2011 predicciones genéticas - ipafcv.files.wordpress.com · 3 podría ser el promedio de la...
TRANSCRIPT

Introducción a la Producción Animal - FCV - UNNE
2011
Predicciones Genéticas
La cantidad de mejoramiento o tasa de cambio genético en una población depende en gran
medida de la precisión de la predicción o más exactamente, en la precisión de los valores de
cría estimados.
La precisión puede ser incrementada hasta un cierto punto al aumentar la heredabilidad de los
caracteres. Manejar los animales de manera uniforme, tomando minuciosas medidas,
ajustando por efectos ambientales conocidos, utilizando grupos contemporáneos – todo
ayuda. Sin embargo, para aumentar la precisión aún más se requiere el uso de tanta
información como sea posible y ponderar cada una apropiadamente. Se requiere el uso de la
tecnología de predicción genética.
Existen dos metodologías estrechamente relacionadas que se utilizan comúnmente para la
predicción genética: el índice de selección y la mejor predicción linear insesgada (BLUP). No
se examinarán estas tecnologías en detalle, pues esto requeriría conocimientos de estadística
y álgebra matricial. El énfasis aquí no es como los índices de selección y la mejor predicción
linear insesgada trabajan, sino más bien cuando deberían ser usadas y en qué y para que
podemos utilizarla. Se tratará también la presentación e interpretación de las predicciones
genéticas producidas con estas tecnologías.
COMPARACIÓN DE ANIMALES UTILIZANDO DATOS DE GRUPOS GENÉTICAMENTE
SIMILARES: EL INDICE DE SELECCIÓN.
La teoría del índice de selección fue desarrollada por primera vez en 1930 y 1940, como
método para la predicción genética y como un medio para combinar caracteres en la selección
de animales de la manera más óptima económicamente. Aquí se considerará el índice de
selección como una metodología de predicción. Su rol en la selección multicarácter ya fue
discutido en métodos de selección.
Un índice de selección es esencialmente una combinación linear de varios tipos de
información fenotípica y factores de ponderación apropiados. Toman la siguiente
forma:
Donde I = un valor índice o predicción genética
bi= un factor de ponderación.
xi= un simple ítem de información fenotípica – un registro de producción o el promedio de los
registros de producción de un grupo.
n= número total de ítems de información fenotípica. En cualquier método de predicción genética la información utilizada en el cálculo de las
predicciones genéticas para un individuo provienen de tres clases de fuentes:
(1) registros de performance del propio individuo,

Introducción a la Producción Animal - FCV - UNNE
2011
(2) registros de producción de antecesores y/o parientes colaterales de los individuos (datos
de pedigrí o genealógicos), y
(3) los registros de producción de los descendientes del individuo (datos de la progenie).
Las cantidades relativas de datos de estas tres fuentes varían. Animales no nacidos o muy
jóvenes tienen solamente datos de pedigrí. Como estos animales crecerán y envejecerán ellos
adquirirán datos de performance propios. Asumiendo, que ellos pueden adquirir datos
propios, ya que algunos caracteres tienen limitación de sexo. Los machos lecheros por ejemplo
no tienen datos de performance propios para caracteres de lactación y leche Si los animales
son seleccionados para convertirse en padres, ellos generarán datos de progenie, y si se
vuelven populares, tendrán una gran cantidad de datos de progenie.
Cualquiera de los tres tipos de registros se puede tomar de cualquier carácter para el cual se
calcularán las predicciones, o pueden ser datos de otros caracteres genéticamente
relacionados. Por ejemplo, en las especies en las cuales la distocia (dificultad al parto) es una
preocupación, las predicciones de la susceptibilidad genética hacia ellas pueden ser
determinadas por la medida directa de los grados de distocia, por el peso al nacimiento (un
carácter genéticamente correlacionado), o por ambos.
Los datos utilizados en el índice de selección – x1 etc – provienen de muchas fuentes dentro de
las categorías: de la propia performance, del pedigrí, y los datos de progenie. Por ejemplo, x1
podría ser un registro de performance individual propia para un carácter, x2 podría ser el
promedio de performance de un medio hermano paterno del individuo para el mismo
carácter, x3 podría ser el promedio de la performance de la progenie del individuo, y x4, x5 y x6
podrían representar la performance del individuo, medio hermano, y progenie
respectivamente para un carácter correlacionado. En todos los casos cada x es un número.
En un índice de selección cada ítem de la información fenotípica es normalmente expresado
como el desvío de la media de un grupo contemporáneo.
Grupos contemporáneos
Ajustando por efectos ambientales conocidos y manejando los animales para minimizar las
diferencias en la performance animal causadas por otros efectos ambientales se aumenta la
precisición de las predicciones ¿Pero qué se puede hacer en situaciones donde no todos los
animales pueden ser manejados de la misma forma? Por ejemplo ¿Qué debería hacer un
criador de animales en sistemas extensivos de pastoreo cuando algunos de los animales en un
rebaño o lote tienen acceso a una pastura de alta calidad, y otros deben ser mantenidos en
una pastura de poca calidad? Y ¿Qué se puede hacer para hacer posible que la performance de
los animales de un criador sea comparada justamente con la performance de los animales de
otro criador cuando el manejo y el ambiente físico difieren en sus establecimientos?
Una respuesta es expresar la performance del animal no en términos absolutos, sino
como una desviación de la media de un grupo contemporáneo.
Un grupo contemporáneo, es un grupo de animales que han experimentado un
ambiente o efectos no genéticos similar con respecto a la expresión de un carácter.
Introducción a la Producción Animal - FCV - UNNE
2011
Un “ambiente similar” en este contexto significa que los animales en el grupo
(contemporáneos) se desempeñan en la mismo lugar físico, son del mismo sexo, de edad
similar y han tenido un mismo manejo.
Por ejemplo un grupo contemporáneo para peso al destete en corderos o terneros
comúnmente serían definidos por: el rebaño o lote, el año, la estación del año, sexo, y efectos
de manejo como la pastura o el régimen de alimentación.
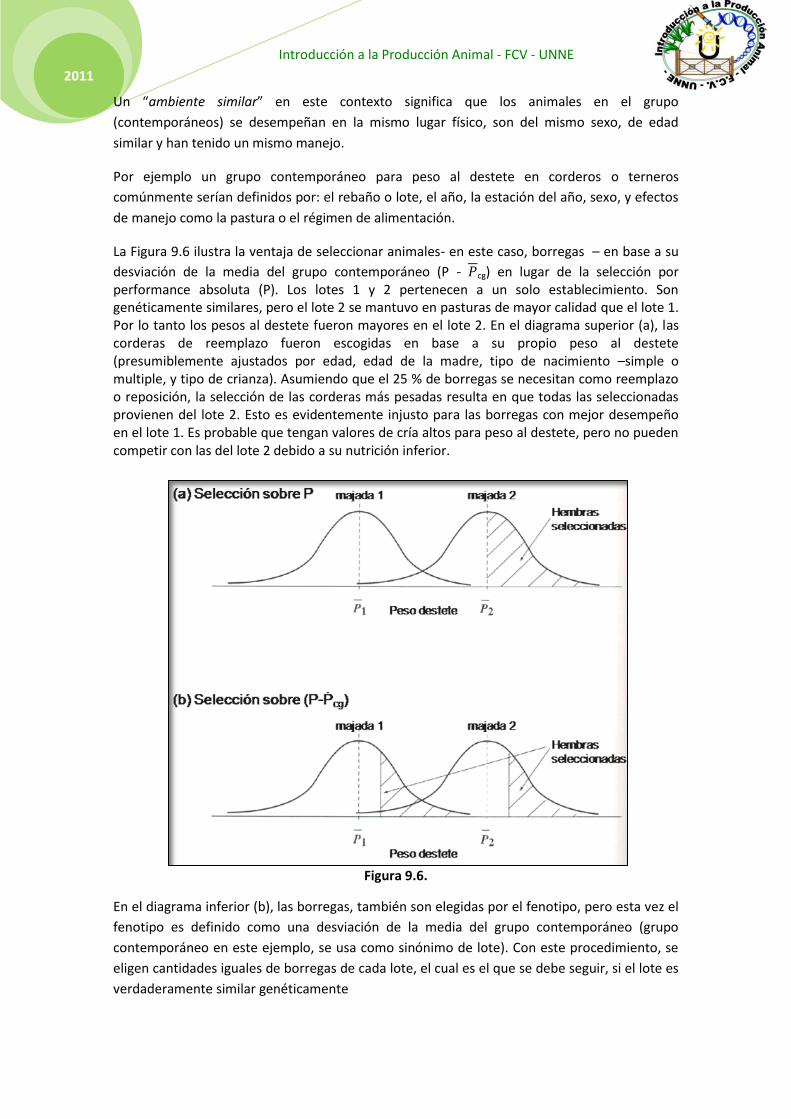
La Figura 9.6 ilustra la ventaja de seleccionar animales- en este caso, borregas – en base a su
desviación de la media del grupo contemporáneo (P - cg) en lugar de la selección por performance absoluta (P). Los lotes 1 y 2 pertenecen a un solo establecimiento. Son genéticamente similares, pero el lote 2 se mantuvo en pasturas de mayor calidad que el lote 1. Por lo tanto los pesos al destete fueron mayores en el lote 2. En el diagrama superior (a), las corderas de reemplazo fueron escogidas en base a su propio peso al destete (presumiblemente ajustados por edad, edad de la madre, tipo de nacimiento –simple o multiple, y tipo de crianza). Asumiendo que el 25 % de borregas se necesitan como reemplazo o reposición, la selección de las corderas más pesadas resulta en que todas las seleccionadas provienen del lote 2. Esto es evidentemente injusto para las borregas con mejor desempeño en el lote 1. Es probable que tengan valores de cría altos para peso al destete, pero no pueden competir con las del lote 2 debido a su nutrición inferior.
Figura 9.6.
En el diagrama inferior (b), las borregas, también son elegidas por el fenotipo, pero esta vez el
fenotipo es definido como una desviación de la media del grupo contemporáneo (grupo
contemporáneo en este ejemplo, se usa como sinónimo de lote). Con este procedimiento, se
eligen cantidades iguales de borregas de cada lote, el cual es el que se debe seguir, si el lote es
verdaderamente similar genéticamente

