6. als igg reduced ca21 channel activity in a side-selective fashion
TRANSCRIPT

Journal of IPhysiology (1993). 461. pp. 103-118 1031T'ith, 7 fiqures
Printed in Great Britain
THE ACTION OF AMYOTROPHIC LATERAL SCLEROSISIMMUNOGLOBULINS ON MAMMALIAN SINGLE SKELETAL MUSCLE
Ca2l CHANNELS
BY VALERIA MAGNELLI*, TADAO SAWADA, OSVALDO DELBONO,R. GLENN SMITHt, STANLEY H. APPELt AND ENRICO STEFANIt
Fronm the Departments of Molecular Physiology and Biophysics and tNeurology,Baylor College of Miedicine, One Baylor Plaza, Houston, TX 77030, USA
(Received 27 January 1992)
SUMMARY
1. The planar phospholipid bilayer technique was used to study the T-tubuleskeletal muscle dihydropyridine (DHP)-sensitive calcium (Ca2") channel. To improvethe signal-to-noise ratio, Ca2" channel activity was recorded using both 800-50 and500-50 mm NaCl gradients.
2. Ca21 channels were characterized by their cation selectivity and pharma-cological profile. The mean open time for channels identified by these techniques wasincreased by the DHP agonist Bay K 8644 (2 /LM), while it was decreased by the DHPantagonist nifedipine (5 /yM). Nifedipine also reduced Ca21 channel amplitude levels.
3. Immunoglobulins G (IgG) from three amyotrophic lateral sclerosis (ALS)patients (n = 14 experiments), one myasthenia gravis (MG) patient (n = 3experiments) and one healthy individual (n = 4 experiments), were tested onCa21 channel activity at a final concentration of 3 mg/ml.
4. Channel mrean open time, mean closed time and time integral for the currentwere not modified by normal IgG (n = 4 experiments). Similarly, MG IgG did notreduce channel activity (n = 3 experiments).
5. ALS IgG reduced the mean open time of DHP-sensitive Ca21 channel activityin twelve out of fourteen experiments. In addition, in five out of twelve experiments,ALS IgG stabilized the channel to a smaller amplitude level.
6. ALS IgG reduced Ca21 channel activity in a side-selective fashion, probablycorresponding to the external side of the channel.
7. These results suggest that ALS IgG action on DHP-sensitive Ca2+ channels isnot mediated by second messengers, thus favouring a direct mechanism for interactionwith the DHP receptor complex.
INTRODUCTION
Amyotrophic lateral sclerosis (ALS) is a devastating human illness which affectsupper and lower motoneurons culminating in respiratory compromise and death
* Fellow of the Department of Human Physiology, UTniversity of Ferrara, Italy.t To whom all correspondence should be addressed.
MIS 1072

14 MAG41NELLI AND OTHERS
(Appel & Stefani, 1991). The etiology of the disease is still unknown, but recent workhas provided evidence for an autoimmune mechanism in the etiology of ALS. Ananimal model of immune-mediated destruction of motoneurons has been developed,which resembles human ALS with respect to the loss of lower and upper
motoneurons (Engelhardt, Appel & Killian, 1989, 1990), the presence of immuno-globulins G (IgG) within motoneurons (Engelhardt & Appel, 1990), and alteredphysiology of the motor axon terminals (Garcia, Engelhardt, Appel & Stefani, 1990).Serum and IgGE from the animal models and human ALS can passively transfer theenhanced miniature endplate postsynaptic potentials (MEPP) frequency of theneuromuscular junction to mice (Appel, Engelhardt, GCarcia & Stefani, 1991). ALSIgG can reduce macroscopic Caa2+ currents (ICa) and charge movement in singlemammalian skeletal muscle fibres (Delbono, Garcia, Appel & Stefani, 1991 a, b).These observations suggest either a direct or an indirect interaction of ALS IgG withdihydropyridine (DHP)-sensitive Ca2+ channels. To further define whether theinteraction of ALS IgG with these channels was direct or indirect, we investigatedthe action of ALS IgG on reconstituted skeletal muscle DHP-sensitive Ca2+ channelsin planar lipid bilayers (Coronado, 1987; Ma & Coronado, 1988; Mejia-Alvarez, Fill& Stefani, 1991). The experiments were designed to test whether ALS IgG actsdirectly on the DHP-sensitive Ca2+ channel or via some second messenger system.Our data suggest a direct action, since ALS IgG reduces Ca2+ channel activityreconstituted in bilayers that contains no cytoplasmic second messenger system.
