2,4-dichlorophenoxyacetic acid 1,1 (monuron) roots1 · absorption of2,4-dichlorophenoxyacetic acid...
TRANSCRIPT

Plant Physiol. (1973) 52, 638-645
Absorption of 2,4-Dichlorophenoxyacetic Acid and3-(p-Chlorophenyl)-1,1-dimethylurea (Monuron)
by Barley Roots1Received for publication July 25, 1972
T. W. DONALDSON,2 D. E. BAYER, AND 0. A. LEONARDDepartment of Botany, University of California, Davis, California 95616
ABSTRACT
Absorption from culture solution of the herbicides 2,4-dichlorophenoxyacetic acid (2,4-D) and 3- (p-chlorophenyl)-1, 1-dimethylurea (Monuron) by excised barley (Hordeumvulgare L.) roots was studied to determine whether absorptionwas due to an active or a passive mechanism. Herbicide absorp-tion was followed at low temperature, under anaerobic condi-tions, and in the presence of metabolic inhibitors and com-pounds of structure similar to that of the herbicide. Totalabsorption was divided into two phases, exchangeable andnonexchangeable herbicide, by washing the roots for 1 hourfollowing absorption. Absorption of both exchangeable and non-exchangeable 2,4-D appeared to depend on a supply of meta-bolic energy which suggests that an active mechanism may beinvolved. A possible conclusion is that 2,4-D is absorbed byroots by an adsorption mechanism and that energy is requiredto maintain the integrity of the absorbing surfaces of the cell.In contrast, absorption of Monuron was independent of anenergy supply. It is concluded that the bulk of the Monuronabsorbed was taken up passively by diffusion.
Absorption of mineral salts by plants has been extensivelystudied and much information on this subject now exists. Al-though many organic compounds, such as insecticides, fungi-cides, and herbicides, are applied either directly to plants, orto the soil from where they are taken up by the roots, less in-formation is available on the absorption of organic substances.
Both physical and metabolically controlled processes havebeen shown to control the absorption of indoleacetic acidby pea epicotyl segments and carrot disks (20), and Avenacoleoptile sections (17). With Lemna minor the initial uptakeof 2,2-dichloropropionic acid (Dalapon) was concluded to belargely under the control of physical factors, whereas the sub-sequent accumulation was metabolically controlled (18). Ab-sorption of mercuric acetate and phenylmercuric acetate byexcised and intact pea roots shows both a passive and an activephase of uptake (19). Hancock (10), using diseased and healthysquash hypocotyls, found that 3-o-methylglucose is taken up in
1This work was supported by National Science Foundation GrantGB 2309. This work is from a thesis submitted by T. W. D. in par-tial fulfillment of the requirements for the Ph.D. degree at the Uni-versity of California, Davis.
2Present address: Keith Turnbull Research Station, Frankston,Victoria, Australia, 3199.
the diseased tissue by an active transport system. Two mecha-nisms of absorption of choline sulfate by excised barley rootshave been demonstrated (15), one being the result of activetransport, while the nature of the other could not be deter-mined.Foy and Yamaguchi (9) found a rapid uptake of 2,4-D by
excised barley roots for 30 min, followed by a loss back to thesolution. It was suggested that accumulation during the 30-minperiod was mainly an adsorption process and, since the 2,4-Dcould be readily leached out, it was apparently held by ratherweak bonding forces. Shortly thereafter, Yamaguchi (22) con-cluded that 2,4-D absorption by roots consists initially of aphysical adsorption and that this adsorbed 2,4-D than actsas a source for metabolic absorption. Crafts and Yamaguchi(5) compared the absorption and translocation of a series ofnine compounds applied to the roots of barley. Of these, 3-(p-chlorophenyl)-l, 1-dimethylurea (Monuron) and 2-chloro-4,6-bis(ethylamino)-s-triazine (Simazine) started moving upwardwithin 2 hr, whereas even after 4 days very little 2,4-D orIAA had entered the tops, although they were concentrated inthe roots. Isensee et al. (13) concluded that the uptake andtranslocation of 4-amino-3, 5, 6-trichloropicolinic acid (Pic-loram) by oats and soybeans is a complex phenomenon gov-erned in part by passive and active processes.
As differences in absorption may account for differences ineffectiveness between herbicides, an understanding of theuptake process is desirable. The studies reported herein wereconducted to determine if 2, 4-D and Monuron are absorbed bybarley roots by a passive or an active mechanism, using thesame criteria and methods as have been used in studies on up-take of inorganic ions.
MATERIALS AND METHODS
For all studies, "4C-labeled herbicides were used. The specificradioactivity of the 2,4-D was 1.24 mc/mmole, while that ofthe Monuron was 0.386 mc/mmole. For most trials, a con-centration of 1 ,uM 2,4-D-14C, or 1 uM Monuron-14C, was used.Solutions of these concentrations were made up by adding 100,ul of a 10 mm stock solution of radioactive herbicide to 1 literof one-quarter strength Hoagland's nutrient solution (11). Forsolutions of higher concentration, additional unlabeled herbi-cide was added.
Culture and Treatment of Roots. Barley (Hordeum vulgareL. var. White California Mariout) roots were grown essentiallyaccording to the method described by Epstein (7). Seeds weregerminated for 24 hr in aerated distilled water at room tem-perature (22-23 C) and then spread on a layer of washedcheesecloth supported by a wire frame over a plastic pancontaining one-quarter strength Hoagland's nutrient solution.
638
www.plantphysiol.orgon July 6, 2018 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.