Introducción a la Producción Animal - FCV - UNNE
2011
Cuando existen diferencias ambientales entre grupos de animales, la relación entre los valores
de cría de los animales y la performance absoluta se debilita. Algunos animales con valores de
cría pobres se desempeñan relativamente bien debido a un ambiente de grupo favorable, y
algunos animales con valores de cría superiores se desempeñan pobremente a causa de un
ambiente de grupo desfavorable
Pero cuando el fenotipo se expresa como una desviación de la media de un grupo
contemporáneo, las diferencias ambientales entre grupos se justifican, y las relaciones entre
los valores de cría de los animales y esta nueva medida de performance es mucho más fuerte.
Expresando la performance de esa forma, se tiene en cuenta las diferencias ambientales entre
grupos contemporáneos. EL problema con este enfoque, es que se asume que todos los grupos
contemporáneos son genéticamente similares. Un desvío de +10 unidades en un grupo
contemporáneo se asume como el equivalente genético de una desviación de +10 unidades en
cualquier otro grupo contemporáneo.
Si la media del valor de cría en cada grupo es la misma, (es decir si los grupos contemporáneos
son realmente genéticamente similares) esta asunción es correcta. Sin embargo, si el valor de
cría medio de los grupos contemporáneos difiere, el uso de las desviaciones de las medias del
grupo contemporáneo crea sesgos en los datos.
Los registros provenientes de los grupos contemporáneos genéticamente más pobres parecen
mejores de lo que deberían, y los registros de los grupos contemporáneos genéticamente
superiores parecen peores de lo que deberían.
Por esta razón, los índices de selección solo deberían ser usados para las predicciones
genéticas cuando los datos de performance provienen de un grupo contemporáneo que se
piensa son genéticamente similares. En la práctica, esto significa que probablemente deberían
ser utilizados dentro de rebaños, rodeos o majadas individuales y no en toda una población. Y
si un determinado rebaño ha experimentado cambios genéticos significativos en el tiempo, los
datos provenientes de los grupos contemporáneos más antiguos – grupos con valores de cría
más “antiguos“y por lo tanto diferentes – deberían ser excluidos del índice.
Los índices de selección podrán ser de aplicación restringida, pero son muy útiles. Suponga,
por ejemplo, que usted está criando ovejas y quiere comparar sus carneros sobre la base de la
performance de sus progenies dentro de su majada. Usted podría usar un índice de selección
para producir progenie, basado en EBVs o EPDs para cada carnero. Los cálculos involucrados
son bastante simples, no requieren nada más que una calculadora de mano. Usted necesitará
una computadora para aplicaciones más complejas de los índices de selección pero aún así una
PC y una hoja de cálculo serán suficientes.
Índice de selección: Una combinación linear de información fenotípica y factores de
ponderación que se utiliza para la predicción genética cuando los datos de performance
provienen de grupos contemporáneos genéticamente similares.
Datos de performance del propio individuo: Información sobre el fenotipo del propio
individuo

