, The University of Oxford, South Parks Road, Oxford OX1 3RB, United Kingdom, anddDepartment of Biology, Darrin Fresh Water Institute and Rensselaer Polytechnic Institute, 5060 Lakeshore Drive, Bolton Landing,
New York 12814, USA
ABSTRACT. Cyclotrichiids are of ecological and evolutionary interest by virtue of their importance in red tide formation, their highlydivergent small subunit (SSU) ribosomal RNA (rRNA) genes, kleptoplastidy, and utility as indicators of eutrophication. However, onlyseven strains have had their SSU rRNA genes sequenced and their environmental diversity and distribution are largely unknown. Weprobed 67 globally dispersed freshwater column/sediment and soil DNA samples (eDNAs) and constructed 24 environmental genelibraries using polymerase chain reaction primers specific to an uncharacterised cyclotrichiid subgroup. We reveal a novel, globallyubiquitous freshwater clade comprising 25 genetically distinct SSU ribosomal DNA (rDNA) sequences (SSU-types). Some identicalSSU-types were detected at globally widely distributed sites. The SSU-types form four distinct phylogenetic clusters according to marineor non-marine provenance, suggesting at least one major marine–freshwater evolutionary transition within the cyclotrichiids. We used thesame primers to sample intensively 18 sampling points in 13 closely situated lakes, each characterised by 14 environmental variables, andshowed that molecular detection or non-detection of cyclotrichiids was most significantly influenced by levels of total phosphorus, dis-solved organic carbon, and chlorophyll a. Within the subset of lakes in which cyclotrichiids were detected, closely related SSU-typesdiffered in their ecological preferences to pH, total phosphorus, and sample depth.
Key Words. Ecological differentiation, Mesodinium, Myrionecta, small subunit (SSU) ribosomal DNA (rDNA).
THE ciliate order Cyclotrichiida (Jankowski 1980) includesthe heterotrophic genus Mesodinium and the mixotrophic
genus Myrionecta, and occurs in diverse and globally dispersedaquatic environments. Other genera in the order—Askenasia,Rhabdoaskenasia, and possibly Pelagovasicola and Liliimorpha(supporting information Table S1)—also vary in nutritional modeand are reported to differ significantly in terms of distribution andecological preferences (Foissner, Berger, and Schaumburg 1999;Krainer and Foissner 1990; Struder-Kypke et al. 2006). My-rionecta rubra (previously Mesodinium rubrum) and Mesodiniumpulex are currently the only two species within the order repre-sented in GenBank by nuclear genes suitable for phylogeneticanalyses. The Mesodinium pulex Morphospecies is common infreshwater and marine pelagial waters and mud. It is rare or absentin acidic/oligotrophic lakes, steadily increasing in frequency withincreasing eutrophication, and because of this has been reported asone of the best ciliate indicator species for assessing the trophicstatus of lakes in Florida (Beaver and Crisman 1989). Another,smaller, Mesodinium species, Mesodinium acarus, is also found inboth marine and non-marine habitats, although less frequently inthe latter than M. pulex, whereas Mesodinium velox is exclusivelymarine and Mesodinium fimbriatum exclusively non-marine (Ta-ble S1). Myrionecta rubra is found in marine and brackish habitatsonly and can form massive non-toxic red tide blooms, which itachieves partly by virtue of its sequestration of cryptophyceanorganelles (Foissner et al. 1999; Johnson et al. 2004).
Sequence data for cyclotrichiids are sparse: very few have beensequenced from cultures or identified cells, and their small subunit(SSU) ribosomal DNA (rDNA) sequences are relatively rarelydetected in eukaryote-wide environmental gene libraries, proba-bly because of their divergent nature (Johnson et al. 2004). How-
ever, cyclotrichiid-like SSU rDNA sequences have been detectedin marine (e.g. Lopez-Garcıa et al. 2001; Stoeck and Epstein2003) and freshwater (e.g. Lake George, Adirondack Park, USAby Richards et al. 2005) libraries, and were initially regarded asnovel deep branching eukaryotes due to their deep and long-branch formed in SSU rDNA phylogenies (Lopez-Garcıa et al.2001; Stoeck and Epstein 2003).
The purpose of this study was to use molecular techniques toinvestigate cyclotrichiid diversity and distribution with a particu-lar emphasis on the novel freshwater lineage revealed by Richardset al. (2005). This lineage was initially referred to as ‘‘LG-D’’ inRichards et al. (2005) and retains that label here for ease of cross-referencing. Because LG-D has thus far only been detected inLake George we probed a widely dispersed set of environmentalDNA samples (eDNAs) to determine its global distribution. Intotal we probed 67 eDNAs samples from freshwater column, sed-iment, and soil habitats from the five global biogeographical re-gions using a highly specific SSU primer set targeting LG-D.Thirty-seven of the eDNAs yielded polymerase chain reaction(PCR) bands, which were used to construct 24 environmental genelibraries. By sequencing clones from these environmental genelibraries we begin to describe the global occurrence of freshwatercyclotrichiids.
Because freshwater cyclotrichiids are morphologically and eco-logically diverse (Table S1), we hypothesised that the cluster ofsequence types we detected using LG-D-specific primers com-prised an ecologically heterogeneous assemblage, perhaps en-compassing several species and genera. To learn more about theenvironmental parameters determining their distribution, andthe nature of niche differences among these closely related strainswe analysed LG-D lineage diversity in the context of multivariateenvironmental data from the Adirondack Effects Assessment Pro-gram at the Darrin Fresh Water Institute, New York, USA. Twelveof our libraries were constructed from different lakes in the A-dirondack Park, each associated with 14 physical/chemical vari-ables measured at the same time as eDNA sampling. We usedcanonical correspondence analyses to determine the environmen-tal parameters within which cyclotrichiids occurred in the lakes,and to gain insight into which variables were the most powerful
1Current address: Centre for Eukaryotic Evolutionary Microbiology,School of Biosciences, University of Exeter, Devon EX4 4QD, UnitedKingdom.
Corresponding Author: T. Richards, School of Biosciences, Univer-sity of Exeter, Geoffrey Pope Building, Stocker Road, Exeter, EX44QD, United Kingdom—Telephone number: 144 0 1392 263756;e-mail: [email protected]
559
J. Eukaryot. Microbiol., 56(6), 2009 pp. 559–567r 2009 The Author(s)Journal compilation r 2009 by the International Society of ProtistologistsDOI: 10.1111/j.1550-7408.2009.00434.x

predictors of the relative distribution of LG-D SSU-types. Byanalysing our new sequences with related ones from GenBank(Table S2) we identified four main phylogenetically distinctgroups of cyclotrichiid-like SSU-types—three apparently exclu-sively marine and one exclusively freshwater. Our results showthat cyclotrichiid taxonomy requires substantial revision in thelight of detailed molecular data, and that ecological partitioningamong strains is more finely divided than could be known previ-ously. We also discuss the contribution of our results to the un-derstanding of protist global biogeography.
