document
TRANSCRIPT

938 NATURE MEDICINE • VOLUME 5 • NUMBER 8 • AUGUST 1999
ARTICLES
Tumor-associated antigens that can be recognized by the im-mune system include the MAGE-family, p53, MUC-1, HER2/neuand p21ras (refs. 1–6). Despite their expression of these dis-tinct antigens, tumor elimination by the immune system isoften inefficient. Postulated mechanisms include insufficientexpression of co-stimulatory or adhesion molecules by tumorcells, or defective processing and presentation of antigens ontheir cell surfaces7–10. Tumor cells may also evade immune at-tack by expressing CD95 (APO-1/Fas) ligand or other mole-cules that induce apoptosis in activated T cells11–13. Here wedescribe RCAS1 (receptor-binding cancer antigen expressedon SiSo cells), a membrane molecule expressed on human can-cer cells. RCAS1 acts as a ligand for a putative receptor presenton various human cell lines and normal peripheral lympho-cytes such as T, B and NK cells. The receptor expression wasenhanced by activation of the lymphocytes. RCAS1 inhibitedthe in vitro growth of receptor-expressing cells and inducedapoptotic cell death. Given these results, tumor cells mayevade immune surveillance by expression of RCAS1, whichwould suppress clonal expansion and induce apoptosis inRCAS1 receptor-positive immune cells.
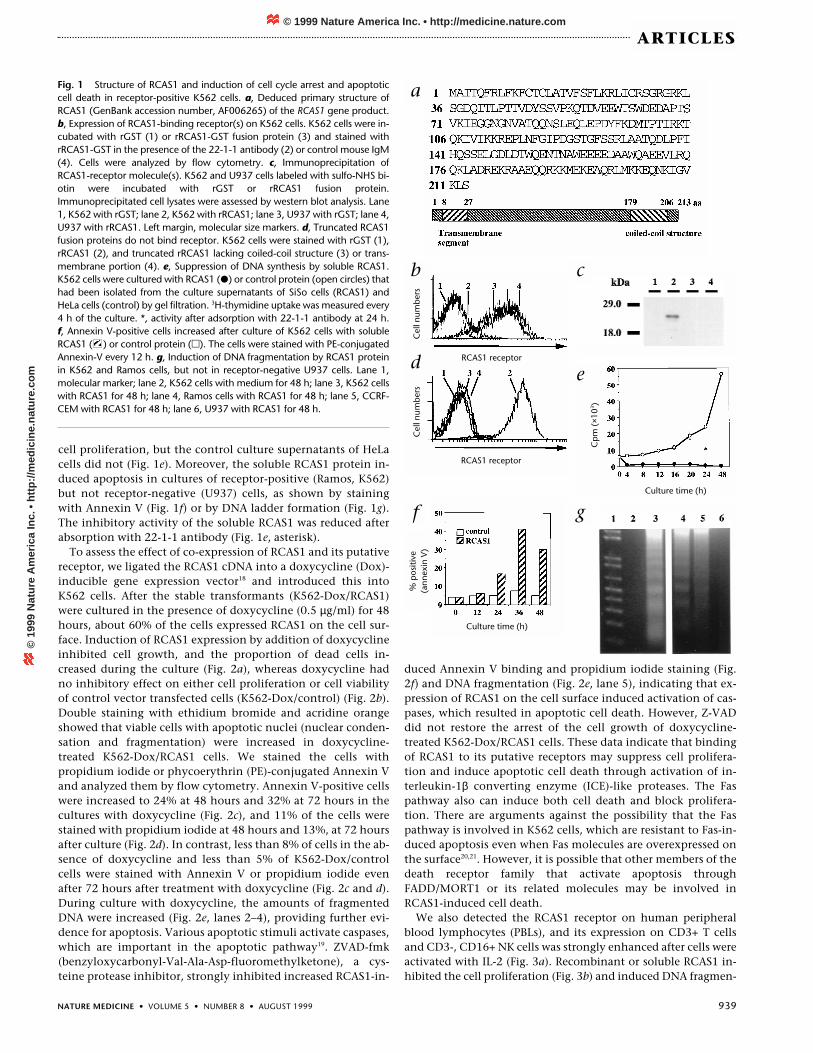
We used a mouse monoclonal antibody (22-1-1 antibody; ref.14) against a human uterine adenocarcinoma cell line, SiSo (ref.15). Immunohistochemical studies have shown that the antigenrecognized by 22-1-1 antibody is expressed on uterine and ovar-ian carcinomas and is especially strongly expressed in invasivecancers14,16. We have now isolated a cDNA encoding the antigenrecognized by the 22-1-1 antibody and called it RCAS1 (recep-tor-binding cancer antigen expressed on SiSo cells) (Fig. 1a). ThecDNA encoding RCAS1 contains a 5′ untranslated region of 242nucleotides, a coding region of 639 nucleotides (213 aminoacids), and a 3′ untranslated region of 179 nucleotides. The ATGinitiation codon is in a standard Kozak consensus sequence(CCCACCATG). RCAS1 has an N-terminal transmembrane seg-ment (8–27 amino acids), as predicted by TMbase (ref. 17), and acoiled-coil structure in the C-terminal portion (179–206 aminoacids), indicating that RCAS1 is a type II membrane protein ableto form oligomers through the coiled-coil structure. In fact, 22-1-1 antibody immunoprecipitates of mouse L cells transfectedwith the RCAS1 cDNA produced 40-kDa and 80-kDa bands, in-dicating the presence of RCAS1 homodimers. The coiled-coilstructure seemed to be essential for homologous complex for-mation, as deletion of this region diminished dimerization(data not shown).
