20years of leptin: role of leptin in cardiomyocyte ... · minireview 20 years of leptin: role of...
TRANSCRIPT

Life Sciences xxx (2015) xxx–xxx
LFS-14314; No of Pages 9
Contents lists available at ScienceDirect
Life Sciences
j ourna l homepage: www.e lsev ie r .com/ locate / l i fesc ie
Minireview
20 years of leptin: Role of leptin in cardiomyocyte physiologyand physiopathology
Feijóo-BandínS. a, PortolésM. b, Roselló-LletíE. b, RiveraM. b, González-JuanateyJ.R. a, LagoF. a,⁎a Cellular and Molecular Cardiology Research Unit and Department of the Institute of Biomedical Research (IDIS), University Clinical Hospital (CHUS-SERGAS), Santiago de Compostela, Spainb Cardiology Unit, Research Centre, Hospital Universitario La Fe, Valencia, Spain
⁎ Corresponding author at: Laboratorio 7, Instituto dSantiago de Compostela (IDIS), Hospital Clínico Univer15706 Santiago de Compostela, Spain. Tel.: +34 981 950
E-mail address: [email protected] (F. Lago)
http://dx.doi.org/10.1016/j.lfs.2015.02.0160024-3205/© 2015 Elsevier Inc. All rights reserved.
Please cite this article as: S. Feijóo-Bandín, e(2015), http://dx.doi.org/10.1016/j.lfs.2015.0
a b s t r a c t
a r t i c l e i n f oArticle history:Received 29 January 2015Accepted 14 February 2015Available online xxxx
Keywords:CardiomyocytesLeptinMetabolismViabilityContractile function
Since the discovery of leptin in 1994 by Zhang et al., there have been a number of reports showing its implicationin the development of a wide range of cardiovascular diseases. However, there exists some controversy abouthow leptin can induce or preserve cardiovascular function, as different authors have found contradictory resultsabout leptin beneficial or detrimental effects in leptin deficient/resistant murine models and in wild type tissueand cardiomyocytes. Here,wewill focus on themain discoveries about the leptin functions at cardiac levelwithinthe last two decades, focusing on its role in cardiac metabolism, remodeling and contractile function.
© 2015 Elsevier Inc. All rights reserved.
Contents
Introduction . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0Leptin signaling and cardiomyocyte metabolism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
Leptin and glucose and fatty acid metabolism . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0Leptin and cardiomyocyte autophagy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
Leptin signaling and cardiac remodeling . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0Leptin and cardiomyocyte hypertrophy . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0Leptin and cardiomyocyte apoptosis . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
Leptin signaling and cardiomyocyte contractility . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0Conclusion . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0Conflict of interest . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0References . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . . 0
Introduction
For many years the human being has adapted its body physiologyto the lack of nourishment in order to survive [10]. Nowadays,overeating and sedentary lifestyle are increasing the prevalenceof obesity worldwide reaching pandemic proportions and becomingan important public health problem [10]. As the prevalence
e Investigaciones Sanitarias desitario, Travesía Choupana s/n,902; fax: +34 981 950 905..
t al., 20 years of leptin: Role2.016
of obesity increases so does the burden of its associated co-morbidities (Fig. 1), which includes type II diabetes, metabolicsyndrome, or cardiovascular diseases such as myocardial infarction,angina pectoris, congestive heart failure, stroke, hypertension, andatrial fibrillation [76].
Many scientists have studied the mechanisms of the energybalance at cellular, tissue, organ and whole body levels in orderto achieve a better knowledge about how to treat or prevent theincidence of obesity and its co-morbidities. One of the mostimportant and widely studied players in the control of energybalance is the hormone leptin [30,71], discovered 20 years ago byZang et al. [111]. Leptin is a 16 kDa protein mostly secreted from
of leptin in cardiomyocyte physiology and physiopathology, Life Sci

Insulin resistanceType II diabetes Heart diseases
DyslipidaemiaCerebrovascular diseaseMetabolic syndrome Pulmonary abnormalities
OsteoarthritisPyschosocial problemsGastrointestinal abnormalitiesCancer
Fig. 1.Morbidities related to obesity [89].
2 S. Feijóo-Bandín et al. / Life Sciences xxx (2015) xxx–xxx
adipose tissue which has a critical role regulating body weight andenergy homeostasis [73,87]. Leptin mediates its effects by bindingto specific leptin receptors (LepRs) expressed in the brain and inperipheral tissues [45]. In the hypothalamus leptin acts as ananorexigenic hormone regulating the melanocortine/neuropeptideY system to reduce food intake, increase energy expenditure, anddecrease body weight [7,71]. However, circulating leptin levels areincreased in obese humans [42], suggesting that obesity may beeither a result or a cause of leptin resistance [19,29,83]. In fact,local effects of leptin can be governed by deregulation of its receptorexpression or downstream signaling components, in particularproteins known to suppress cytokine, and leptin signaling [38].
In an effort to better understand the pathophysiology of humanobesity and its co-morbidities, several rodent models of obesityhave been developed and implemented including high fat dietfeeding and spontaneous mutants of leptin or its receptor such asob/ob (mutant for leptin gene, leptin deficient) and db/db (loss-offunction mutation in the leptin receptor, leptin resistant) miceor Zucker fa/fa rats (loss-of function mutation in the leptin receptor,leptin resistant) [57]. These animals have the common featureof compromised cardiac contractile function [85] and in humanscirculating leptin levels are elevated in vascular and coronaryheart diseases [84], favoring a contemporary perception ofhyperleptinemia as an independent risk factor for the developmentof cardiovascular diseases.
Although the adipose tissue is the main source of leptin, it is alsoproduced by other peripheral tissues, such as the liver, the skeletalmuscle or the kidneys [53,100,102]. Within the heart, leptin andits receptor are abundantly expressed in cardiomyocytes [62,75]where it can regulate the baseline physiology of the heart,including cardiomyocyte contractility, hypertrophy, apoptosis,and metabolism [63]. In this review we will summarize the maindiscoveries about the leptin functions at cardiac level within thelast two decades, focusing on its actions on cardiac tissue andcultured cardiomyocytes.
Leptin signaling and cardiomyocyte metabolism
Leptin and glucose and fatty acid metabolism
The constant pumping activity of the heart requires a permanentsupply of energy [55]. It is widely accepted that fatty acids arethe predominant energy substrates used in the normal adultmyocardium, providing ~70% of adenosine triphosphate (ATP)necessary for the heart to maintain contractile function [55].However, the cardiac metabolic network is highly flexible in usingother substrates when they become abundantly available [47].Thus, depending on the energetic context, the heart is capableof using different substrates (including carbohydrates, lipids,amino acids, and ketone bodies) for ATP production in themitochondria (Fig. 2.A), a concept known as metabolic flexibility ofthe heart [47]. In a normal heart, mitochondria are largely fuelledby acyl-coenzyme A (CoA) and pyruvate, which are the primary
Please cite this article as: S. Feijóo-Bandín, et al., 20 years of leptin: Role(2015), http://dx.doi.org/10.1016/j.lfs.2015.02.016
metabolites of fatty acids and carbohydrates, respectively [47].Energy production from fatty acids requires oxygen consumption,whereas carbohydrate-derived ATP is produced by both glycolysis(oxygen independent) and glucose oxidation [69]. So that, althoughglucose represents a small component of total myocardial energysource, it is the most efficient means of energy production,particularly in conditions of ischemia/hypoxia [69]. During exerciselactate becomes the predominant energy substrate [33], andprolonged fasting or a ketogenic diet increases circulating levels ofketone bodies resulting in an enhanced use by the heart [104]. Theability of the myocardium to switch from one energy substrate toanother (or to use multiple substrates simultaneously) is lost inobesity and diabetes, a state of metabolic inflexibility in which glu-cose transport, glycolysis, and glucose oxidation in cardiomyocytesdecrease, while fatty acid uptake and oxidation increase [20,35,47].
Despite the fact that there exist a number of studies regardingleptin function in modulating systemic and skeletal muscle metabo-lism, little is known about its implication in regulating cardiomyo-cyte metabolism. Some groups have shown that ob/ob and db/dbmice, and fa/fa rats show a metabolic profile in which carbohydrateuptake and utilization are reduced both in cardiac tissue [6,17,27,32,61,92,99] and in cultured cardiomyocyte [27,61] by diminishingglucose transporters GLUT4 translocation to plasma membraneor its protein and mRNA levels, and by reducing pyruvate dehydro-genase and oxoglutarate dehydrogenase activity. In contrast, fattyacid uptake rates are increased in these leptin deficient or resistantanimal models through a mechanism that involves the increasein the expression and membrane localization of the fatty acidtranslocase (FAT)/CD36 and the stimulation of peroxisomeproliferator-activated receptor α (PPARα) signaling (Fig. 2.B) [1,12–14,61,69]. While in leptin deficient/resistant mice the increasein fatty acid uptake is accompanied by an increase in fatty acidoxidation, in fa/fa rats the increase on fatty acid uptake is uncoupledwith the oxidation and yields to lipotoxicity [113]. These findingssuggest a role for the disruption of leptin signaling in the develop-ment of the metabolic inflexibility observed in cardiac metabolismunder pathological conditions, favoring fatty acid utilization anddiminishing cardiac efficiency.
