3.6 a primer in morphogenesis and developmental biology
TRANSCRIPT

3.6 A primer in morphogenesis and developmental biology

What are the big questions in developmental biology?

Phylotaxis – leafs on plants are usually arranged in specific geometries (according to the golden mean).

Limb development – what determines when and how limbs are formed?

Scaling – How come that animals always have the same proportions no matter their size?

Growth – How does an organism know when to stop growing (by the way note the scaling in the picture below even though it doesn‘t work physically)?

Morphogenesis - How do you get from a spherical egg to say a frog?

"It is not birth, marriage or death, but gastrulation, which is truly the most important time in your life."
Lewis Wolpert

First developmental experiments: Willhelm Roux on sea urchins
3.6.1 Morphogen gradients

Driesch repeats the experiments and gets very different results

Spemann Mangold experiment – bringing both sides back together

And now for some physics: Enter Alan Turing
Turing, Phil. Trans. Roy. Soc. B 237, 37 (1952)

The activator-inhibitor system shows an instability to fluctuations.

An application to this may be in Phylotaxis or why do plants know the Fibonacci series.

In 1969 the world changed...

Lewis Wolpert takes up Turing‘s ideas experimentally and produces his own mathematical treatment.

Morphogen diffusion with breakdown
stationary state
with the solution
Wolpert, Journal of theoretical biology 25, 1 (1969)
Take a source at one end of the embryo and let the morphogen diffuse through it.
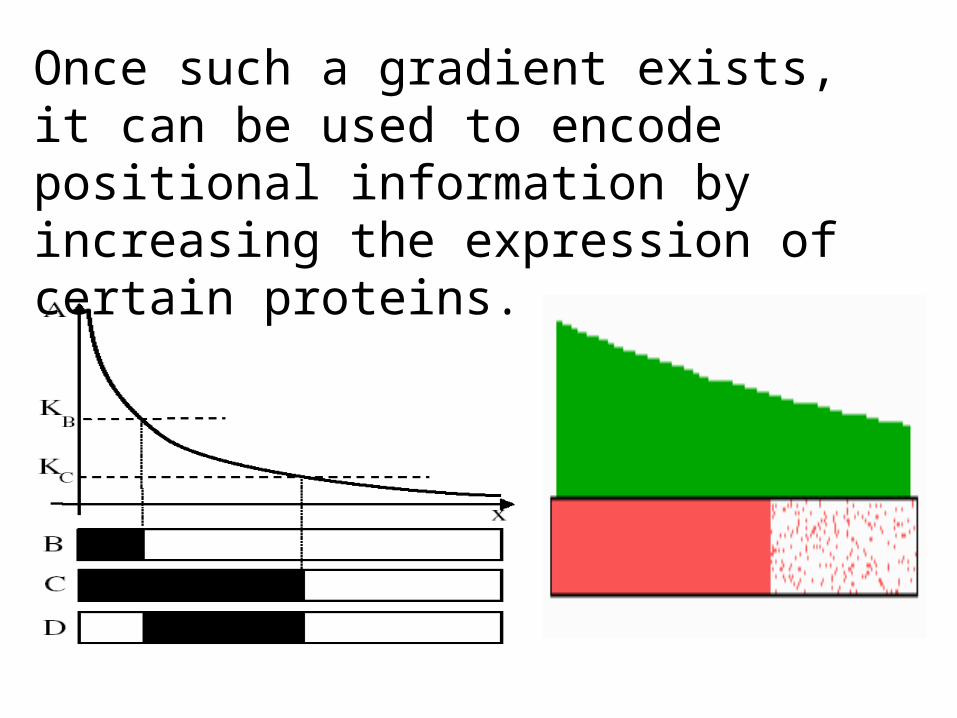
Once such a gradient exists, it can be used to encode positional information by increasing the expression of certain proteins.