Based on RNA blot analysis, the RCAS1 gene is expressed inovary, testis, prostate, thymus, muscle and heart.Immunohistochemically, however, RCAS1 protein has not
been detected in normal ovary tissues or any of the other aboveorgans15,16. Neither transcript nor protein was detected in smallintestine, colon, lymph nodes or peripheral blood lymphocytes(data not shown). To determine the biological function of theRCAS1 antigen, we synthesized recombinant RCAS1 (rRCAS1)protein as a GST fusion protein. The affinity-purifiedrRCAS1–GST fusion protein bound to the surface of manyhuman and mouse cell lines, such as K562 (human chronicmyelogeneous leukemia), CCRF-CEM (human T lymphoblast),Ramos (Burkitt lymphoma), WI-38 (human diploid cell line),IMR-90 (human diploid cell line), 293 (human transformed pri-mary embryonal kidney cell line), mouse L cells and NIH3T3cells. One exception was U937, a human histiocytic leukemiacell line. Addition of 22-1-1 antibody inhibited the binding ofrRCAS1 to those cell lines (Fig 1b). Immunoprecipitation in re-ducing conditions with rRCAS1 specifically identified a mole-cule with a molecular weight of 25 kDa as an rRCAS1-bindingprotein from surface-biotinylated K562 cells, but not fromU937 cells (Fig. 1c). These data indicate the presence of recep-tor-like molecule(s) for RCAS1 on various cell lines except forU937. Truncated RCAS1 protein lacking the C-terminal coiled-coil structure did not bind to the receptor-positive cells (Fig1d). Beads coated with this truncated protein failed to formrosettes with K562 cells. In contrast, beads coated with a mu-tant protein lacking the transmembrane domain but contain-ing the coiled-coil portion were able to form rosettes withreceptor-expressing K562 cells (data not shown). This result in-dicates that formation of homologous RCAS1 complex may berequired for maintenance of binding activity, and that thecoiled-coil region may be involved in the binding of RCAS1 toreceptor-positive cells through the formation of the complex.Addition of the rRCAS1–GST fusion protein to K562 cells in-duced tyrosine phosphorylation of several cytoplasmic pro-teins within 5 mintues of incubation, but recombinant GST(rGST) protein and truncated rRCAS1 lacking the coiled-coilstructure did not induce any phosphorylation (data notshown). These data also indicate the existence of a signal-trans-ducing RCAS1 receptor on various cell types. The expression ofRCAS1 and its receptor molecule were mutually exclusive.None of the human and mouse cells simultaneously expressedboth molecules. K562 cells expressed the receptor molecule,but not RCAS1 on the cell surface. Normal cells such ashematopoietic cells and lymphocytes expressed only receptormolecules.
SiSo cells secreted RCAS1 antigen into the culture medium,and the soluble RCAS1 protein was partially purified by gel fil-tration from the culture supernatants. Addition of solubleRCAS1 protein into cultures of K562 cells strongly suppressed
Inhibition of cell growth and induction of apoptotic celldeath by the human tumor-associated antigen RCAS1
MANABU NAKASHIMA1, KENZO SONODA2 & TAKESHI WATANABE1
1Department of Molecular Immunology, Medical Institute of Bioregulation, and 2Department of Gynecology andObstetrics, Faculty of Medicine, Kyushu University, 3-1-1 Maidashi, Higashi-ku, Fukuoka 812-8582, Japan
Correspondence should be addressed to T.W.; email: [email protected]
© 1999 Nature America Inc. • http://medicine.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://m
edic
ine.
nat
ure
.co
m
NATURE MEDICINE • VOLUME 5 • NUMBER 8 • AUGUST 1999 939
ARTICLES
cell proliferation, but the control culture supernatants of HeLacells did not (Fig. 1e). Moreover, the soluble RCAS1 protein in-duced apoptosis in cultures of receptor-positive (Ramos, K562)but not receptor-negative (U937) cells, as shown by stainingwith Annexin V (Fig. 1f) or by DNA ladder formation (Fig. 1g).The inhibitory activity of the soluble RCAS1 was reduced afterabsorption with 22-1-1 antibody (Fig. 1e, asterisk).