METHODS
Single-channel recordingsSingle-channel experiments were performed using the planar lipid bilayer technique with the
Mueller-Rudin method. A mixture of lipids dissolved in a non-polar liquid (n-decane), was paintedacross a small hole (200 ,tm) through a thin partition (500 Itm) between twocups. The lipid mixturecontained phosphatidylethanolamine, phosphatidylserine and phosphatidylcholine (5:3:2,50 mg/ml; Avanti Polar-Lipids Inc., Birmingham, AL, USA). The side to which the potential was
applied is referred to as cis, while the ground side is designated the trans side. The vesicles were
added to the cis side. Since vesicles from tubular membranes usually have the inside-outorientation (Pelzer, Grant, Cavali6, Pelzer, Sieber, Hofmann & Trautwein, 1989), the intracellularchannel side would normally face the cis chamber, while the extracellular side should face the transchamber. Single-channel activity was recorded with the following solutions: trans side: 50 mm
NaCl,1 mm EGTA and 10 mm Na-Hepes and cis side: 500 mm NaCl,1 mm EGTA and 10 mM
Na-Hepes or 800 mm NaCl,1 mm EGTA and 10 mm Na-Hepes, pH 7-4. The theoretical reversalpotential for Na+ ions (ENa) is -58-0 mV (500-50 mm Na+) or -69-8 mV (800-50 mm Na'). In mostexperiments Bay K 8644 (final concentration of 2,IM) was added to both chambers aliquoting froma 1 mM stock solution in ethanol. All experiments were performed at room temperature (20-22°C).Ag-AgCl electrodes were connected to the chambers with 50 mm NaCl-agar bridges. CommandVoltages to the bilayer and data acquisition were performed using pClamp software with a TL-tAxon Instruments interface (Axon Instruments Inc., Foster City, CA, USA). The bilayer amplifierwas designed to obtain the best signal-to-noise ratio and a complete description of the circuit hasalready been shown elsewhere (Hamilton, Mejia-Alvarez, Fill, Hawkes, Brush, Schilling & Stefani,1989). Data were filtered at 500 Hz through an 8-pole Bessel filter and acquired at 400 Its per point.Channel records were analysed with a custom-made software that corrected for dead time andsampling frequency. Dwell times were fitted to the probability density function using themaximum likelihood method (Colquhoun & Sigworth, 1983). Dwell times were plotted usinglogarithmic binning of the time axis (Sigworth & Sine, 1987). To evaluate the time course ofchannel activity, diary plots were obtained by plotting the time integral of the current in each trace(819-2ms) vs. time. The software was kindly provided by A. M. Brown, A. M. T. Van Dongen, H.
104

ACTION OF ALS IgG ON Ca2+ CHANNELS
B
2 gIM Bay K 8644
NaCI, 500-50 mM; 0 mV
c
cl
2 pA
c
1 s
C
5000 -
j100 200
Bay K 8644
300
vL1,400 500 600
Time (s)
Fig. 1. Action of Bay K 8644 on Ca2l channel. A, traces recorded at 0 mV steady-statepotential in 500-50 mM cis-trans NaCl gradient. B, traces recorded from the same channelin the presence of 2 /LM Bay K 8644 in the cis chamber. In this and following figures thec level marks the closed state. C, in this and following figures the diary plot is the timeintegral of the current vs. time. Each point corresponds to an interval of 819-2 ms. In thisplot the arrow marks the addition of Bay K 8644.
A
Control
QRE
CL
C.,-c
c
00)
a,4--
CD
E
4000-
3000 -
2000 -
1000 -
.I.00
ii
105
I
V. MAGNELLI AND OTHERS
B
t#Contro I 45 Jm nifedipine
c~~~~~~~~~~~~c
aCl, 800-50 mM; 0 mV1 s
C
4000
C'aE< 3000-
0.
1 000
0
0 100 200 300 400Time (s)
Nifedipine
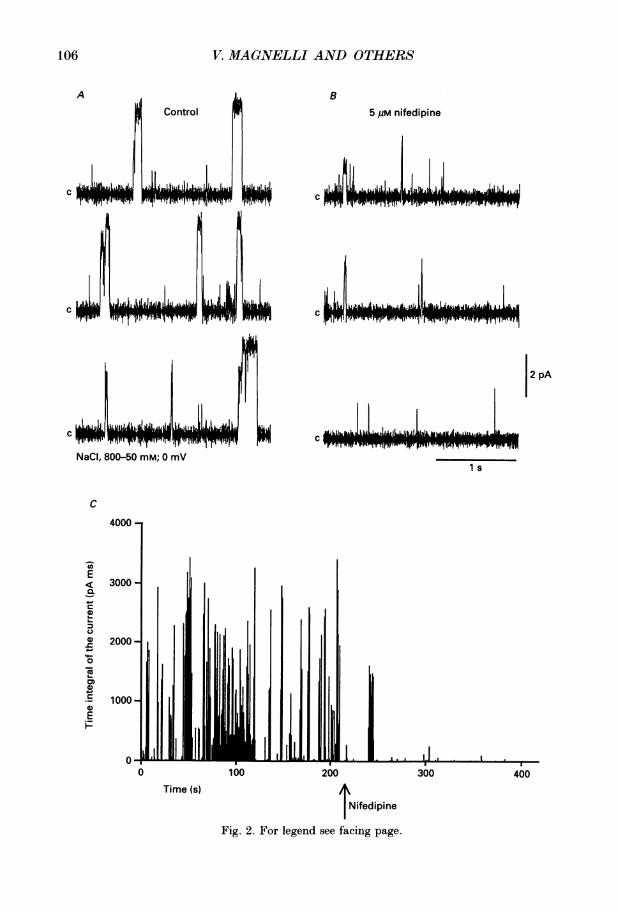
Fig. 2. For legend see facing page.
106
N

ACTION OF ALS IgG ON Ca2+ CHANNELS
Nguyen and B. Steiner of the Department of Molecular Physiology and Biophysics, Baylor Collegeof Medicine, Houston, and R. Latorre of the University of Chile.