How morphogens actually work we‘ll see in example 2...
But there‘s more: positional information is kept when different genes are expressed – and development is robust (sea urchins always look the same no matter what you take away from them... So there‘s scaling.

Chick limb development: the morphogen sonic hedgehog in the early limb determines the later fate.
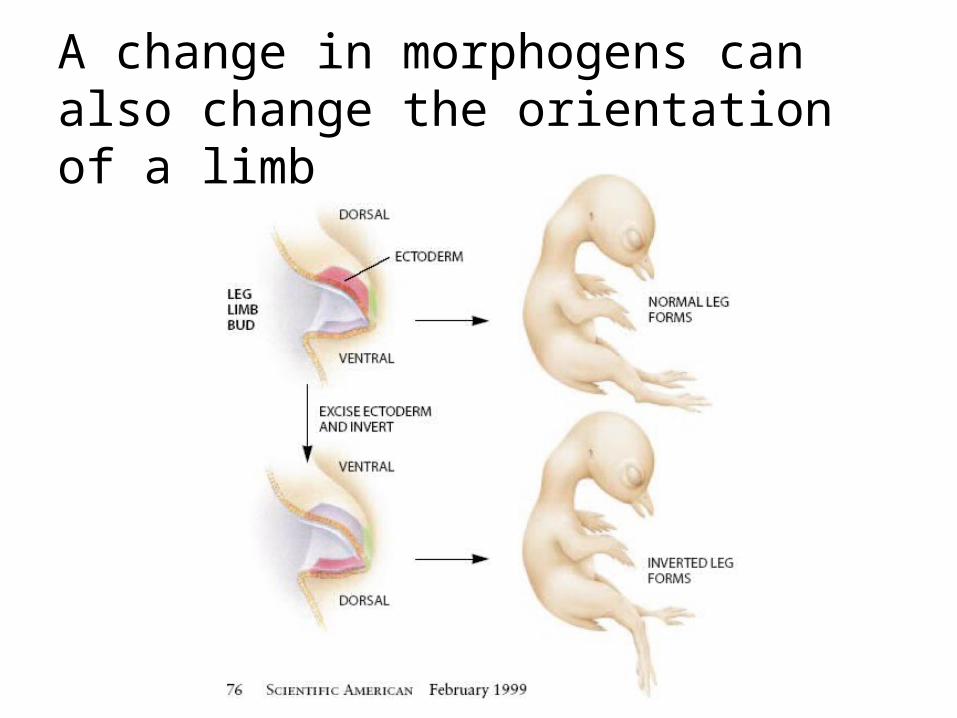
A change in morphogens can also change the orientation of a limb

Extremity development is crucially dependent on the right positional information at a very early stage.