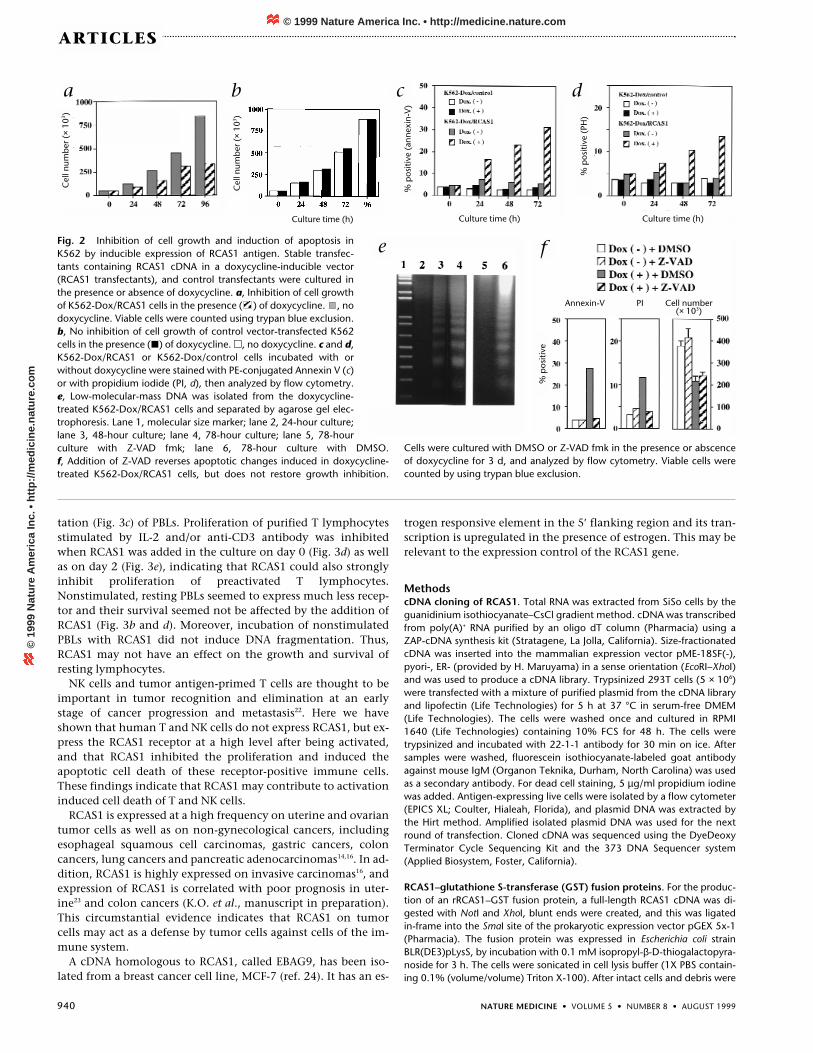
To assess the effect of co-expression of RCAS1 and its putativereceptor, we ligated the RCAS1 cDNA into a doxycycline (Dox)-inducible gene expression vector18 and introduced this intoK562 cells. After the stable transformants (K562-Dox/RCAS1)were cultured in the presence of doxycycline (0.5 µg/ml) for 48hours, about 60% of the cells expressed RCAS1 on the cell sur-face. Induction of RCAS1 expression by addition of doxycyclineinhibited cell growth, and the proportion of dead cells in-creased during the culture (Fig. 2a), whereas doxycycline hadno inhibitory effect on either cell proliferation or cell viabilityof control vector transfected cells (K562-Dox/control) (Fig. 2b).Double staining with ethidium bromide and acridine orangeshowed that viable cells with apoptotic nuclei (nuclear conden-sation and fragmentation) were increased in doxycycline-treated K562-Dox/RCAS1 cells. We stained the cells withpropidium iodide or phycoerythrin (PE)-conjugated Annexin Vand analyzed them by flow cytometry. Annexin V-positive cellswere increased to 24% at 48 hours and 32% at 72 hours in thecultures with doxycycline (Fig. 2c), and 11% of the cells werestained with propidium iodide at 48 hours and 13%, at 72 hoursafter culture (Fig. 2d). In contrast, less than 8% of cells in the ab-sence of doxycycline and less than 5% of K562-Dox/controlcells were stained with Annexin V or propidium iodide evenafter 72 hours after treatment with doxycycline (Fig. 2c and d).During culture with doxycycline, the amounts of fragmentedDNA were increased (Fig. 2e, lanes 2–4), providing further evi-dence for apoptosis. Various apoptotic stimuli activate caspases,which are important in the apoptotic pathway19. ZVAD-fmk(benzyloxycarbonyl-Val-Ala-Asp-fluoromethylketone), a cys-teine protease inhibitor, strongly inhibited increased RCAS1-in-
duced Annexin V binding and propidium iodide staining (Fig.2f) and DNA fragmentation (Fig. 2e, lane 5), indicating that ex-pression of RCAS1 on the cell surface induced activation of cas-pases, which resulted in apoptotic cell death. However, Z-VADdid not restore the arrest of the cell growth of doxycycline-treated K562-Dox/RCAS1 cells. These data indicate that bindingof RCAS1 to its putative receptors may suppress cell prolifera-tion and induce apoptotic cell death through activation of in-terleukin-1β converting enzyme (ICE)-like proteases. The Faspathway also can induce both cell death and block prolifera-tion. There are arguments against the possibility that the Faspathway is involved in K562 cells, which are resistant to Fas-in-duced apoptosis even when Fas molecules are overexpressed onthe surface20,21. However, it is possible that other members of thedeath receptor family that activate apoptosis throughFADD/MORT1 or its related molecules may be involved inRCAS1-induced cell death.