Transverse tubule preparationT-tubule membranes were obtained from pieces of skeletal muscle weighing 250-300 g from hind
legs and back of adult New Zealand White rabbits following the procedures of our laboratory(Hamilton et at. 1989). The animals were killed before muscle dissection in a CO2 chamber. In short,the tissue was minced and homogenized at different speeds in a buffer of 0 3 M sucrose and 20 mmTris maleate (pH= 72). A mixture of protease inhibitors was added to the buffer (100,UMphenylmethylsulphonyl fluoride, 1 ,UM pepstatin A, 100 /aM p-aminobenzamidine, 1 /ug/ml aprotininand 1 ,ug/ml leupeptin) prior to tissue homogenization, and it was present in all subsequentsolutions in the preparation. The muscle homogenate was centrifuged (3000 g, 30 min) and thepellet was rehomogenized with the same buffer. The first and the second supernatants were collectedtogether and centrifuged at a higher speed (8600 y, 30 min). The pellet was discarded, and thesupernatant was first incubated for 1 h with 0-6 M KCl and then was centrifuged at 33000 g for45 min. This pellet was dissolved in 0 4 M KCl solution containing 5 mm Pipes-K (the dipotassiumsalt of piperazine-N-N'-bis(2-ethanesulphonic acid); pH 7 2), and sedimented through a sucrosegradient (67000 g, 18 h). The layers at the different interfaces were collected in 5 mm Pipes buffer(pH 7 4) and again centrifuged at 42000 g for 45 min. Finally the pellet was resuspended in 0 3 Msucrose and 100 mm KCl (pH 7 4) and stored at -80 °C until the experiments were performed.T-tubule skeletal muscle DHP-sensitive Ca2+ channel microsomes sedimented at 20-25% sucroseinterface.
Immunoglobulin preparationALS IgGs were obtained both with rivanol and ammonium sulphate precipitation (Engelhardt
et al. 1989; Appel et al. 1991). Serum proteins were precipitated with 40% ammonium sulphatesolubilized in water, dialysed against water followed by 0-1 mm NH4HC03. The material was thenlyophilized, dissolved in water, centrifuged to discard insoluble matter and then applied to proteinA-agarose columns (Pearce). The IgG fraction was eluted with 1 M glycine buffer (pH = 2) and thenneutralized with 1 M Tris to a pH of 7-5. This buffer was then exchanged with 10 mM 3-(N-morpholino)propanesulphonic acid (MOPS) buffer by dialysis before storage at -80 'C. Both ALSIgG and control IgG were used at a final concentration of 3 mg/ml.
RESULTS
Characterization of DHP-sensitive Ca2+ channels from T-tubule skeletal muscleSingle Ca2+ channel currents recorded in Na' gradients and in the absence of
divalent ions are larger and better resolved than those observed in a Ba2+ gradient(Coronado & Affolter, 1986; Coronado, 1987). Thus, we used Na+ gradients and weinitially confirmed the nature of the channels recorded by measuring the effects ofpharmacological agents.To define the nature of the recorded channels we tested the action of the DHP
agonist Bay K 8644 and the DHP antagonist nifedipine. Figure 1 shows channelopenings as upward deflections when measured at 0 mV and in 500-50 mm NaClgradient indicating that they are Na+ selective; the reversal potential was close tothe theoretical Na+ reversal potential. Figure 1 shows records in control conditions(A) and after the addition to the cis chamber of 2 aM Bay K 8644 (B). In the absence
Fig. 2. Action of nifedipine reduced on Ca2+ channels. Single channels recorded in800-50 mM cis-trans NaCl gradient at 0 mV in control conditions (A) and after theaddition of 5 am nifedipine to trans chamber (B). C, diary plot of the experiment, in whichthe arrow marks the addition of nifedipine. Bay K 8644 (2 /tM) was present throughoutthe experiment in both chambers.
107

V. MAGNELLI AND OTHERS
4 /
2O-10-5
Control
n
I---------Y------ Ut~10-3 lo- 101 10-
Time (s)
600
500
400
300
200
100 j
5,uM nifedipine
10 10 101I
10-3 lo-' 1 1
-2 0 2 4 6 -2 0 2 4 6
Amplitude (pA)Fig. 3. Nifedipine reduced Ca2+ channel activity by acting on channel open time andamplitude. A, open time histograms and fitted probability density functions (Sigworth &Sine, 1987) in control (left panel) and after adding 5 fM nifedipine to c6s and transchambers (right panel). Fitted mean values in control were 71 = 2-2 ms, A1 = 0-80 andT2 = 231 ms, A2 = 0-20. In the presence ofnifedipine the values were r1 = 1P2 ms, A1 = 0-80and r2 = 5-4 ms, A2 = 0*20. B, current amplitude histograms and fitted sum of Gaussianfunctions in control (left) and with nifedipine (right). Fitted mean values in control were
1 0 + 0 3, 3 0+ 0 3 and 4-3 ± 0-2 pA. The PO values (probability of the channel being open)corresponding to the closed state and the different current amplitude peaks were: 0 755,
A
14
12-
10
8
6-
4000L.4)
cru)
G)40
0
0.0
L-
o
Ez
B
400
100
108

ACTION OF ALS IgG ON Ca2+ CHANNELS
of Bay K 8644 channel openings are infrequent and brief (Vilven & Coronado, 1988).As expected for a DHP-sensitive Ca2+ channel, Bay K 8644 increased the mean opentime and decreased the mean closed time, in this experiment from 9-85 ms to 21V7 msand from 218 ms to 73-5 ms, respectively. The open channel probability increasedfrom 0 04 to 0-23.To evaluate channel activity we measured the time integral of the current since
DHP-sensitive tubular Ca2+ channel had frequent substates and this measurementtakes into account channel amplitude (Ma & Coronado, 1988; Mejia-Alvarez et al.1991). Figure 1 C shows the diary of the same experiment in which the time integralof the current in each trace (819-2 ms) was plotted vs. time. The arrow marks therecordings after Bay K 8644 and 1 min of continuous stirring. Channel activityincrease was detected as soon as the recording started. The average time integral ofthe current was 74-4 pA ms in control and 336 pA ms after adding Bay K 8644.To further verify the identity of the channel the effect of the DHP channel
antagonist nifedipine was tested (Fig. 2). The inhibitory effect of nifedipine wasmeasured in the presence of Bay K 8644 and in a 800-50 mm NaCl gradient.The records demonstrate that control channel activity (A) was greatly reduced by
5 /1m nifedipine (B). The mean open time was reduced from 59-3 to 1-6 ms and longerperiods without activity were observed. Furthermore nifedipine increased thefrequency of channel openings with smaller current amplitude. The time course ofthe action of nifedipine is shown in the diary plot of Fig. 2 C. The average timeintegral of the current was 681 pA ms in control conditions, and 44-8 pA ms after theaddition of nifedipine.