ABSORPTION OF 2,4-D AND MONURON BY ROOTS
The culture solution was continuously aerated and growth tookplace in the dark at room temperature. By the 5th day from theinitial soaking of the seed, the roots were 7 to 10 cm long andready for use.
Immediately prior to the commencement of an absorptionexperiment, the roots were excised just below the wire screenand placed in 1 liter of aerated one-quarter Hoagland's solu-tion. As required, roots were gently blotted dry, and 0.5-g sam-ples were weighed and placed in 50 ml of one-quarter strengthHoagland's solution in a 125-ml Erlenmeyer flask. At the startof the absorption period, the nutrient solution was replaced by50 ml of the treatment solution. All solutions were continuouslyaerated throughout the absorption period unless stated other-wise.The absorption period was terminated by decanting the
solution and washing the roots by one of the following methods:(a) roots were washed for approximately 10 sec in 25 to 30 mlof water, or (b) roots were washed as above, then tied in asmall square of cheesecloth and suspended for 1 hr in a largevolume of nonradioactive herbicide solution, at approximatelythe same strength as used during the absorption period, madeup in one-quarter strength Hoagland's solution. At the end ofthe washing period, the roots were rinsed for 1 min in tapwater. Following washing, the roots were blotted dry, weighed,and dried at 60 C.
Radioactive Assay. The amount of herbicide absorbed bythe roots was measured by a count of the radioactivity present.The Schoniger oxygen combustion technique, as described byWang and Willis (21), was employed for sample preparation.Combustion of the root samples, weighed and wrapped inblack, ashless filter paper wrappers in a flask of oxygen wasaccomplished by momentarily focusing a beam of infrared lightfrom a Thomas-Ogg oxygen flask igniter, Model 11, onto theblack wrapper. Following combustion, 20 ml of an ethanol-amine-absolute ethanol mixture (1:2 v/v) was injected intothe flask. After about 1 hr, during which time the CO2 pro-duced by combustion was absorbed by the ethanolamine, a 5-mlsample was taken and added to 10 ml of scintillation fluid,which consisted of 5 g of PPO and 0.3 g of dimethyl POPOPdissolved in 1 liter of toluene. The samples were counted in aPackard Tri-Carb liquid scintillation spectrometer.
250-
o 200-
or.Yc 150E-.. .
E.cq ~~T-7
Extraction of Root Material. To determine whether 2,4-Dand Monuron were present in an unchanged form at the endof the absorption period one trial was carried out in which theradioactivity was extracted in 80% ethanol. The treatmentconsisted of 1.0 g of roots in 25 ml of 1 tuM 2,4-D-"C, or 10pM Monuron-'4C, and absorption occurred for 4 hr. Followingwashing, the roots were placed in 25 ml of boiling 80% ethanoland boiled for 1 min. After standing overnight the sampleswere ground, filtered, and the extract was evaporated to dry-ness. The residue remaining after evaporation was taken up in3 ml of 95% ethanol. The dried root residues and 1 ml samplesof the concentrated extracts were placed in separate planchetsand the radioactivity counted in a windowless gas flow GMcounter.Each extract was also chromatographed. For 2,4-D extracts,