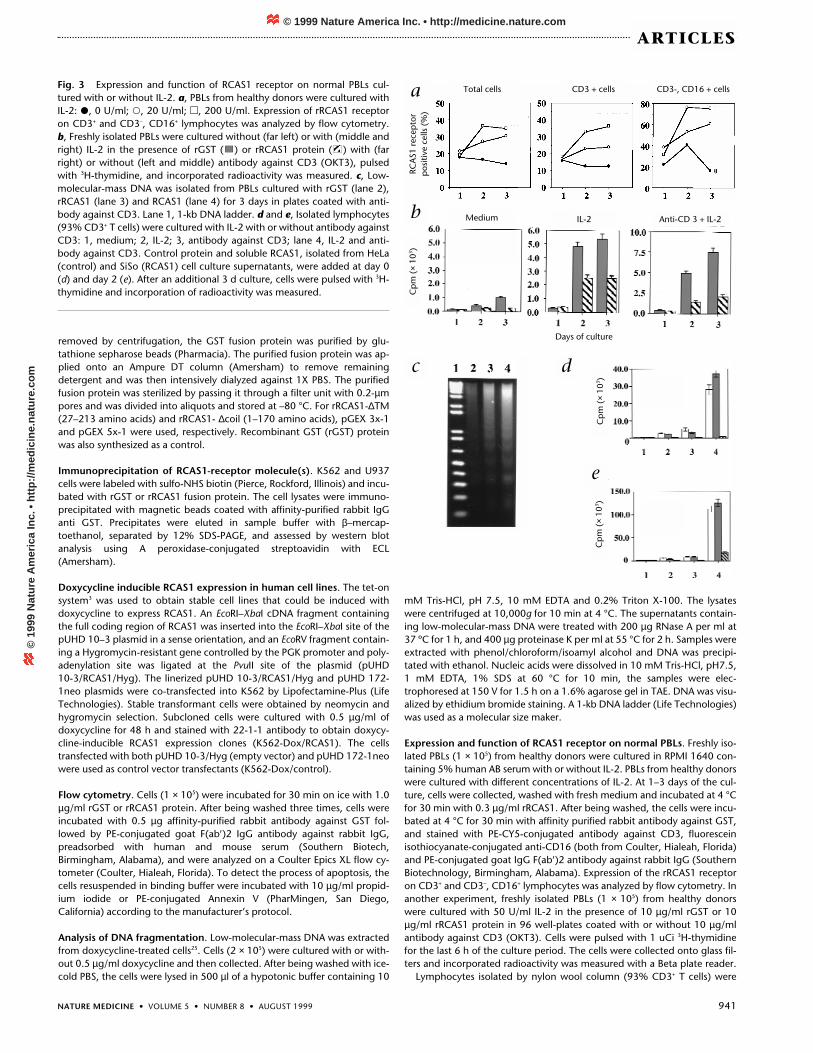
We also detected the RCAS1 receptor on human peripheralblood lymphocytes (PBLs), and its expression on CD3+ T cellsand CD3-, CD16+ NK cells was strongly enhanced after cells wereactivated with IL-2 (Fig. 3a). Recombinant or soluble RCAS1 in-hibited the cell proliferation (Fig. 3b) and induced DNA fragmen-
Fig. 1 Structure of RCAS1 and induction of cell cycle arrest and apoptoticcell death in receptor-positive K562 cells. a, Deduced primary structure ofRCAS1 (GenBank accession number, AF006265) of the RCAS1 gene product.b, Expression of RCAS1-binding receptor(s) on K562 cells. K562 cells were in-cubated with rGST (1) or rRCAS1-GST fusion protein (3) and stained withrRCAS1-GST in the presence of the 22-1-1 antibody (2) or control mouse IgM(4). Cells were analyzed by flow cytometry. c, Immunoprecipitation ofRCAS1-receptor molecule(s). K562 and U937 cells labeled with sulfo-NHS bi-otin were incubated with rGST or rRCAS1 fusion protein.Immunoprecipitated cell lysates were assessed by western blot analysis. Lane1, K562 with rGST; lane 2, K562 with rRCAS1; lane 3, U937 with rGST; lane 4,U937 with rRCAS1. Left margin, molecular size markers. d, Truncated RCAS1fusion proteins do not bind receptor. K562 cells were stained with rGST (1),rRCAS1 (2), and truncated rRCAS1 lacking coiled-coil structure (3) or trans-membrane portion (4). e, Suppression of DNA synthesis by soluble RCAS1.K562 cells were cultured with RCAS1 (�) or control protein (open circles) thathad been isolated from the culture supernatants of SiSo cells (RCAS1) andHeLa cells (control) by gel filtration. 3H-thymidine uptake was measured every4 h of the culture. *, activity after adsorption with 22-1-1 antibody at 24 h. f, Annexin V-positive cells increased after culture of K562 cells with solubleRCAS1 ( ) or control protein (�). The cells were stained with PE-conjugatedAnnexin-V every 12 h. g, Induction of DNA fragmentation by RCAS1 proteinin K562 and Ramos cells, but not in receptor-negative U937 cells. Lane 1,molecular marker; lane 2, K562 cells with medium for 48 h; lane 3, K562 cellswith RCAS1 for 48 h; lane 4, Ramos cells with RCAS1 for 48 h; lane 5, CCRF-CEM with RCAS1 for 48 h; lane 6, U937 with RCAS1 for 48 h.