Figure 3 shows the effect of nifedipine on channel open time (A) and amplitudedistributions (B). Open times were plotted using logarithmic binning of the time axisin which the mean time values correspond to the time at the peaks (Sigworth & Sine,1987). The control open-time histogram was fitted with two exponential functionsgiving the following time constant (r) and amplitude factor (A) values: sr = 2-2 ms,A1= 0-80 and T2 = 231 ms, A2 = 0-20. Nifedipine shifted both time constantstowards shorter times: Tl= 1-2 ms, A1 = 0-80 and T2 = 5-4 ms, A2 = 0-20.
Channel amplitude recorded in 800-50 mm NaCl gradient at 0 mV had a maximumaverage value of 4-3 pA. In many instances, openings to smaller amplitudes andincomplete closings were detected. This suggested, as previously reported, thepresence of substates (Ma & Coronado, 1988). Nifedipine, in addition to reducingchannel open probability, also diminished channel amplitude. Figure 3B shows thetotal amplitude histograms (bars) and fitted Gaussian functions (continuous line)before (left) and after nifedipine (right). To visually enhance the smaller amplitudesthe ordinate is depicted as the square root of the number of events. The peak centredat 0 pA corresponds to the closed state. In the control histogram three differentchannel amplitudes could be distinguished, with the mean values centred at 1D0 pA(15 pS), 3 0 pA (53 pS) and 4-3 pA (62 pS). Nifedipine reduced open channelprobability and channel amplitude in such a way that only one-open level could be
0064, 0095 and 0086, respectively. In the presence of nifedipine the fitted value wasts7 +02 pA. PO for the closed state and the peak at 1-7 pA were 0986 and 0014respectively.
109
V. MAGNELLI AND OTHERS
B3 mg/ml normal IgG
c; I. IL.j..
NaCI, 500-50 mM; 10 mV
III Ii J].,., Ill.Jo' I 11 an Ii L II,iJLIII I ..4L... .. l *|l .ll I lA l L M11 LISLLJ1O1 .AlliJ 1 I 11N1 AL L 1 IL I
Tin
200 400 6(
ne (s)
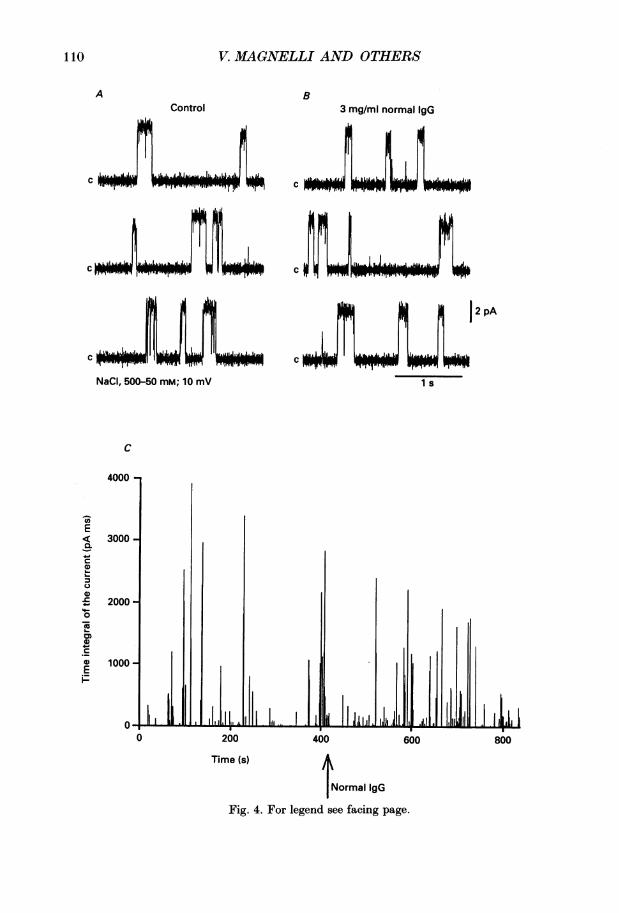
Normal IgGFig. 4. For legend see facing page.
