4.3 nucleic acids 4.3a dna structure 4.3b rna...
TRANSCRIPT

Part IV => DNA and RNA
§4.3 Nucleic Acids
§4.3a DNA Structure
§4.3b RNA Structure

I will climb up this
DNA ladder to see
where life takes me!
Section 4.3a:
DNA Structure

Synopsis 4.3a
- DNA is a double-stranded biopolymer comprised of four constituent units or building blocks referred to as “deoxyribonucleotides”—the base components of which in general are adenine (A), cytosine (C), guanine (G), and thymine (T)—wherein A base pairs with T, and G with C
- DNA carries genetic information in the form of its sequence of deoxyribonucleotides—linked together via phosphodiester bonds
- In the so-called DNA double helix, two antiparallel polynucleotide strands wind around each other—they are stabilized by van der Waals forces (hydrogen bonding & stacking interactions), and ionic interactions between opposite strands
- DNA double-helix can adopt three distinct conformations: A-DNA, B-DNA, or Z-DNA
- DNA can be denatured by heating and renatured through annealing
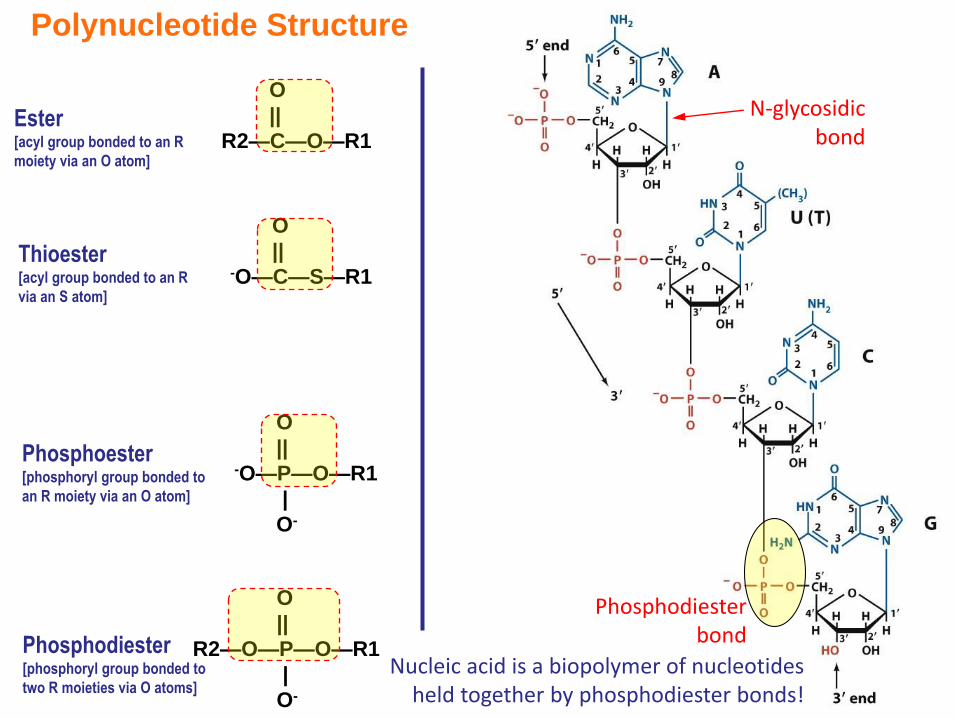
Phosphodiester bond
Nucleic acid is a biopolymer of nucleotides held together by phosphodiester bonds!
N-glycosidicbond
O
||
R2—C—O—R1Ester[acyl group bonded to an R
moiety via an O atom]
O
||-O—P—O—R1
|
O-
Phosphoester[phosphoryl group bonded to
an R moiety via an O atom]
Thioester[acyl group bonded to an R
via an S atom]
O
||-O—C—S—R1
O
||
R2—O—P—O—R1
|
O-
Phosphodiester[phosphoryl group bonded to