More reaction-diffusion systems and more physics: Hans Meinhardt
Gierer & Meinhardt, Kybernetik 12, 30 (1972).
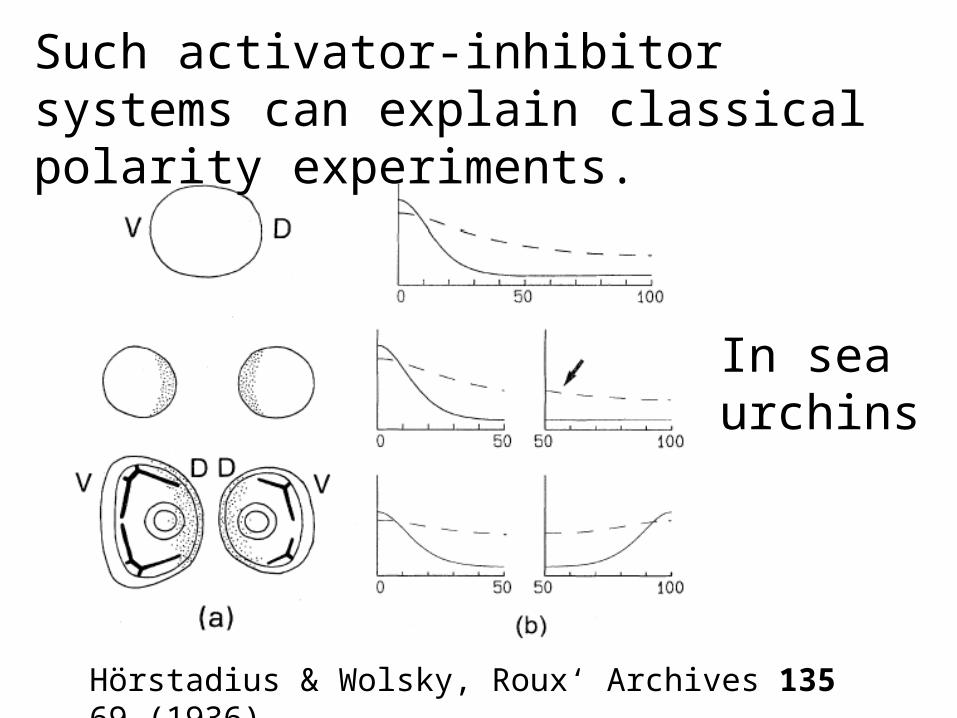
Hörstadius & Wolsky, Roux‘ Archives 135 69 (1936).
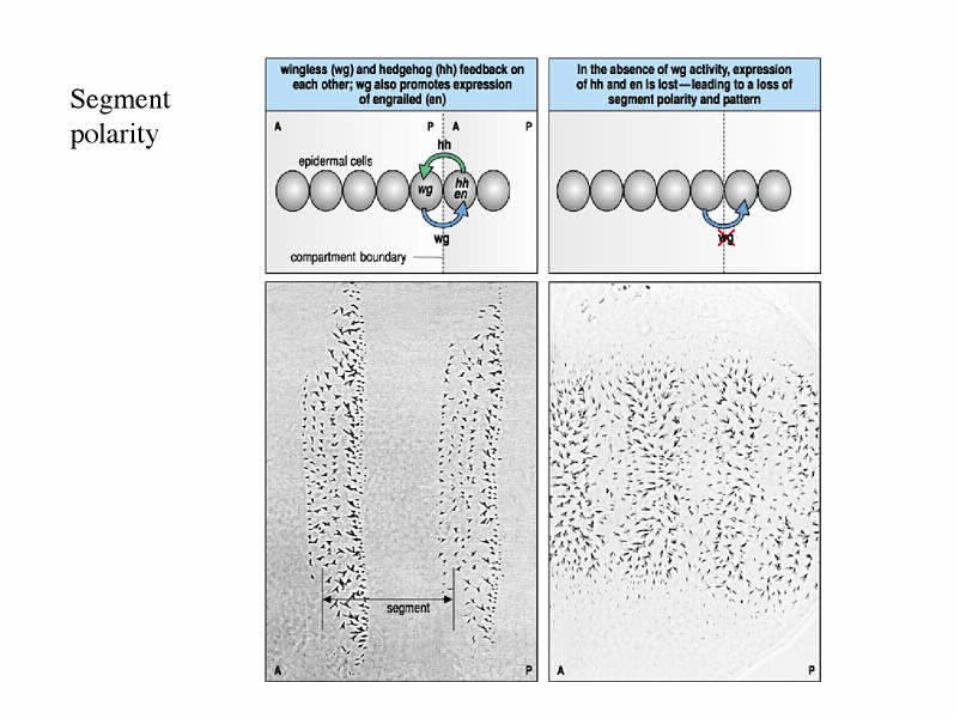
Such activator-inhibitor systems can explain classical polarity experiments.
In sea urchins
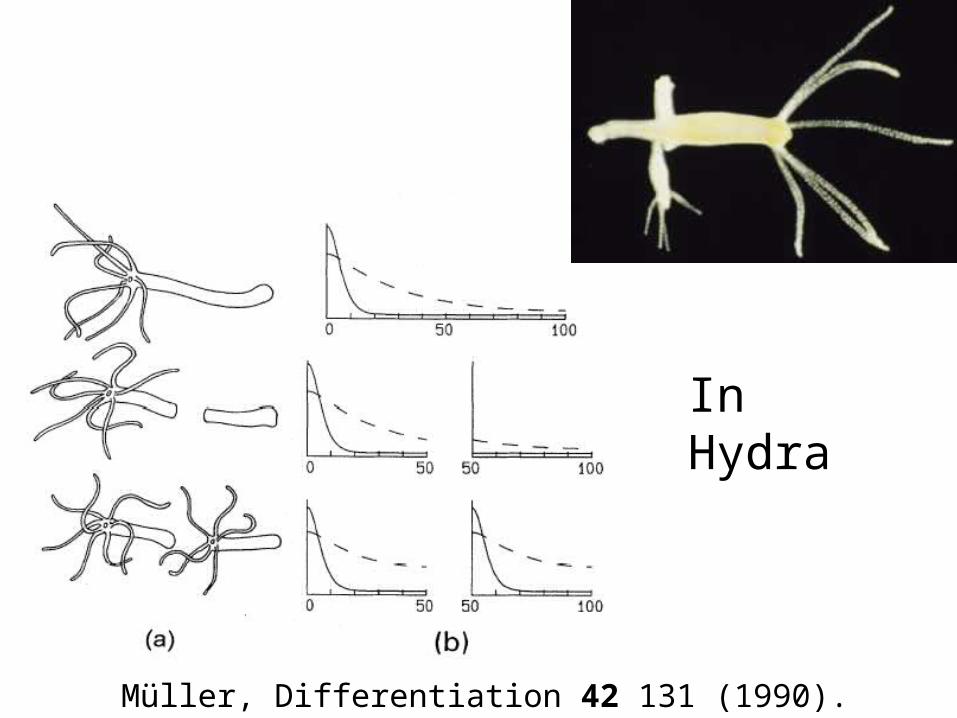
Müller, Differentiation 42 131 (1990).
In Hydra