MATERIALS AND METHODS
DNA extraction, PCR, and environmental gene library con-struction. Environmental DNA was extracted from 67 freshwatercolumn, freshwater sediment, and soil samples collected fromPalaearctic, Australasia, and Neotropic ecoregions (Table 1, Fig.1), as described previously (Bass and Cavalier-Smith 2004). Allnon-Adirondack eDNA samples had been used previously to am-plify SSU ribosomal RNA (rRNA) gene sequences using a varietyof PCR primer combinations (Bass and Cavalier-Smith 2004;Bass et al. 2007). For all Adirondack eDNA samples we per-formed a series of control experiments to confirm the PCR resultsas discussed below.
Eighteen water samples from 13 lakes in the Adirondack Park(Nearctic) were collected in September 2002 as part of the long-term abiotic/biotic surveys of Adirondack Park freshwater envi-ronments (http://www.rpi.edu/dept/DFWI) (Table S3). Water
samples from lakes that exhibited thermal stratification were col-lected from the well-mixed epilimnion and the hypolimnion (at1 m above the lake bottom). Epilimnetic samples and samplesfrom lakes that were not stratified at the time of sampling werecollected using a wide-diameter hose to provide a depth-inte-grated composite sample. Hypolimnetic samples were collected at1 m above the lake bottom using a van Dorn sampler. Thirteen
Table 1. Sampling and provenance of SSU-types.
Biogeographicalregion (numbers referto Fig. 1 ecoregions)
Provenance Number eDNAsscreened/
number successful/number cloned
Samplename
Habitat SSU-types SSU-typesuniqueto locality
Australasia (4) New Zealand 13/11/5 NZ119 Beech forest streamsediment, South Island
A, E, F, Y 0
TA-NZ15/16/12 Hamilton Lake watercolumn, North Island
K, P, R
TB-NZ103/101 Stream near Te TotoGorge, North Island
Y
E. Palearctic (2) Sakhalin Island 5/4/3 SKL-RX f/w column B, D, H, X, Z, Y 2 (I, D)SKL-Y f/w column A, C, D, I, ESKL-P f/w column C, E, F, H
W. Palearctic (5) United Kingdomand Germany
11/6/4 BRI-74 f/w (from submergedlily stem)
F, X, Z, Y 0
BRI-69 f/w column (0.5 m deep) EGER-F2 Soil YMARB-1 Soil Y
Neotropic (1) Panama 10/3/3 P40 f/w sediment K, L, V 0P25 f/w sediment A, B, FP39 f/w sediment C, E, F
Nearctic (3) United States(Adirondacks)
20/12/12 BM1 f/w columna J, K, M, N, V 8, (W, U, T,Q, O, N, M,J, G)
BT2 f/w columna O, P, W, VIL8 f/w columna K, Q, VIL9 f/w columna M, N, K, VLP10 f/w columna U, V, K, QNL11 f/w columna K, T, VNL12 f/w columna J, K, VSL13 f/w columna K, VSL14 f/w columna W, G, JWL15 f/w columna K, VWL16 f/w columna K, L, O, U, VWL17 f/w columna K, R, V
aSampled with 5mm filter selection.SSU, small subunit.
Fig. 1. Map of the world showing six terrestrial ecoregions. Num-bered circles mark sampling localities for the cyclotrichiid ciliates andcorrespond to phylogenetic labels shown in Fig. 2.
560 J. EUKARYOT. MICROBIOL., 56, NO. 6, NOVEMBER–DECEMBER 2009

physical and chemical properties were determined on replicatewater samples. Secchi depth was also estimated in lakes at thetime of sampling. Water samples were processed, preserved(where appropriate), and analysed for variables listed in Table S3.
All analyses were conducted at the Darrin Fresh Water Insti-tute’s New York State Environmental Laboratory Approval Pro-gram-certified laboratories following appropriate analytical andquality assurance/quality control procedures approved by the USEnvironmental Protection Agency (USEPA 1983) The specificanalytical methods used are as follows: pH (Electrometric USEPAMethod 150.1); Acid Neutralising Capacity (Gran Titration USE-PA Method 310.1); Dissolved Organic Carbon (Infrared IR Spec-troscopy USEPA Method 415.2); Na, K, Fe, Mg (AtomicAbsorption USEPA Method 200.0); total Al (Inductively CoupledPlasma Emission); NH4 (Flow Injection Analysis-Lachat); NO3,SO4 (Ion Chromatography USEPA Method 300.0); total P (Col-orimetric USEPA Method 365.2); and chlorophyll a (chl a) [Flu-orimetric-Turner Designs, Sunnyvale, CA (Turner 1985)].Environmental Laboratory Approval certification is required forall laboratories performing environmental analyses on samplesoriginating from New York State to ensure the accuracy andreliability of these analyses. For molecular studies one-litre sam-ples were pre-filtered through a nylon mesh, filtered through 5 mmcellulose membrane filters (Millipore, Billerica, MA), and theremaining plankton collected on 0.2 mm cellulose membranefilters (Millipore). DNAs were extracted using The FastPreps
System (Q-Biogene, Irvine, CA).For PCR amplifications we designed a pair of highly selective
LG-D group-specific primers based on environmental SSU rRNAgene sequences from Lake George (Richards et al. 2005): LGD-698—50-GCTTAGGTTTCTCGTCTTAGGA-30 and LGD-1351—50-TAAGACGACGATTGCTGATTT-30 (primer numbers referto corresponding positions in GenBank sequence AY919681).Primers were targeted to sample a � 650-nt region suggestedto include the longest hypervariable region of the cyclotrichiidSSU rRNA gene when compared with SSU genes from differenttaxa (Johnson et al. 2004). The cyclotrichiid V4 region has un-dergone a number of deletions of hypervariable regions (Johnsonet al. 2004); consequently our PCR sampling focused on samplingthe two cyclotrichiid variable regions between the V4 and V7 re-gions of the SSU gene.