two R moieties via O atoms]
Polynucleotide Structure
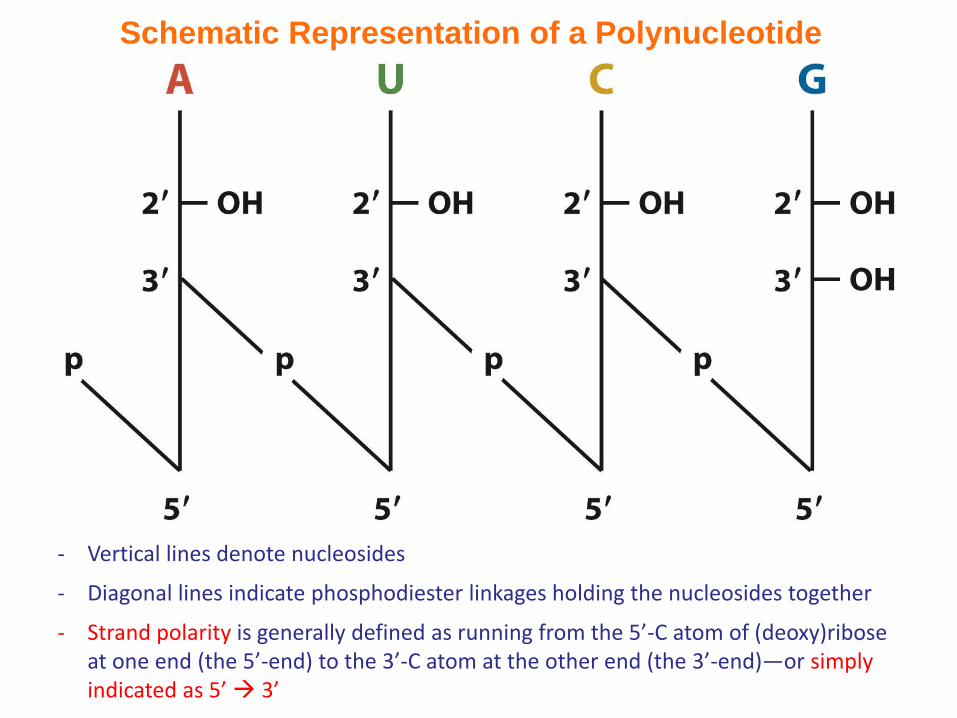
Schematic Representation of a Polynucleotide
- Vertical lines denote nucleosides
- Diagonal lines indicate phosphodiester linkages holding the nucleosides together
- Strand polarity is generally defined as running from the 5’-C atom of (deoxy)ribose at one end (the 5’-end) to the 3’-C atom at the other end (the 3’-end)—or simply indicated as 5’ 3’
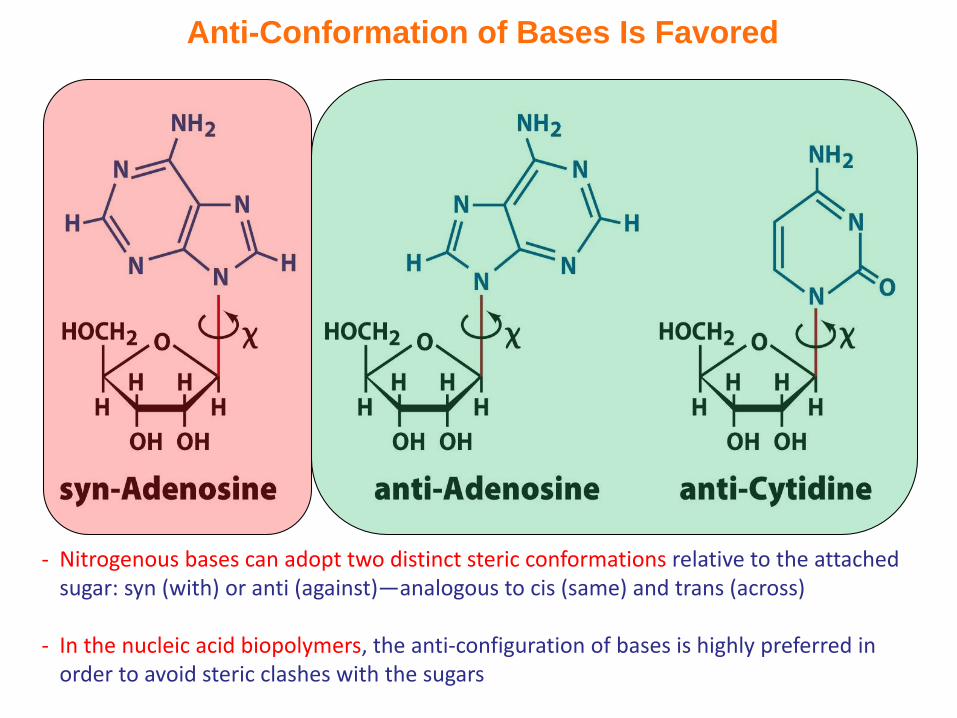
Anti-Conformation of Bases Is Favored
- Nitrogenous bases can adopt two distinct steric conformations relative to the attached sugar: syn (with) or anti (against)—analogous to cis (same) and trans (across)
- In the nucleic acid biopolymers, the anti-configuration of bases is highly preferred in order to avoid steric clashes with the sugars

The Double Helix Discovery (1953)
Rosalind Franklin(1920-1958)
DNA Model (1953)First formulated by Watson and Crick—
but on the basis of x-ray crystallographic data generated by Franklin (in collaboration with Wilkins)
Maurice Wilkins(1916-2004)
James Watson(1928-date)
Francis Crick(1916-2004)
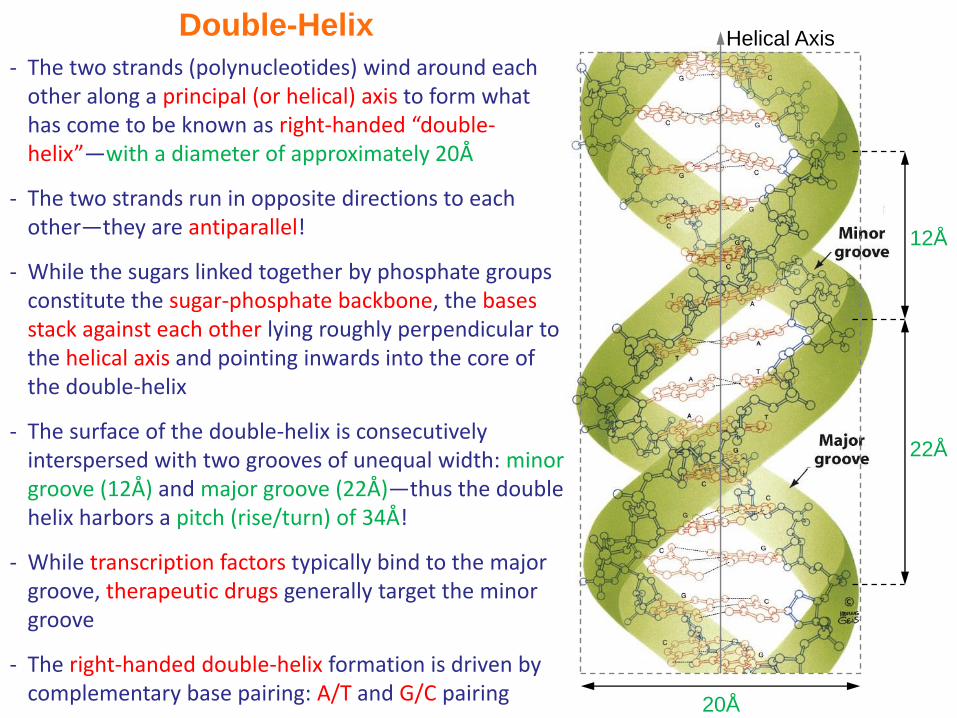
Double-Helix
20Å
Helical Axis
12Å
22Å
- The two strands (polynucleotides) wind around each other along a principal (or helical) axis to form what has come to be known as right-handed “double-helix”—with a diameter of approximately 20Å
- The two strands run in opposite directions to each other—they are antiparallel!