110
AControl
c
12 pA
C
1 s
4000 -
Q
Co
E
F0C0
0
0
0
0Ei
3000 -
2000 -
1000 -
11, Ii .1.,1I III 1 1IILL0
iOO 800I
1.A4

ACTION OF ALS IgG ON Ga2+ CHANLS 111
adequately resolved. The total amplitude histogram with nifedipine was fitted bytwo Gaussian curves with a mean channel amplitude of 1P7 pA. This channelamplitude reduction cannot be explained by the reduction of mean open time, sincelong and small openings were noticeable in the presence of nifedipine (see Fig. 2B).In other experiments, Cd21 (5 mM) added to the external side of the channelcompletely abolished channel activity (data not shown).As already mentioned, the single channel opens upwardly at 0 mV with the
500-50 mM Na' gradient as is expected for being Na' selective. This was establishedby measuring the average channel amplitudes at different potentials and withdifferent Na' gradients. The reversal potential was close to the expected Na' reversalpotential of -58 mV in 500-50 mm NaCi, -70 mV in 800-50 mm NaCl, and 0 mV in800-800 mm NaCl. These findings indicate that these channels were Na' selectiveand are consistent with the view that they are Ca21 channels, since they becameselective for monovalent cations in the absence of divalent cations (Almers &McCleskey, 1984; Coronado & Smith, 1987).
In summary, these results agree with previous findings in the same preparationand confirm the nature of DHP-sensitive Ca21 channels (Affolter & Coronado, 1985;Coronado. 1987; Mejia-Alvarez et al. 1991).
Effects of control IgG on Ca21 channelWe initially tested the action of IgG from a healthy individual on Ca21 channel
activity in the presence of 2 aM Bay K 8644. Figure 4 shows that Ca2+ channelactivity was unaltered by the addition of normal IgG (3 mg/ml) when consecutivelyadded to both cis and trans chambers. The mean open time, the mean closed time,channel amplitude and the diary of channel activity were not modified by the normalIgG (n = 4). For example in Fig. 4C the average time integral values of the currentwere 97 4 and 86 pA ms respectively in control and after the addition of normal IgG.We also tested IgG obtained from one patient with myasthenia gravis (MG). In twoout of three experiments MG IgG did not affect channel activity. In the remainingcase the mean open time was increased.
Effects of ALS IgG on Ca2+ channelsALS IgG was tested in the trans chamber that should correspond to the
extracellular side of the channel since in most cases tubular membrane vesicles havethe inside-out orientation (Pelzer et al. 1989). Figure 5 shows the effects of ALS IgGon DHP-sensitive Ca2+ channels. Traces in control (A) and after adding 3 mg/mlALS IgG (B) to the trans side are shown. Records in the presence of ALS IgG wereobtained immediately after stirring the chamber solution. ALS IgG clearlydiminished channel activity by reducing the mean open time from 44-9 to 7 2 ms. Asdid nifedipine, ALS IgG made more frequent channel levels with smaller amplitudes.The average current amplitude was decreased from 3-5 to 2 2 pA. As in the case for
Fig. 4. Normal human IgG did not affect Ca2+ channel activity. A, traces recorded at10 mV steady-state potential in 500-50 mM cis-trans NaCl gradient. B, traces recordedfrom the same channel after the addition of normal human IgG both at a finalconcentration of 3 mg/ml in cis and trans chambers. C, diary plot of the experiment. BayK 8644 (2 pm) was present throughout the experiment in both chambers.
III

2V. MAGNELLI AND OTHERS
B3 mg/ml ALS IgGControl
c
c
C
C., .,. k
NaCI, 500-50 mM; 0 mV
11jil li0
TimE
IhI .11j L ...1 I ILl I. ,,. Lll.I1 I
100 200 30e (S) /
I1ALS IgG
Fig. 5. For legend see facing page.
A
..1...C "I_ _ tMNW P- r - WM4
.LIJL.UIiL6,1,1_ '_Ii- l
i2PA
C
1 s
7000 -
E 6000-E
5000-a)
24000 -c
F00)
I0 2000 -
E
ioo
400
U.1-all LL16-.F,m Irn
Io- --- ..-- - WONM""n|rM77MMr 1mr
I
112
'. L.
INTWIrm. 'ITFF-ll
u-
30
.. -I

ACTION, OF ALS IgG ON (,a2+ CHANNELS 113
nifedipine, the amplitude reduction induced by ALS IgG cannot be attributed to arecording artifact due to the filtering of brief openings since long small states weredetected (see Fig. 5B, bottom trace). The time integral of the current (Fig. 5C) wasdecreased by ALS IgG from 672 to 126 pA ms. The diary plot shows that thisreduction could be detected as soon as the stirring ended.
Figure 6 shows the effect of 3 mg/ml ALS IgG on channel mean open time (A) andchannel amplitude distribution (B). The upper panels show the open time histogrambefore (left) and after (right) adding ALS IgG. In control conditions the histogramwas fitted by two exponential functions with T1 = 19 ms, Al = 0-85 and T2= 130 ms,A2= 0 15. In the presence of ALS IgG the peaks were shifted to shorter mean opentimes with T1= 1P5 ms, A1 = 058 and T2 = 107 ms, A2 = 042.