In vitro experiments with cardiac cells have shown that short-term(1 h) leptin treatment has no effect on glucose uptake and oxidationin HL-1 cardiomyocytes and in perfused rat hearts, while fatty aciduptake and oxidation are increased [1,69]. Long term leptin treatment(24 h) also has no effect on glucose uptake and oxidation in HL-1cardiomyocytes and increases fatty acid uptake, however longterm treatment induces a decrease in fatty acid oxidation leading tointracellular lipid accumulation [69], confirming the results obtainedby Zhou et al. in fa/fa rats [113].
Leptin and cardiomyocyte autophagy
Autophagy is an evolutionarily conserved lysosome-mediatedcatabolic pathway that maintains cellular homeostasis through therenewal/recycling of cytoplasmic materials and organelles (such as
of leptin in cardiomyocyte physiology and physiopathology, Life Sci

H+H+
Acetyl-CoA
TCAcycle
I II III IVQ
CytC
NADH
H+
FADH2
ADP ATP
MOM
MIM
Mitochondria
GLUT4Vesicle
Glucose
Glucose Piruvate
GLUT4 CD36
FA
B-oxidation
FA
Lactate
Lactate
Ketonebodies
Ketone bodies
AA
AA
AB
ob/ob db/dbfa/fa
Carbohydratemetabolism
Fatty acidmetabolism
Leptin treatmentin wild type
Fatty acidmetabolism
Fig. 2.Metabolic network in cardiomyocytes. A: catabolism of the different energetic substrates to produce ATP [47]. B: effect of leptin dysfunction (in deficient/resistant murinemodels)and treatment (inwild type) on cardiacmetabolism. GLUT4: glucose transporter 4, CD36: fatty acid translocase, AA: aminoacid, FA: fatty acid,MOM:mitochondrial outermembrane,MIM:mitochondrial inner membrane, CytC: cytochrome C.
3S. Feijóo-Bandín et al. / Life Sciences xxx (2015) xxx–xxx
mitochondria) and protein aggregates that could become toxic to thecell [49]. Such cellular refreshing is particularly important in quies-cent and terminally differentiated cells, like cardiomyocytes, inwhich damaged components are not diluted by cell replication, andin which aging increases its accumulation, making the function ofthe cells less efficient and decreasing their adaptability [96]. Thebreakdown products derived from autophagy have a dual role,providing substrates for both biosynthesis and energy generation,and being of special importance to assure substrate availability andenergetic maintenance under stress [54]. Autophagy can be activatedby a number of diverse stimuli including caloric restriction, oxidativestress, hormones, or other developmental signals [41], and it istypically measured by the stimulation of a post-translationalmodification of microtubule-associated protein 1 light chain 3(LC3I) that increases its electrophoretic mobility due to proteolysisand lipidation (LC3II), the increase in AMP-activated protein kinase(AMPK) and acetyl-CoA carboxylase (ACC) phosphorylation, themammalian target of rapamycin (mTOR) dephosphorylation, theincrease in proteins derived from autophagy-related genes (Atg) 5and 7, and the reduction of the autophagic substrate p62 [59].Deregulation of autophagy in cardiomyocytes is associated withvarious cardiac diseases, such as isquemic heart disease, hyperten-sive heart disease, arrhythmia, isquemia/reperfusion injury or heartfailure [88]. Particularly, cardiomyocytes have extremely highmitochondrial density compared with other tissues due to itscontinuous high demand for energy. And, because of that, they
Please cite this article as: S. Feijóo-Bandín, et al., 20 years of leptin: Role(2015), http://dx.doi.org/10.1016/j.lfs.2015.02.016
need a continuous basal-level autophagic turnover of mitochondria(mitophagy) to assure the proper functioning of the heart [41].
In leptin deficient or resistant mice, it has been shown an increasein autophagy in peripheral tissues such as the liver or the skeletalmuscle, measured by the increase in LC3II leves and the diminutionof p62 [59]. These results could indicate that leptin signaling disrup-tion may induce autophagy; however, external application of leptinalso stimulates autophagy in those tissues [59]. According to this,leptin has been shown to have a pro-autophagic effect in differentcell types, including human cervical carcinoma HeLa cells, humanHCT116 colorectal cancer cells, human U2OS osteosarcoma cells,mouse embryonic fibroblast, rat bone marrow-derived mesenchymalstem cells and piglet intestinal epithelial cells [59,93,101,103]. Never-theless, leptin treatment also seems to protect from autophagy progres-sion in human T cells [16].
With respect to leptin effect on cardiac autophagy, it has beenreported that db/db mice show increased levels of myocardial au-tophagy (increased levels of LC3II and decreased of p62) [59], andleptin injection induces a higher increase of autophagy in myo-cardium, effect also observed in liver and skeletal muscle [59].Moreover, leptin injection in C57BL/6 mice also induce an in-crease in autophagy in cardiac tissue, similar to the autophagy in-ducers spermidine, resveratrol or rapamycin [59], as well asleptin treatment of left ventricle cultured cardiomyocytes also in-duces autophagy [43]. Taking all together, those results suggest acomplex relationship between autophagy and leptin signaling,
of leptin in cardiomyocyte physiology and physiopathology, Life Sci

4 S. Feijóo-Bandín et al. / Life Sciences xxx (2015) xxx–xxx
where any deviation from leptin homeostasis may affect autoph-agy progression.
Leptin signaling and cardiac remodeling
Leptin and cardiomyocyte hypertrophy
Cardiac hypertrophy is one of the main ways in whichcardiomyocytes respond to mechanical and neurohormonal stimulito increase their work output and improve cardiac pump function[15]. However, this compensatory mechanism can becomeoverwhelmed by biomechanical stress, thereby resulting in thedevelopment of cardiac diseases such as ischemic heart disease,hypertension, heart failure or valve disease [15]. For instance,chronic excessive accumulation of body fat causes adaptations ofthe cardiovascular system to maintain whole body homeostasissuch as increased cardiac output and a decrease in peripheralresistance [5]. Stroke volume, the major determinant in theincreased cardiac output in the obese patient, increases due to theaugmentation of circulating blood volume, induces an enlargementof the cardiac cavities and increases wall tension, leading to leftventricular hypertrophy (LVH) [5].
Deregulation of the leptin signaling pathway within the hearthas been suggested to be one of themechanisms that cause LVH, howev-er there exist contradictory studies about leptin role in the developmentof LVH. A number of publications have shown a pro-hypertrophic effectof leptin treatment in neonatal rat cardiomyocytes [31,37,39,44,79–81,106,108-110] and in human pediatric ventricular myocytes [58]. Also,it has been shown that the neutralization of LepR using antibodiesabrogates hypertrophy in surviving myocardium after coronary ar-tery ligation in rats [77]. According to this, clinical studies demon-strated a positive correlation between serum leptin levels and leftventricular mass or wall thickness in obese and in insulin resistantpatients [70,74]. The mechanisms described through which leptincan induce cardiomyocyte hypertrophy seem to be quite different:induction of mTOR signaling [108], calcineurin activation and nucle-ar factor of activated T-cells (NFAT) nuclear translocation [80], acti-vation of PPARα signaling [37], mitogen-activated protein kinase14 (p38) activation and translocation into the nuclei [79,109], acti-vation of Rho and actin dynamics [110] or increased intracellularlevels of reactive oxygen species [39,106]. However, it has alsobeen shown that leptin treatment of neonatal rat and HL-1cardiomyocytes has no effect on the development of hypertrophy[75].
On the contrary, mice lacking leptin (ob/ob) or its receptor (db/db)develop LVH when they become morbidly obese [3,4,50,82], whileleptin repletion in ob/ob mice restores left ventricle normal thicknessindependently of body weight [4]. In hearts from diet induced obesemice showing hyperleptinemia, LepR continue to respond to elevatedcirculating or cardiac leptin, which seems to protect from cardiachypertrophy via LepR-induced signal transducer and activator oftranscription 3 (STAT3) activation compared to LepR mutant or db/dbmice [50].
In summary, it is unclear whether cardiac hypertrophy is theconsequence of pro-hypertrophic effects of leptin or rather the resultof a resistance toward leptin's preventive effects on hypertrophiccardiac remodeling.