- While the sugars linked together by phosphate groups constitute the sugar-phosphate backbone, the bases stack against each other lying roughly perpendicular to the helical axis and pointing inwards into the core of the double-helix
- The surface of the double-helix is consecutively interspersed with two grooves of unequal width: minor groove (12Å) and major groove (22Å)—thus the double helix harbors a pitch (rise/turn) of 34Å!
- While transcription factors typically bind to the major groove, therapeutic drugs generally target the minor groove
- The right-handed double-helix formation is driven by complementary base pairing: A/T and G/C pairing

Double-Helix: Handedness
- In each case, the fingers curl in the direction the helix turns—provided that the thumb is pointing parallel to the principal axis of the helix!
- The handedness is retained irrespective of the orientation of the helix—turning it upside down has no effect!
Clockwise
Spiral
Counterclockwise
Spiral
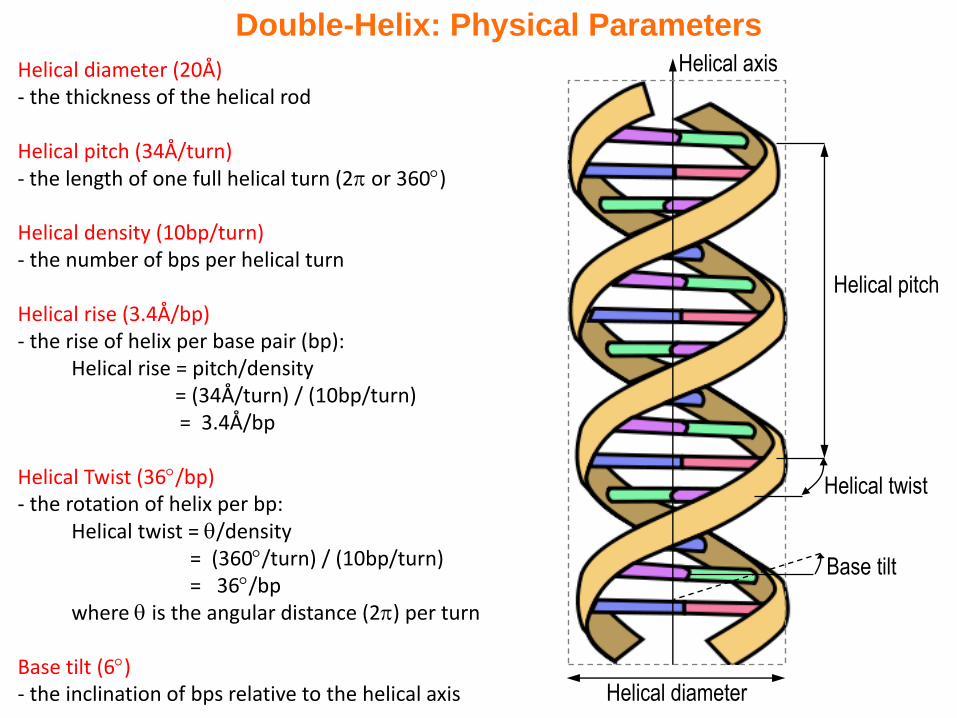
Double-Helix: Physical Parameters
Helical diameter (20Å)- the thickness of the helical rod
Helical pitch (34Å/turn)- the length of one full helical turn (2 or 360)
Helical density (10bp/turn)- the number of bps per helical turn
Helical rise (3.4Å/bp)- the rise of helix per base pair (bp):
Helical rise = pitch/density = (34Å/turn) / (10bp/turn)= 3.4Å/bp
Helical Twist (36/bp)- the rotation of helix per bp:
Helical twist = /density= (360/turn) / (10bp/turn)= 36/bp
where is the angular distance (2) per turn
Base tilt (6)- the inclination of bps relative to the helical axis Helical diameter
Helical pitch
Helical twist
Helical axis
Base tilt
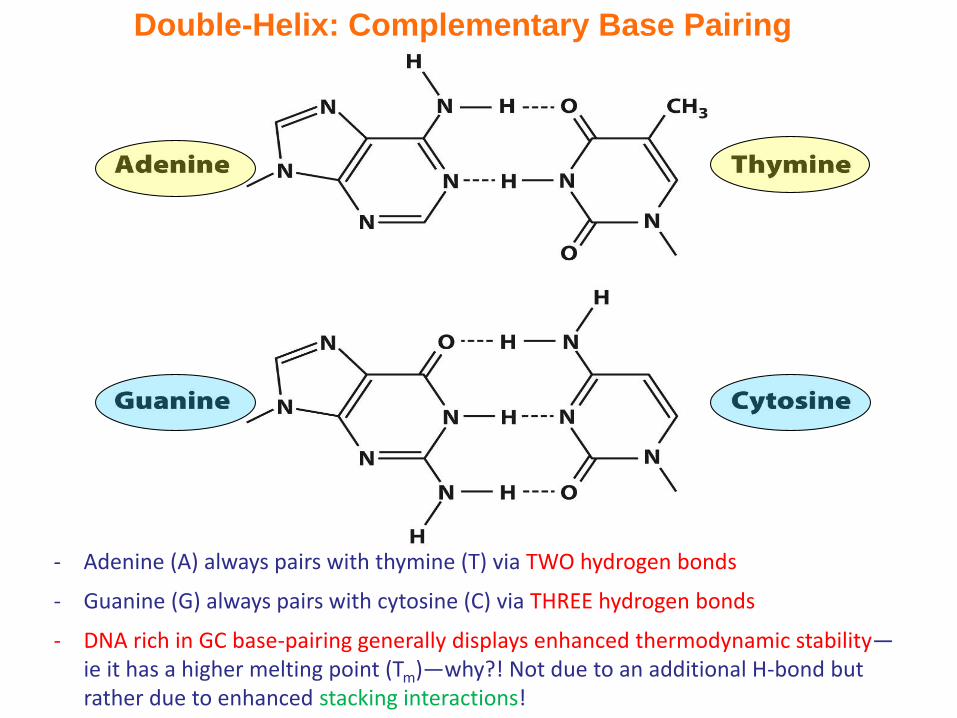
Double-Helix: Complementary Base Pairing
- Adenine (A) always pairs with thymine (T) via TWO hydrogen bonds
- Guanine (G) always pairs with cytosine (C) via THREE hydrogen bonds
- DNA rich in GC base-pairing generally displays enhanced thermodynamic stability—ie it has a higher melting point (Tm)—why?! Not due to an additional H-bond but rather due to enhanced stacking interactions!