Such reaction diffusion systems of three different morphogens can also lead to spatial stabilization.

This isn‘t just an academic plaything – the proteins MinC, MinD and MinE, which are important in the division of E. coli show exactly these oscillations. Thus leading to an accurate splitting.
Raskin & de Boer, PNAS 96 4971 (1999).

3.6.2 A primer in pattern formation
Start with the Gierer-Meinhardt equations as an example:
For simplicity, we set a = h = 0

dimensionless variables:
gives simpler equations:

Solve them for the homogeneous steady state (i.e. = 0 and t = 0):
Then perturb this state with a harmonic function and only keep terms linear in a0 and h0:

This gives the linear system of equations:
with

There is only a solution with non-zero a and h if the discriminant of the Matrix is zero:
with

The fluctuations only grow if the real part of > 0. The critical value is thus given by Re() = 0. If has complex values (i.e. > (/2)2), the real part is given by /2 and hence the condition is = 0. Thus

On the other hand, if is real valued, then it is only zero if = 0. This yields:
A spatial pattern can therefore only develop in an embryo, if ist size exceeds Lc. As long as the length is close to Lc, this also implies a polarity, since the cosine does not recover on this length scale.

We can do this more generally by assuming that k is continuous. Then we look at which wave number disturbance grows fastest:
while Re() > 0

Again we start with the case that w is complex: then Re() = -/2 and the fastest growing wavenumber is k = 0. The fact that is complex and that Re() > 0 lead to conditions for m where we are in this case of a growing homogeneous state that oscillates.