Leptin and cardiomyocyte apoptosis
The myocardium is comprised of terminally differentiatedcardiomyocytes that are responsible for contractile function and thathave a limited capacity to efficiently regenerate [68]. Maintenance ofcardiac homeostasis depends on cardiomyocyte death and renewal,and excessive loss of cardiomyocytes has been implicated incardiovascular diseases such as myocardial ischemia/reperfusion injury
Please cite this article as: S. Feijóo-Bandín, et al., 20 years of leptin: Role(2015), http://dx.doi.org/10.1016/j.lfs.2015.02.016
and congestive heart failure, situationswhere, with fewermyocytes, theheart is unable to sustain efficient contraction [68]. Among the mainmechanisms that promote loss of cells in the heart is apoptosis, a highlyregulated process by which activation of specific signaling cascadesultimately leads to cell death [68]. An apoptotic cell undergoesstructural changes including cell shrinkage, plasma membraneblebbing, nuclear condensation, and fragmentation of the DNA andnucleus, followed by fragmentation into apoptotic bodies that arequickly removed by phagocytes [68]. Apoptosis is mediated by twocentral pathways: the extrinsic or death receptor pathway, that isinduced via activation of a death domain-containing receptor locatedat the plasma membrane, and the intrinsic or mitochondrial pathway,which is activated by intracellular stress signals such as loss of growthfactors, hypoxia, oxidative stress, or DNA damage (Fig. 3.A) [68]. Theimmediate objectives of apoptotic signaling are the activation ofcaspases and the disabling of mitochondrial function to induce the celldeath [21].
In obese Zucker rats (fa/fa), cardiomyocytes show increased levels ofapoptosis which involve both the extrinsic and the intrinsic apoptoticpathways, pointing to the existence of local and systemic stresses in theZucker fa/fa hearts to induce cell death [48,56]. Similar to Zucker fa/farats, db/db and ob/obmice have increased total cardiac triglyceride levelsand increased cardiomyocyte apoptosis, however, it has been shown thatthe triglyceride accumulation and the high levels of cardiac apoptosis inob/ob mice can be returned toward normal with leptin repletion [3], aswell as in db/dbmice the rescue of cardiac leptin receptors preventsmyo-cardial triglyceride accumulation and improves cardiac function [34], pro-viding a direct role for leptin in preventing excess of cardiac lipidaccumulation and in ameliorating cardiac function. Thesefindings suggestthat increased cell death in these obese models is not just a reflection ofsenescence or injury, but rather specific pathways directly related withleptin signaling disruption (Fig. 3.B). According to this, the generation ofa cardiomyocyte-specific leptin receptor knock-out mouse showed thatleptin signaling disruption exhacerbates cardiac injury in the post-miocardial infarction failing heart by acting directly on cardiomyocytesto increase cardiac hypertrophy, apoptosis, and inflammation, as well asdeleterious changes in cardiac structure, function, and glycolytic metabo-lism [63].
One of the mechanisms that induce apoptosis in the heart of theleptin resistant Zucker rats is the lipoapoptosis. Triacylglycerol contentin nonadipose cells is normally quite low and is maintained within anarrow range, whereas in adipocytes it can vary considerablydepending on the composition and quantity of food intake [97]. Thisreveals a limited ability of nonadipose tissue to accommodate excessof triglycerides and a strict regulation of triglyceride levels in bothadipose and nonadipose cells [97]. In this line, in the Zucker fa/fa heartsa sharp increase in lipid accumulation that leads to cardiomyocyteapoptosis and cardiac dysfunction occurs (Fig. 3.B) [113]. Althoughtriglycerides are themselves harmless, they are hydrolyzed tofatty acyl-CoA, providing increased substrate for synthesis of the pro-apoptotic sphingolipid ceramide, which triggers the intrinsic apoptosispathway in cardiomyocytes [11,72]. In fa/fa Zucker cardiomyocytesthe increase of triglyceride accumulation is correlated with increasedceramide levels and increased DNA damage, leading to lipoapoptosis[113].
Therefore, it could seem clear that at cardiac level leptin must havean anti-apoptotic effect. In several studies it has been shown that leptintreatment protects cultured cardiomyocytes from apoptosis in differentways (Fig. 3.B) [22,28,91,94,95,107,112]. Leptin has been shown toinhibit the mitochondrial permeability transition channel pore (mPTP)opening in murine cardiomyocytes [22,94,95], which is one of themechanisms that triggers apoptosis due to the release of cytochromeC into the cytoplasm [8]. In the subclone of the original clonal cell linederived from embryonic BD1X rat heart tissue (H9c2), leptin treatmentexerts a protective effect against H2O2-induced apoptosis bypreventing the activation of components of the intrinsic pathway of
of leptin in cardiomyocyte physiology and physiopathology, Life Sci

Death receptors(Extrinsic pathway)
DNA damage
ER stress
Oxidative stress
CytoCCytoC
CytoCCytoC
mPTP
Mitochondria(Intrinsic pathway)
Ceramide
BIDtBIDBaxBak
Apoptosis
Apoptosome
Caspase 8
Caspase 9
Caspase 3/7
Lipiddroplet
CytoC
Hypoxia/ReoxigenationHigh glucose
Ca2+
Ca2+Ca2+
Ca2+
Calcium accumulation
Aob/ob db/db
fa/fa
Apoptosis
Apoptosis under:Calcium accumulation
High glucose
B
Leptin treatmentin wild type
Apoptosis induced by:mPTP opening
Hypoxia/ReoxigenationH2o2
Serum deprivationTNFa
NO
NO NO
Fig. 3. Extrinsic and intrinsic apoptosis pathways. A: extrinsic and intrinsic apoptosis pathways and some of themechanism that triggers them in cardiomyocytes [67,68]. B: effect of leptindysfunction (in deficient/resistant murinemodels) and treatment (inwild type) on cardiac apoptosis. BID: BH3 interacting domain death agonist, tDID: truncated BH3 interacting domaindeath agonist, CytoC: cytochrome C, Bax: BCL2-associated X protein, Bak: BCL2-antagonist/killer, mPTP: mitochondrial permeability transition pore, NO: nitric oxide, ER: endoplasmicreticulum.
5S. Feijóo-Bandín et al. / Life Sciences xxx (2015) xxx–xxx
apoptosis by reducing Bax integration in the mitochondrial membraneand cytochrome C release frommitochondria [28] and it also attenuateshypoxia/reoxygenation-induced activation of the intrinsic pathway ofapoptosis [91]. In neonatal rat cardiomyocytes leptin exerts a directanti-apoptotic effect in serum-deprived cardiomyocytes by relievingoxidative stress and inactivating the intrinsic apoptotic pathway [112],and it also abrogates tumor necrosis factorα (TNFα)-induced apoptosisby blocking both the intrinsic mitochondrial pathway of apoptosis andthe extrinsic apoptotic pathway upregulated by TNFα [107].
In the opposite, other research groups have suggested a pro-apoptotic function of leptin at cardiac level under damagingconditions (Fig. 3.B). In the presence of excessive intracellularcalcium accumulation, leptin may contribute to mitochondrialdysfunction by inducing the mPTP opening and the development ofcardiomyocyte apoptosis [60]; also, under high glucose conditions,the inhibition of leptin signaling protects H9c2 cells from mPTPopening [18] suggesting that, depending on the cellular context,leptin signaling may exert protective or detrimental actions oncardiomyocytes viability.