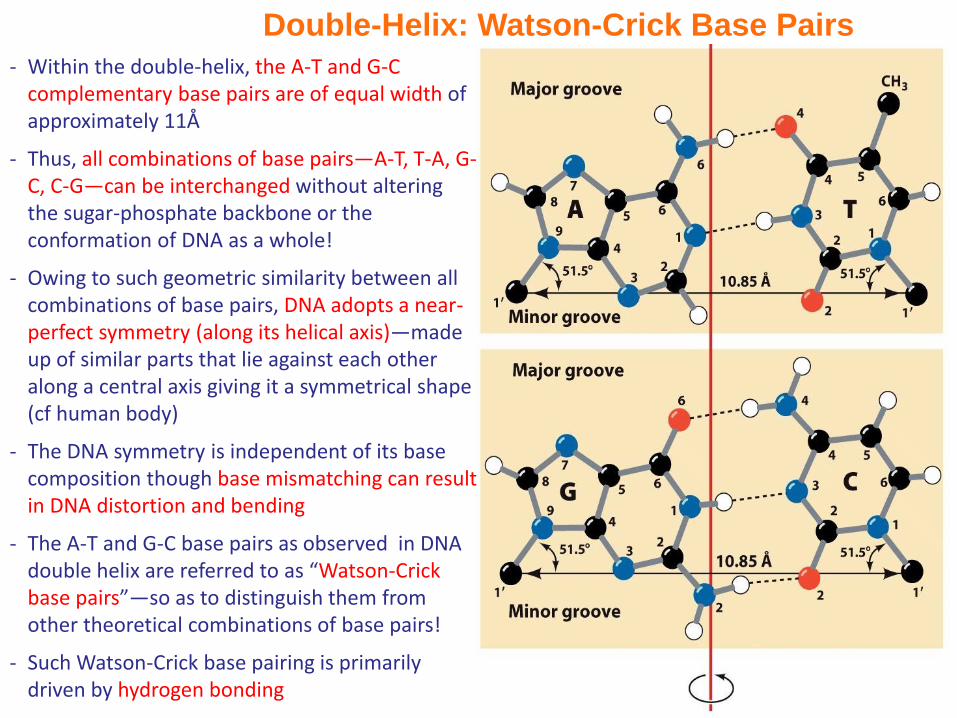
Double-Helix: Watson-Crick Base Pairs
- Within the double-helix, the A-T and G-C complementary base pairs are of equal width of approximately 11Å
- Thus, all combinations of base pairs—A-T, T-A, G-C, C-G—can be interchanged without altering the sugar-phosphate backbone or the conformation of DNA as a whole!
- Owing to such geometric similarity between all combinations of base pairs, DNA adopts a near-perfect symmetry (along its helical axis)—made up of similar parts that lie against each other along a central axis giving it a symmetrical shape (cf human body)
- The DNA symmetry is independent of its base composition though base mismatching can result in DNA distortion and bending
- The A-T and G-C base pairs as observed in DNA double helix are referred to as “Watson-Crick base pairs”—so as to distinguish them from other theoretical combinations of base pairs!
- Such Watson-Crick base pairing is primarily driven by hydrogen bonding
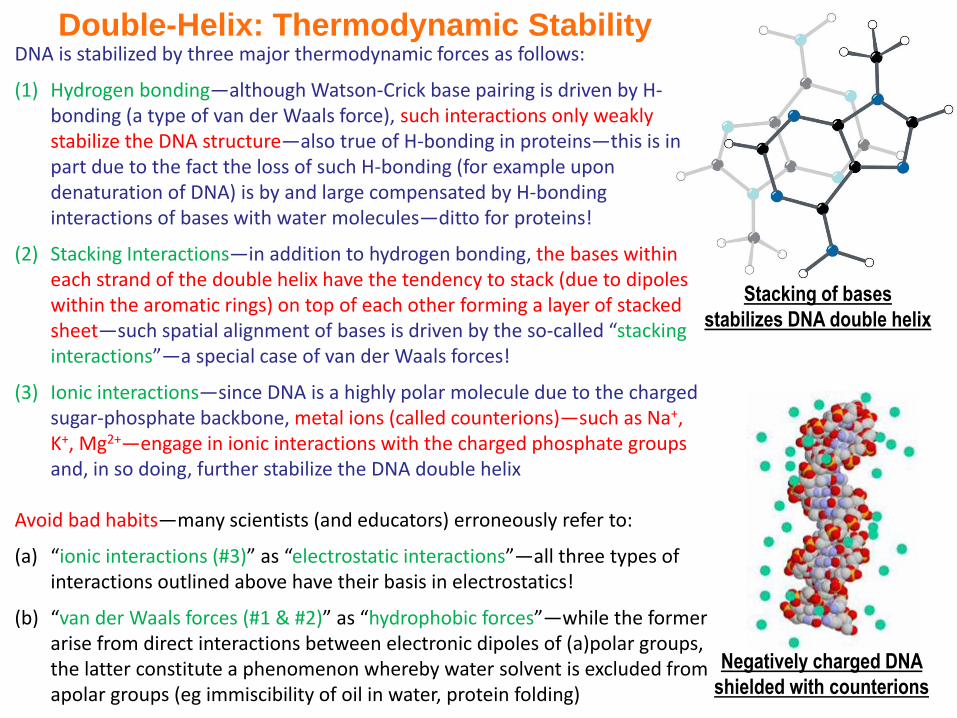
Double-Helix: Thermodynamic Stability
Stacking of bases
stabilizes DNA double helix
Negatively charged DNA
shielded with counterions
DNA is stabilized by three major thermodynamic forces as follows:
(1) Hydrogen bonding—although Watson-Crick base pairing is driven by H-bonding (a type of van der Waals force), such interactions only weakly stabilize the DNA structure—also true of H-bonding in proteins—this is in part due to the fact the loss of such H-bonding (for example upon denaturation of DNA) is by and large compensated by H-bonding interactions of bases with water molecules—ditto for proteins!