All 67 eDNA samples were subjected to LG-D-specific PCR. Thereaction and cycling conditions were as follows: 30 ng templateDNA, 1 � TaqExtender PCR buffer (Stratagene, Santa Clara, CA),3.5 mM MgCl2, 200mM of each dNTP (Fisher Scientific, Pittsburgh,PA), 0.2mM of each primer, 0.6mg of T4 gene 32 Protein (Ambion,Austin, TX), 1.0 U of TaqExtender (Stratagene), and 1.0 U of TaqDNA polymerase (New England Biolabs, Ipswich, MA) were com-bined in a total volume of 25ml and incubated for 5 min at 94 1C,followed by 35 cycles of 30 s at 94 1C, 45 s at 57 1C, 2 min at 72 1Cwith an additional 5 min extension at 72 1C.
All Adirondack lake eDNA samples were verified first to beSSU-amplifiable using general SSU primers 3Fphp and 1749Rphp(Richards et al. 2005). In all cases the general SSU primersproduced a PCR product of the correct size ( � 1,700 bp). Usingthe LG-D primers we then performed five separate PCR amplifi-cations on each Adirondack DNA sample. Six of the 18 sampleswere LG-D PCR negative. To confirm negative amplifications wethen performed nested PCR by subjecting the general SSU PCRproducts obtained with 3Fphp and 1749Rphp (Richards et al.2005) to a second round of amplification using the LG-D prim-ers. The distribution of LG-D PCR results from the Adirondackenvironmental DNA samples did not differ between direct andnested amplifications.
Products from three independent PCR amplifications werepooled, gel-extracted, purified with QIAquick spin columns
(Qiagen, Valencia, CA), and cloned using TOPO TA CloningKit (Invitrogen, Carlsbad, CA). Sixteen clones were picked fromeach Adirondack library for sequencing. Plasmid DNA was pre-pared using QIAprep spin columns (Qiagen).
Sequence analysis and phylogeny. Large-scale molecularscreening projects are susceptible to methodological errors thatoverestimate molecular diversity and/or underestimate the overlapbetween different environments. These include nucleotide mis-incorporation, chimaera formation, and/or differences in the se-quence among different copies of the SSU rRNA gene within asingle nucleus. To minimise these potential sources of artefact/error, we did the following: (1) used multiple individual PCRproducts from the same eDNA in our cloning reactions; (2) con-firmed all unique clones by double-stranded sequencing; (3) tar-geted a short but hypervariable fragment ( � 650 bp) to minimisechimaera formation [assisted by prolonging the PCR extensionstep time (Judo, Wedel, and Wilson 1998)] and to enable double-stranded sequencing with a single primer set; and (4) checked thealignment for patterns of sequence variation that could bethe products of chimaeras, both by visual analysis of signaturesequences and by constructing trees from the first two-fifths andthe last two-fifths of the cyclotrichiid alignment. These latter twoanalyses showed a different topology among the terminal nodes ofthe phylogeny, however, neither alternative topology was sup-ported by a bootstrap value 450% and these topological differ-ences were restricted to nodes that were weakly supported in thefinal analyses (Fig. 2). No evidence of recombination in the frag-ment analysed was detected by either method.
We only designated sequence types as unique SSU-types if theyshared the same single nucleotide substitutions across more thanone library. Because libraries are independent samples, such se-quences cannot come from the same nucleus. Only SSU-typesthus defined and singleton sequences that are o99% similar toany other across the sequences analysed (647 bp) were included inthe phylogenetic analyses shown in Fig. 2. Thus singleton se-quences differ from each other at seven or more positions, muchmore than would be expected by PCR and sequencing error (Basset al. 2007), and are therefore very likely to derive from differentlineages. In other protist groups studied in detail, even singledifferences in SSU sequence between strains can represent differ-ent species (e.g. Chantangsi and Lynn 2008; Howe et al. 2009;Lynn and Struder-Kypke 2006), and species boundaries in a widerange of biological species have been shown to coincide withdifferences in particular substitutions in ITS2 rDNA (i.e. betweenstrains with potentially identical SSU rDNA—Coleman 2007).After the clustering of sequences according to shared substitutionsamong libraries, only one singleton SSU-type (I) remainedthat was not present in more than one environmental gene library(Fig. 2). This clustering process reduced the dataset of 235 se-quences to 25 SSU-types (A–Z, excluding S, Fig. 2). Examples ofeach SSU-type are available in GenBank (accession numbers aredisplayed on Fig. 2).
BLASTn searches were used to sample all available cyclotri-chiid-like sequences available in GenBank (08/08). The great ma-jority of these sequences were from marine environmental genelibraries, 29 of which were excluded because the sequences werenot complete across the section of SSU rRNA gene sequencesampled using our LG-D primer set (Table S2). The remainingsequences were aligned manually and the 50- and 30- ends notwithin the LG-D primer sampling range were removed. An align-ment data matrix of 69 sequences and 647 nucleotide characterswas analysed using the program PHYML (Guindon and Gascuel2003) with the following model parameters: GTR substitutionmatrix, base frequencies estimated by ML, a four category G dis-tribution (a5 0.63), and a proportion of invariant sites (0.176))selected using MODELGENERATOR analysis (Keane et al.