the chromatogram was developed in isopropanol-20% (w/v)ammonium acetate-glacial acetic acid (180:20:1 v/v) and forMonuron extracts in 1 -butanol-glacial acetic acid-water (4: 1: 1v/v). The location of the radioactive spots was determined byexposing the chromatograms to Kodak blue brand medicalx-ray film.
RESULTS
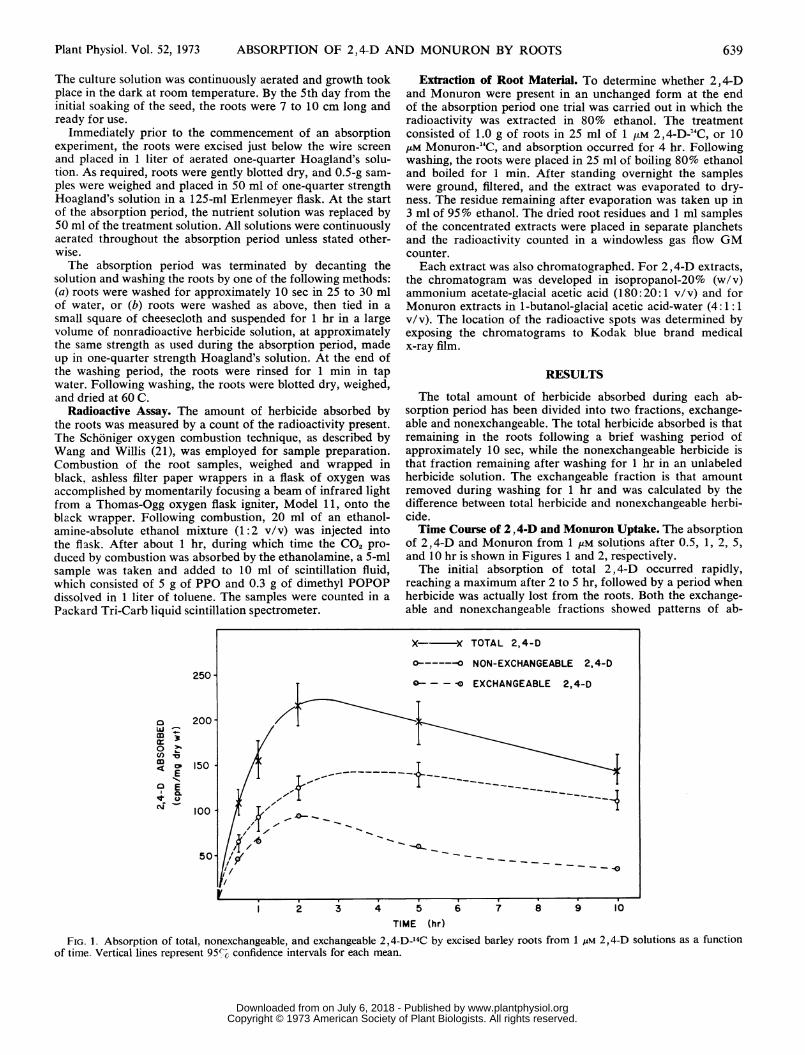
The total amount of herbicide absorbed during each ab-sorption period has been divided into two fractions, exchange-able and nonexchangeable. The total herbicide absorbed is thatremaining in the roots following a brief washing period ofapproximately 10 sec, while the nonexchangeable herbicide isthat fraction remaining after washing for 1 hr in an unlabeledherbicide solution. The exchangeable fraction is that amountremoved during washing for 1 hr and was calculated by thedifference between total herbicide and nonexchangeable herbi-cide.Time Course of 2, 4-D and Monuron Uptake. The absorption
of 2,4-D and Monuron from 1 ,uM solutions after 0.5, 1, 2, 5,and 10 hr is shown in Figures 1 and 2, respectively.The initial absorption of total 2, 4-D occurred rapidly,
reaching a maximum after 2 to 5 hr, followed by a period whenherbicide was actually lost from the roots. Both the exchange-able and nonexchangeable fractions showed patterns of ab-
TIME (hr)
FIG. 1. Absorption of total, nonexchangeable, and exchangeable 2,4-D-14C by excised barley roots from 1 liM 2,4-D solutions as a functionof time. Vertical lines represent 95%G confidence intervals for each mean.
Plant Physiol. Vol. 52, 1973 639
IliUI
www.plantphysiol.orgon July 6, 2018 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.
DONALDSON, BAYER, AND LEONARD
7-
6-
5-
4-
3.
2-
-
22 3
I 1
4 5
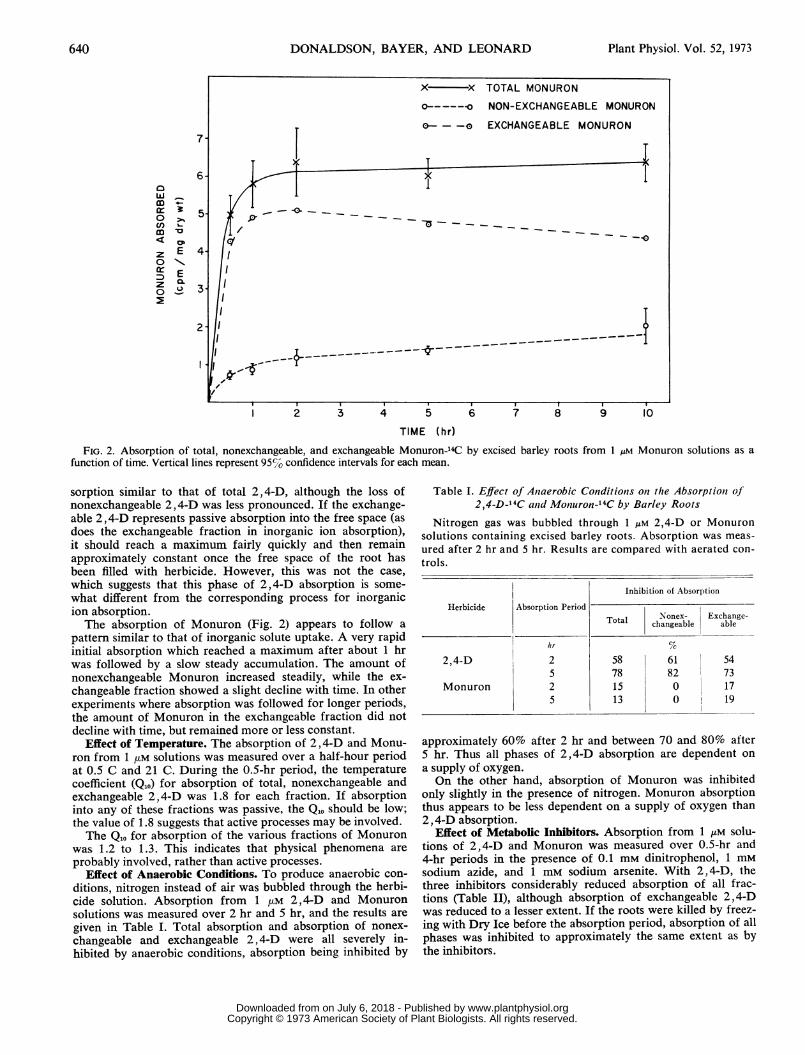
TIME (hr)FIG. 2. Absorption of total, nonexchangeable, and exchangeable Monuron-14C by excised barley roots from 1 AM Monuron solutions as a
function of time. Vertical lines represent 95% confidence intervals for each mean.
sorption similar to that of total 2,4-D, although the loss ofnonexchangeable 2,4-D was less pronounced. If the exchange-able 2,4-D represents passive absorption into the free space (asdoes the exchangeable fraction in inorganic ion absorption),it should reach a maximum fairly quickly and then remainapproximately constant once the free space of the root hasbeen filled with herbicide. However, this was not the case,
which suggests that this phase of 2,4-D absorption is some-
what different from the corresponding process for inorganicion absorption.The absorption of Monuron (Fig. 2) appears to follow a
pattern similar to that of inorganic solute uptake. A very rapidinitial absorption which reached a maximum after about 1 hrwas followed by a slow steady accumulation. The amount ofnonexchangeable Monuron increased steadily, while the ex-changeable fraction showed a slight decline with time. In otherexperiments where absorption was followed for longer periods,the amount of Monuron in the exchangeable fraction did notdecline with time, but remained more or less constant.