(2) Stacking Interactions—in addition to hydrogen bonding, the bases within each strand of the double helix have the tendency to stack (due to dipoles within the aromatic rings) on top of each other forming a layer of stacked sheet—such spatial alignment of bases is driven by the so-called “stacking interactions”—a special case of van der Waals forces!
(3) Ionic interactions—since DNA is a highly polar molecule due to the charged sugar-phosphate backbone, metal ions (called counterions)—such as Na+, K+, Mg2+—engage in ionic interactions with the charged phosphate groups and, in so doing, further stabilize the DNA double helix
Avoid bad habits—many scientists (and educators) erroneously refer to:
(a) “ionic interactions (#3)” as “electrostatic interactions”—all three types of interactions outlined above have their basis in electrostatics!
(b) “van der Waals forces (#1 & #2)” as “hydrophobic forces”—while the former arise from direct interactions between electronic dipoles of (a)polar groups, the latter constitute a phenomenon whereby water solvent is excluded from apolar groups (eg immiscibility of oil in water, protein folding)
max
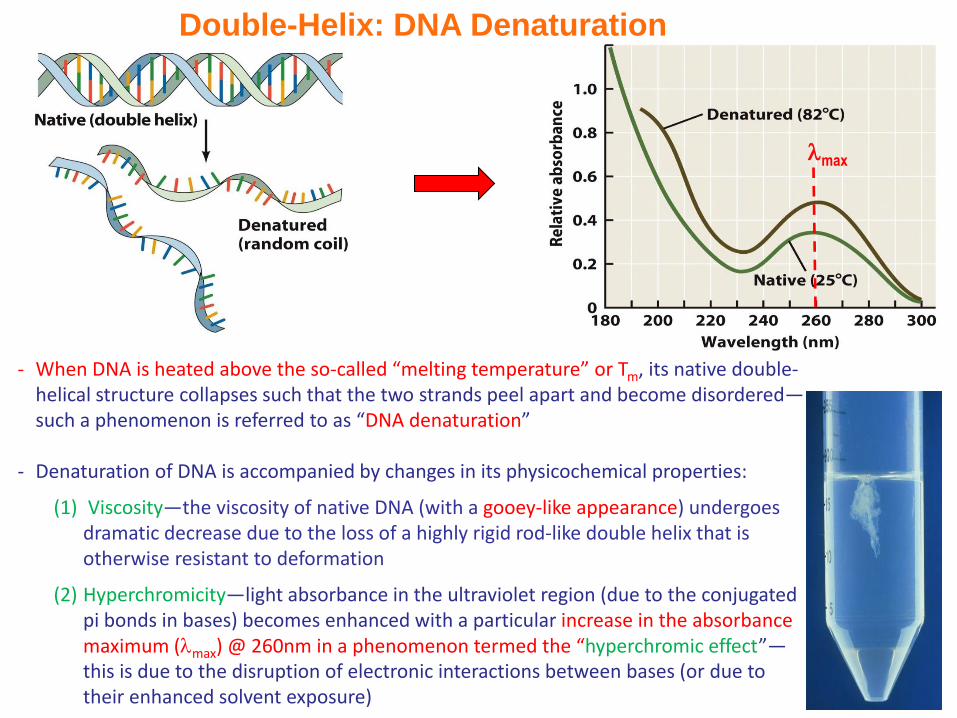
Double-Helix: DNA Denaturation
- When DNA is heated above the so-called “melting temperature” or Tm, its native double-helical structure collapses such that the two strands peel apart and become disordered—such a phenomenon is referred to as “DNA denaturation”
- Denaturation of DNA is accompanied by changes in its physicochemical properties:
(1) Viscosity—the viscosity of native DNA (with a gooey-like appearance) undergoes dramatic decrease due to the loss of a highly rigid rod-like double helix that is otherwise resistant to deformation
(2) Hyperchromicity—light absorbance in the ultraviolet region (due to the conjugated pi bonds in bases) becomes enhanced with a particular increase in the absorbance maximum (max) @ 260nm in a phenomenon termed the “hyperchromic effect”—this is due to the disruption of electronic interactions between bases (or due to their enhanced solvent exposure)
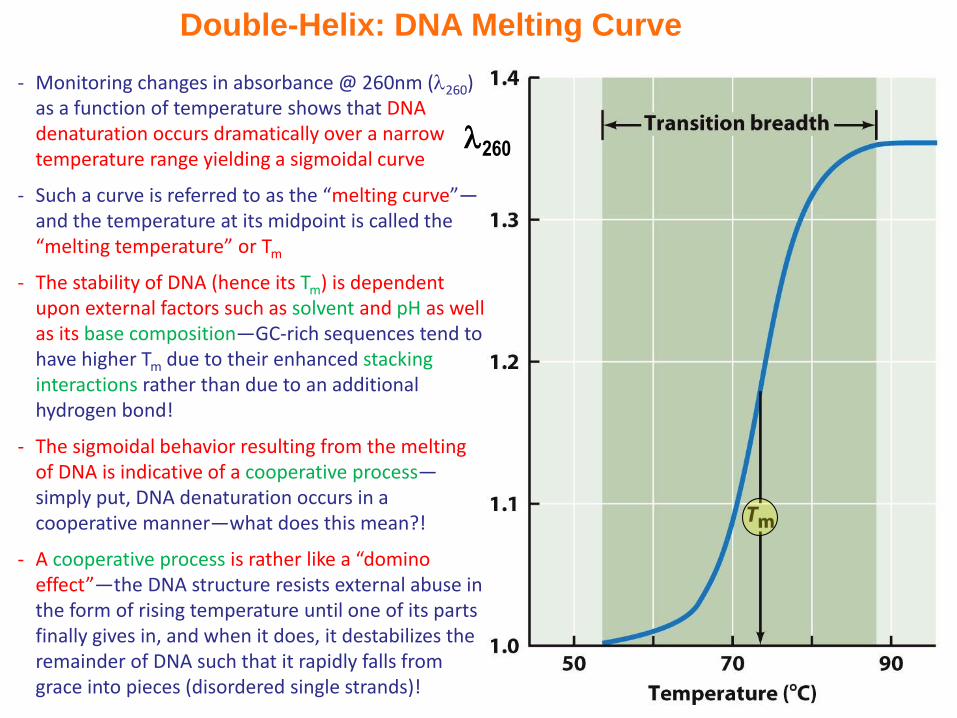
Double-Helix: DNA Melting Curve
- Monitoring changes in absorbance @ 260nm (260) as a function of temperature shows that DNA denaturation occurs dramatically over a narrow temperature range yielding a sigmoidal curve