561BASS ET AL.—MOLECULAR ECOLOGY OF CYCLOTRICHIIDS
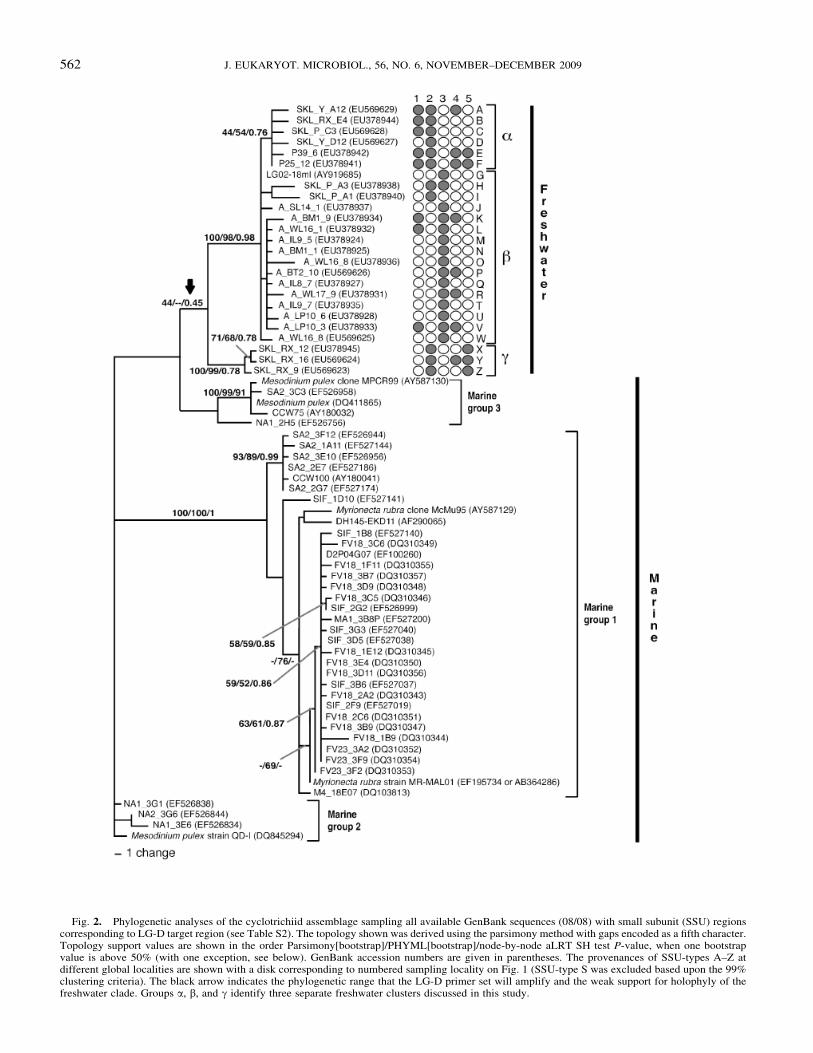
Fig. 2. Phylogenetic analyses of the cyclotrichiid assemblage sampling all available GenBank sequences (08/08) with small subunit (SSU) regionscorresponding to LG-D target region (see Table S2). The topology shown was derived using the parsimony method with gaps encoded as a fifth character.Topology support values are shown in the order Parsimony[bootstrap]/PHYML[bootstrap]/node-by-node aLRT SH test P-value, when one bootstrapvalue is above 50% (with one exception, see below). GenBank accession numbers are given in parentheses. The provenances of SSU-types A–Z atdifferent global localities are shown with a disk corresponding to numbered sampling locality on Fig. 1 (SSU-type S was excluded based upon the 99%clustering criteria). The black arrow indicates the phylogenetic range that the LG-D primer set will amplify and the weak support for holophyly of thefreshwater clade. Groups a, b, and g identify three separate freshwater clusters discussed in this study.
562 J. EUKARYOT. MICROBIOL., 56, NO. 6, NOVEMBER–DECEMBER 2009

2004). To test the support for this topology we performed 100bootstrap replicates and node-by-node aLRT SH-tests using thePHYML program (Anisimova and Gascuel 2006).
The DNA alignment showed a very low level of variabilityacross the alignment with multiple insertion/deletion sites thatwere potentially informative for resolving the cyclotrichiid phy-logeny. However, the PHYML method ignores these potentiallyinformative insertion/deletion characters during tree calculations.We therefore re-analysed our dataset using a parsimony methodthat counted alignment gaps (insertions and deletions) as a fifthcharacter state. Although parsimony methods generally do not in-corporate correction for site rate variation, they were justified herebecause of the low sequence variation in our dataset. A parsimonytopology was calculated using 100 random replicate heuristicsearches with the tree-bisection-reconnection (TBR) branch-swapping algorithm (Swofford 2002). A further 100 bootstrappseudo-replicate parsimony analyses were conducted using thesame settings but with a single TBR search per replicate.
Ecological statistics. We used canonical correspondence anal-ysis (CCA) to investigate the effect of the environmental variableson the distribution of cyclotrichiid SSU-types. We combined de-tailed physical and chemical variable measurements taken fromeach lake at the time of sampling (Table S3) with presence andimplied absence points (negative PCR results verified as describedabove) for each of the SSU-types detected in the Adirondack lakes.Presence–absence data were Hellinger transformed (Legendre andGallagher 2001). In practice the Hellinger transformation had al-most no effect on the ordination. No sample had more than fiveSSU-types present and it is known that if sites do not differ in taxonrichness Hellinger transformation has no effect (Legendre, Borcard,and Peres-Neto 2005).
Six of the 18 sample sites yielded no PCR products; CCA doesnot permit inclusion of samples where no study species werefound. As we were interested in understanding the environmentalconditions associated with absence/rarity of cyclotrichiids,we created a dummy SSU-type representing absence of anyPCR reaction product, which we recorded as present at all six ofthese sites. We normalised the environmental data sets (mean 5 0,variance 5 1.0), then tested for multicolinearity because theresults obtained by CCA (a multiple regression method) may beunreliable if the environmental variables are highly correlated(Ter Braak 1987). Three environmental variables (i.e. total Al,NH4-N, and SO4-S) with coefficients of determination (R2) 40.9were sequentially removed from the analysis. Canonical corre-spondence analysis was carried out with Ecological CommunityAnalysis version 2.0 (Seaby, Henderson, and Prendergast 2004).A Monte Carlo permutation test (1,000 replicates) was used toestablish which of the ordination axes explained significantlymore of the variation than expected by chance.
The data were then re-analysed, excluding those samples inwhich no PCR reaction products were found, in order to explorethe relationship between LG-D SSU-type community structureand environmental variables. As this reduced the dataset and pro-duced a much narrower range of environmental conditions therewas a much higher level of multicolinearity in the environmentaldata. Therefore, redundancy analysis (Legendre and Legendre1998) was used to select those environmental variables that con-tributed most to variability in the SSU-type data. Three variables(sample depth, pH, and total P) were selected for the CCA. AMonte Carlo test (1,000 replicates) was used to test the signifi-cance of the canonical axes. We constructed a matrix of similaritybetween samples based on their SSU-type community composi-tion, using the S�rensen similarity index. The Euclidean distancesbetween all pairs of sites were calculated by plotting the positionof all lakes in a GIS (MapInfo v8.0, PitneyBowes, Troy, NY). AMantel test was used to look for a correlation between composi-
tional similarity and geographical separation in order to test thehypothesis that those sample sites that were close together wouldbe more similar in SSU-type community composition.
RESULTS
The phylogenetic analysis of cyclotrichiid sequences fromGenBank and those generated by this study by our group-specificprimers designed to the LG-D clade showed that all 24 of our newSSU-types can be assigned to three non-marine phylogeneticgroups—a, b, and g (Fig. 2; the LG-D primer range is markedby a bold arrow). Fourteen LG-D SSU-types were recovered fromglobally dispersed sampling sites separated by oceans: two SSU-types were found in four global sites (E, F, Fig. 2), four werefound in three global sites (A, K, V, Y, Fig. 2), and eight in twoglobal sites (B, C, H, L, P, R, X, Z, Fig. 2). In contrast, 11 SSU-types were detected in only one biogeographical region.