Effect of Temperature. The absorption of 2,4-D and Monu-ron from 1 /-M solutions was measured over a half-hour periodat 0.5 C and 21 C. During the 0.5-hr period, the temperaturecoefficient (Q1o) for absorption of total, nonexchangeable andexchangeable 2,4-D was 1.8 for each fraction. If absorptioninto any of these fractions was passive, the Q1o should be low;the value of 1.8 suggests that active processes may be involved.The Q1o for absorption of the various fractions of Monuron
was 1.2 to 1.3. This indicates that physical phenomena are
probably involved, rather than active processes.Effect of Anaerobic Conditions. To produce anaerobic con-
ditions, nitrogen instead of air was bubbled through the herbi-cide solution. Absorption from 1 tcM 2, 4-D and Monuronsolutions was measured over 2 hr and 5 hr, and the results are
given in Table I. Total absorption and absorption of nonex-
changeable and exchangeable 2, 4-D were all severely in-hibited by anaerobic conditions, absorption being inhibited by
Table I. Effect of Aniaerobic Conditionis on thle Absorptioni of2,4-D-14C anid Moniuron-' 4C by Barley Roots
Nitrogen gas was bubbled through 1 ,uM 2,4-D or Monuronsolutions containing excised barley roots. Absorption was meas-ured after 2 hr and 5 hr. Results are compared with aerated con-trols.
Inhibition of Absorption
Herbicide Absorption PeriodTtl Nonex- Exchange-Total changeable able
hr %
2,4-D 2 58 61 545 78 82 73
Monuron 2 15 0 175 13 0 a 19
approximately 60% after 2 hr and between 70 and 80% after5 hr. Thus all phases of 2,4-D absorption are dependent on
a supply of oxygen.On the other hand, absorption of Monuron was inhibited
only slightly in the presence of nitrogen. Monuron absorptionthus appears to be less dependent on a supply of oxygen than2, 4-D absorption.
Effect of Metabolic Inhibitors. Absorption from 1 uM solu-tions of 2,4-D and Monuron was measured over 0.5-hr and4-hr periods in the presence of 0.1 mm dinitrophenol, 1 mMsodium azide, and 1 mm sodium arsenite. With 2,4-D, thethree inhibitors considerably reduced absorption of all frac-tions (Table II), although absorption of exchangeable 2,4-Dwas reduced to a lesser extent. If the roots were killed by freez-ing with Dry Ice before the absorption period, absorption of all
phases was inhibited to approximately the same extent as bythe inhibitors.
awcr
0C')m
z0cr
z0
4-
0'
QL-NNE0.u
X X TOTAL MONURON
o------o NON-EXCHANGEABLE MONURON
o- --o EXCHANGEABLE MONURON
ST i f~~~~/X
9 p-v
1-/-- _
6 7 8 9 10
Plant Physiol. Vol. 52, 1973640
www.plantphysiol.orgon July 6, 2018 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.

ABSORPTION OF 2,4-D AND MONURON BY ROOTS
Table II. Effect of Several Metabolic Inihibitors anid of KillingRoots by Freeziitg oni Absorptioni of 2,4-D-'4C antd
Monuiironi-' 4C by Barley RootsThe excised barley roots were supplied with 1 ,M 2,4-D or
Monuron. Absorption was measured after 0 5 hr and 4 hr. Resultsare compared with living roots without metabolic inhibition.
Inhibition of Absorption
Root Treatment Total \onex- Exchange-
O.5 hr 4 hr 0.5 hr 4 hr '0.5 hr 4 hr
2,4-D IDinitrophenol, 0.1 mm 63 68 75 77 40 43Sodium azide, 1 mm 74 81 86 88 53 63Sodium arsenite, 1 mm 80 89 91 93 60 77Dead roots 87 90 97 97 63 41
MonuronDinitrophenol, 0.1 mm 0 4 27 0 0 7Sodium azide,1mMI 0 23 24 3 0 28Sodium arsenite, 1 mm 0 0 0 0 0 6Dead roots 0 0 0 0 0 5
Monuron absorption was affected to only a limited extentby the three inhibitors. Even killing the roots by freezing withDry Ice did not inhibit absorption. Monuron absorption, there-fore, is apparently not dependent on a supply of metabolicenergy, whereas energy is essential for 2, 4-D absorption.