- Such a curve is referred to as the “melting curve”—and the temperature at its midpoint is called the “melting temperature” or Tm
- The stability of DNA (hence its Tm) is dependent upon external factors such as solvent and pH as well as its base composition—GC-rich sequences tend to have higher Tm due to their enhanced stacking interactions rather than due to an additional hydrogen bond!
- The sigmoidal behavior resulting from the melting of DNA is indicative of a cooperative process—simply put, DNA denaturation occurs in a cooperative manner—what does this mean?!
- A cooperative process is rather like a “domino effect”—the DNA structure resists external abuse in the form of rising temperature until one of its parts finally gives in, and when it does, it destabilizes the remainder of DNA such that it rapidly falls from grace into pieces (disordered single strands)!
260
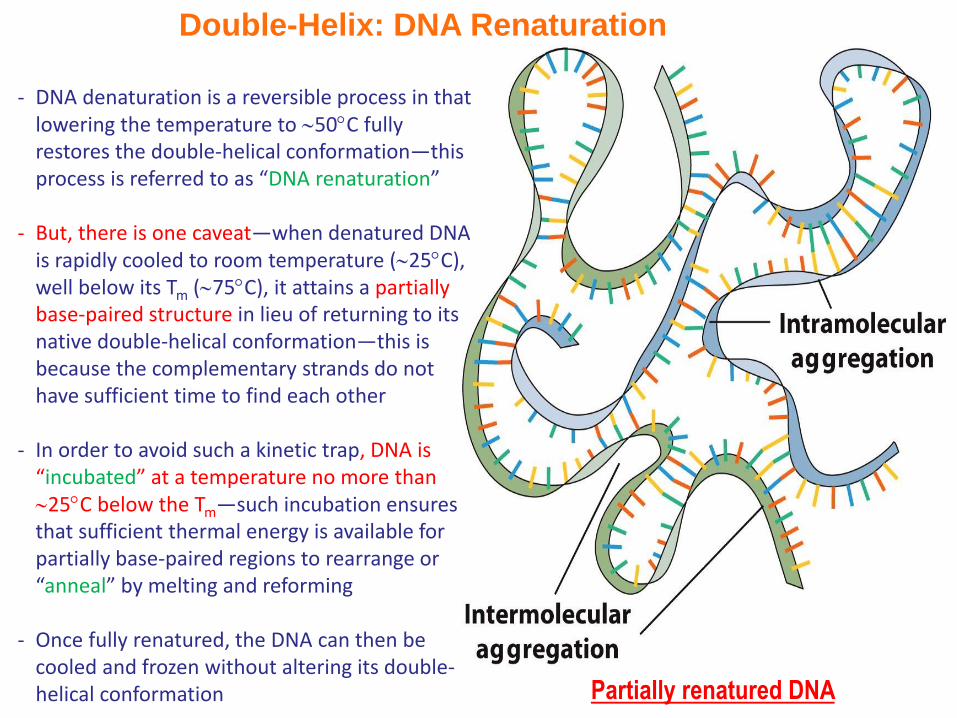
Double-Helix: DNA Renaturation
- DNA denaturation is a reversible process in that lowering the temperature to 50C fully restores the double-helical conformation—this process is referred to as “DNA renaturation”
- But, there is one caveat—when denatured DNA is rapidly cooled to room temperature (25C), well below its Tm (75C), it attains a partially base-paired structure in lieu of returning to its native double-helical conformation—this is because the complementary strands do not have sufficient time to find each other
- In order to avoid such a kinetic trap, DNA is “incubated” at a temperature no more than 25C below the Tm—such incubation ensures that sufficient thermal energy is available for partially base-paired regions to rearrange or “anneal” by melting and reforming
- Once fully renatured, the DNA can then be cooled and frozen without altering its double-helical conformation Partially renatured DNA

DNA Conformations
- Owing to its conformational flexibility (albeit somewhat restricted relative to proteins), DNA adopts distinct structural conformations in solution depending on the solvent and base composition
- Three major conformations of DNA are referred to as the A-DNA, B-DNA, and Z-DNA—of these three conformations, B-DNA is thermodynamically most stable and hence the biologically most common form
- The discussion on double helix has hitherto been solely focused on B-DNA—how does such biologically most prevalent form of DNA differ from others?