Our analysis of environmental sequences suggests clear phylo-genetic separations exist between the marine and the freshwatergroups (Fig. 2). Groups a, b, and g were recovered from 26 en-vironmental gene libraries (i.e. 24 reported here plus two fromRichards et al. (2005)) and are mainly freshwater with two soilsamples collected at close proximity to freshwater; all other se-quences in Fig. 2 were from eight independent marine studies. OurLG-D-specific (freshwater) primers failed to produce PCR prod-ucts from any marine eDNA sample suggesting that organismsdetected using the LG-D primer set are absent or rare in marineenvironments. Reciprocal experiments using marine group-spe-cific primers are needed to confirm the marine and non-marineexclusivity of these groups.
The available marine-derived sequences fall into three looselygrouped phylogenetic clusters, labelled Marine Groups 1–3 (Fig. 2).Marine Groups 2 and 3 contain three M. pulex isolates, showing thisspecies (if ‘‘correctly’’ identified in each case) to be paraphyletic.Marine Group 1 comprises sequences from cultured isolates namedM. rubra plus 25 environmental sequences and further environmen-tal sequences branching closely to this ‘‘M. rubra’’ cluster.
Within the Adirondack lake system, a CCA ordination plot ofsample scores shows the lake water environmental samples thathave a similar composition of SSU-types relative to environmen-tal gradients (Fig. 3). The five lakes (six eDNAs) that yielded noPCR products have a markedly different position from LG-D-positive lakes (12 eDNA samples) on the ordination plot. The LG-D-negative lakes have a high loading on dissolved organic carbon,total P, and chl a. Only the first canonical axis was significant(Monte Carlo permutation test, P 5 0.044), and explained 22% ofthe total variability. A separate CCA was carried out on only thosesites producing LG-D-positive PCR products. Again, only thisfirst axis was significant (Monte Carlo permutation test,P 5 0.009), and explained 19.7% of the total variability (Fig. 4).This axis loaded strongly for sample depth, pH, and total P, sug-gesting that these abiotic factors are important for governing theenvironmental distribution of subgroups within the LG-D group.A Mantel test showed that physical distance between the samplesites did not explain any differences in terms of SSU-type detec-tion (r 5 0.20, P 5 0.118).
DISCUSSION
Our use of highly group-specific primers has shown that theLG-D clade represents a diverse and differentiated cluster offreshwater ciliates. Groups a, b, and g are each moderately tostrongly supported and the lengths of the branches separatingthese phylogenetic clusters suggest that they may turn out to rep-resent distinct cyclotrichiid genera. There is now a strong case forisolating and describing as many of these as possible and inves-
563BASS ET AL.—MOLECULAR ECOLOGY OF CYCLOTRICHIIDS

tigating their roles in freshwater ecology. Data from other genesare required to resolve the phylogenetic relationships of thesefreshwater subgroups, and to resolve more robustly the number ofmarine–freshwater transitions discussed below. Perhaps, like M.pulex (Beaver and Crisman 1989) other cyclotrichiid species willprove to be useful as indicators of environmental change.
Ecological biogeography and differentiation of LG-D lin-eages. There is highly robust marine and non-marine clustering incyclotrichiids suggesting that the LG-D group is not viable inmarine conditions. Our data suggest that there have been one(groups a1b1g) or two (a1b; g) freshwater–marine transitions,depending on whether the freshwater group is holophyletic. Thisrelationship could not be strongly supported or rejected by ourphylogenetic analyses. Therefore, it is not possible to infer theancestral state for cyclotrichiids or polarise the number of fresh-water/marine transitions within this clade from the results re-ported here. However, the clear freshwater/marine divide isconsistent with results for some other protist groups (von derHeyden and Cavalier-Smith 2005; von der Heyden, Chao, andCavalier-Smith 2004) and inferred from a meta-analysis of envi-
ronmental SSU-types present in GenBank (Richards and Bass2005).
To investigate the possibility that individual cyclotrichiid SSU-types have different ecological preferences/requirements at ahigher level of phylogenetic resolution we intensively sampledone geographical region—the Adirondack Park, USA. Becausehighly specific primers were used for replicated LG-D PCRs on 18identically processed eDNAs from 13 Adirondack lakes and thePCR amplifications with negative signal were stringently tested(see ‘‘Materials and Methods’’), we infer that PCR reactions fromAdirondack samples yielding no PCR products represent sam-pling sites in which members of the LG-D groups a, b, and g areeither relatively rare or absent. This inference is supported bylakes in which both epilimnion and hypolimnion layers were sam-pled: PCR products were obtained from either both, or neitherlayer in all six cases.
The CCA ordination plot in Fig. 3 implies that lakes with rel-atively high dissolved organic carbon, total P, and chl a are notsuitable habitat for the abundant LG-D populations. These resultssuggest a different ecological trend from that described for a
Environment Axis 1
Env
ironm
ent A
xis
1
Species/Samples Axis 1
Spe
cies
/Sam
ples
Axi
s 2
2.6
2.4
2.2
2
1.8
1.6
1.4
1.2
1
0.8
0.6
0.4
0.2
0
–0.2
–0.4
–0.6
–0.8
–1
–1.2
–1.4
–1.6
–1.8
–2
–2.2
–2.4
–2.6
0.65
0.6
0.55
0.5
0.45
0.4
0.35
0.3
0.25
0.2
0.15
0.1
0.05
0
–0.05
–0.14
–0.15
–0.2
–0.25
–0.3
–0.35
–0.4
–0.45
–0.5
–0.55
–0.6
–0.65
L10L16
L8L2
L11L15L13L17
Fe mg/LL14sample depth
NO -N (mg N/L) 3
Na (mg/L)
L1
L9
L12
0
K (mg/L)
L4 L3 L5
L6 L7 L18
pH
total P
DOC (mg/L)
Mg (mg/L)
chl a
–3 –2 –1 0 1 2 3
no cyclotrichiidsdetected
Fig. 3. Canonical correspondence analysis (CCA) ordination plot of all sampling sites in the Adirondacks lakes region (L1–L18: see Table S3 fordetails of the lake samples), including lakes in which no cyclotrichiid polymerase chain reaction signal was detected. Only the first canonical axis, loadingstrongly for dissolved organic carbon, total phosphorus, and chlorophyll a, is significant.