If is real, we obtain:
and is is positive if:

All of this is summarized in the Stability diagram:
Homogeneous, static pattern
growing, inhomogeneouspattern
Oscillating, homogeneous pattern

Another set of differential equations describes a threshold switch

Simulation of animal coatings using reaction diffusion and a switch

lepard cheetagiraffe
Simulation results for pigmentation

3.6.3 An example: The anterio-posterior axis in Drosophila.
Nüsslein-Vollhard & Wieschaus, Nature 287 795 (1980).

Three different sets of genes
Nüsslein-Vollhard & Wieschaus, Nature 287 795 (1980).

So there‘s a hierarchy of genes and proteins in the early development



Reminder – where are we in the developmental stages...

Lets have a closer look at the gap-genes – their positions determine the stripes of the pair-rule genes

Interactions (as transcription factors) of the different gap genes

This can be visualised using fluorescence probes in vivo....

For instance stripe 2 is given by the competition of Hunchback, Giant and Krüppel

All in all there will be seven stripes of eve expression controlled by different combinations of the gap-genes

Meinhardt has modelled this much more elegantly than nature with less complicated feedback cycles – but nature sometimes isn‘t elegant...
Meinhardt, J. Cell Sci. Suppl. 4 357 (1986).

But how do proteins act as transcription factors on a molecular level such that they can be viewed as morphogens?
See section 3.5 on transcription

Some enhancer sequences for transcription

Krüppel for instance uses such a transcription factor.
Others form fingers which stick out specific DNA binding sites using Zinc groups.

How does one know all this?
Electrophoresis of digested RNA
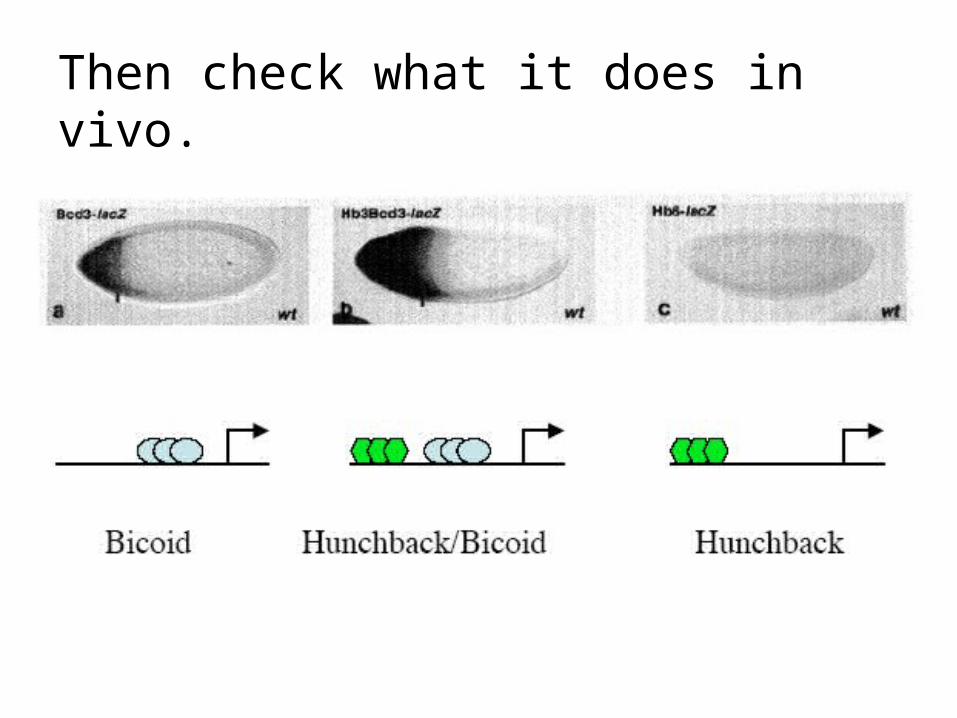
Then check what it does in vivo.