Effect of External Concentration of Herbicide. Absorptionof 2, 4-D from solutions ranging in concentration from 0.1 uMto 10 aM was determined for 0.5-hr and 4-hr periods. Theabsorption isotherms, shown in Figure 3 for the 4-hr period,indicate a hyperbolic relationship between absorption and ex-
'-
a 3:
o E1(nax o<o
CX o,v E-
r* -
ternal concentration for the three fractions of 2, 4-D. If dif-fusion or mass flow is involved, a linear relationship betweenabsorption and external concentration would be expected.However, if adsorption or an active process involving a carrieris the means whereby absorption occurs, the relationship be-tween uptake and external concentration would be hyper-bolic; i.e. as the absorption sites or the carrier system becomesaturated, absorption becomes independent of external con-centration.The absorption of Monuron from solutions ranging in con-
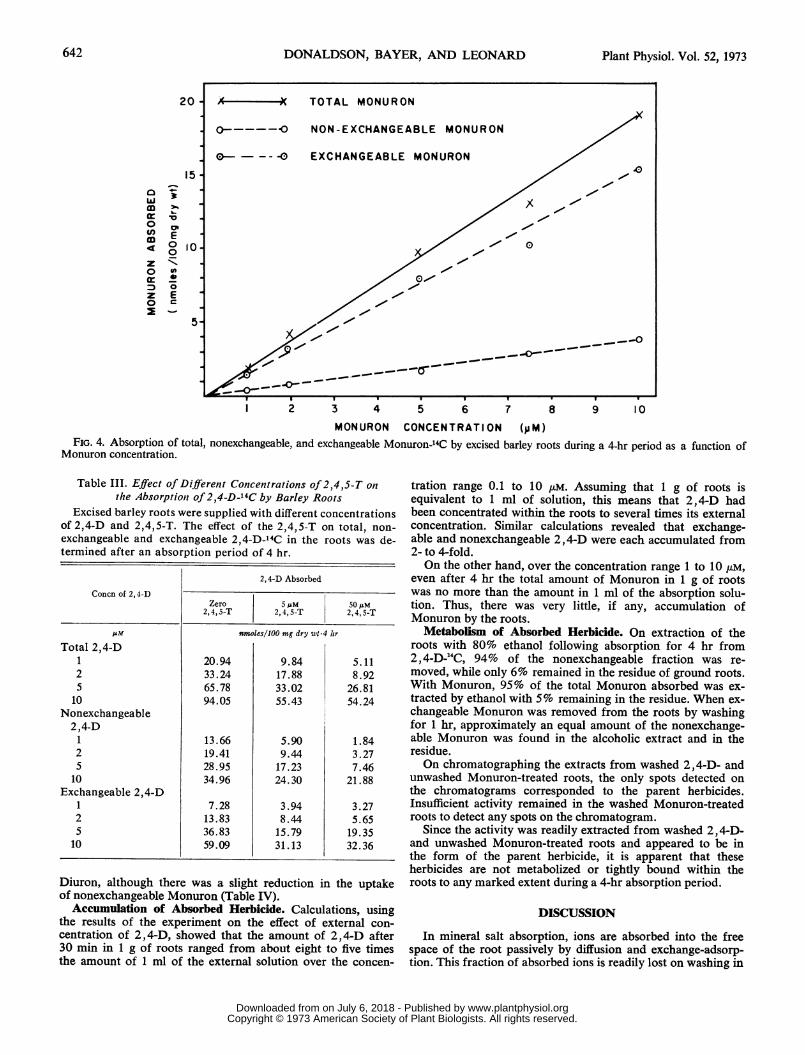
centration from 1 Mm to 10 ,M was also determined for 0.5-hrand 4-hr periods. In this case, the results shown in Figure 4for the 4-hr period indicate a linear relationship between ab-sorption and the external concentration for the various Mon-uron fractions. This suggests that diffusion may be the mainmode of entry into the roots.
Effect of Compounds of Similar Structure. The absorptionof 2, 4-D over a 4-hr period from solutions ranging in con-centration from 1 /cM to 10 Mm was determined in the presenceof various compounds. When 2,4,5-T at 5 jtM or 50 utM wasadded, the absorption of 2,4-D was severely inhibited (TableIII). By using methods similar to those used in studying in-hibition of enzyme reactions it can be determined whether theinhibition of absorption of one compound by another is com-petitive or noncompetitive. A plot of the reciprocal of rate ofabsorption against the reciprocal of 2,4-D concentration atthe different 2,4,5-T concentrations shows that the inhibitionis competitive in nature (Fig. 5).With IAA at 50 ,uM absorption of 2,4-D was inhibited and
this also appeared to be competitive. In the presence of 5 Mmor 50 /AM DL-tryptophan the absorption of 2,4-D was not in-hibited. With Monuron, absorption was studied over a 4-hrperiod in the presence of 5 gm and 50 tLM 3-(3 , 4-dichloro-phenyl)-1 , 1-dimethylurea (Diuron). Absorption of total andexchangeable Monuron showed no inhibition in the presence of
2 3 4 5 6 7 8 9 10
2, 4 - D CONCENTRATION ( pM)FIG. 3. Absorption of total, nonexchangeable, and exchangeable 2, 4-D-'4C by excised barley roots during a 4-hr period as a function of 2,4-D
concentration.
Plant Physiol. Vol. 52, 1973 641
www.plantphysiol.orgon July 6, 2018 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.
DONALDSON, BAYER, AND LEONARD
20-
aw
0
Ur0
z
o
z0
15
-
O 10
o
0
N.
(0
0
EC
5.
I 2 3 4 5 6 7 8 9 10
MONURON CONCENTRATION (y M)FIG. 4. Absorption of total, nonexchangeable, and exchangeable Monuron-'4C by excised barley roots during a 4-hr period as a function of
Monuron concentration.
Table III. Effect of Different Concentrations of 2,4,5-T oznthe Absorptiont of 2,4-D-'4C by Barley Roots
Excised barley roots were supplied with different concentrationsof 2,4-D and 2,4,5-T. The effect of the 2,4,5-T on total, non-
exchangeable and exchangeable 2,4-D-'4C in the roots was de-termined after an absorption period of 4 hr.