Introducción a la Producción Animal - FCV - UNNE
2011
Datos de pedigrí: Información sobre el genotipo o la performance de antecesores y/o
parientes colaterales de un individuo
Datos de la progenie: Información sobre el genotipo o la performance de los descendientes de
un individuo.
Sesgo: Cualquier factor que causa la distorsión de las predicciones genéticas.
COMPARACIÓN DE ANIMALES USANDO DATOS DE DIFERNTES GRUPOS GENÉTICOS:
MEJOR PREDICCIÓN LINEAR INSESGADA. (BLUP)
El índice de selección es un poderoso método para la predicción genética. Sin embargo una
asunción básica del índice de selección es que la información de performance utilizada
proviene de grupos contemporáneos genéticamente similares. ¿Y si queremos hacer
predicciones usando datos de grupos contemporáneos genéticamente diferentes? – grupos de
diferentes establecimientos o de diferentes décadas.
Una extensión de la metodología de los índices de selección conocida como mejor predicción
linear insesgada o BLUP por su sigla en inglés (Best Linear Unbiased Predicition) está
diseñada para este tipo de información. La distinción hecha aquí entre índice de selección y
BLUP refleja diferencias en como las dos tecnologías han sido aplicadas históricamente.
La teoría estadística detrás del BLUP es casi tan vieja como la teoría detrás del índice de
selección. Sin embargo no fue hasta principios de 1980 que las computadoras y los algoritmos
matemáticos han avanzado hasta el punto de que la difusión de la aplicación del BLUP para las
evaluaciones genéticas se convirtió en factible.
BLUP requiere de computación intensivamente. Tal como los índices de selección que utilizan
múltiples fuentes de información, BLUP involucra la solución simultánea de un gran número de
ecuaciones y provee predicciones para una población entera de animales – no solo para un
animal a la vez (que es el caso de la mayoría de las aplicaciones del índice de selección).
Debido a su habilidad para contabilizar o tomar en cuenta las diferencias entre grupos
contemporáneos y debido a que provee predicciones genéticas para muchos animales a la vez
BLUP es el método preferido para las evaluaciones genéticas a gran escala: la evaluación
genética de poblaciones grandes, típicamente razas enteras. Los registros de performance
utilizados en dichas evaluaciones usualmente provienen de datos de campo, datos que
regularmente son reportados por criadores individuales a las asociaciones de cría de las razas o
a las agencias gubernamentales.
Mejor predicción linear insesgada BLUP: Un método de predicción genética que es
particularmente apropiado cuando los datos de performance provienen de grupos
contemporáneos genéticamente diversos.
Evaluaciones genéticas a gran escala: la evaluación genética de poblaciones grandes,
típicamente razas enteras.
Datos de campo: datos que regularmente son reportados por criadores individuales a las
asociaciones de cría de las razas o a las agencias gubernamentales.

Introducción a la Producción Animal - FCV - UNNE
2011
Tipos de modelos de BLUP
BLUP es una técnica que puede ser concebida como una familia de modelos estadísticos, es
decir, representaciones matemáticas de performance animal que incluye varios efectos
ambientales y genéticos y que son usados para las predicciones genéticas.
Hay modelos padre, modelos padre-abuelo materno, modelo animal, modelo de medidas
repetidas, modelos materno-directo, modelos multicarácter- la lista continua y continua.
Las diferencias entre modelos tienen que ver primariamente con que animales recibirán las
predicciones genéticas (solamente los padres, todos los progenitores, o todos los animales), el
número y clase de predicciones generadas y la dificultad computacional. En general cuanto más
sofisticado el modelo, involucra más ecuaciones y más cantidad de recursos informáticos
requiere.
Modelo estadístico: Representación matemática de la performance de un animal que incluye
varios efectos genéticos y ambientales y es utilizado para la predicción genética.
Potencial de los modelos BLUP avanzados
A continuación se presentan los atributos del BLUP aunque no se debería generalizar acerca
de los modelos BLUP. Cada uno difiere del otro de una forma u otra, y dos de ellos no tienen
exactamente las mismas capacidades. El modelo particular aquí es el más básico, el más
avanzado y actualmente el tipo más popular de modelo BLUP – el modelo animal.
El modelo animal tiene una cantidad de cualidades deseables de las cuales la más obvia es su
habilidad para evaluar “todos los animales” (en contraposición a solo los machos) en una
población.
Modelo Animal: Modelo estadístico avanzado para la predicción genética que es utilizado para
evaluar todos los animales (en contraposición a solo los padres) en una población.
Niveles genéticos de los grupos contemporáneos
Los modelos BLUP tienen en cuenta las diferencias en los valores de cría promedio de los
grupos contemporáneos. En otras palabras, tienen en cuenta el hecho de que la performance
superior en un grupo contemporáneo genéticamente inferior no es equivalente a la
performance superior en un grupo contemporáneo superior.
En un análisis BLUP la performance de un caballo ganador en su tercera carrera en una
competencia común no es considerada tan impresionante como la performance del ganador
del Derbi. El índice de selección no puede hacer esta distinción. A causa de que usa desvíos de
la media de los grupos contemporáneos (en el ejemplo del caballo de carrera, desviaciones del
tiempo medio para correr una carrera dada) el índice de selección no tiene en cuenta el nivel
de la competencia. Una desviación de - 0,5 es una desviación de -5 segundos si la carrera es en
el Derbi una de las tantas ferias comunes.
Los modelos BLUP no utilizan desviaciones de las medias de los grupos contemporáneos. En
cambio incluyen ecuaciones que realmente resuelven los efectos del grupo contemporáneo