Figure 6B shows the total amplitude histograms and fitted Gaussian functionsbefore (left) and after (right) ALS IgG. ALS IgG made more frequent smaller channelamplitudes. As in Fig. 3B, the channel in control conditions had different amplitudelevels, and in this case the largest level was predominant. The mean values were 0 99,2 77 and 4 14 pA. In the presence of ALS IgG the mean amplitude levels were 0 67,1P56 and 3-82 pA.
The effects of ALS IgG on Ca21 channels may be selective for the external sideALS IgG added to the trans chamber inhibited channel activity in nine out of
twelve experiments. In the three negative experiments, ALS IgG became effectivewhen consecutively added to the cis side. One of these experiments is shown in Fig.7. Panel A shows Ca21 channel recordings in control conditions (left panel), with ALSIgG in trans (middle panel) and with ALS IgG in both cis and trans (right panel).Panel B shows in the same sequence as panel A the diary plot of channel activity. Theaverage time integral of the current was lowered from 667 in control to 123 pA msby cis ALS IgG. The lack of effect in these experiments of ALS IgG in the trans sidecan be explained by an opposite sidedness of the channel insertion in the bilayer. Inthe experiments in Fig. 7 the extracellular side would be facing the cis chamber inwhich the ALS IgG was effective. These observations are consistent with singlemuscle fibre experiments in which ALS IgG was effective when added to the externalsolution (Delbono et al. 1991 a, b).
DISCUSSION
T-tubule skeletal muscle Ca21 channels are characterized by heterogeneity of theirconductance states (Talvenheimo, Worley & Nelson, 1987; Ma & Coronado, 1988;Mejia-Alvarez et al. 1991). This heterogeneity has been previously described withBa2+ as charge carrier and attributed to different open states of the same DHP-sensitive Ca2+ channel (Ma & Coronado, 1988). The same heterogeneity is evidentwhen channel activity is recorded in a NaCl gradient (Coronado, 1987) and in the
Fig. 5. ALS IgG decreases DHP-sensitive Ca2+ channel activity. A, traces recorded at0 mV steady-state potential in 500-50 mM cis-trans NaCl gradient. B, traces recordedfrom the same channel after the addition of ALS IgG at a final concentration of 3 mg/mlin the trans chamber. C, diary plot of the experiment. Bay K 8644 (2 ,UM) was presentthroughout the experiment in both chambers.
113

X. MAGNVELLI AAND OTHERS
3 mg/ml ALS IgG
-2 0 2 4
Amplitude (pA)
Fig. 6. ALS IgG alters Ca2+ channel activity by reducing mean open time and channelamplitude. A, open time histograms and fitted probability density functions (Sigworth &Sine, 1987) in control (left panel) and after adding 3 mg/ml ALS IgG in the trans chamber(right panel). The fitted values in control were T, = 19 ms, A1 = 0-85 and T2 = 130 ms,A2 =0-15. In the presence of ALS IgG the fitted values were T= 1-5 ms, A, = 0-58 andT2= 10-7 ms, A2 = 0-42. B, current amplitude histograms and fitted sum of Gaussianfunctions in control (left) and with ALS IgG (right). The mean values in control were
0-99+0-35, 2-77 +0-37 and 4-14+0-16 pA. The P. values for the closed state and the otherfour peaks were 0-383, 0-008, 0-008 and 0-601, respectively. In the presence ofALS IgG thevalues were 0-67 +0-15, 1-56+0-30 and 3-82+0-35 pA. The P. values for the closed stateand the other four peaks were 0 779, 0-132, 0-084 and 0-005 respectively.
114
A
14-
12 -
10 -
8-
6-
4-
Control
/A
10-3 10-l
20-
10-
00L,0)Co
ao
n
CT-S
a)
a)0Ez
101
Time (s)
B
120
100
10'
80
60 1
40-
20-
IIl I II-2 0 2 4 6
II^ I .LL

ACTION OF ALS IgG ON (,a21 CHANNELS 11
absence of divalent cations. In this case the channel becomes selective for monovalentcations (Almers & McCleskey, 1984; Coronado & Smith, 1987) and the conductancelevels detected are greater than in a BaCl2 gradient (Coronado. 1987). Ourexperiments have shown that in a NaCl gradient at 0 mV steady-state potential thesechannels show different substates which could be resolved in three main conductancelevels from the amplitude distribution. The nature of the recorded channels has beenconfirmed by using pharmacological agents which allowed us to identify functionalCa21 channels from transverse tubule vesicles. The agonist Bay K 8644 increasedopen channel probability by prolonging the mean open time as predicted for DHP-sensitive Ca21 channels (Affolter & Coronado, 1985). The DHP antagonist nifedipinereduced channel activity by acting both on channel mean open time and amplitude(Affolter & Coronado, 1985; Talvenheimo et al. 1987; Ma & Coronado, 1988; Mejia-Alvarez et al. 1991). The amplitude distribution showed only one level in the presenceof nifedipine compared to the three amplitude levels in control conditions. The actionof nifedipine on channel amplitude has been previously reported on the samepreparation (Talvenheimo et al. 1987). These authors showed that nifedipine addedin the presence of Bay K 8644 decreased channel open probability and also causeda small reduction in unitary current amplitude. This prompted the suggestion thatnifedipine affects the conformation of the open channel in addition to reducingchannel mean open time.ALS IgG reduced the mean open time and diminished channel amplitude. The
action of ALS IgG has been compared with normal human IgG and with MG IgG totest the specificity of action, and it was found that control IgG and MG IgG did notaffect Ca21 channel activity as already reported in single fibre experiments (Delbonoet al. 1991 a, b). ALS IgG modified channel activity by acting on mean open time andchannel amplitude. The open time distribution showed a shift of time constantstowards shorter openings, resulting in a reduction of the probability of channelopenings.