2, 4-D Absorbed
Concn of 2,4-DZero 5 JM 50 Mm
2,4,5-T 2,4,5-T 2,4,5-T
AM nmoles/100 mg dry wt-4 hr
Total 2,4-D1 20.94 9.84 5.112 33.24 17.88 8.925 65.78 33.02 26.81
10 94.05 55.43 54.24Nonexchangeable2,4-D1 13.66 5.90 1.842 19.41 9.44 3.275 28.95 17.23 7.46
10 34.96 24.30 21.88Exchangeable 2,4-D
1 7.28 3.94 3.272 13.83 8.44 5.655 36.83 15.79 19.35
10 59.09 31.13 32.36
Diuron, although there was a slight reduction in the uptakeof nonexchangeable Monuron (Table IV).
Accumulation of Absorbed Herbicide. Calculations, usingthe results of the experiment on the effect of external con-centration of 2,4-D, showed that the amount of 2, 4-D after30 min in 1 g of roots ranged from about eight to five timesthe amount of 1 ml of the external solution over the concen-
tration range 0.1 to 10 [LM. Assuming that 1 g of roots isequivalent to 1 ml of solution, this means that 2, 4-D hadbeen concentrated within the roots to several times its externalconcentration. Similar calculations revealed that exchange-able and nonexchangeable 2,4-D were each accumulated from2- to 4-fold.On the other hand, over the concentration range 1 to 10 /jM,
even after 4 hr the total amount of Monuron in 1 g of rootswas no more than the amount in 1 ml of the absorption solu-tion. Thus, there was very little, if any, accumulation ofMonuron by the roots.
Metabolism of Absorbed Herbicide. On extraction of theroots with 80% ethanol following absorption for 4 hr from2, 4-D-'4C, 94% of the nonexchangeable fraction was re-moved, while only 6% remained in the residue of ground roots.With Monuron, 95% of the total Monuron absorbed was ex-tracted by ethanol with 5% remaining in the residue. When ex-changeable Monuron was removed from the roots by washingfor 1 hr, approximately an equal amount of the nonexchange-able Monuron was found in the alcoholic extract and in theresidue.On chromatographing the extracts from washed 2,4-D- and
unwashed Monuron-treated roots, the only spots detected onthe chromatograms corresponded to the parent herbicides.Insufficient activity remained in the washed Monuron-treatedroots to detect any spots on the chromatogram.
Since the activity was readily extracted from washed 2, 4-D-and unwashed Monuron-treated roots and appeared to be inthe form of the parent herbicide, it is apparent that theseherbicides are not metabolized or tightly bound within theroots to any marked extent during a 4-hr absorption period.
DISCUSSION
In mineral salt absorption, ions are absorbed into the freespace of the root passively by diffusion and exchange-adsorp-tion. This fraction of absorbed ions is readily lost on washing in
A-X X TOTAL MONURON
0 -----.O NON-EXCHANGEABLE MONURON
0---- e- EXCHANGEABLE MONURON
-0,
X _____-0 1-~~~~~~~o
4 // _r____- I~~~~~~~~4D
642 Plant Physiol. Vol. 52, 1973
www.plantphysiol.orgon July 6, 2018 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.

ABSORPTION OF 2,4-D AND MONURON BY ROOTS
2020 20-
10- 2,z4,5T 100. / t2,4,5-T
02 04 06 08 0 02 04 06 08 1-0 0-2 0-4 06 08 1.0
SFIG. 5. Effect of 2, 4, 5-T on the absorption of total, nonexchangeable and exchangeable 2,4-D-14C by excised barley roots during a 4-hr period,
plotted as the reciprocal of the rate of absorption against the reciprocal of 2, 4-D concentration. V equals nmoles of 2, 4-D absorbed per mg drymatter in 4 hr. S is the AM concentration of 2, 4-D.
Table IV. Effect of Differenit Concenitrations of Diuron on theAbsorption of Monzuroni-'4C by Barley Roots
Excised barley roots were supplied with different concentrationsof Monuron and Diuron. The effect of Diuron on total, nonex-changeable, and exchangeable Monuron-14C in the roots wasdetermined after an absorption period of 4 hr.
Concn ofMonurononuron AbsorbedConcn of Mlonuron
Zero Diuron 5 jSAe Diuron 50 jm Diuron
;AM nmtoles/lOO mg dry wI 4 hr
Total Monuron1 1.94 1.97 1.852 3.93 3.65 3.945 9.78 9.79 10.3110 20.67 20.62 19.32
NonexchangeableMonuron1 0.52 0.42 0.392 0.89 0.86 0.875 2.37 2.50 2.13
10 5.41 5.01 4.12Exchangeable Monuron
1 1.42 1.55 1.462 3.04 2.79 3.075 7.41 7.29 8.1810 15.26 15.61 15.20
a solution of an exchangeable ion and is frequently referred toas the exchangeable fraction. Ions held in the inner space arenot lost on washing and are called the nonexchangeable frac-tion. Absorption of this fraction is usually considered to be anactive process which is metabolically controlled. On the otherhand, diffusion and adsorption are physical phenomena and,
thus, absorption of the exchangeable fraction should be in-dependent of a supply of metabolic energy.To demonstrate that a compound is being absorbed by an
active mechanism, it is usually required that the followingconditions be fulfilled: (a) the temperature coefficient of ab-sorption should be high, i.e., a Q., of 2 or more; (b) oxygenshould be required, while anaerobic conditions should severelyinhibit absorption; (c) the rate of absorption should not bea linear function of the external concentration but rather hy-perbolic; (d) compounds of similar structure should competefor absorption in a manner similar to competitive inhibitionof enzyme activity; (e) inhibition of absorption should bepossible with a variety of enzyme inhibitors; (f) the absorbedcompound should be accumulable against a concentrationgradient, resulting in a higher concentration within the tis-sue than in the external solution, and be present within the cellin an unchanged form.On considering the absorption of 2, 4-D into the non-
exchangeable and the exchangeable fractions, it is seen thatall the above criteria are satisfied. This indicates that energyis required for absorption into both fractions, which is unex-pected as absorption of the exchangeable fraction into the freespace should be energy-independent. Absorption of the ex-changeable fraction would be expected to show an initial rapiduptake, after which the amount absorbed would remain con-stant with increasing absorption periods, while the total amountabsorbed gradually increased. Thus, the percentage of ex-changeable 2,4-D would gradually decrease. However, duringthe first 2 hr, the exchangeable 2,4-D is a constant fraction(40%) of the total herbicide absorbed (Fig. 1). This may bedue to the nonexchangeable fraction contributing to the ex-changeable fraction during the 1-hr wash period.