a
b c
de
f gCulture time (h)
Culture time (h)
RCAS1 receptor
RCAS1 receptor
Cp
m (
×103 )
% p
ositi
ve(a
nnex
in V
)C
ell n
umb
ers
Cel
l num
ber
s
© 1999 Nature America Inc. • http://medicine.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://m
edic
ine.
nat
ure
.co
m
940 NATURE MEDICINE • VOLUME 5 • NUMBER 8 • AUGUST 1999
ARTICLES
Fig. 2 Inhibition of cell growth and induction of apoptosis inK562 by inducible expression of RCAS1 antigen. Stable transfec-tants containing RCAS1 cDNA in a doxycycline-inducible vector(RCAS1 transfectants), and control transfectants were cultured inthe presence or absence of doxycycline. a, Inhibition of cell growthof K562-Dox/RCAS1 cells in the presence ( ) of doxycycline. , nodoxycycline. Viable cells were counted using trypan blue exclusion.b, No inhibition of cell growth of control vector-transfected K562cells in the presence (�) of doxycycline. �, no doxycycline. c and d,K562-Dox/RCAS1 or K562-Dox/control cells incubated with orwithout doxycycline were stained with PE-conjugated Annexin V (c)or with propidium iodide (PI, d), then analyzed by flow cytometry.e, Low-molecular-mass DNA was isolated from the doxycycline-treated K562-Dox/RCAS1 cells and separated by agarose gel elec-trophoresis. Lane 1, molecular size marker; lane 2, 24-hour culture;lane 3, 48-hour culture; lane 4, 78-hour culture; lane 5, 78-hourculture with Z-VAD fmk; lane 6, 78-hour culture with DMSO. f, Addition of Z-VAD reverses apoptotic changes induced in doxycycline-treated K562-Dox/RCAS1 cells, but does not restore growth inhibition.
tation (Fig. 3c) of PBLs. Proliferation of purified T lymphocytesstimulated by IL-2 and/or anti-CD3 antibody was inhibitedwhen RCAS1 was added in the culture on day 0 (Fig. 3d) as wellas on day 2 (Fig. 3e), indicating that RCAS1 could also stronglyinhibit proliferation of preactivated T lymphocytes.Nonstimulated, resting PBLs seemed to express much less recep-tor and their survival seemed not be affected by the addition ofRCAS1 (Fig. 3b and d). Moreover, incubation of nonstimulatedPBLs with RCAS1 did not induce DNA fragmentation. Thus,RCAS1 may not have an effect on the growth and survival ofresting lymphocytes.
NK cells and tumor antigen-primed T cells are thought to beimportant in tumor recognition and elimination at an earlystage of cancer progression and metastasis22. Here we haveshown that human T and NK cells do not express RCAS1, but ex-press the RCAS1 receptor at a high level after being activated,and that RCAS1 inhibited the proliferation and induced theapoptotic cell death of these receptor-positive immune cells.These findings indicate that RCAS1 may contribute to activationinduced cell death of T and NK cells.
RCAS1 is expressed at a high frequency on uterine and ovariantumor cells as well as on non-gynecological cancers, includingesophageal squamous cell carcinomas, gastric cancers, coloncancers, lung cancers and pancreatic adenocarcinomas14,16. In ad-dition, RCAS1 is highly expressed on invasive carcinomas16, andexpression of RCAS1 is correlated with poor prognosis in uter-ine23 and colon cancers (K.O. et al., manuscript in preparation).This circumstantial evidence indicates that RCAS1 on tumorcells may act as a defense by tumor cells against cells of the im-mune system.
A cDNA homologous to RCAS1, called EBAG9, has been iso-lated from a breast cancer cell line, MCF-7 (ref. 24). It has an es-
trogen responsive element in the 5′ flanking region and its tran-scription is upregulated in the presence of estrogen. This may berelevant to the expression control of the RCAS1 gene.
MethodscDNA cloning of RCAS1. Total RNA was extracted from SiSo cells by theguanidinium isothiocyanate–CsCl gradient method. cDNA was transcribedfrom poly(A)+ RNA purified by an oligo dT column (Pharmacia) using aZAP-cDNA synthesis kit (Stratagene, La Jolla, California). Size-fractionatedcDNA was inserted into the mammalian expression vector pME-18SF(-),pyori-, ER- (provided by H. Maruyama) in a sense orientation (EcoRI–XhoI)and was used to produce a cDNA library. Trypsinized 293T cells (5 × 106)were transfected with a mixture of purified plasmid from the cDNA libraryand lipofectin (Life Technologies) for 5 h at 37 °C in serum-free DMEM(Life Technologies). The cells were washed once and cultured in RPMI1640 (Life Technologies) containing 10% FCS for 48 h. The cells weretrypsinized and incubated with 22-1-1 antibody for 30 min on ice. Aftersamples were washed, fluorescein isothiocyanate-labeled goat antibodyagainst mouse IgM (Organon Teknika, Durham, North Carolina) was usedas a secondary antibody. For dead cell staining, 5 µg/ml propidium iodinewas added. Antigen-expressing live cells were isolated by a flow cytometer(EPICS XL; Coulter, Hialeah, Florida), and plasmid DNA was extracted bythe Hirt method. Amplified isolated plasmid DNA was used for the nextround of transfection. Cloned cDNA was sequenced using the DyeDeoxyTerminator Cycle Sequencing Kit and the 373 DNA Sequencer system(Applied Biosystem, Foster, California).