Leptin signaling and cardiomyocyte contractility
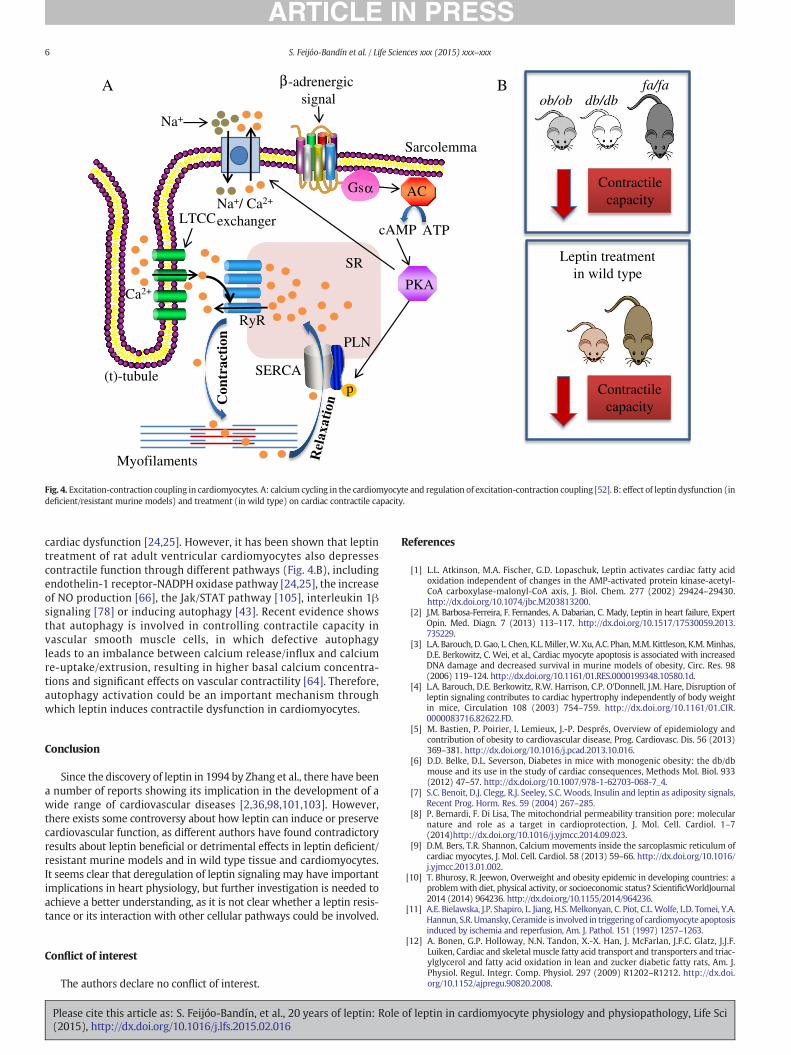
In the heart, Ca2+ influx acts as a multi-functional signal that triggersmuscle contraction, controls action potential duration, and regulates geneexpression [90]. The sarcoplasmic reticulum (SR) in striated muscle is ahighly specialized form of endoplasmic reticulum which surrounds themyofilaments and operates in collaboration with deep invaginations of
Please cite this article as: S. Feijóo-Bandín, et al., 20 years of leptin: Role(2015), http://dx.doi.org/10.1016/j.lfs.2015.02.016
the plasma membrane (or sarcolemma), called transverse (t)-tubules,to regulate the release of calcium from the SR lumen into the cytoplasm,where it regulates myocyte contraction [26]. During cardiac excitation–contraction coupling, β-adrenergic signaling induces the activation ofNa+/Ca2+ exchanger channels through PKA signaling, leading tosarcolemma depolarization [9]. This depolarization stimulates theopening of high-voltage-activated L-type calcium channels (LTCC) in thesarcolemma, generating a Ca2+ current into the cytosol, which inducesCa2+ release from the sarcoplasmic reticulum(SR) via ryanodine receptor(RyR) channels to initiate myocyte contraction (Fig. 4.A) [9]. On thecontrary, sarcoplasmic/endoplasmic reticulum calcium ATPase 2(SERCA2) and phospholamban activation (PLN) have a key role insarcoplasmic reticulumCa2+ sequestration from the cytosol andmyocyterelaxation [46]. Abnormalities in SR Ca2+ cycling are hallmarks of heartdiseases such as heart failure or atrial fibrillation and contribute to thepathophysiology and progression of these diseases [40,46].
Ob/ob and db/db mice, and fa/fa rats develop cardiac contractiledysfunction, showing slowed intracellular Ca2+ decay rate and re-duced contractile capacity in myocytes (Fig. 4.B) [24,25,51,65,85,86]. SERCA2 and Na+/Ca2+ exchanger channel activity aredepressed in leptin deficient mice [23], while leptin treatment ofcardiomyocytes from ob/obmice improves β-adrenergic responsive-ness with increased protein expression of the stimulatory guaninenucleotide-binding protein α subunit (Gsα), enhanced PKA activityand enhanced phosphorylation of PLN [65]. These data provide aconvincing link between adequate leptin signaling and cardiac func-tion and suggest a mechanism by which leptin deficiencymay lead to
of leptin in cardiomyocyte physiology and physiopathology, Life Sci
SR
(t)-tubule
Sarcolemma
Myofilaments
Ca2+
SERCA
RyR
LTCCNa+/ Ca2+
exchanger
Na+
p
-adrenergic signal
Gs AC
PLN
ATPcAMP
PKA
Con
trac
tion
Bob/ob db/db
fa/fa
Contractilecapacity
Leptin treatmentin wild type
Contractilecapacity
A β
α
Fig. 4. Excitation-contraction coupling in cardiomyocytes. A: calcium cycling in the cardiomyocyte and regulation of excitation-contraction coupling [52]. B: effect of leptin dysfunction (indeficient/resistant murine models) and treatment (in wild type) on cardiac contractile capacity.
6 S. Feijóo-Bandín et al. / Life Sciences xxx (2015) xxx–xxx
cardiac dysfunction [24,25]. However, it has been shown that leptintreatment of rat adult ventricular cardiomyocytes also depressescontractile function through different pathways (Fig. 4.B), includingendothelin-1 receptor-NADPH oxidase pathway [24,25], the increaseof NO production [66], the Jak/STAT pathway [105], interleukin 1βsignaling [78] or inducing autophagy [43]. Recent evidence showsthat autophagy is involved in controlling contractile capacity invascular smooth muscle cells, in which defective autophagyleads to an imbalance between calcium release/influx and calciumre-uptake/extrusion, resulting in higher basal calcium concentra-tions and significant effects on vascular contractility [64]. Therefore,autophagy activation could be an important mechanism throughwhich leptin induces contractile dysfunction in cardiomyocytes.
Conclusion
Since the discovery of leptin in 1994 by Zhang et al., there have beena number of reports showing its implication in the development of awide range of cardiovascular diseases [2,36,98,101,103]. However,there exists some controversy about how leptin can induce or preservecardiovascular function, as different authors have found contradictoryresults about leptin beneficial or detrimental effects in leptin deficient/resistant murine models and in wild type tissue and cardiomyocytes.It seems clear that deregulation of leptin signaling may have importantimplications in heart physiology, but further investigation is needed toachieve a better understanding, as it is not clear whether a leptin resis-tance or its interaction with other cellular pathways could be involved.
Conflict of interest
The authors declare no conflict of interest.
Please cite this article as: S. Feijóo-Bandín, et al., 20 years of leptin: Role(2015), http://dx.doi.org/10.1016/j.lfs.2015.02.016
References
[1] L.L. Atkinson, M.A. Fischer, G.D. Lopaschuk, Leptin activates cardiac fatty acidoxidation independent of changes in the AMP-activated protein kinase-acetyl-CoA carboxylase-malonyl-CoA axis, J. Biol. Chem. 277 (2002) 29424–29430.http://dx.doi.org/10.1074/jbc.M203813200.
[2] J.M. Barbosa-Ferreira, F. Fernandes, A. Dabarian, C. Mady, Leptin in heart failure, ExpertOpin. Med. Diagn. 7 (2013) 113–117. http://dx.doi.org/10.1517/17530059.2013.735229.
[3] L.A. Barouch, D. Gao, L. Chen, K.L.Miller,W. Xu, A.C. Phan,M.M. Kittleson, K.M.Minhas,D.E. Berkowitz, C. Wei, et al., Cardiac myocyte apoptosis is associated with increasedDNA damage and decreased survival in murine models of obesity, Circ. Res. 98(2006) 119–124. http://dx.doi.org/10.1161/01.RES.0000199348.10580.1d.
[4] L.A. Barouch, D.E. Berkowitz, R.W. Harrison, C.P. O'Donnell, J.M. Hare, Disruption ofleptin signaling contributes to cardiac hypertrophy independently of body weightin mice, Circulation 108 (2003) 754–759. http://dx.doi.org/10.1161/01.CIR.0000083716.82622.FD.
[5] M. Bastien, P. Poirier, I. Lemieux, J.-P. Després, Overview of epidemiology andcontribution of obesity to cardiovascular disease, Prog. Cardiovasc. Dis. 56 (2013)369–381. http://dx.doi.org/10.1016/j.pcad.2013.10.016.
[6] D.D. Belke, D.L. Severson, Diabetes in mice with monogenic obesity: the db/dbmouse and its use in the study of cardiac consequences, Methods Mol. Biol. 933(2012) 47–57. http://dx.doi.org/10.1007/978-1-62703-068-7_4.
[7] S.C. Benoit, D.J. Clegg, R.J. Seeley, S.C. Woods, Insulin and leptin as adiposity signals,Recent Prog. Horm. Res. 59 (2004) 267–285.
[8] P. Bernardi, F. Di Lisa, The mitochondrial permeability transition pore: molecularnature and role as a target in cardioprotection, J. Mol. Cell. Cardiol. 1–7(2014)http://dx.doi.org/10.1016/j.yjmcc.2014.09.023.
[9] D.M. Bers, T.R. Shannon, Calcium movements inside the sarcoplasmic reticulum ofcardiac myocytes, J. Mol. Cell. Cardiol. 58 (2013) 59–66. http://dx.doi.org/10.1016/j.yjmcc.2013.01.002.