In preliminary experiments on the length of the washingperiod, most loss (the exchangeable fraction) occurred within15 to 30 min, but a slow loss continued over the next 30 min.
Plant Physiol. Vol. 52, 1973 643
www.plantphysiol.orgon July 6, 2018 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.

DONALDSON, BAYER, AND LEONARD
With inorganic ions, 30 min washing in a solution of an ex-changeable ion is sufficient to completely remove all adsorbedions in the free space (8). Foy and Yamaguchi (9) found thatover 90% of the 2, 4-D absorbed by excised barley roots during0.5 hr could be removed by washing for 5 hr. As 2,4-D wasstill being lost slowly after 1 hr in the experiments described inthis paper, it is possible that prolonged washing would havecompletely removed all the chemical. This suggests that allthe 2,4-D in the roots may be slowly exchangeable, and thatthere is no distinct nonexchangeable fraction, i.e. a singlemechanism of absorption may be involved. If this is the case,washing for 1 hr would remove only a portion of the herbicide,thus dividing the single mechanism into two apparent "phases"of absorption. Also, if a single mechanism is involved, thevarious criteria studied would have similar effects on bothapparent phases of this mechanism. This was found to be thecase.The requirement for energy suggests that uptake of 2,4-D
is an active process. However, the failure to distinguish apassive phase of absorption into the free space suggests that2,4-D uptake may be different from uptake of inorganic ions.It is possible that there is both active and passive uptake butthe cells may be affected by the herbicide which then "leaks"out and contributes to the exchangeable fraction. Alternatively,2,4-D could be absorbed by an adsorption mechanism andmetabolic energy is required to maintain the integrity of mem-branes and other surfaces on which adsorption could occur.From their experiments, Foy and Yamaguchi (9) concluded thatloss on washing was a desorption process and, therefore, thatabsorption must be an adsorption process. Crafts (3) con-siders 2 ,4-D may be adsorbed at the cell wall cytoplasm inter-face and suggests that this interface may be under the influenceof living cytoplasm and can reversibly bind and release com-pounds. Nooden (16) found that binding of maleic hydrazideby corn roots requires metabolic energy and suggests thatbinding is an active process. Thus, factors such as low tem-perature, metabolic inhibitors, and anaerobic conditions whichreduce the supply of energy within the cell would reduce itscapacity to adsorb 2, 4-D.The hyperbolic relationship between absorption and external
concentration and competition for absorption by similar com-pounds can apply equally to physical adsorption and activeaccumulation. Also, since physical adsorption can result inmultilayer formation (1), the concentration of adsorbed com-pound at the adsorbing surface can be greater than its con-centration in the ambient solution.
According to the Crafts-Broyer theory of salt accumulation(4), ions which are accumulated by the cortex, "leak" out of thecells within the stele because of a less favorable environment,due to lower oxygen levels. If 2,4-D is actively accumulatedby the cells of the cortex, it should also leak out inside thestele and move up in the xylem. However, at low concentra-tions this does not happen. Therefore, it would appear that2,4-D either does not enter the cortical cells or, if it does, itis bound to some cell constituent, perhaps by adsorption, andis not capable of leaking out. At high concentrations 2,4-Dapplied to the roots can move up in the xylem (6). This maybe due to a saturation of the adsorption sites so that excess2,4-D leaks out of the cells in the stele and moves up in thexylem.
In the light of the above experimental evidence, a possibleconclusion is that 2,4-D is absorbed by roots by an adsorptionmechanism and that energy is required to maintain the in-tegrity of the adsorbing surfaces of the cell.
In contrast to absorption of 2,4-D, the absorption of ex-changeable and nonexchangeable Monuron does not appear tobe an active process, as metabolic energy was not required.With exchangeable Monuron, the linear increase in absorption
with increasing concentration, and the lack of competition byDiuron suggests that a passive diffusion mechanism is involvedrather than adsorption. Since between 80 and 90% of the totalMonuron absorbed is in the exchangeable fraction, diffusion isapparently the main means of absorption of this herbicide.As herbicide absorbed by diffusion would be lost during
washing, absorption into the nonexchangeable fraction prob-ably occurs by a passive mechanism other than diffusion. Al-though there was a linear increase with increasing concentra-tion, slight competition by Diuron was observed and thus theuptake of nonexchangeable Monuron could be due to ad-sorption.