Introducción a la Producción Animal - FCV - UNNE
2011
(Ecg), es decir efectos ambientales comunes a todos los miembros de un grupo
contemporáneo. Y lo hacen mediante la comparación de la performance de parientes en
grupos contemporáneos diferentes.
Si los animales en un grupo contemporáneo en particular no producen como deberían de
acuerdo a lo determinado por la performance de sus parientes en otros grupos
contemporáneos, entonces, el efecto ambiental común a ese grupo contemporáneo se estima
que es más bajo que la media. Igualmente, si los animales en el grupo producen mejor que lo
que deberían, el Ecg se estima que es superior a la media.
A través de la solución simultánea de ecuaciones, las estimaciones de los efectos de los grupos
contemporáneos proveen la información que se utiliza para producir una predicción genética
más confiable para animales individuales y viceversa.
Así los procedimientos BLUP tienen en cuenta el hecho de que la información de performance
utilizada en las predicciones proviene de grupos contemporáneos que difieren por razones
ambientales y genéticas.
Se puede comparar directamente las predicciones genéticas provenientes de los modelos
BLUP, aún cuando esas predicciones derivan mayormente de registros producidos en
ambientes muy diferentes y/o en grupos contemporáneos que difieren ampliamente en el
mérito genético promedio.
Tendencia genética
Un corolario de las capacidades de los modelos BLUP para contabilizar los niveles genéticos de
los grupos contemporáneos es su habilidad para tomar en cuenta la tendencia genética.
Si una población han sido objeto de una selección efectiva por un período de tiempo
considerable (es decir si ha experimentado una tendencia genética significativa), el valor de
cría promedio de los grupos contemporáneos más nuevos deberían ser mejores que los
valores de cría promedio de los grupos contemporáneos más viejos. Pero a causa de que los
procedimientos BLUP toman en cuenta las diferencias genéticas entre grupos
contemporáneos, las tendencias genéticas usualmente no son un problema (las tendencias
genéticas causarían sesgos en un índice de selección utilizando desvíos de las medias de los
grupos contemporáneos).
Con los modelos BLUP podemos utilizar registros de performance provenientes de animales de
diferentes épocas – desde aquellos que desde hace mucho tiempo que están muertos hasta
los recién nacidos
Uso de todos los datos
Las aplicaciones tradicionales de los índices de selección utilizan registros de relativamente
pocas fuentes. Los modelos BLUP pueden usar información de muchas más fuentes. De hecho,
a menudo utilizan información proveniente de todos los animales en una población.
El beneficio de esto puede no ser grande; los registros de los parientes lejanos potencialmente
no contribuyen mucho a una predicción. Aún así la habilidad de los BLUP de incorporar

Introducción a la Producción Animal - FCV - UNNE
2011
información de toda clase de parientes suma precisión a la predicción, aunque solamente
fuera poco.
También clarifica las relaciones genealógicas. Lo que podría ser considerado el registro de un
medio hermano en un índice de selección típico podría ser representado más correctamente
en un análisis BLUP como el registro de un pariente que es medio hermano y primo hermano
(vea la figura 11.13 y la leyenda que la acompaña). Este registro por lo tanto podría recibir una
ponderación más apropiada con BLUP que con los índices de selección.
El uso de todos los datos también puede ser interpretado como medio para utilizar los datos
de caracteres correlacionados.
Los modelos BLUP multicarácter, modelos estadísticos utilizados para predecir valores para
más de un carácter a la vez, permite que la información sobre un carácter sea utilizada en la
predicción de valores para otro carácter. Esto es especialmente útil cuando la información en
el segundo carácter es escasa. Por ejemplo, generalmente hay mucha más información sobre
pesos al destete que sobre pesos al nacimiento tomados a campo en el ganado de carne.
Cuando estos caracteres son incorporados en un modelo multicarácter, la información del
peso al destete ayuda a predecir valores del peso al nacimiento y viceversa. El resultado es
más preciso para ambos caracteres.
Modelos BLUP multicarácter: modelos estadísticos utilizados para predecir valores para más
de un carácter a la vez.

Introducción a la Producción Animal - FCV - UNNE
2011
Apareamientos no aleatorios
El problema con los apareamientos no al azar desde el punto de vista de la predicción genética
es que le da ventaja a algunos animales sobre los otros – incrementa el sesgo.
Aquellos machos que se piensan que son mejores son apareados con las mejores hembras, y
por lo tanto es factible que obtengan una progenie con las mejores performances. Los
restantes machos son relegados a las peores hembras y por lo tanto es probable que tengan
una progenie con una performance más pobre. Los procedimientos BLUP tienen en cuenta los
apareamientos no aleatorios de ésta clase, en esencia ajustando las predicciones por el mérito
genético de los progenitores (padre y madre).Las predicciones genéticas para los machos y
hembras se producen simultáneamente y la solución simultánea de ecuaciones provoca que
cada predicción sea tenida en cuenta para todas las otras. El resultado es que no es posible
hacer lucir a un individuo mejor de lo que es asignándole un macho o una hembra superior.
Apareamiento no aleatorio: Cualquier sistema de apareamiento en el cual los machos no son
asignados de manera aleatoria a las hembras.
Refugo por baja performance
Otra clase de sesgo es causado por el refugo por baja performance en caracteres repetidos o
en un carácter que es registrado antes del registro de otro carácter genéticamente
relacionado. Un ejemplo de este tipo de sesgo, es el que afecta a los caballos de carrera como
se ilustra la figura 11.4. Los equinos pura sangre de carrera usualmente comienzan a correr a
los dos años de edad. Si tienen éxito en su primera temporada, continúan corriendo. De otra
manera son refugados por baja performance. Refugar de esa manera causa sesgo si la
progenie de alguno de los macho es rechazada más severamente que la progenie de otros
machos.
En la figura 11.4 la performance de la progenie del macho B es mucho mejor que la progenie
del macho A, a los dos años de edad. Claramente el macho B debería tener el mejor valor de
cría para performance en la carrera. Entre las temporadas de carrera, algunos de ambos
machos son refugados por baja performance, pero a causa de que la progenie del macho A
tiene una performance más baja de lo normal, una proporción mucho más grande de su
progenie que de la progenie del macho B es refugada. La poca progenie remanente del macho
A corre bien en años posteriores, entonces la diferencia en la media de la performance
subsecuente de la progenie de los dos machos ( ) es relativamente pequeña. Note que
esta diferencia es mucho más pequeña que si no hubiera habido refugo. Si muchos de los
datos utilizados para predecir los valores de cría para estos machos proviene de la progenie
que mayor de tres años o más, el macho A tendrá una ventaja relativa y el macho B una
desventaja relativa.

Introducción a la Producción Animal - FCV - UNNE
2011
Los modelos BLUP multicarácter tienen en cuenta este sesgo causado por el refugo por baja
performance. Si el carácter en el cual se basa el refugo es incluido en el análisis y si las
relaciones genéticas (correlaciones genéticas) entre ese carácter y caracteres subsecuentes se
incorporan también, las predicciones para el segundo carácter no son afectadas por el refugo.
Si el BLUP es utilizado para evaluar caballos de carrera, las predicciones de los valores de cría
para la performance de una carrera a edades mayores no sufrirán sesgo por el refugo por
performance a los dos años de edad.
Predicción de los componentes genéticos directos y maternos de los caracteres
Todos los caracteres tienen lo que es denominado un componente directo, el efecto de los
genes de un individuo sobre su propia performance. Algunos caracteres tienen un
componente materno, el efecto de los genes en la madre de un individuo que influencia en la
performance de este a través del ambiente provisto por la madre.
El ejemplo clásico de un carácter con ambos componentes directo y materno es el peso al
destete. El peso al destete de un animal es una función de su habilidad heredada para la tasa
de crecimiento y la producción de leche y habilidad materna de su madre. La tasa de
crecimiento heredada está determinada por los genes del animal. Este comprende el
componente directo del peso al destete.
La producción de leche y la habilidad materna de la madre están determinadas por los genes
de ésta (así como por el ambiente). Los genes de la madre para estos caracteres no afectan la