[10] T. Bhurosy, R. Jeewon, Overweight and obesity epidemic in developing countries: aproblemwith diet, physical activity, or socioeconomic status? ScientificWorldJournal2014 (2014) 964236. http://dx.doi.org/10.1155/2014/964236.
[11] A.E. Bielawska, J.P. Shapiro, L. Jiang, H.S. Melkonyan, C. Piot, C.L.Wolfe, L.D. Tomei, Y.A.Hannun, S.R. Umansky, Ceramide is involved in triggering of cardiomyocyte apoptosisinduced by ischemia and reperfusion, Am. J. Pathol. 151 (1997) 1257–1263.
[12] A. Bonen, G.P. Holloway, N.N. Tandon, X.-X. Han, J. McFarlan, J.F.C. Glatz, J.J.F.Luiken, Cardiac and skeletal muscle fatty acid transport and transporters and triac-ylglycerol and fatty acid oxidation in lean and zucker diabetic fatty rats, Am. J.Physiol. Regul. Integr. Comp. Physiol. 297 (2009) R1202–R1212. http://dx.doi.org/10.1152/ajpregu.90820.2008.
of leptin in cardiomyocyte physiology and physiopathology, Life Sci

7S. Feijóo-Bandín et al. / Life Sciences xxx (2015) xxx–xxx
[13] J. Buchanan, P.K. Mazumder, P. Hu, G. Chakrabarti, M.W. Roberts, J.Y. Ui, R.C.Cooksey, S.E. Litwin, E.D. Abel, Reduced cardiac efficiency and altered substratemetabolism precedes the onset of hyperglycemia and contractile dysfunction intwo mouse models of insulin resistance and obesity, Endocrinology 146 (2005)5341–5349. http://dx.doi.org/10.1210/en.2005-0938.
[14] A.N. Carley, D.L. Severson, Fatty acid metabolism is enhanced in type 2 diabetichearts, Biochim. Biophys. Acta 1734 (2005) 112–126. http://dx.doi.org/10.1016/j.bbalip.2005.03.005.
[15] J.E. Carreño, F. Apablaza, M.P. Ocaranza, J.E. Jalil, Cardiac hypertrophy: molecularand cellular events, Rev. Esp. Cardiol. (English Ed. 59 (2006) 473–486. http://dx.doi.org/10.1016/S1885-5857(06)60796-2.
[16] S. Cassano, V. Pucino, C. La Rocca, C. Procaccini, V. De Rosa, G. Marone, G.Matarese, Leptin modulates autophagy in human CD4 + CD25- conventionalT cells, Metabolism 63 (2014) 1272–1279. http://dx.doi.org/10.1016/j.metabol.2014.06.010.
[17] J. Chatham, Cardiac carbohydrate metabolism in zucker diabetic fatty rats,Cardiovasc. Res. 55 (2002) 104–112. http://dx.doi.org/10.1016/S0008-6363(02)00399-1.
[18] J. Chen, H. Mo, R. Guo, Q. You, R. Huang, K. Wu, Inhibition of the leptin-induced ac-tivation of the p38 MAPK pathway contributes to the protective effects of naringinagainst high glucose-induced injury in H9c2 cardiac cells, Int. J. Mol. Med. 33(2014) 605–612. http://dx.doi.org/10.3892/ijmm.2014.1614.
[19] R.V. Considine, M.K. Sinha, M.L. Heiman, A. Kriauciunas, T.W. Stephens, M.R. Nyce,J.P. Ohannesian, C.C. Marco, L.J. McKee, T.L. Bauer, Serum immunoreactive-leptinconcentrations in normal-weight and obese humans, N. Engl. J. Med. 334 (1996)292–295. http://dx.doi.org/10.1097/00019616-199607000-00020.
[20] C. Crewe, M. Kinter, L.I. Szweda, Rapid inhibition of pyruvate dehydrogenase: aninitiating event in high dietary fat-induced loss of metabolic flexibility in theheart, PLoS One 8 (2013) e77280. http://dx.doi.org/10.1371/journal.pone.0077280.
[21] M.T. Crow, K.Mani, Y.J. Nam, R.N. Kitsis, Themitochondrial death pathway and car-diac myocyte apoptosis, Circ. Res. 95 (2004) 957–970. http://dx.doi.org/10.1161/01.RES.0000148632.35500.d9.
[22] R.A. Dixon, S.M. Davidson, A.M. Wynne, D.M. Yellon, C.C.T. Smith, Thecardioprotective actions of leptin are lost in the zucker obese (fa/fa) rat, J.Cardiovasc. Pharmacol. 53 (2009) 311–317. http://dx.doi.org/10.1097/FJC.0b013e31819d6152.
[23] F. Dong, X. Yang, N. Sreejayan, J. Ren, Chromium (D-phenylalanine)3 improvesobesity-induced cardiac contractile defect in ob/ob mice, Obesity (Silver Spring)15 (2007) 2699–2711. http://dx.doi.org/10.1038/oby.2007.322.
[24] F. Dong, X. Zhang, J. Ren, Leptin regulates cardiomyocyte contractile functionthrough endothelin-1 receptor-NADPH oxidase pathway, Hypertension 47(2006) 222–229. http://dx.doi.org/10.1161/01.HYP.0000198555.51645.f1.
[25] F. Dong, X. Zhang, X. Yang, L.B. Esberg, H. Yang, Z. Zhang, B. Culver, J. Ren, Impairedcardiac contractile function in ventricular myocytes from leptin-deficient ob/obobese mice, J. Endocrinol. 188 (2006) 25–36. http://dx.doi.org/10.1677/joe.1.06241.
[26] S. Doroudgar, C.C. Glembotski, New concepts of endoplasmic reticulum function inthe heart: programmed to conserve, J. Mol. Cell. Cardiol. 55 (2013) 85–91. http://dx.doi.org/10.1016/j.yjmcc.2012.10.006.
[27] J. Eckel, A. Wirdeier, L. Herberg, H. Reinauer, Insulin resistance in the heart: studieson isolated cardiocytes of genetically obese zucker rats, Endocrinology 116 (1985)1529–1534. http://dx.doi.org/10.1210/endo-116-4-1529.
[28] M. Eguchi, Y. Liu, E.-J. Shin, G. Sweeney, Leptin protects H9c2 rat cardiomyocytesfrom H2O2-induced apoptosis, FEBS J. 275 (2008) 3136–3144. http://dx.doi.org/10.1111/j.1742-4658.2008.06465.x.
[29] R.C. Frederich, A. Hamann, S. Anderson, B. Löllmann, B.B. Lowell, J.S. Flier, Leptinlevels reflect body lipid content in mice: evidence for diet-induced resistance toleptin action, Nat. Med. 1 (1995) 1311–1314. http://dx.doi.org/10.1038/nm1295-1311.
[30] J.M. Friedman, J.L. Halaas, Leptin and the regulation of body weight in mammals,Nature 395 (1998) 763–770. http://dx.doi.org/10.1038/27376.
[31] X.T. Gan, G. Zhao, C.X. Huang, A.C. Rowe, D.M. Purdham, M. Karmazyn, Identifica-tion of fat mass and obesity associated (FTO) protein expression incardiomyocytes: regulation by leptin and its contribution to leptin-induced hyper-trophy, PLoS One 8 (2013) e74235. http://dx.doi.org/10.1371/journal.pone.0074235.
[32] L.S. Golfman, C.R. Wilson, S. Sharma, M. Burgmaier, M.E. Young, P.H. Guthrie, M.Van Arsdall, J.V. Adrogue, K.K. Brown, H. Taegtmeyer, Activation of PPARgammaenhances myocardial glucose oxidation and improves contractile function in iso-lated working hearts of ZDF rats, Am. J. Physiol. Endocrinol. Metab. 289 (2005)E328–E336. http://dx.doi.org/10.1152/ajpendo.00055.2005.
[33] G.W. Goodwin, H. Taegtmeyer, Improved energy homeostasis of the heart in themetabolic state of exercise, Am. J. Physiol. Heart Circ. Physiol. 279 (2000)H1490–H1501.
[34] M.E. Hall, M.W. Maready, J.E. Hall, D.E. Stec, Rescue of cardiac leptin receptors indb/db mice prevents myocardial triglyceride accumulation, Am. J. Physiol.Endocrinol. Metab. 307 (2014) E316–E325. http://dx.doi.org/10.1152/ajpendo.00005.2014.
[35] L.C. Heather, K. Clarke, Metabolism, hypoxia and the diabetic heart, J. Mol. Cell.Cardiol. 50 (2011) 598–605. http://dx.doi.org/10.1016/j.yjmcc.2011.01.007.