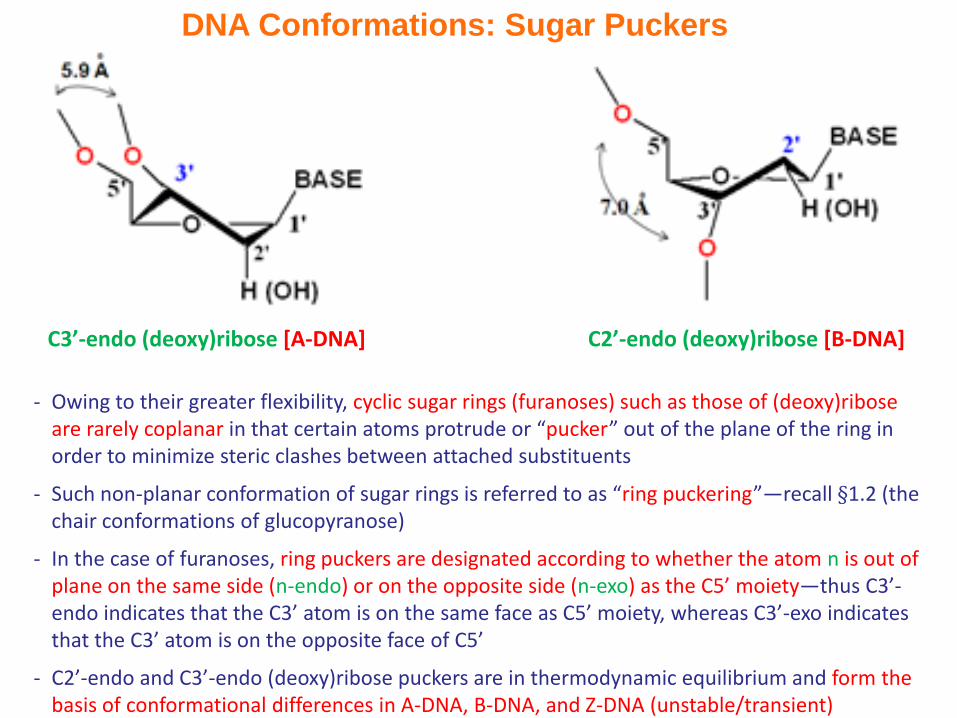
DNA Conformations: Sugar Puckers
- Owing to their greater flexibility, cyclic sugar rings (furanoses) such as those of (deoxy)ribose are rarely coplanar in that certain atoms protrude or “pucker” out of the plane of the ring in order to minimize steric clashes between attached substituents
- Such non-planar conformation of sugar rings is referred to as “ring puckering”—recall §1.2 (the chair conformations of glucopyranose)
- In the case of furanoses, ring puckers are designated according to whether the atom n is out of plane on the same side (n-endo) or on the opposite side (n-exo) as the C5’ moiety—thus C3’-endo indicates that the C3’ atom is on the same face as C5’ moiety, whereas C3’-exo indicates that the C3’ atom is on the opposite face of C5’
- C2’-endo and C3’-endo (deoxy)ribose puckers are in thermodynamic equilibrium and form the basis of conformational differences in A-DNA, B-DNA, and Z-DNA (unstable/transient)
C2’-endo (deoxy)ribose [B-DNA]C3’-endo (deoxy)ribose [A-DNA]

DNA Conformations: Structural Comparison
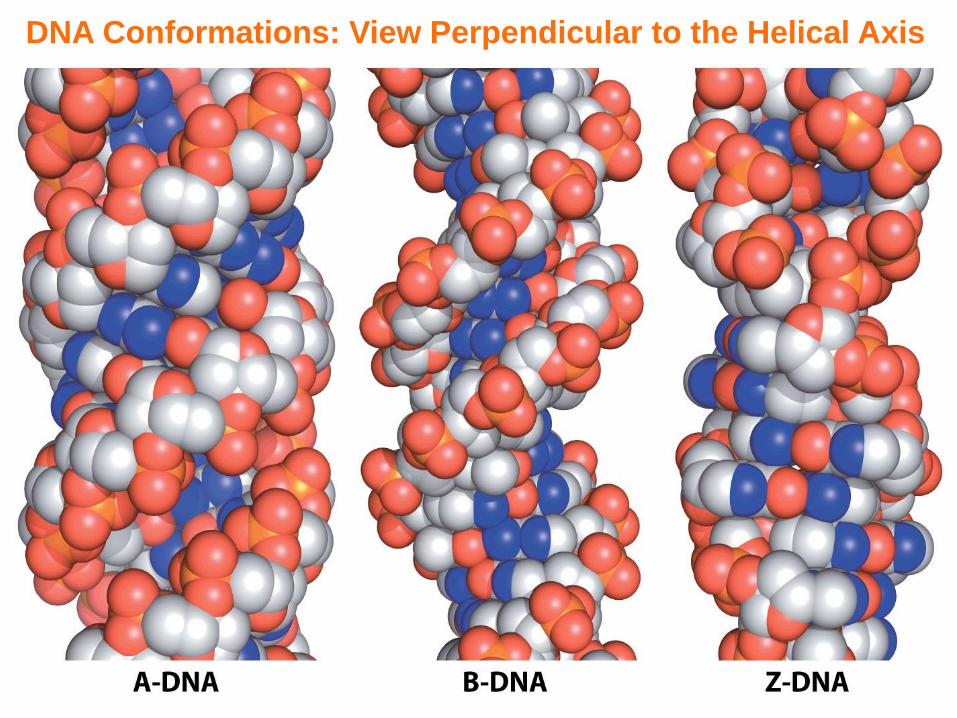
DNA Conformations: View Perpendicular to the Helical Axis
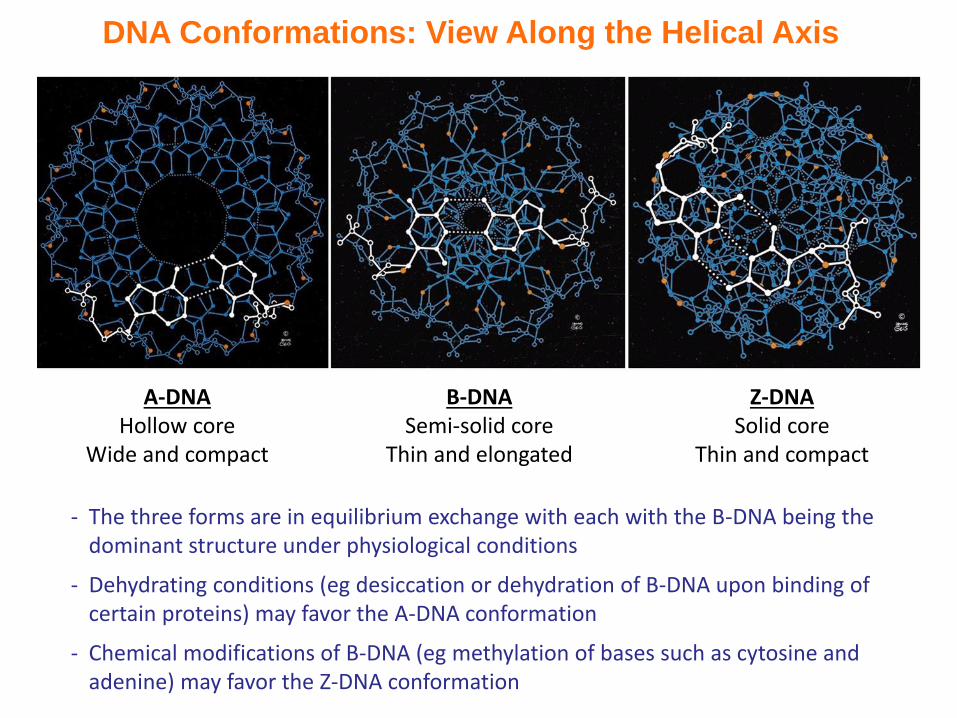
DNA Conformations: View Along the Helical Axis
A-DNAHollow core
Wide and compact
B-DNASemi-solid core
Thin and elongated
Z-DNASolid core
Thin and compact
- The three forms are in equilibrium exchange with each with the B-DNA being the dominant structure under physiological conditions
- Dehydrating conditions (eg desiccation or dehydration of B-DNA upon binding of certain proteins) may favor the A-DNA conformation
- Chemical modifications of B-DNA (eg methylation of bases such as cytosine and adenine) may favor the Z-DNA conformation

Exercise 4.3a
- Draw the complete structure of a nucleoside triphosphate before and after
it becomes incorporated into a polynucleotide chain
- Describe the structural differences between A-, B-, and Z-DNA, including
handedness, diameter, and presence of grooves
- Explain why most nucleotides adopt the anti-conformation
- In what nucleic acid conformation(s) does the ribose pucker with C2’ endo?
C3’ endo?
- Describe the forces that stabilize nucleic acid structure. Which is most
important?
- Explain the molecular events of nucleic acid denaturation and renaturation
Section 4.3b:
RNA Structure
Synopsis 4.3b