Or specifically Bicoid binding sites
Map of the hb gene indicating the locations of bcd-binding sites. The 2.9 kb hb transcript is expressed in an anterior domain which extends from 55-100% egg length, whereas the 3.2 kb transcript (which is expressed maternally and zygotically) is localized to 0-25% egg length. A, B and C are the fragments identified in the experiment shown in Fig. 2. b, Nucleotide sequences of the regions where bcd protein binds to hb regulatory regions. Base pairs protected against DNaseI digestion as referred from Fig. 4 are indicated by a bar below the sequence.
Driever & Nüsslein-Vollhard, Nature 337 138 (1989).

Lets get back to the development of the anerioposterior axis

The first step is the most important – get a gradient going à la Wolpert!
Driever & Nüsslein-Vollhard, Cell 54 95 (1988).

Bicoid is known to act as an activator for hunchback expression.
Driever & Nüsslein-Vollhard, Nature 337 138 (1989).

Bicoid protein also has the exponential gradient one expects from a maternally deposited morphogen!

However things are a little more complicated than that – Hunckback is expressed too precise in order to just be determined by a Wolperian Bicoid...
Houchmandzadeh, Wieschaus and Leibler, Nature 415, 798 (2002)

Furthermore, the boundary is always in the middle of the embryo irrespective of ist size – this would not be expected from an exponential gradient, which sets a length scale.


On with the development of the fly…

During metamorphosis the imaginal discs turn into the proper organs for example the wing is folded out from the wing disc

How does this influence the setting of scales and coordinates in the wing disc, i.e. How is the later shape of the wing encoded in this?

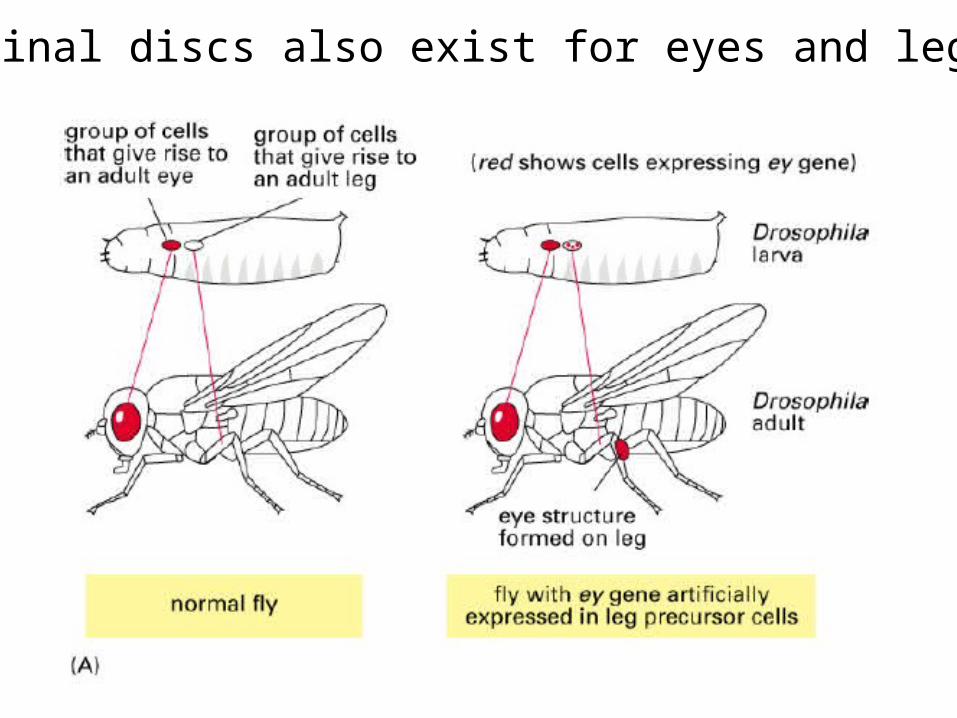
Imaginal discs also exist for eyes and legs

Expressing the 'wrong' genes in the leg disc leads to legs with eyes