[36] N. Hou, J.-D. Luo, Leptin and cardiovascular diseases, Clin. Exp. Pharmacol. Physiol.38 (2011) 905–913. http://dx.doi.org/10.1111/j.1440-1681.2011.05619.x.
[37] N. Hou, M.-S. Luo, S.-M. Liu, H.-N. Zhang, Q. Xiao, P. Sun, G.-S. Zhang, J.-D. Luo, M.-S.Chen, Leptin induces hypertrophy through activating the peroxisome proliferator-activated receptor α pathway in cultured neonatal rat cardiomyocytes, Clin. Exp.
Please cite this article as: S. Feijóo-Bandín, et al., 20 years of leptin: Role(2015), http://dx.doi.org/10.1016/j.lfs.2015.02.016
Pharmacol. Physiol. 37 (2010) 1087–1095. http://dx.doi.org/10.1111/j.1440-1681.2010.05442.x.
[38] J.K. Howard, J.S. Flier, Attenuation of leptin and insulin signaling by SOCS proteins,Trends Endocrinol. Metab. 17 (2006) 365–371. http://dx.doi.org/10.1016/j.tem.2006.09.007.
[39] T.-P. Hu, F.-P. Xu, Y.-J. Li, J.-D. Luo, Simvastatin inhibits leptin-induced hypertrophyin cultured neonatal rat cardiomyocytes, Acta Pharmacol. Sin. 27 (2006) 419–422.http://dx.doi.org/10.1111/j.1745-7254.2006.00300.x.
[40] M. Ibrahim, J. Gorelik, M.H. Yacoub, C.M. Terracciano, The structure and function ofcardiac t-tubules in health and disease, Proc. Biol. Sci. 278 (2011) 2714–2723.http://dx.doi.org/10.1098/rspb.2011.0624.
[41] M. Iglewski, J.A. Hill, S. Lavandero, B.A. Rothermel, Mitochondrial fission and au-tophagy in the normal and diseased heart, Curr. Hypertens. Rep. 12 (2010)418–425. http://dx.doi.org/10.1007/s11906-010-0147-x.
[42] E. Jéquier, Leptin signaling, adiposity, and energy balance, Ann. N. Y. Acad. Sci. 967(2002) 379–388. http://dx.doi.org/10.1111/j.1749-6632.2002.tb04293.x.
[43] M.R. Kandadi, N.D. Roe, J. Ren, Autophagy inhibition rescues against leptin-inducedcardiac contractile dysfunction, Curr. Pharm. Des. 20 (2014) 675–683.
[44] M. Karmazyn, D.M. Purdham, V. Rajapurohitam, A. Zeidan, Leptin as a cardiachypertrophic factor: a potential target for therapeutics, Trends Cardiovasc.Med. 17 (2007) 206–211. http://dx.doi.org/10.1016/j.tcm.2007.06.001.
[45] T. Kelesidis, I. Kelesidis, S. Chou, C.S. Mantzoros, Narrative review: the role ofleptin in human physiology: emerging clinical applications, Ann. Intern. Med.152 (2010) 93–100. http://dx.doi.org/10.7326/0003-4819-152-2-201001190-00008.
[46] C. Kho, A. Lee, R.J. Hajjar, Altered sarcoplasmic reticulum calcium cycling—targetsfor heart failure therapy, Nat. Rev. Cardiol. 9 (2012) 717–733. http://dx.doi.org/10.1038/nrcardio.2012.145.
[47] S.C. Kolwicz, S. Purohit, R. Tian, Cardiac metabolism and its interactions with con-traction, growth, and survival of cardiomyocytes, Circ. Res. 113 (2013) 603–616.http://dx.doi.org/10.1161/CIRCRESAHA.113.302095.
[48] S.-D. Lee, B.-S. Tzang, W.-W. Kuo, Y.-M. Lin, A.-L. Yang, S.-H. Chen, F.-J. Tsai, F.-L.Wu, M.-C. Lu, C.-Y. Huang, Cardiac fas receptor-dependent apoptotic pathway inobese zucker rats, Obesity (Silver Spring) 15 (2007) 2407–2415. http://dx.doi.org/10.1038/oby.2007.286.
[49] Y. Lee, H.-Y. Lee, A.B. Gustafsson, Regulation of autophagy by metabolic and stresssignaling pathways in the heart, J. Cardiovasc. Pharmacol. 60 (2012) 118–124.http://dx.doi.org/10.1097/FJC.0b013e318256cdd0.
[50] M. Leifheit-Nestler, N.-M. Wagner, R. Gogiraju, M. Didié, S. Konstantinides, G.Hasenfuss, K. Schäfer, Importance of leptin signaling and signal transducerand activator of transcription-3 activation in mediating the cardiac hypertro-phy associated with obesity, J. Transl. Med. 11 (2013) 170. http://dx.doi.org/10.1186/1479-5876-11-170.
[51] S.-Y. Li, X. Yang, A.F. Ceylan-Isik, M. Du, N. Sreejayan, J. Ren, Cardiac contractile dys-function in Lep/Lep obesity is accompanied by NADPH oxidase activation, oxida-tive modification of sarco(endo)plasmic reticulum Ca2+−ATPase and myosinheavy chain isozyme switch, Diabetologia 49 (2006) 1434–1446. http://dx.doi.org/10.1007/s00125-006-0229-0.
[52] C.-C. Liew, V.J. Dzau, Molecular genetics and genomics of heart failure, Nat. Rev.Genet. 5 (2004) 811–825. http://dx.doi.org/10.1038/nrg1470.
[53] T.-C. Lin, T.-C. Lee, S.-L. Hsu, C.-S. Yang, The molecular mechanism of leptin secre-tion and expression induced by aristolochic acid in kidney fibroblast, PLoS One 6(2011) e16654. http://dx.doi.org/10.1371/journal.pone.0016654.
[54] B. Loos, A.-M. Engelbrecht, R.A. Lockshin, D.J. Klionsky, Z. Zakeri, The variabilityof autophagy and cell death susceptibility: unanswered questions, Autophagy9 (2013) 1270–1285. http://dx.doi.org/10.4161/auto.25560.
[55] G.D. Lopaschuk, J.R. Ussher, C.D.L. Folmes, J.S. Jaswal, W.C. Stanley, Myocardial fattyacid metabolism in health and disease, Physiol. Rev. 90 (2010) 207–258. http://dx.doi.org/10.1152/physrev.00015.2009.
[56] M.-C. Lu, B.-S. Tzang, W.-W. Kuo, F.-L. Wu, Y.-S. Chen, C.-H. Tsai, C.-Y. Huang, S.-D.Lee, More activated cardiac mitochondrial-dependent apoptotic pathway in obesezucker rats, Obesity (Silver Spring) 15 (2007) 2634–2642. http://dx.doi.org/10.1038/oby.2007.315.
[57] T.A. Lutz, S.C. Woods, Overview of animal models of obesity, Curr. Protoc.Pharmacol. (2012)http://dx.doi.org/10.1002/0471141755.ph0561s58 (Chapter 5,Unit5.61).
[58] S. Madani, S. De Girolamo, D.M. Muñoz, R.-K. Li, G. Sweeney, Direct effects of leptinon size and extracellular matrix components of human pediatric ventricularmyocytes, Cardiovasc. Res. 69 (2006) 716–725. http://dx.doi.org/10.1016/j.cardiores.2005.11.022.
[59] S.A. Malik, G. Mariño, A. BenYounès, S. Shen, F. Harper, M.C. Maiuri, G. Kroemer,Neuroendocrine regulation of autophagy by leptin, Cell Cycle 10 (2014)2917–2923. http://dx.doi.org/10.4161/cc.10.17.17067.
[60] E. Martinez-Abundis, V. Rajapurohitam, J.V. Haist, X.T. Gan, M. Karmazyn, Theobesity-related peptide leptin sensitizes cardiac mitochondria to calcium-induced permeability transition pore opening and apoptosis, PLoS One 7 (2012)e41612. http://dx.doi.org/10.1371/journal.pone.0041612.
[61] P.K. Mazumder, B.T. O'Neill, M.W. Roberts, J. Buchanan, U.J. Yun, R.C. Cooksey, S.Boudina, E.D. Abel, Impaired cardiac efficiency and increased fatty acid oxidationin insulin-resistant ob/ob mouse hearts, Diabetes 53 (2004) 2366–2374. http://dx.doi.org/10.2337/diabetes.53.9.2366.
[62] K.R. McGaffin, C.S. Moravec, C.F. McTiernan, Leptin signaling in the failing andmechanically unloaded human heart, Circ. Heart Fail. 2 (2009) 676–683. http://dx.doi.org/10.1161/CIRCHEARTFAILURE.109.869909.