Introducción a la Producción Animal - FCV - UNNE
2011
tasa de crecimiento de la progenie directamente, pero si afectan el ambiente experimentado
por la cría. La producción de leche y la habilidad materna comprende el componente materno
del peso al destete.
Otros caracteres que tienen componente materno importante incluyen la distocia y la
sobrevivencia. El componente directo de la distocia está relacionado con el tamaño y la forma
del feto. El componente materno está asociado con el tamaño y la conformación de la pelvis
de la madre y otros factores fisiológicos y psicológicos más sutiles. El componente directo de la
sobrevivencia es una función de esos genes en los animales jóvenes que afectan la fortaleza
física, la respuesta inmune, y el instinto de supervivencia. El componente materno se relaciona
con la habilidad de la madre para amamantar y proteger a su cría.
Los procedimientos BLUP son capaces de separar los componentes directos y maternos de un
carácter, proveyendo predicciones para ambos. En el caso del peso al destete, las predicciones
están disponibles para el componente de crecimiento, así como para el componente leche/
habilidad materna.
Se puede cuestionar cómo es posible eso, dado que el componente materno del peso al
destete no se mide directamente. Nadie ordeña cerdas, ovejas o vacas de carne y registra sus
rendimientos lecheros. En su lugar la información para la predicción del componente materno
proviene de los mismos pesos al destete. La verdadera explicación de esta aparente paradoja
yace en la solución simultánea de ecuaciones, pero se puede pensar en ellos y no
incorrectamente como una simple cuestión de substracción (resta).
Supongamos por ejemplo, que un macho produce un gran número de hijos y de los pesos al
destete de éstos obtenemos una predicción confiable de los valores de cría de ese macho para
el carácter tasa de crecimiento predestete - el componente directo del peso al destete. Las
hijas de este macho son entonces apareadas y producen su propia progenie. Nosotros ya
tenemos una expectativa de los pesos al destete de esa progenie – nietos del macho – basada
en lo que ya conocemos sobre el potencial crecimiento de sus madres. Si los pesos al destete
de estos nietos exceden dichas expectativas, entonces se puede asumir que la diferencia es
debida a la habilidad materna superior de sus madres (producción de leche superior).
Por el otro lado, si los pesos al destete de los nietos no alcanzan las expectativas, la diferencia
es atribuida a la habilidad materna inferior de sus madres. Así la predicción para el
componente materno del peso al destete, está determinado por la diferencia entre la
performance real y las expectativas de performance al destete. Toda la situación es
complicada por el hecho de que el macho mismo pudo haber sido apareado con las mejores o
peores hembras que la media, y sus hijas con machos peores o mejores que la media, pero
BLUP toma en cuenta eso, y otras consideraciones igualmente confusas -
Una discusión de los componentes directos y maternos de los caracteres no estaría completa
sin mencionar un tercer componente, el componente paterno. Una definición de componente
paterno, análoga a la de componente materno diría que el componente paterno de un
carácter es el efecto de los genes en el padre de un individuo que influyen sobre su
performance a través del ambiente provisto por el padre. Sin embargo, para los animales
domésticos, una definición de este tipo es rara, los machos tienen muy poco que ver con la

Introducción a la Producción Animal - FCV - UNNE
2011
crianza de sus hijos y por lo tanto no tienen efecto ambiental sobre la performance de estos.
Efectos paternos sí existen para especies de pájaros silvestres y para algunos – jacanas y
pingüino emperador, por ejemplo – los componentes paternos de incubabilidad y
sobrevivencia de los pichones pueden ser hasta más importantes que el componente materno.
Una segunda definición de componente paterno se aplica a los caracteres de fertilidad. Las
medidas de fertilidad, consideradas caracteres de la madre o de la cría, pero que están
afectadas por la fertilidad del macho y su habilidad física para el servicio, se dice que tienen un
componente paterno. Hablamos por ejemplo, del componente paterno para la tasa de
concepción. Esta segunda definición tiene una mayor aplicación práctica en el mejoramiento
de las especies domésticas.
Componente directo: el efecto de los genes de un individuo sobre su propia performance.
Componente materno: el efecto de los genes de la madre de un individuo que influencia en la
performance de este a través del ambiente provisto por su madre.
Componente paterno: los caracteres de la madre o de la cría que son afectados por la
fertilidad y habilidad para reproducirse del macho también se dice que tienen un componente
paterno.
Predicción de varios tipos de valores
Los procedimientos BLUP, típicamente producen EBVs o EPDs para los componentes directos
de los caracteres, como así también para caracteres con componentes maternales
importantes, (EBVs o EPDs para estos también). Sin embargo, las predicciones de los análisis
BLUP no se limitan solamente a eso.
También es posible la predicción de los efectos ambientales permanentes para caracteres
repetidos, y eso significa que BLUP puede ser usado para predecir la habilidad de producción.
Así
PA = BV + Ep
Donde se asume que Ep contiene el efecto permanente y el valor combinatorio de los genes,
entonces
= +
Podemos calcular la MPPA de una hembra lechera para producción de leche asumiendo su
valor de cría estimado y la predicción de su efecto ambiental permanente. En ese cálculo, la
información utilizada pertenece a la propia performance de lactación de la hembrea, y los
registros de lactación de todo tipo de parientes.
Otro valor comúnmente estimado por los procedimientos BLUP es el llamado valor materno
total (BVtm). EL valor materno total combina los valores de cría para los componentes maternos
y directos del carácter.
Por ejemplo, el valor materno total de una vaca de carne para peso al destete representa su
habilidad genética para producir peso al destete. Como tal, incluye los efectos de sus genes

Introducción a la Producción Animal - FCV - UNNE
2011
para la producción de leche y su habilidad materna (el componente maternal para peso al
destete) así como también los efectos de sus genes para la tasa de crecimiento predestete que
ella transmite a su ternero – el componente directo del peso al destete. (Se usa una vaca de
carne en este ejemplo. Predecimos valores maternos totales para machos también. Pero
debido a que los genes del padre para el componente materno del peso al destete no son
expresados en el peso al destete de su hijo, un ejemplo utilizando un macho es más difícil de
entender).
En forma de ecuación,
Donde los subíndices WW, m y d representan el peso al destete, maternal y directo
respectivamente. El valor materno total de la vaca contiene todo su valor de cría para el
componente materno de peso al destete y solo la mitad de su valor de cría para el
componente directo de peso al destete. Esto es debido a que todos sus genes que influencian
el componente producción de leche / habilidad materna afectan a la performance de su
ternero, pero debido a que transmite solo la mitad de los genes al ternero, solo la mitad de sus
genes que influencian la tasa de crecimiento predestete afecta la performance del ternero. En
términos de predicciones genéticas
Y
Los EBVs maternos totales y los EPDs maternos totales son fáciles de confundir. EL EBV
materno total de una hembra es una predicción de su habilidad genética para producir algo
medido en su hijo. Es nuestra expectativa de su propia producción (excluyendo los efectos
ambientales). El EPD materno total de un individuo, por el otro lado, es nuestra expectativa
sobre la producción de las hijas de un individuo.
El EVB materno total de una vaca para peso al destete es una predicción de la parte heredable
de su habilidad para producir el peso al destete del ternero – su valor de cría para la
producción de peso al destete. Podemos ir un paso más allá y calcular el MPPA de una vaca
para peso al destete. Esto podría incluir cualquier efecto ambiental permanente en la
producción de peso al destete. Matemáticamente