RCAS1–glutathione S-transferase (GST) fusion proteins. For the produc-tion of an rRCAS1–GST fusion protein, a full-length RCAS1 cDNA was di-gested with NotI and XhoI, blunt ends were created, and this was ligatedin-frame into the SmaI site of the prokaryotic expression vector pGEX 5x-1(Pharmacia). The fusion protein was expressed in Escherichia coli strainBLR(DE3)pLysS, by incubation with 0.1 mM isopropyl-β-D-thiogalactopyra-noside for 3 h. The cells were sonicated in cell lysis buffer (1X PBS contain-ing 0.1% (volume/volume) Triton X-100). After intact cells and debris were
Cells were cultured with DMSO or Z-VAD fmk in the presence or abscenceof doxycycline for 3 d, and analyzed by flow cytometry. Viable cells werecounted by using trypan blue exclusion.
Cel
l num
ber
(×
103 )
Cel
l num
ber
(×
103 )
Culture time (h) Culture time (h) Culture time (h)
% p
ositi
ve (
anne
xin-
V)
% p
ositi
ve (
PH)
Annexin-V PI Cell number(× 103)
a b c d
e f
% p
ositi
ve
© 1999 Nature America Inc. • http://medicine.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://m
edic
ine.
nat
ure
.co
m
NATURE MEDICINE • VOLUME 5 • NUMBER 8 • AUGUST 1999 941
ARTICLES
removed by centrifugation, the GST fusion protein was purified by glu-tathione sepharose beads (Pharmacia). The purified fusion protein was ap-plied onto an Ampure DT column (Amersham) to remove remainingdetergent and was then intensively dialyzed against 1X PBS. The purifiedfusion protein was sterilized by passing it through a filter unit with 0.2-µmpores and was divided into aliquots and stored at –80 °C. For rRCAS1-∆TM(27–213 amino acids) and rRCAS1- ∆coil (1–170 amino acids), pGEX 3x-1and pGEX 5x-1 were used, respectively. Recombinant GST (rGST) proteinwas also synthesized as a control.
Immunoprecipitation of RCAS1-receptor molecule(s). K562 and U937cells were labeled with sulfo-NHS biotin (Pierce, Rockford, Illinois) and incu-bated with rGST or rRCAS1 fusion protein. The cell lysates were immuno-precipitated with magnetic beads coated with affinity-purified rabbit IgGanti GST. Precipitates were eluted in sample buffer with β–mercap-toethanol, separated by 12% SDS-PAGE, and assessed by western blotanalysis using A peroxidase-conjugated streptoavidin with ECL(Amersham).
Doxycycline inducible RCAS1 expression in human cell lines. The tet-onsystem3 was used to obtain stable cell lines that could be induced withdoxycycline to express RCAS1. An EcoRI–XbaI cDNA fragment containingthe full coding region of RCAS1 was inserted into the EcoRI–XbaI site of thepUHD 10–3 plasmid in a sense orientation, and an EcoRV fragment contain-ing a Hygromycin-resistant gene controlled by the PGK promoter and poly-adenylation site was ligated at the PvuII site of the plasmid (pUHD10-3/RCAS1/Hyg). The linerized pUHD 10-3/RCAS1/Hyg and pUHD 172-1neo plasmids were co-transfected into K562 by Lipofectamine-Plus (LifeTechnologies). Stable transformant cells were obtained by neomycin andhygromycin selection. Subcloned cells were cultured with 0.5 µg/ml ofdoxycycline for 48 h and stained with 22-1-1 antibody to obtain doxycy-cline-inducible RCAS1 expression clones (K562-Dox/RCAS1). The cellstransfected with both pUHD 10-3/Hyg (empty vector) and pUHD 172-1neowere used as control vector transfectants (K562-Dox/control).
Flow cytometry. Cells (1 × 105) were incubated for 30 min on ice with 1.0µg/ml rGST or rRCAS1 protein. After being washed three times, cells wereincubated with 0.5 µg affinity-purified rabbit antibody against GST fol-lowed by PE-conjugated goat F(ab′)2 IgG antibody against rabbit IgG,preadsorbed with human and mouse serum (Southern Biotech,Birmingham, Alabama), and were analyzed on a Coulter Epics XL flow cy-tometer (Coulter, Hialeah, Florida). To detect the process of apoptosis, thecells resuspended in binding buffer were incubated with 10 µg/ml propid-ium iodide or PE-conjugated Annexin V (PharMingen, San Diego,California) according to the manufacturer’s protocol.