Previous studies of the DHP receptor complex have shown that Ca21 channelantagonists like nitrendipine and nifedipine bind to the al subunit which contains ahigh affinity receptor site (Catterall, Seagar & Takahashi, 1988). The location of thiscomplex was initially thought to include a region of segment 6 domain IV found onthe cytoplasmic side of the Ca21 channel a1 subunit (Regulla, Schneider, Nastainezyk,Meyer & Hofmann, 1991). However, more recently it has been reported to be formedby the close association of two discontinuous regions of the a1 subunit sequence indomains III and IV that reside on the external face of the channel, near thetransmembrane pore (Nakayama, Taki, Striessnig, Glossmann, Catterall & Kanaoka,1991). ALS IgG reduces single Ca2+ channel activity in a manner similar to nifedipineby affecting the mean open time and channel amplitude. Furthermore, ALS IgG andnifedipine reduced skeletal muscle macroscopic Ca2+ currents in a similar way andwith the same voltage dependency (Delbono et al. 1991 a). This similarity in actionis interesting, in the light of potential similar domains for the action of ALS and
Fig. 7. ALS IgG affected only one side of the Ca2+ channel. A. traces recorded at 0 ml'holding potential in 500-50 mm NaCl gradient in control (left sweeps), after adding3 mg/ml ALS IgG in trans chamber (middle sweeps) and with ALS IgG in both cis andtrans chambers (right sweeps). B, diary plot of the experiment. The arrows mark theaddition of ALS IgG consecutively in both the chambers.
115

V. MAGNELLI AND OTHERS
ALS IgC trans
A.h i h.wim l' iri
ALS IgG cis + trans
woALL Ll-j16L-
12 pA
1 s
B
7000
6000
5000
4000
3000
2000
1000 -
0 200 400 600 800
Time (s) / |
T ALS IgG trans |fALS IgG cis
Fig. 7. For legend see previous page.
116
AControl
c
ci
c
c
NaCI, 500-50 mM; O mV
E
CLQ
a)0
4)-i6Lo0
C.0E
wrrrlr",M-lr,,Fr"wmF
lw
s
-.. I I..,1,
..
I 1.1- II L., I -I

ACTION OF ALS IgG ON C,a2ECHANNELS1
nifedipine on the external side of the channel. This does not exclude the possibilityof separate external domains for the binding of these two agents to the DHP-sensitive Ca2" channel. Furthermore, we cannot clearly distinguish whether ALS IgGstabilizes and increases the frequency of smaller substates or whether ALS IgG actsas a fast open channel blocker with a general reduction of channel amplitude.Further experiments will clarify this point.We have shown that ALS IgG appears to affect channel activity only when applied
to what is likely to be the extracellular side of the Ca2+ channel as has already beenshown for single mammalian skeletal muscle fibres (Delbono et al. 1991 b). Theobservation that ALS IgG modifies Ca21 channel activity suggests that pathogenicantibodies can act by altering single channel characteristics as has been proposed forthe action of Lambert-Eaton antibodies (Kim & Neher, 1988). Points that remain tobe investigated are whether ALS IgG can act on Ca2+ channels from motoneuronswhich are the main cells affected in ALS, and whether such action is selective for oneor more of the Ca2+ channel subunits. The similarity in the primary sequence betweenthe skeletal muscle DHP-sensitive Ca21 channel and various types of neuronal Ca2+channels suggests that the same epitope may be present in these channels (Hui,Ellinor, Krizanova, Wang, Diebold & Schwartz, 1991).
This study was supported by a National Institutes of Health (USA) grant R01-AR38970, bygrants from the Muscular Dystrophy Association to Enrico Stefani, MDA fellowship to OsvaldoDelbono and through a MDA ALS Center grant, by the M. H. 'Jack' Wagner Memorial Fund, theRobert C. and Helen C. Kleberg Foundation and by a grant from Cephalon, Inc.
REFERENCES
AFFOLTER, H. & CORONADO, R. (1985). Agonists Bay K-8644 and CGP-28392 open calciumchannels reconstituted from skeletal muscle transverse tubules. Biophysical Journal 48. 341-347.
ALMERS, W. & MCCLESKEY, E. W. (1984). Non-selective conductance in calcium channels of frogmuscle: calcium selectivity in a single-file pore. Journal of Physiology 353, .58.5--608.
APPEL, S. H., ENGELHARDT, J. I., GARCIA, ,J. & STEFANI, E. (1991). Immunoglobulins from animalmodels of motor neuron disease and from human amyotrophic lateral sclerosis patients passivelytransfer physiological abnormalities to the neuromuscular junction. Proceedings of the NationalAcademy of Science.s of the U7SA 88, 647-651.
APPEL, S. H. & STEFANI, E. (1991). Amyotrophic lateral sclerosis: etiology and pathogenesis. InCurrent Neurology, ed. APPEL, S. H., pp. 287-310. Mosby Year Book Inc. St Louis, MO, USA.