Introducción a la Producción Animal - FCV - UNNE
2011
Los análisis BLUP también producen estimativas de los efectos ambientales comunes a todos
los miembros del grupo contemporáneo (Ecg). A pesar de que no son rutinariamente
utilizados, podrían ser útiles desde el punto de vista del manejo. Podrían indicar situaciones
donde los calendarios sanitarios son inefectivos, las pasturas están siendo sobrepastoreadas,
etc. Cuando las estimaciones de los efectos de grupos contemporáneos en una población son
trazadas a través del tiempo, revelan la tendencia ambiental – cambio en la performance
media de una población a través del tiempo causada por los cambios en el ambiente.
Valor materno total (BVtm): Una combinación de los valores de cría para los componentes
directo y materno de un carácter. El valor materno total de una hembra representa la parte
heredable de su habilidad para producir una cantidad que es medida en sus hijos.
Tendencia ambiental: cambio en la performance media de una población a través del tiempo
causada por los cambios en el ambiente.
Predicciones para todos los animales
Los procedimientos BLUP pueden ser utilizados para generar predicciones de cualquier animal
en una población. Las predicciones están disponibles para machos y hembras, padres y no
padres, animales que todavía no han sido concebidos y animales que han muerto hace mucho
tiempo, y animales con registros de performance o animales sin ellos.

Introducción a la Producción Animal - FCV - UNNE
2011
Evaluación genética a gran escala: SUMARIO DE PADRES
A continuación se tratan las evaluación genética a gran escala, como se ve un sumario de
padres y de donde proviene la información de los mismos, que información hay disponibles
sobre los no padres, cómo debe ser interpretada la información de la evaluación genética, que
trampas hay que tener en cuenta, y que alternativas hay a los programas convencionales de
evaluación a gran escala.
La evaluación genética a gran escala se refiere a la evaluación genética de grandes
poblaciones. Típicamente, estas poblaciones son razas enteras en un país o incluso dentro de
un área geográfica más grande (ej: PanAm, INTERBULL).
Debido a que la información utilizada para la evaluación genética a gran escala proviene de
muchos criadores y es procesada centralmente, la evaluación genética a gran escala es un
esfuerzo corporativo que involucra a los criadores, las asociaciones de criadores, y
profesionales en tecnología de mejoramiento animal.
El propósito de la evaluación genética a gran escala es simple – permitir la comparación
genética de animales en diferentes rebaños o lotes. ¿Por qué es esto importante? Suponga que
usted es dueño de un padre que piensa que es sobresaliente. Su propia performance y los
registros de progenie en su rebaño o lote son excelente, y usted está convencido que es el
mejor que jamás ha criado. Pero sin algún mecanismo para compararlo con los padres de otros
criadores, no tiene una forma objetiva de saber que tan bueno es dentro de la raza entera. La
evaluación genética a gran escala provee ese mecanismo.
La evacuación genética a gran escala acelera la tasa de cambio genético en una población.
Permitiendo una comparación directa de los animales de diferentes rebaños o lotes,
efectivamente, permite a los criadores seleccionar individuos de un pool mayor de candidatos.
En vez de estar limitados a los animales sus animales, los criadores pueden seleccionar de una
población más grande – una raza entera. Y así como es mucho más fácil formar grupos de
atletismo en una escuela grande que en una escuela pequeña porque en la grande hay más
atletas de los cuales elegir, también es más fácil encontrar reproductores verdaderamente
sobresalientes en una población grande que en una pequeña. La evaluación genética a gran
escala permite un incremento en la intensidad de selección.
La evaluación genética a gran escala también aumenta la tasa de cambio o progreso genético
incrementando la precisión de la selección. Las bases de datos de las razas contienen una
enorme cantidad de información, mucho más que la cantidad de información disponible en
cualquier rebaño o lote. Cuando los registros de razas enteras son utilizados para la predicción,
la exactitud de la predicción aumenta en virtud al volumen completo de información
disponible.
Evaluación genética a gran escala: La evaluación genética de grandes poblaciones –
típicamente razas enteras

Introducción a la Producción Animal - FCV - UNNE
2011
HISTORIA DE LAS COMPARACIONES GENETICAS
Sin contar las carreras de caballos, que han sido usadas para comparar animales de los
diferentes rebaños por milenios, las más recientes comparaciones en rodeos fueron
probablemente aquellas desarrolladas en el ganado lechero europeo cerca del comienzo del
siglo veinte. Estos involucraban test de progenie para ganado lechero. Test de progenie
similares se iniciaron en la lechería en Estados Unidos en la década del 30.
Algunas de las primeras comparaciones en cerdos, ovejas y bovinos de carne fueron provistos
por las centrales de prueba. Los verracos, carneros y toros de diferentes establecimientos se
llevan a un mismo lugar, la central de prueba, donde son alimentados y manejados juntos por
un determinado período de tiempo. Luego son comparados para caracteres como ganancia
diaria de peso, conversión alimenticia y otras medidas físicas como en lanares peso de vellón
etc. Las centrales de prueba permiten a los animales de diferentes rebaños o lotes competir
directamente entre ellos, y han sido usadas históricamente como una herramienta de
marketing para los criadores de reproductores y como foro para promover el uso de
información de performance. Pero la habilidad de los test realizados en centrales para
comparar el mérito genético de los animales es limitada. Las comparaciones están restringidas
a unos pocos caracteres medidos en la estación de prueba y están basados en la información
de performance del individuo solamente, y el ambiente previo al inicio de la prueba puede
afectar la performance en el test, dando una ventaja a los animales de algunos sitios y
desventajas a animales de otros. Desde la llegada de las BLUPs – con su habilidad de explicar
las diferencias ambientales y genéticas entre grupos contemporáneos – y la evaluación a gran
escala utilizando BLUP, los test de progenie en centrales de prueba han perdido mucha de su
justificación genética, sobre todo en algunas especies.
Las primeras evaluaciones de padres para ganado de carne aparecieron en los EEUU, y Canadá
en los años 70. Con el advenimiento de computadoras de gran velocidad, el análisis usando
BLUP y los procedimientos parecidos al BLUP se convirtieron en la norma para la cría de
ganado en los 80 en esos países y Europa. El cambio ocurrió rápido, y la evaluación genética a
gran escala se torno común en países industrializados para la producción de leche y carne
bovina, y los programas para ovejas y cerdos se desarrollaron en los últimos años. Las
evaluaciones de caracteres de performance en equinos también están disponibles – algunas
técnicamente más sofisticadas en Europa, y menos sofisticadas en Estados Unidos.
Las primeras evaluaciones eran test diseñados, cuidadosamente monitoreados, test de
progenie, libres de algunas fuentes de sesgo, como el apareamiento no al azar y el refugo por
baja performance. Los test diseñados eran caros y necesariamente limitados en cantidad.
Con el poder de los procedimientos estadísticos modernos, de explicar un mayor número
fuentes de sesgos, la tendencia general en la evaluación a gran escala ha sido hacia el uso de
datos de campo, datos que son regularmente reportados por criadores individuales a las
asociaciones de cría o agencia gubernamentales. Los datos de campo provienen de enorme
cantidad de información – mucha más información de la que los test diseñados pueden
proveer.