564 J. EUKARYOT. MICROBIOL., 56, NO. 6, NOVEMBER–DECEMBER 2009
freshwater form of M. pulex, which is represented by MarineGroups 2 and/or 3 in our study. Beaver and Crisman (1989) notedthat this species becomes steadily more common with increasingeutrophication in freshwater Floridian lakes (i.e. trophic statusclassified by mean annual chl a levels). However, the paraphyly ofthe marine ‘‘M. pulex’’ ‘‘species’’ as shown in our phylogenetictrees is revealing of the danger of using genetically uncircum-scribed morphospecies as environmental indicators—clearly bothof these ‘‘marine’’ groups cannot include the true M. pulex; there-fore, which (if either) contains the indicator organism referred toby Beaver and Crisman (1989) is unknown. If these groups turnout to be exclusively marine, then ‘‘M. pulex’’ of Beaver andCrisman (1989) very probably represents a third genotype asso-ciated with this ‘‘species’’. Again, phylogenies based on genesother than (or in addition to) SSU rDNA are required to resolvethe relationship between these groups. Therefore, and in the lightof increasing awareness of the mutual exclusivity of sets of protistgenotypes found in marine and non-marine environments (Bass etal. 2009; von der Heyden and Cavalier-Smith 2005), it is probablethat ‘‘Mesodinium pulex’’ as referred to in much of the literatureis not a single species.
The same may be true for M. rubra, the taxonomic status ofwhich is unclear. It occurs in four statistically supported sizeclasses, generally considered to be environmentally determinedvariants of the same taxon, and is found in water bodies differingsignificantly in environmental parameters (Montagnes et al.2008). It is therefore possible that the diversity of genotypes inMarine Group 1 may be directly related to the morphological andecological diversity increasingly reported for this ‘‘species’’.
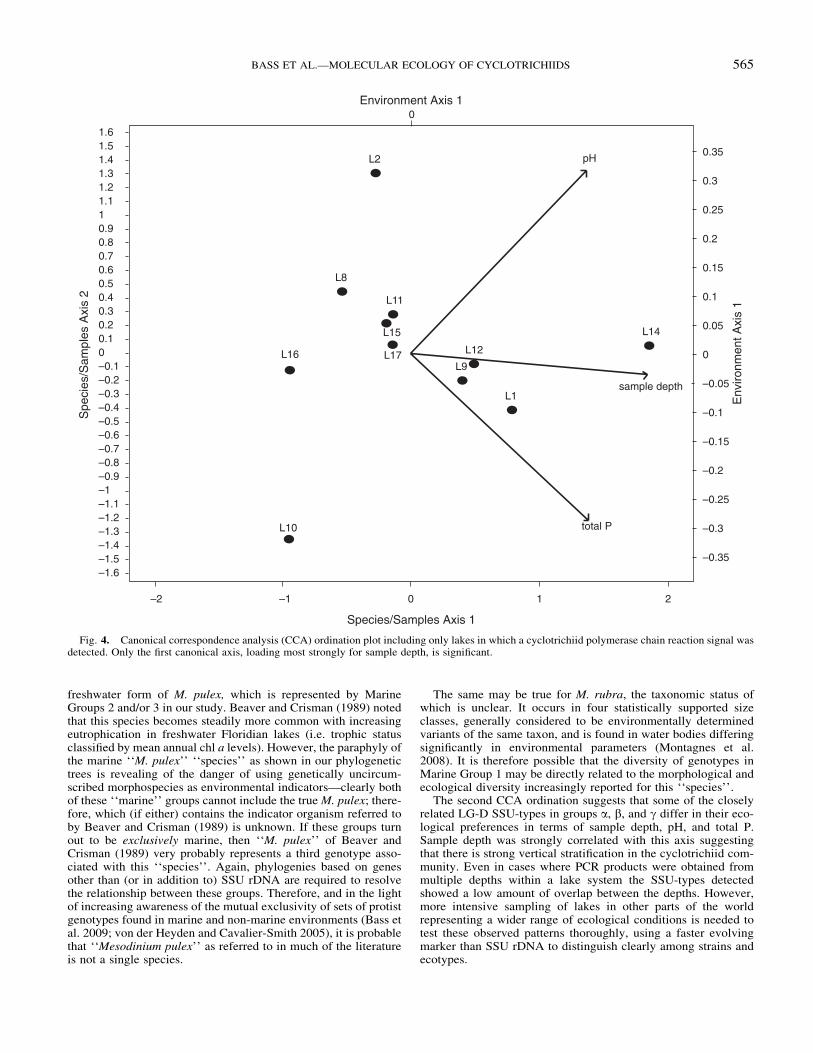
The second CCA ordination suggests that some of the closelyrelated LG-D SSU-types in groups a, b, and g differ in their eco-logical preferences in terms of sample depth, pH, and total P.Sample depth was strongly correlated with this axis suggestingthat there is strong vertical stratification in the cyclotrichiid com-munity. Even in cases where PCR products were obtained frommultiple depths within a lake system the SSU-types detectedshowed a low amount of overlap between the depths. However,more intensive sampling of lakes in other parts of the worldrepresenting a wider range of ecological conditions is needed totest these observed patterns thoroughly, using a faster evolvingmarker than SSU rDNA to distinguish clearly among strains andecotypes.
Species/Samples Axis 1
Spe
cies
/Sam
ples
Axi
s 2
Environment Axis 1
Env
ironm
ent A
xis
1
1.61.51.41.31.21.110.90.80.70.60.50.40.30.20.10–0.1–0.2–0.3–0.4–0.5–0.6–0.7–0.8–0.9–1–1.1–1.2–1.3–1.4–1.5–1.6
0.35
0.3
0.25
0.2
0.15
0.1
0.05
0
–0.05
–0.1
–0.15
–0.2
–0.25
–0.3
–0.35
0
–2 –1 1 2
L10
L16
L8
L2
L17
L15
L11
L14
L1
L9
L12
total P
sample depth
pH
0
Fig. 4. Canonical correspondence analysis (CCA) ordination plot including only lakes in which a cyclotrichiid polymerase chain reaction signal wasdetected. Only the first canonical axis, loading most strongly for sample depth, is significant.