[63] K.R. McGaffin, W.G. Witham, K.A. Yester, L.C. Romano, R.M. O'Doherty, C.F.McTiernan, C.P. O'Donnell, Cardiac-specific leptin receptor deletion exacerbates
of leptin in cardiomyocyte physiology and physiopathology, Life Sci

8 S. Feijóo-Bandín et al. / Life Sciences xxx (2015) xxx–xxx
ischaemic heart failure in mice, Cardiovasc. Res. 89 (2011) 60–71. http://dx.doi.org/10.1093/cvr/cvq288.
[64] C.F. Michiels, P. Fransen, D.G. De Munck, G.R.Y. De Meyer, W. Martinet, Defectiveautophagy in vascular smooth muscle cells alters contractility and calciumhomeostasis in mice, Am. J. Physiol. Heart Circ. Physiol. (2015)http://dx.doi.org/10.1152/ajpheart.00659.2014 (ajpheart.00659.2014).
[65] K.M. Minhas, S.A. Khan, S.V.Y. Raju, A.C. Phan, D.R. Gonzalez, M.W. Skaf, K. Lee,A.D. Tejani, A.P. Saliaris, A.P. Saliaris, et al., Leptin repletion restores depressed{beta}-adrenergic contractility in ob/ob mice independently of cardiac hyper-trophy, J. Physiol. 565 (2005) 463–474. http://dx.doi.org/10.1113/jphysiol.2005.084566.
[66] M.W. Nickola, L.E. Wold, P.B. Colligan, G.J. Wang, W.K. Samson, J. Ren, Leptinattenuates cardiac contraction in rat ventricular myocytes. Role of NO, Hyperten-sion 36 (2000) 501–505. http://dx.doi.org/10.1161/01.HYP.36.4.501.
[67] G. Niu, X. Chen, Apoptosis imaging: beyond annexin V, J. Nucl. Med. 51 (2010)1659–1662. http://dx.doi.org/10.2967/jnumed.110.078584.
[68] A.M. Orogo, Å.B. Gustafsson, Cell death in the myocardium: my heart won't go on,IUBMB Life 65 (2013) 651–656. http://dx.doi.org/10.1002/iub.1180.
[69] R. Palanivel, M. Eguchi, I. Shuralyova, I. Coe, G. Sweeney, Distinct effects of short-and long-term leptin treatment on glucose and fatty acid uptake and metabolismin HL-1 cardiomyocytes, Metabolism 55 (2006) 1067–1075. http://dx.doi.org/10.1016/j.metabol.2006.03.020.
[70] G. Paolisso, M.R. Tagliamonte, M. Galderisi, G.A. Zito, A. Petrocelli, C. Carella, O. deDivitiis, M. Varricchio, Plasma leptin level is associated with myocardial wall thick-ness in hypertensive insulin-resistant men, Hypertension 34 (1999) 1047–1052.http://dx.doi.org/10.1161/01.HYP.34.5.1047.
[71] H.-K. Park, R.S. Ahima, Leptin signaling, F1000Prime Rep. 6 (2014) 73. http://dx.doi.org/10.12703/P6-73.
[72] V. Parra, F. Moraga, J. Kuzmicic, C. López-Crisosto, R. Troncoso, N. Torrealba, A.Criollo, J. Díaz-Elizondo, B.A. Rothermel, A.F.G. Quest, et al., Calcium and mito-chondrial metabolism in ceramide-induced cardiomyocyte death, Biochim.Biophys. Acta 1832 (2013) 1334–1344. http://dx.doi.org/10.1016/j.bbadis.2013.04.009.
[73] M.A. Pelleymounter, M.J. Cullen, M.B. Baker, R. Hecht, D. Winters, T. Boone, F.Collins, Effects of the obese gene product on body weight regulation in ob/obmice, Science 269 (1995) 540–543. http://dx.doi.org/10.1126/science.7624776.
[74] L. Perego, P. Pizzocri, D. Corradi, F. Maisano, M. Paganelli, P. Fiorina, M. Barbieri, A.Morabito, G. Paolisso, F. Folli, et al., Circulating leptin correlates with left ventricu-lar mass in morbid (grade III) obesity before and after weight loss induced by bar-iatric surgery: a potential role for leptin in mediating human left ventricularhypertrophy, J. Clin. Endocrinol. Metab. 90 (2005) 4087–4093. http://dx.doi.org/10.1210/jc.2004-1963.
[75] R.J.M.I. Piñeiro, S. Eiras, J. Viñuela, F. Lago, J.R. González-Juanatey, Leptin does notinduce hypertrophy, cell cycle alterations, or production of MCP-1 in cultured ratand mouse cardiomyocytes, Endocr. Res. 31 (2005) 375–386. http://dx.doi.org/10.1080/07435800500456937.
[76] X. Pi-Sunyer, Themedical risks of obesity, Postgrad. Med. 121 (2009) 21–33. http://dx.doi.org/10.3810/pgm.2009.11.2074.
[77] D.M. Purdham, V. Rajapurohitam, A. Zeidan, C. Huang, G.J. Gross, M. Karmazyn,A neutralizing leptin receptor antibody mitigates hypertrophy and hemody-namic dysfunction in the postinfarcted rat heart, Am. J. Physiol. Heart Circ.Physiol. 295 (2008) H441–H446. http://dx.doi.org/10.1152/ajpheart.91537.2007.
[78] M.J. Radin, B.J. Holycross, S.A. McCune, R.A. Altschuld, Crosstalk between leptin andinterleukin-1β abrogates negative inotropic effects in a model of chronichyperleptinemia, Exp. Biol. Med. (Maywood). 236 (2011) 1263–1273. http://dx.doi.org/10.1258/ebm.2011.011144.
[79] V. Rajapurohitam, X.T. Gan, L.A. Kirshenbaum, M. Karmazyn, The obesity-associated peptide leptin induces hypertrophy in neonatal rat ventricularmyocytes, Circ. Res. 93 (2003) 277–279. http://dx.doi.org/10.1161/01.RES.0000089255.37804.72.
[80] V. Rajapurohitam, F. Izaddoustdar, E. Martinez-Abundis, M. Karmazyn, Leptin-induced cardiomyocyte hypertrophy reveals both calcium-dependent andcalcium-independent/RhoA-dependent calcineurin activation and NFAT nucleartranslocation, Cell. Signal. 24 (2012) 2283–2290. http://dx.doi.org/10.1016/j.cellsig.2012.07.025.
[81] V. Rajapurohitam, S. Javadov, D.M. Purdham, L.A. Kirshenbaum, M. Karmazyn, Anautocrine role for leptin in mediating the cardiomyocyte hypertrophic effects ofangiotensin II and endothelin-1, J. Mol. Cell. Cardiol. 41 (2006) 265–274. http://dx.doi.org/10.1016/j.yjmcc.2006.05.001.
[82] S.V.Y. Raju, M. Zheng, K.H. Schuleri, A.C. Phan, D. Bedja, R.M. Saraiva, O.Yiginer, K. Vandegaer, K.L. Gabrielson, C.P. O'donnell, et al., Activation ofthe cardiac ciliary neurotrophic factor receptor reverses left ventricularhypertrophy in leptin-deficient and leptin-resistant obesity, Proc. Natl.Acad. Sci. U. S. A. 103 (2006) 4222–4227. http://dx.doi.org/10.1073/pnas.0510460103.
[83] A.S. Reed, E.K. Unger, L.E. Olofsson, M.L. Piper, M.G. Myers, A.W. Xu, Functional roleof suppressor of cytokine signaling 3 upregulation in hypothalamic leptin resis-tance and long-term energy homeostasis, Diabetes 59 (2010) 894–906. http://dx.doi.org/10.2337/db09-1024.
[84] J. Ren, Leptin and hyperleptinemia— from friend to foe for cardiovascular function,J. Endocrinol. 181 (2004) 1–10. http://dx.doi.org/10.1677/joe.0.1810001.
[85] J. Ren, F. Dong, G.-J. Cai, P. Zhao, J.M. Nunn, L.E.Wold, J. Pei, Interaction between ageand obesity on cardiomyocyte contractile function: role of leptin and stress signal-ing, PLoS One 5 (2010) e10085. http://dx.doi.org/10.1371/journal.pone.0010085.
Please cite this article as: S. Feijóo-Bandín, et al., 20 years of leptin: Role(2015), http://dx.doi.org/10.1016/j.lfs.2015.02.016
[86] J. Ren, J.R. Sowers, M.F. Walsh, R.A. Brown, Reduced contractile response to insulinand IGF-I in ventricular myocytes from genetically obese zucker rats, Am. J. Physiol.Heart Circ. Physiol. 279 (2000) H1708–H1714.
[87] M. Rosenbaum, R.L. Leibel, 20 years of leptin: role of leptin in energy homeostasis inhumans, J. Endocrinol. 223 (2014) T83–T96. http://dx.doi.org/10.1530/JOE-14-0358.