Analysis of DNA fragmentation. Low-molecular-mass DNA was extractedfrom doxycycline-treated cells25. Cells (2 × 105) were cultured with or with-out 0.5 µg/ml doxycycline and then collected. After being washed with ice-cold PBS, the cells were lysed in 500 µl of a hypotonic buffer containing 10
mM Tris-HCl, pH 7.5, 10 mM EDTA and 0.2% Triton X-100. The lysateswere centrifuged at 10,000g for 10 min at 4 °C. The supernatants contain-ing low-molecular-mass DNA were treated with 200 µg RNase A per ml at37 ºC for 1 h, and 400 µg proteinase K per ml at 55 °C for 2 h. Samples wereextracted with phenol/chloroform/isoamyl alcohol and DNA was precipi-tated with ethanol. Nucleic acids were dissolved in 10 mM Tris-HCl, pH7.5,1 mM EDTA, 1% SDS at 60 °C for 10 min, the samples were elec-trophoresed at 150 V for 1.5 h on a 1.6% agarose gel in TAE. DNA was visu-alized by ethidium bromide staining. A 1-kb DNA ladder (Life Technologies)was used as a molecular size maker.
Expression and function of RCAS1 receptor on normal PBLs. Freshly iso-lated PBLs (1 × 105) from healthy donors were cultured in RPMI 1640 con-taining 5% human AB serum with or without IL-2. PBLs from healthy donorswere cultured with different concentrations of IL-2. At 1–3 days of the cul-ture, cells were collected, washed with fresh medium and incubated at 4 °Cfor 30 min with 0.3 µg/ml rRCAS1. After being washed, the cells were incu-bated at 4 °C for 30 min with affinity purified rabbit antibody against GST,and stained with PE-CY5-conjugated antibody against CD3, fluoresceinisothiocyanate-conjugated anti-CD16 (both from Coulter, Hialeah, Florida)and PE-conjugated goat IgG F(ab′)2 antibody against rabbit IgG (SouthernBiotechnology, Birmingham, Alabama). Expression of the rRCAS1 receptoron CD3+ and CD3–, CD16+ lymphocytes was analyzed by flow cytometry. Inanother experiment, freshly isolated PBLs (1 × 105) from healthy donorswere cultured with 50 U/ml IL-2 in the presence of 10 µg/ml rGST or 10µg/ml rRCAS1 protein in 96 well-plates coated with or without 10 µg/mlantibody against CD3 (OKT3). Cells were pulsed with 1 uCi 3H-thymidinefor the last 6 h of the culture period. The cells were collected onto glass fil-ters and incorporated radioactivity was measured with a Beta plate reader.
Lymphocytes isolated by nylon wool column (93% CD3+ T cells) were
Fig. 3 Expression and function of RCAS1 receptor on normal PBLs cul-tured with or without IL-2. a, PBLs from healthy donors were cultured withIL-2: �, 0 U/ml; �, 20 U/ml; �, 200 U/ml. Expression of rRCAS1 receptoron CD3+ and CD3–, CD16+ lymphocytes was analyzed by flow cytometry. b, Freshly isolated PBLs were cultured without (far left) or with (middle andright) IL-2 in the presence of rGST ( ) or rRCAS1 protein ( ) with (farright) or without (left and middle) antibody against CD3 (OKT3), pulsedwith 3H-thymidine, and incorporated radioactivity was measured. c, Low-molecular-mass DNA was isolated from PBLs cultured with rGST (lane 2),rRCAS1 (lane 3) and RCAS1 (lane 4) for 3 days in plates coated with anti-body against CD3. Lane 1, 1-kb DNA ladder. d and e, Isolated lymphocytes(93% CD3+ T cells) were cultured with IL-2 with or without antibody againstCD3: 1, medium; 2, IL-2; 3, antibody against CD3; lane 4, IL-2 and anti-body against CD3. Control protein and soluble RCAS1, isolated from HeLa(control) and SiSo (RCAS1) cell culture supernatants, were added at day 0(d) and day 2 (e). After an additional 3 d culture, cells were pulsed with 3H-thymidine and incorporation of radioactivity was measured.
Total cells CD3 + cells CD3-, CD16 + cells
RCA
S1 r
ecep
tor
pos
itive
cel
ls (
%)
a
b
c d
e
Medium IL-2 Anti-CD 3 + IL-2
Days of culture
Cp
m (
×10
3 )
Cp
m (
×10
3 )C
pm
(×
103 )
© 1999 Nature America Inc. • http://medicine.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://m
edic
ine.
nat
ure
.co
m

942 NATURE MEDICINE • VOLUME 5 • NUMBER 8 • AUGUST 1999
ARTICLES
cultured with IL-2 in 96 well-plates coated with or without antibody againstCD3 antibody. Control protein and soluble RCAS1, isolated from HeLa andSiSo cell culture supernatants, respectively, were added at day 0 and day 2.After an additional 3 days of culture, the cells were pulsed with 3H-thymi-dine for the last 8 h of culture.
AcknowledgmentsWe thank P.D. Burrows and T.H. Bester for their critical reading andcomments. We also thank H. Katsuta for his help. The work is supported by theGrant-in-Aid for Cancer Reserch from The Ministry of Education, Science andCulture of Japan (Grant number: 10153245).