565BASS ET AL.—MOLECULAR ECOLOGY OF CYCLOTRICHIIDS

Other variables are also likely to be important. For exampleMontagnes et al. (2008) showed that nitrate concentration was thevariable most strongly correlated with changes in size distributionin M. rubra. Our study suggests that nitrate concentration is im-portant in determining the ecological distribution of at least someLG-D SSU-types: lake sample 14 (L14—Sagamore Lake hypo-limnion layer; Table S3) has a uniquely high nitrate loading, inwhich SSU-types W, G, and J were detected. Interestingly, SSU-type G was detected only in this sample, and J also in L1 and L12;these lakes also have slightly higher nitrate loading than all othersexcept L14, suggesting that some SSU-types may become moreabundant with increased nitrate loading.
Historical biogeography. Our data demonstrate wide dis-persal of at least some members of groups a, b, and g, and sug-gest that others have restricted distributions and/or are variablyabundant across a wider geographical range. Whichever of thesescenarios is true, our results support the growing acceptance thatthe distribution of many protists fits a ‘‘moderate endemicitymodel’’ (Bass et al. 2007; Foissner 2006; Telford, Vandvik, andBirks 2006; Vyverman et al. 2007). However, the majority ofthese ‘‘restricted’’ SSU-types were found only in the Adirondacklakes, which were by far the most thoroughly sampled of oursampling sites. Thus their appearance here and nowhere else mayreflect this disparity in sampling effort, and/or that some ecolog-ical requirements of the ‘‘restricted’’ strains were met moststrongly at some Adirondack lake sampling sites.
Groups a, b, and g differ in their global distribution profiles.Groups a and g were not recovered from the intensively sampledAdirondacks, even though the highly specific primers were capa-ble of detecting them. Because members of each group showbroad global distributions, this absence is unlikely to be historicbiogeographical structuring. Rather, it probably reflects as yet un-known ecological factors. In support of this, groups a and g com-prise sequences from eutrophic freshwater sediment samples fromSakhalin Island and freshwater sediment samples from Panama,New Zealand, and Europe. As members of group b were exclu-sively from freshwater columns, it may be that members of thisgroup b show a greater preference for sediment-free open waterthan groups a and g.
CONCLUSIONS
To our knowledge this study is the first to combine highly spe-cific environmental gene libraries detecting a phylogeneticallydefined set of protist strains in a sensitive and relatively bias-freeway and multiple co-measured environmental variables to inferthe strains’ ecological preferences in a freshwater lake system. Weshow clear environmental differences between lakes in whichcyclotrichiids are easily detected and those in which we detectednone. This study also demonstrates that even in water bodies sit-uated close to each other and sharing many physio-chemical prop-erties, relatively small differences in ecologically importantvariables can have significant impacts on microbial distribution.These differences in turn may affect the mode of action of themicrobial loop, community assembly, and larger, filter-feedinganimals. In fact, the relatively small proportion of variability ex-plained by the first canonical axes in the CCA plots suggests thatenvironmental factors other than those measured here are equallyor more important in explaining cyclotrichiid community assem-bly. We suggest that these are likely to be biological interactionsin a diverse and perhaps unexpectedly highly compartmentalisedlacustrine water column.
ACKNOWLEDGMENTS
T.A.R. is supported by BBSRC, NERC, and the LeverhulmeTrust, DB NERC. This research was supported by the NaturalHistory Museum, London grant-in-aid. This study was in partsupported by a grant from the U.S. Environmental ProtectionAgency (contract number 68D20171) as part of the AdirondackEffects Assessment Program (AEAP) and the Helen V. FroehlichFoundation to S.A,N.-B. This work has not been subjected to U.S.EPA Agency review and, therefore, does not necessarily reflectthe views of the agency, and no official endorsement should beinferred. We are indebted to our colleagues in the AEAP that col-lected and processed water samples, in particular, Robert Bom-bard, Larry Eichler, Laurie Ahrens-Franklin, Jim Sutherland, andDavid Winkler. We also thank Peter Henderson for advice re-garding statistical methods and software.
LITREATURE CITED
Anisimova, M. & Gascuel, O. 2006. Approximate likelihood-ratio test forbranches: a fast, accurate, and powerful alternative. Syst. Biol., 55:539–552.
Bass, D. & Cavalier-Smith, T. 2004. Phylum-specific environmental DNAanalysis reveals remarkably high global biodiversity of Cercozoa (Pro-tozoa). Int. J. Syst. Evol. Microbiol., 54:2393–2404.
Bass, D., Richards, T. A., Matthai, L., Marsh, V. & Cavalier-Smith, T.2007. DNA evidence for global dispersal and probable endemicity ofprotozoa. BMC Evol. Biol., 7:162, doi: 10.1186/1471-2148-7-162.
Bass, D., Howe, A. T., Mylnikov, A. P., Vickerman, K., Chao, E. E.-Y.,Edwards Smallbone, J., Snell, J., Cabral, J. C. & Cavalier-Smith, T.2009. Phylogeny and classification of Cercomonadida: Cercomonas,Eocercomonas, Paracercomonas, and Cavernomonas gen. n. Protist, inpress. doi: 10.1016/j.protis.2009.01.004.
Beaver, J. R. & Crisman, T. L. 1989. Analysis of the community structureof planktonic ciliated protozoa relative to trophic state in Florida lakes.Hydrobiologia, 174:177–184.
Chantangsi, C. & Lynn, D. H. 2008. Phylogenetic relationships within thegenus Tetrahymena inferred from the cytochromes c oxidase subunit 1and the small subunit ribosomal RNA genes. Mol. Phylogenet. Evol.,49:979–987.
Coleman, A. W. 2007. Pan-eukaryote ITS2 homologies revealed by RNAsecondary structure. Nucleic Acids Res., 35:3322–3329.
Foissner, W. 2006. Biogeography and dispersal of micro-organisms: a re-view emphasizing protists. Acta Protozool., 45:111–136.
Foissner, W., Berger, H. & Schaumburg, J. 1999. Identification and ecol-ogy of limnetic plankton ciliates. Informationberichte des BayerischenLandesamtes fur Wasserwirtschaft, 3/99, 793 p.
Guindon, S. & Gascuel, O. 2003. A simple, fast, and accurate algorithm toestimate large phylogenies by maximum likelihood. Syst. Biol., 52:696–704.
Howe, A. T., Bass, D., Vickerman, K., Chao, E. E. & Cavalier-Smith, T.2009. Phylogeny, taxonomy, and astounding genetic diversity ofGlissomonadida ord. nov., the dominant gliding zooflagellates in soil(Protozoa: Cercozoa). Protist, 160:159–189.