Introducción a la Producción Animal - FCV - UNNE
2011
Las evaluaciones genéticas a gran escala usando información de campo requieren experiencia
técnica avanzada, programas informáticos complejos y computadoras poderosas. Son
conducidos por lo general una vez al año por especialistas en las asociaciones de cría, en el
gobierno, en universidades, o (menos comúnmente) en compañías o cabañas privadas.
Test de progenie en centrales de prueba: Test diseñado para comparar la performance de
animales (usualmente machos jóvenes) de diferentes rebaños o lotes para caracteres como
tasa de crecimiento y conversión alimenticia, alimentándolos en un sitio central.
Test diseñados: Test de progenie, cuidadosamente monitoreado libre de fuentes de sesgo
como apareamiento al no al azar y refugo por baja performance.
Datos de campo: datos que son regularmente reportados por criadores individuales a las
asociaciones de cría o agencia gubernamentales.
SUMARIOS DE PADRES
El producto más viable de la evaluación genética a gran escala es el sumario de padres. Los
sumarios de padres son listas de predicciones genéticas, valores de precisión, y otra
información útil sobre padres en una raza.
Los sumarios varían en formato de especie en especie y de raza en raza. Sin embargo,
típicamente, están compuestos de una sección introductoria, seguido de una lista de datos de
los padres.
La sección introductoria de un sumario de padres es muy informativa. Podría contener una
explicación de los datos, incluyendo un glosario de términos, las características que deben
reunir los machos, necesarias para aparecer en el sumario, una tabla de parámetros genéticos
estimados (heredabilidades y correlaciones usadas para calcular las predicciones),
distribuciones de predicciones dentro de una raza (en forma de gráficos que usualmente se
ven como distribuciones normales y/o tablas de percentiles), una tabla que convierte las
precisiones a intervalos de confianza o posibles valores de cambio (definido luego en esta
sección), y gráficos de tendencia genética.
La lista de padres típicamente incluye tres tipos de datos:
¤ identificación animal;
¤ diversa información fenotípica sobre el padre incluyendo características de herencia
simple como el color del manto, defectos genéticos, etc.;
¤ predicciones y medidas de precisión.
En la 12.1 se presenta una muestra de una lista de un supuesto sumario de padres de toros
de aptitud carnicera. Los toros son identificados por nombre, nombre del padre, y abuelo
materno, número de registro y dueño(s). Otros ítems de información incluyen la fecha de
nacimiento del animal y un código o sufijo indicando características de herencia simple
como los nombres del segundo, tercero y cuarto toro en la lista que poseen el sufijo “R”
(portadores de color de capa rojo). El carácter en la primera columna indica si el toro se
encuentra en una jerarquía particularmente alta dentro de la raza, para un carácter

Introducción a la Producción Animal - FCV - UNNE
2011
específico. RNC Crescendo 538 tiene un EPD especialmente alto para peso al año y es un
líder para ese carácter. RNC Prelude 732 tiene un EPD especialmente bajo para peso al
nacimiento y es líder en esa categoría. Un sumario de padre de una raza para carne real,
presenta información – incluyendo más EPDs – de lo que se muestra en este ejemplo
simplificado. Los sumarios de padres lecheros presentan mucha más información.
Tabla 12.1 Segmento de muestra de la lista de un sumario de padres de bovinos
para carne Nombre (sufijo)
Reg # Fecha de nacimiento
Dueño, Estado o provincia
Trt. Ldr.
Peso al nacer EPD ACC
Peso al destete EPD ACC
Peso al año EPD ACC
Leche EPD ACC
Total mat
RAB George Washington
129755 16/3/81 N. Maclean MT
8,4 0,93
31 0,93
53 0,92
16 0,91
31
RNC Crescendo 538
181650 11/3/85 T.Morrison NY Y 0.9 0.87
39 0,86 71 0,85
-12 0,80
7
RCN Intonation 338
153082 09/3/83 W. Stegner, VT 1,3 0,86
24 0,85
43 0,84
11 0,81
23
RCN Prelude 732
274698 11/3/87 T. Williams, UT M. Golden, CO
B -5,6 0,87
5 0,86
10 0,85
20 0,78
22
RD Madison Ave 6X
329877 08/3/90 J. Salinger,NH E. Hemingway, ID
1,1 0,87
39 0,63
54 0,51
8 0,40 27
Sumario de padres: listas de predicciones genéticas, valores de precisión, y otra información
útil sobre los padres en una raza.
Predicciones
Las predicciones genéticas publicadas en los sumarios de padres modernos y actualizados son
las diferencias esperadas en la progenie o EPDs (DEPs). Los sumarios de animales lecheros
utilizan diferentes términos: Diferencias predichas (PDs) y habilidades transmitidas esperadas
(ETAs o HTP), pero EPD, PD y ETA significan lo mismo. (Aquí se utiliza EPD). Las diferencias
esperadas de progenie (en lugar de valores de cría estimados o EBVs) son comparativamente
fáciles de interpretar. Representan la performance esperada en la progenie del padre.
La lista de padres mostrada en la tabla 12.1 provee ejemplos de EPDs para los diferentes
componentes de los caracteres: directo, materno, y materno total. Los EPD para peso al año y
peso al destete son medidas de componentes directos. Predicen la performance de la progenie
que es atribuible a los genes heredados del padre. Los EPDs de peso al año y peso al destete
predicen el potencial de crecimiento, en diferentes edades, de los terneros de un padre.
El EPD leche es una medida del componente materno del carácter peso al destete. Predice la
habilidad lechera y materna (medida en libras de peso al destete) de las hijas de ese padre. El
EPD materno total mide la combinación de los componentes directo y materno del peso al
destete conocido como peso al destete materno total. Predice la habilidad total de las hijas de
un padre para producir el peso al destete del ternero.