CATTERALL, W. A., SEAGAR, M. J. & TAKAHASHI, M. (1988). Molecular properties of dihydro-pyridine-sensitive calcium channels in skeletal muscle. Journal of Biological Chemistry 263,3535-3538.
COLQIUHOUN, D. & SIGWORTH, F. J. (1983). Fitting and statistical analysis of single-channelrecords. In Single-ChannelRecording, ed. SAKMANN, B. & NEHER, E., pp. 191-263. Plenum Press,New York.
CORONADO, R. (1987). Planar bilayer reconstitution of calcium channels: lipid effects on single-channel kinetics. Circulation Research 61, 146-52.
CORONADO, R. & AFFOLTER, H. (1986). Insulation of the conduction pathway of muscle transversetubule c(alcium channels from the surface charge of bilayer phospholipid. Journal of GeneralPhysiology 87, 933-953.
CORONADO, R. & SMITH, J. S. (1987). Monovalent ion current through single calcium channels ofskeletal muscle transverse tubules. Biophysical Journal 51, 497-502.
DELBONO, O., GARCIA, FJ., APPEL, S. H. & STEFANI, E. (1991 a). IgG from amyotrophic lateralsclerosis affects tubular calcium channels of skeletal muscle. American Journal of Physiology 260,C1347-1351.
.5 PHY 461
117

11.I.JAGNELLI AND OTHERS
DELBONO, O., GARCIA, J., APPEL, S. H. & STEFANI, E. (1991b). Calcium current and chargemovement of mammalian skeletal muscle: action of amyotrophic lateral sclerosis immuno-globulins. Journal of Physiology 444, 723-742.
ENGELHARDT, J. I. & APPEL, S. H. (1990). IgG reactivity in the spinal cord and motor cortex inamyotrophic lateral sclerosis. Archives of Neurology 47, 1210-1216.
ENGELHARDT, J. I., APPEL, S. H. & KILLIAN, J. M. (1989). Experimental autoimmune motoneurondisease. Annals of Neurology 26, 368-376.
ENGELHARDT, J. I., APPEL, S. H. & KILLIAN, J. M. (1990). Motor neuron destruction in guinea pigsimmunized with bovine spinal cord ventral horn homogenate: experimental autoimmune graymatter disease. Journal of Neuroimmunology 27, 21-31.
GARCIA, J., ENGELHARDT, J. I., APPEL, S. H. & STEFANI, E. (1990). Increased mepp frequency asan early sign of experimental immune mediated motor neuron disease. Annals of Neurology 28,329-334.
HAMILTON, S. L., MEJIA-ALVAREZ, R., FILL, M., HAWKES, M. J., BRUSH, K. L., SCHILLING, W. P.& STEFANI, E. (1989). [3H]PN299-119 and [3H]ryanodine binding and reconstitution of ionchannel activity with skeletal muscle membranes. Analytical Biochemistry 183, 31-41.
HuI, A., ELLINOR, P. T., KRIZANOVA, O., WANG, J. J., DIEBOLD, R. J. & SCHWARTZ, A. (1991).Molecular cloning of multiple subtypes of a novel rat brain isoform of the a, subunit of thevoltage-dependent calcium channel. Neuron 7, 35-44.
KIM, Y. I. & NEHER, E. (1988). IgG from patients with Lambert-Eaton syndrome blocks voltage-dependent calcium channels. Science 239, 405-408.
MA, J. & CORONADO, R. (1988). Heterogeneity of conductance states in calcium channels of skeletalmuscle. Biophysical Journal 53, 387-395.
MEJIA-ALVAREZ, R., FILL, M. & STEFANI, E. (1991). Voltage-dependent inactivation of T-tubularskeletal calcium channels in planar lipid bilayers. Journal of General Physiology 97, 393-412.
NAKAYAMA, H., TAKI, M., STRIESSNIG, J., GLOSSMANN, H., CATTERALL, W. A. & KANAOKA, Y.(1991). Identification of 1,4-dihydropyridine binding regions within the a, subunit of skeletalmuscle Ca2+ channels by photoaffinity labeling with diazipine. Proceedings of the NationalAcademy of Sciences of the USA 88, 9203-9207.
PELZER, D., GRANT, A. O., CAVALIE, A., PELZER, S., SIEBER, M., HOFMANN, F. & TRAUTWEIN, W.(1989). Calcium channels reconstituted from the skeletal muscle dihydropyridine receptorprotein complex and its a peptide subunit in lipid bilayers. Annals of the New York Academy ofSciences 560, 138-154.
REGULLA, S., SCHNEIDER, T., NASTAINCZYK, W., MEYER, H. E. & HOFMANN, F. (1991).Identification of the site of interaction of the dihydropyridine channel blockers nitrendipine andazidopine with the calcium-channel al subunit. EMBO Journal 10, 45--49.
SIGWORTH, F. J. & SINE, S. M. (1987). Data transformations for improved display and fitting ofsingle-channel dwell time histograms. Biophysical Journal 52, 1047-1054.
TALVENHEIMO, J. A., WORLEY, J. F. & NELSON, M. T. (1987). Heterogeneity of calcium channelsfrom a purified dihydropyridine receptor preparation. Biophysical Journal 52, 891-899.
VILVEN, J. & CORONADO, R. (1988). Opening of dihydropyridine calcium channels in skeletalmuscle membranes by inositol trisphosphate. Nature 336, 587-589.
118