[88] J.K. Salabei, D.J. Conklin, Cardiovascular autophagy: crossroads of pathology, phar-macology and toxicology, Cardiovasc. Toxicol. 13 (2013) 220–229. http://dx.doi.org/10.1007/s12012-013-9200-8.
[89] D. Segula, Complications of obesity in adults: a short review of the literature,Malawi Med. J. 26 (2014) 20–24.
[90] R.M. Shaw, H.M. Colecraft, L-type calcium channel targeting and local signalling incardiac myocytes, Cardiovasc. Res. 98 (2013) 177–186. http://dx.doi.org/10.1093/cvr/cvt021.
[91] E.-J. Shin, K. Schram, X.-L. Zheng, G. Sweeney, Leptin attenuates hypoxia/reoxygenation-induced activation of the intrinsic pathway of apoptosis in ratH9c2 cells, J. Cell. Physiol. 221 (2009) 490–497. http://dx.doi.org/10.1002/jcp.21883.
[92] K.I. Shoghi, R.J. Gropler, T. Sharp, P. Herrero, N. Fettig, Y. Su, M.S. Mitra, A. Kovacs,B.N. Finck, M.J. Welch, Time course of alterations in myocardial glucose utilizationin the zucker diabetic fatty rat with correlation to gene expression of glucose trans-porters: a small-animal PET investigation, J. Nucl. Med. 49 (2008) 1320–1327.http://dx.doi.org/10.2967/jnumed.108.051672.
[93] M. Słupecka, J. Woliński, M. Gajewska, S.G. Pierzynowski, Enteral leptin admin-istration affects intestinal autophagy in suckling piglets, Domest. Anim.Endocrinol. 46 (2014) 12–19. http://dx.doi.org/10.1016/j.domaniend.2013.09.007.
[94] C.C.T. Smith, R.A. Dixon, A.M. Wynne, L. Theodorou, S.-G. Ong, S. Subrayan, S.M.Davidson, D.J. Hausenloy, D.M. Yellon, Leptin-induced cardioprotection involvesJAK/STAT signaling that may be linked to the mitochondrial permeability transi-tion pore, Am. J. Physiol. Heart Circ. Physiol. 299 (2010) H1265–H1270. http://dx.doi.org/10.1152/ajpheart.00092.2010.
[95] C.C.T. Smith, M.M. Mocanu, S.M. Davidson, A.M. Wynne, J.C. Simpkin, D.M. Yellon,Leptin, the obesity-associated hormone, exhibits direct cardioprotective effects,Br. J. Pharmacol. 149 (2006) 5–13. http://dx.doi.org/10.1038/sj.bjp.0706834.
[96] A. Terman, T. Kurz, M. Navratil, E.A. Arriaga, U.T. Brunk, Mitochondrial turnoverand aging of long-lived postmitotic cells: the mitochondrial-lysosomal axis theoryof aging, Antioxid. Redox Signal. 12 (2010) 503–535. http://dx.doi.org/10.1089/ars.2009.2598.
[97] P.S. Trivedi, L.A. Barouch, Cardiomyocyte apoptosis in animal models of obesity,Curr. Hypertens. Rep. 10 (2008) 454–460. http://dx.doi.org/10.1007/s11906-008-0085-z.
[98] M. Trovati, G. Doronzo, C. Barale, C. Vaccheris, I. Russo, F. Cavalot, Leptin and vascu-lar smooth muscle cells, Curr. Pharm. Des. 20 (2014) 625–634.
[99] I. Uphues, T. Kolter, B. Goud, J. Eckel, Failure of insulin-regulated recruitment of theglucose transporter GLUT4 in cardiac muscle of obese zucker rats is associatedwithalterations of small-molecular-mass GTP-binding proteins, Biochem. J. 311 (Pt 1)(1995) 161–166.
[100] M.N. Vansaun, A.M. Mendonsa, D. Lee Gorden, Hepatocellular proliferation corre-lates with inflammatory cell and cytokine changes in a murine model ofnonalchoholic fatty liver disease, PLoS One 8 (2013) e73054. http://dx.doi.org/10.1371/journal.pone.0073054.
[101] H. Wang, W. Luo, D.T. Eitzman, Leptin in thrombosis and atherosclerosis, Curr.Pharm. Des. 20 (2014) 641–645.
[102] J.Wang, R. Liu, M. Hawkins, N. Barzilai, L. Rossetti, A nutrient-sensing pathway reg-ulates leptin gene expression in muscle and fat, Nature 393 (1998) 684–688.http://dx.doi.org/10.1038/31474.
[103] L. Wang, X. Hu, W. Zhu, Z. Jiang, Y. Zhou, P. Chen, J. Wang, Increased leptin byhypoxic-preconditioning promotes autophagy of mesenchymal stem cells andprotects them from apoptosis, Sci. China Life Sci. 57 (2014) 171–180. http://dx.doi.org/10.1007/s11427-014-4607-4.
[104] A.E. Wentz, D.A. D'Avignon, M.L. Weber, D.G. Cotter, J.M. Doherty, R. Kerns, R.Nagarajan, N. Reddy, N. Sambandam, P.A. Crawford, Adaptation of myocardial sub-strate metabolism to a ketogenic nutrient environment, J. Biol. Chem. 285 (2010)24447–24456. http://dx.doi.org/10.1074/jbc.M110.100651.
[105] L.E. Wold, D.P. Relling, J. Duan, F.L. Norby, J. Ren, Abrogated leptin-induced cardiaccontractile response in ventricular myocytes under spontaneous hypertension:role of Jak/STAT pathway, Hypertension 39 (2002) 69–74.
[106] F.-P. Xu, M.-S. Chen, Y.-Z. Wang, Q. Yi, S.-B. Lin, A.F. Chen, J.-D. Luo, Leptin induceshypertrophy via endothelin-1-reactive oxygen species pathway in cultured neona-tal rat cardiomyocytes, Circulation 110 (2004) 1269–1275. http://dx.doi.org/10.1161/01.CIR.0000140766.52771.6D.
[107] L. Yu, Y. Zhao, S. Xu, C. Jin, M.Wang, G. Fu, Leptin confers protection against TNF-α-induced apoptosis in rat cardiomyocytes, Biochem. Biophys. Res. Commun. 455(2014) 126–132. http://dx.doi.org/10.1016/j.bbrc.2014.10.134.
[108] A. Zeidan, J.C. Hunter, S. Javadov, M. Karmazyn, mTOR mediates RhoA-dependentleptin-induced cardiomyocyte hypertrophy, Mol. Cell. Biochem. 352 (2011)99–108. http://dx.doi.org/10.1007/s11010-011-0744-2.
[109] A. Zeidan, S. Javadov, S. Chakrabarti, M. Karmazyn, Leptin-induced cardiomyocytehypertrophy involves selective caveolae and RhoA/ROCK-dependent p38 MAPKtranslocation to nuclei, Cardiovasc. Res. 77 (2008) 64–72. http://dx.doi.org/10.1093/cvr/cvm020.
[110] A. Zeidan, S. Javadov, M. Karmazyn, Essential role of Rho/ROCK-dependent pro-cesses and actin dynamics in mediating leptin-induced hypertrophy in rat neona-tal ventricular myocytes, Cardiovasc. Res. 72 (2006) 101–111. http://dx.doi.org/10.1016/j.cardiores.2006.06.024.
of leptin in cardiomyocyte physiology and physiopathology, Life Sci

9S. Feijóo-Bandín et al. / Life Sciences xxx (2015) xxx–xxx
[111] Y. Zhang, R. Proenca, M. Maffei, M. Barone, L. Leopold, J.M. Friedman, Positionalcloning of the mouse obese gene and its human homologue, Nature 372 (1994)425–432. http://dx.doi.org/10.1038/372425a0.
[112] J. Zheng, J. Fang, Y.-J. Yin, X.-C. Wang, A.-J. Ren, J. Bai, X.-J. Sun, W.-J. Yuan, L. Lin,Leptin protects cardiomyocytes from serum-deprivation-induced apoptosis by
Please cite this article as: S. Feijóo-Bandín, et al., 20 years of leptin: Role(2015), http://dx.doi.org/10.1016/j.lfs.2015.02.016
increasing anti-oxidant defence, Clin. Exp. Pharmacol. Physiol. 37 (2010)955–962. http://dx.doi.org/10.1111/j.1440-1681.2010.05415.x.
[113] Y.T. Zhou, P. Grayburn, A. Karim, M. Shimabukuro, M. Higa, D. Baetens, L. Orci, R.H.Unger, Lipotoxic heart disease in obese rats: implications for human obesity, Proc.Natl. Acad. Sci. U. S. A. 97 (2000) 1784–1789.
of leptin in cardiomyocyte physiology and physiopathology, Life Sci