Jankowski, A. W. 1980. Conspectus of a new system of the phylumCiliophora. Trudy Zool. Inst. Leningr., 107:80–115.
Johnson, M. D., Tengs, T., Oldach, D. W., Delwiche, C. F. & Stoecker, D.K. 2004. Highly divergent SSU rRNA genes found in the marine ciliatesMyrionecta rubra and Mesodinium pulex. Protist, 155:347–359.
Judo, M. S. B., Wedel, A. B. & Wilson, C. 1998. Stimulation and sup-pression of PCR-mediated recombination. Nucleic Acids Res., 26:1819–1825.
Keane, T. M., Creevey, C. J., Naughton, T. J., Pentony, M. M., Naughton,T. J. & Mcinerney, J. O. 2004. Assessment of methods for amino acidmatrix selection and their use on empirical data shows that ad hoc as-sumptions for choice of matrix are not justified. BMC Evol. Biol., 6:29.
Krainer, K.-H. & Foissner, W. 1990. Revision of the genus AskenasiaBlochmann, 1895, wih proposal of two new species, and descriptionof Rhabdoaskenasia minima n.g., n.sp. (Ciliophora, Cyclotrichida).J. Protozool., 37:414–427.
Legendre, P. & Gallagher, E. 2001. Ecologically meaningful transforma-tions for ordination of species data. Oecologia, 129:271–280.
566 J. EUKARYOT. MICROBIOL., 56, NO. 6, NOVEMBER–DECEMBER 2009

Legendre, P. & Legendre, L. 1998. Numerical Ecology. 2nd ed. ElsevierScience, Elsevier, Amsterdam. 853 p.
Legendre, P., Borcard, D. & Peres-Neto, P. 2005. Analyzing beta diver-sity: partitioning the spatial variation of community composition data.Ecol. Monogr., 75:435–450.
Lopez-Garcıa, P., Rodriguez-Valera, F., Pedros-Alio, C. & Moreira, D.2001. Unexpected diversity of small eukaryotes in deep-sea Antarcticplankton. Nature, 409:603–607.
Lynn, D. H. & Struder-Kypke, M. C. 2006. Species of Tetrahymena iden-tical by small subunit rRNA gene sequences are discriminated by mi-tochondrial cytochrome c oxidase I gene sequences. J. Eukaryot.Microbiol., 53:385–387.
Montagnes, D. J., Allen, J., Brown, L., Bulit, C., Davidson, R., Dıaz-Ava-los, C., Fielding, S., Heath, M., Holliday, N. P., Rasmussen, J., Sanders,R., Waniek, J. J. & Wilson, D. 2008. Factors controlling the abundanceand size distribution of the phototrophic ciliate Myrionecta rubra inopen waters of the North Atlantic. J. Eukaryot. Microbiol., 55:457–465.
Richards, T. A. & Bass, D. 2005. Molecular screening of free-living mi-crobial eukaryotes: diversity and distribution using a meta-analysis.Curr. Opin. Microbiol., 8:240–252.
Richards, T. A., Vepritskiy, A. A., Gouliamova, D. & Nierzwicki-Bauer,S. A. 2005. The molecular diversity of freshwater picoeukaryotes froman oligotrophic lake reveals diverse, distinctive and globally dispersedlineages. Environ. Microbiol., 7:1413–1425.
Seaby, R., Henderson, P. & Prendergast, J. R. 2004. Community AnalysisPackage. Pisces Conservation Ltd., Lymington, UK.
Stoeck, T. & Epstein, S. 2003. Novel eukaryotic lineages inferred fromsmall-subunit rRNA analyses of oxygen-depleted marine environments.Appl. Environ. Microbiol., 69:2657–2663.
Struder-Kypke, M. C., Wright, A. D., Foissner, W., Chatzinotas, A. &Lynn, D. H. 2006. Molecular phylogeny of litostome ciliates (Cilio-phora, Litostomatea) with emphasis on free-living haptorian genera.Protist, 157:261–278.
Swofford, D. L. 2002. PAUP�. Phylogenetic Analysis Using Parsimony(�and Other Methods), Version 4. Sinauer Associates, Sunderland, MA.
Telford, R. J., Vandvik, V. & Birks, H. J. 2006. Dispersal limitations mat-ter for microbial morphospecies. Science, 312:1015.
Ter Braak, C. J. F. 1987. The analysis of vegetation-environment rela-tionships by canonical correspondence analysis. Vegetatio, 64:69–77.
Turner, G. K. 1985. Fluorometry. In: Van Dyke, K. (ed.), Bioluminescenceand Chemiluminescence: Instruments and Applications. CRC Press,Boca Raton, LA. 1:43–78.
USEPA. 1983. Methods for Chemical Analysis of Water and Wastes. US-EPA 600/4-79-020, Cincinnati, OH. 552 p.
von der Heyden, S. & Cavalier-Smith, T. 2005. Culturing and environ-mental DNA sequencing uncover hidden kinetoplastid biodiversity anda major marine clade within ancestrally freshwater Neobodo designis.Int. J. Syst. Evol. Microbiol., 55:2605–2621.
von der Heyden, S., Chao, E. E. & Cavalier-Smith, T. 2004. Genetic di-versity of goniomonads: an ancient divergence between marine andfreshwater species. Eur. J. Phycol., 39:343–350.
Vyverman, W., Verleyen, E., Sabbe, K., Vanhoutte, K., Sterken, M.,Hodgson, D. A., Mann, D. G., Juggins, S., Van de Vijver, B., Jones,V., Flower, R., Roberts, D., Chepurnov, V. A., Kilroy, C., Van-ormelingen, P. & De Wever, A. 2007. Historical processes constrainpatterns in global diatom diversity. Ecology, 88:1924–1931.
SUPPORTING INFORMATION
Additional Supporting Information may be found in the onlineversion of this article:
Table S1. Summary of described (putative) cyclotrichiid spe-cies. Names underlined in the left-hand column are those forwhich sequences are available and shown in Fig. 2 and Supple-mentary Fig. 2. Dashes indicate no data available.
Table S2. Cyclotrichiid-like sequences in GenBank (12/02/2008).
Table S3. Adirondack lake water sampling.Please note: Wiley-Blackwell are not responsible for the con-
tent or functionality of any supporting materials supplied by theauthors. Any queries (other than missing material) should be di-rected to the corresponding author for the article.
Received: 09/28/08, 05/05/09, 06/07/09; accepted: 06/13/09
567BASS ET AL.—MOLECULAR ECOLOGY OF CYCLOTRICHIIDS