- RNA (ribonucleic acid) is a single-stranded biopolymer comprised of four constituent units or building blocks referred to as “ribonucleotides”—the base components of which in general are adenine (A), cytosine (C), guanine (G), and uracil (U)—tRNA also contains hypoxanthine (I)
- Like DNA, phosphodiester bonds link ribonucleotide residues in RNA
- Unlike DNA, RNA molecules are usually single-stranded but may adopt secondary structure by virtue of their ability to engage in intra-strandbase pairs—wherein A base pairs with U, and C with G/I
- Of the many forms that it adopts, messenger RNA (mRNA) is the only coding RNA—ie it is translated into proteins!
- In addition to mRNA, RNA also exists in the form of so-called “noncoding RNA” or ncRNA species involved in orchestrating numerous cellular functions such as RNA translation, RNA splicing, and gene regulation
- Two most abundant forms of ncRNA in cells include:(1) Ribosomal RNA (rRNA)—a constituent of ribosomes(2) Transfer RNA (tRNA)—a component of translational machinery
Single-Stranded RNA (ssRNA)
- RNA differs from DNA in two major respects:(1) Deoxyribose (2’-H) is replaced with ribose (2’-OH)(2) Thymine (5-methyluracil) is replaced with uracil (5-H)
- The fact that the RNA is predominantly a single-stranded (ssRNA) polymer is largely due to the ribose sugar—the steric clashes between 2’-OH moieties on the ribose sugar of ribonucleotides prevent the formation of a double helix!

stem
loop
- RNA can also adopt secondary structure via intramolecular base-pairing of long stretches of complimentary ribonucleotides within the same strand to form double-stranded (dsRNA) structures
- Such intra-strand base-pairing (due to A-U and G-C base pairs)—as opposed to inter-strand base-pairing in DNA—results in the formation of RNA structures referred to as “stem-loop” or “hairpin loop” fold
Double-Stranded RNA (dsRNA)

RNA-DNA Double Helix
RNA strand
DNA strand
In the presence of complementary DNA strands, RNA can also assume double helix conformation that usually resembles that of A-DNA or a mixture of A-DNA/B-DNA
Short stretches of such hybrid RNA-DNA double helices are required during the:
(1) Initiation of DNA replication, where they serve as RNA primers (see §4.4)
(2) Transcription of RNA on DNA templates (see §4.5)

Exercise 4.3b
- List the structural differences between DNA and RNA
- What structures can RNA adopt alone and in complex with
complementary strands of DNA?