RECEIVED 12 APRIL; ACCEPTED 9 JUNE 1999
1. van de Bruggen, P. et al. A gene encoding an antigen recognized by cytolytic Tlymphocytes on a human melanoma. Science 254, 1643–1647 (1991).
2. Melief, C.J. & Kast, W.M. T-cell immunotherapy of cancer. Res. Immunol. 142,425–429 (1991).
3. Barnd, D.L., Lan, M.S., Metzgar, R.S. & Finn, O.J. Specific, major histocompatibilitycomplex-unrestricted recognition of tumor-associated mucins by human cytotoxicT cells. Proc. Natl. Acad. Sci. USA 86, 7159–7163 (1989).
4. Peiper, M. et al. The HER2/neu-derived peptide p654-662 is a tumor-associatedantigen in human pancreatic cancer recognized by cytotoxic T lymphocytes. Eur. J.Immunol. 27, 1115–1123 (1997).
5. Van den Eynde, B.J. & Van der Bruggen, P. T cell defined tumor antigens. Curr.Opin. Immunol. 9, 684–693 (1997).
6. Sahin, U. Tureci, O. & Pfreundschuh, M. Serological identification of human tumorantigens. Curr. Opin. Immunol. 9, 709–716 (1997).
7. Becker, J.C. et al. Tumor escape mechanisms from immunosurveillance: inductionof unresponsiveness in a specific MHC-restricted CD4+ human T cell clone by theautologous MHC class II+ melanoma. Int. Immunol. 5, 1501–1508 (1993).
8. Runger, T.M., Klein, C.E., Becker, J.C. & Brocker, E.B. The role of genetic instability,adhesion, cell motility, and immune escape mechanisms in melanoma progression.Curr. Opin. Oncol. 6, 188–196 (1994).
9. Edward, M. Integrins and other adhesion molecules involved in melanocytic tumorprogression. Curr. Opin. Oncol. 7, 185–191 (1995).
10. Ferrone, S. & Marincola, F.M. Loss of HLA class I antigens by melanoma cells: mole-cular mechanisms, functional significance and clinical relevance. Immunol. Today16, 487–494 (1995).
11. Susanne, S. et al. Lymphocyte apoptosis induced by CD95(APO-1/Fas) ligand-ex-pressing tumor cells - A mechanism of immune evasion? Nature Med. 2, 1361–1366(1996).
12. Claude, D.G. et al. Breast cancer-associated antigen, DF3/MUC1, induces apoptosisof activated human T cells. Nature Med. 2, 1367–1370 (1996).
13. Hahne, M. et al. Melanoma cell expression of Fas(Apo-1/CD95) ligand: Implicationsfor tumor immune escape. Science 274, 1363–1366 (1996).
14. Sonoda, K. et al. A novel tumor-associated antigen expressed in human uterine andovarian carcinomas. Cancer 77, 1501–1509 (1996).
15. Sonoda, K. et al. Establishment of a new human uterine cervical adenocarcinomacell line, SiSo, and its reactivity to anti-cancer reagents. Int. J. Oncol. 6, 1099–1104(1995).
16. Sonoda, K. et al. Tumor-associated antigen 22-1-1 expression in the uterine cervicalsquamous neoplasias. Clinical Cancer Research 4, 1517–1520 (1998).
17. K. Hofmann and W. Stoffel. TMBASE – A database of membrane spanning proteinsegments. Biol. Chem. Hoppe-Seyler 374, 166 (1993).
18. Gossen, M. et al. Transcriptional activation by tetracyclines in mammalian cells.Science 268, 1766–1769 (1995).
19. Henkart, P.A. ICE family proteases: mediators of all apoptotic cell death? Immunity,4, 195–201 (1996).
20. Munker, R. et al. Expression of CD95(FAS) by gene transfer does not sensitize K562to Fas-killing. Hematol. Cell Ther. 39, 75–78 (1997).
21. Owen-Schaub L.B. et al. Wild-type human p53 and a temperture-sensitive mutantInduce Fas/APO-1 Expression. Mol. Cell. Biol. 15, 3032–3040 (1995).
22. Whiteside, T.L. & Herberman, R.B. The role of natural killer cells in immune surveil-lance of cancer. Curr. Opin. Immunol. 7, 704–710 (1995).
23. Kaku T. et al. The prognostic significance of tumor-associated antigen 22-1-1 ex-pression in adenocarcinoma of the uterine cervix. Clinical Cancer Research (in thepress).
24. Watanabe T. et al. Isolation of estrogen-responsive genes with a CpG island library.Mol. Cell. Biol. 18, 442–449 (1998).
25. Fukuda, T. et al. Restoration of surface IgM-mediated apoptosis in an anti-IgM-re-sistant variant of WEHI-231 lymphoma cells by HS-1, a protein-tyrosine kinase sub-strate. Proc. Natl. Acad. Sci. USA 92, 7302–7306 (1995).
© 1999 Nature America Inc. • http://medicine.nature.com©
199
9 N
atu
re A
mer
ica
Inc.
• h
ttp
://m
edic
ine.
nat
ure
.co
m