Calculation of the amount of exchangeable Monuron in 1 gof roots indicates that it is about the same as the amount in 1ml of the external solution. If absorption into this fraction isby diffusion, accumulation would not occur and, therefore,most of the root volume is accessible to Monuron. Most cal-culations of the free space of tissues give values between 8and 14% (12, 14). Thus, Monuron must be penetrating intomore than the free space of the roots. As energy is not requiredfor absorption, the Monuron molecule, being relatively non-polar, may be able to "dissolve" in the lipid of the cell mem-brane and passively penetrate the cell. Once inside the cell, itcould move through the cortex to the stele and from there tothe top of the plant via the transpiration stream. This wouldexplain how Monuron is able to bypass the Casparian strip ofthe endodermis.
In their studies on the absorption of herbicides by roots,Crafts and Yamaguchi (5) postulated that Monuron must enterthe living cells of the root to be able to cross from the cortexto the xylem. But from foliar applications of this herbicide,there is no evidence of penetration into cells and subsequentmovement in the symplast. However, Monuron is able to inhibitphotosynthesis (2) and, therefore, it must enter the cells ofleaves, although perhaps at extremely low concentrations.
Acknowledgment-Appreciation is expressed to E. I. du Pont de Nemours andCompany for the supply of radioactive Monuron.
LITERATURE CITED
1. ADAMSON, A. IV. 1960. Physical Chemistry of Surfaces. Interscience Publishers,Inc., New York.
2. COOKE, A. R. 1956. A possible mechanism of action of the urea type herbi-cides. Weeds 4: 397-398.
3. CRAFTS, A. S. 1961. Translocation in Plants. Holt, Rinehart and Winston, NewYork.
4. CRAFTS, A. S. AND T. C. BROYER. 1938. 'Migration of salts and water intoxylem of the roots of higher plants. Amer. J. Bot. 25: 529-535.
5. CRAFTS, A. S. AN-D S. YA.MAGUCHI. 1960. Absorption of herbicides by roots.Amer. J. Bot. 47: 248-255.
6. CRAFTS, A. S. AND S. YAMAGUCHI. 1964. The Autoradiography of PlantMIaterials. Univ. of California, Division of Agricultural Sciences, Manual35.
7. EPSTEIN, E. 1961. The essential role of calcium in selective cation transport byplant cells. Plant Physiol. 36: 437-444.
8. EPSTEIN, E., WV. E. SCHnID, AN-D D. W. RAIN_S. 1963. Significance and tech-nique of short term experiments on solute absorption by plant tissues. PlantCell Physiol. 4: 79-84.
9. Foy, C. L. AND S. YANIAGUCHI. 1964. Mechanisms of root absorption of organicmolecules. In: J. Hacskaylo, ed., Absorption and Translocation of OrganicSubstances in Plants. 7th Annual Symposium Southern Sec., Amer. Soc.Plant Physiol. pp. 5-28.
10. HANCOCK, J. G. 1969. Uptake of 3-o-methylglucose by healthy and Hypo-myces-infected squash hypocotyls. Plant Physiol. 44: 1267-1272.
11. HOAGLAND, D. R. AND D. I. ARNON. 1950. The water culture method for grow-ing plants without soil. Calif. Agr. Expt. Sta. Circ. 347, Revised by D. I.Arnon.
12. INGELSTEN, B. AXD B. HYLMO. 1961. Apparent free space and surface filmdetermined by a centrifugation method. Plhysiol. Plant. 14: 157-170.
13. ISENSEE, A. R., G. E. JONES A-ND B. C. TU-RNER. 1971. Root absorption andtranslocation of picloram by oats and soybeans. Weed Sci. 19: 727-731.
14. LEvIrr, J. 1957. The significance of "Apparent Free Space" (AFS) in ionabsorption. Physiol. Plant. 10: 882-888.
15. NISSEN, P. AND A. A. BENSON. 1964. Active transport of choline sulfate bybarley roots. Plant Physiol. 39: 586-589.
644 Plant Physiol. Vol. 52, 1973
www.plantphysiol.orgon July 6, 2018 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.

Plant Physiol. Vol. 52, 1973 ABSORPTION OF 2, 4-D AND MONURON BY ROOTS 645
16. NOODEN, L. D. 1970. 'Metabolism and binding of 14C-maleic hydrazide. Plant 19. RAO, A. V., E. FALLIN, AND S. C. FANG. 1966. Comparative study of uptakePhysiol. 45: 46-52. and cellular distribution of 203Hg-labeled phenyl-mercuric acetate and
17. POOLE, R. J. AND K. V. THIMAAN'N-. 1964. Uptake of indole-3-acetic acid and mercuric acetate by pea roots. Plant Physiol. 41: 443-446.indole-3-acetonitrile by Avena coleoptile sections. Plant Physiol. 39: 98- 20. REINHOLD, L. 1954. The uptake of indole-3-acetic acid by pea epicotyl seg-103. ments and carrot disks. New Phytol. 53: 217-239.
18. PRASAD, R. AND G. E. BLACKNIAN. 1965. Studies in the physiological action of 21. WANG, C. H. AND D. L. WILLIS. 1965. Radiotracer Methodology in2,2-dichloropropionic acid. III. Factors affecting the level of accumulation Biological Sciences. Prentice-Hall, Inc., Englewood Cliffs, N.J.and mode of action. J. Exp. Bot. 16; 545-568. 22. YAM1AGECHI, S. 1965. Analysis of 2,4-D transport. Hilgardia 36(9): 349-378.
www.plantphysiol.orgon July 6, 2018 - Published by Downloaded from Copyright © 1973 American Society of Plant Biologists. All rights reserved.