Introducción a la Producción Animal - FCV - UNNE
2011
Medidas de precisión
La precisión es definida como una medida de la fuerza de la relación entre valores reales y sus
predicciones, y en términos matemáticos, es la correlación entre valores verdaderos y sus
predicciones. Podríamos llamar a la precisión definida así precisión clásica (el término para
precisión que comúnmente aparece en publicaciones lecheras es repetibilidad – no confundir
con el concepto de Repetibilidad dado con anterioridad).
En los sumarios, la precisión clásica sería la correlación entre diferencias de progenie
verdaderas y EPDs ( ). Sin embargo, los valores de precisión publicados en los sumarios
de padres raramente representan esta correlación. En su lugar, son funciones de precisiones
clásicas. La razón para esto es que las precisiones clásicas se acercan a 1 con relativamente
poca información y no diferencian suficientemente entre padres que tienen muchos datos y
aquellos que tienen una cantidad moderada.
En la tabla 12.2 están los EPD y precisiones publicadas de algunos padres catalogados en la
tabla 12.1, así como algunas medidas de precisión alternativas incluyendo las precisiones
clásicas. Note cuán altas son las precisiones clásicas para los primeros cuatro padres de la lista.
De hecho son tan altas, que es difícil ver mucha diferencia en precisión entre el primer toro,
cuyos EPDs son derivados de cantidades grandes de información, y los siguientes tres toros
cuyos EPDs derivan de cantidades de información moderada. Las precisiones publicadas son
mucho mejores al respecto. Al igual que las precisiones clásicas, van de 0 a 1 de manera que se
puede interpretar de la misma forma que éstas.
Otra forma de expresar precisión es con intervalos de confianza, un rango de valores dentro
del cual esperamos –con una probabilidad dada, un grado determinado de confianza – que se
encuentre un valor verdadero de interés.
Rangos del 68% de confianza para los EPDs de peso al año de dos padres de la tabla 12.2 son
ilustrados en la figura 12.1.

Introducción a la Producción Animal - FCV - UNNE
2011
RAB George Washington está bien evaluado, teniendo una precisión publicada para EPD de
peso al año de 0,92. El 68% de rango de confianza de este EPD es pequeño – de 51,3 a 54,7 lb.
En otras palabras, las chances de su diferencia de progenie verdadera entre 51,3 y 54,7 lb son
un poco mejor que 2/3 (dos de tres). La verdadera diferencia de progenie de RAB George
Washington podría estar fuera de este rango, pero incluso si está, es poco probable que esté
muy por fuera de este.
RD Madison Ave 6X tiene relativamente poca información como evidencia para su precisión
publicada de 0,51 para peso al año. El rango de confianza del 68% para este EPD para peso al
año es mucho más amplio – de 43,7 a 64,3 lb. Claramente hay más error potencial asociado
con el EPD para peso al año de RD Madison Ave 6X que con RAB George Washington.
Las predicciones genéticas para individuos cambian a través del tiempo, dependiendo de
cuanta más información se incluya en los cálculos usados en las evaluaciones sucesivas.
Una medida de predicción que indica la cantidad potencial de un futuro cambio en una
predicción es el llamado cambio posible (PC). El término estadístico de cambio posible es el
error estándar de predicción. Una predicción ± cambio posible, es simplemente una forma
alternativa de expresar un rango de confianza del 68%. Por ejemplo, el EPD de RAB George
Washington ± PC para peso al año es 53 ±1,7, lo que corresponde a su rango de confianza de
68% de 51,3 a 54,7 lb (vea la figura 12.1). Igualmente, RD Madison Ave 6X ± PC para peso al
año es 54 ± 10,3 lb, correspondiente al rango de confianza de 68%, de 43,7 a 54,3 lb. Los
rangos de confianza y los posibles cambios para los cinco padres listados en la tabla 12.1 son
mostrados en la tabla 12.2.

Introducción a la Producción Animal - FCV - UNNE
2011
Las precisiones, rangos de confianza, y los valores de cambio posibles están estrechamente
relacionados. Cuanto mayor (mejor) es la precisión de una predicción, menor es el intervalo de
confianza y los cambios posibles asociados.
Las precisiones tiene la ventaja sobre los rangos de confianza y los cambios posibles en que no
se necesita saber nada acerca de la variabilidad del carácter para interpretar una precisión.
Una precisión publicada de 0,95 es una precisión alta independientemente del carácter. No se
puede decir lo mismo para un rango de confianza o un cambio posible. Por ejemplo, un cambio
posible de ± 2,5 significa una cosa si el carácter es peso maduro en ganado de carne y algo
bastante diferente si el carácter es días para alcanzar las 230 lb en cerdos. En el primer caso,
sugiere una precisión muy alta; en el segundo caso, sugiere una precisión muy baja. Por otro
lado, si usted no está familiarizado con el carácter que está siendo medido, los rangos de
confianza y los posibles cambios son más gráficos y probablemente indicadores más
informativos de la confianza de la predicción.
Tabla 12.2 Medidas de precisión alternativas para padres listados en la tabla 12.1
Nombre del padre
Peso al nacimiento
Peso al destete Peso al año Leche
RAB George Washington
EPD 8,4 31 53 16
Precisión publicada
0,93 0,93 0,92 0,91
Precisión ( )
0,998 0,998 0,997 0,996
Rango de confianza al 68%
8,2 a 8,6 30 a 32 51,3 a 54,7 15,4 a 16,6
Cambio posible ± 0,20 ± 1,0 ±1,7 ±0,6
RNC Crescendo 538
EPD 0,9 39 71 -12
Precisión publicada
0,87 0,86 0,85 0,80
Precisión ( )
0,992 0,990 0,989 0,980
Rango de confianza al 68%
0,54 a 1,26 37 a 41 67,8 a 74,2 -13,4 a -10,6
Cambio posible ± 0,36 ±2 ±3,2 ±1,4
RCN Intonation 338
EPD 1,3 24 43 11
Precisión publicada
0,86 0,85 0,84 0,81
Precisión( ) 0,990 0,989 0,987 0,982
Rango de confianza al 68%
0,91 a 1,69 21,9 a 26,1 39,6 a 46,4 9,6 a 12,4
Cambio posible ±0,39 ±2,5 ±3,4 ±1,4
RCN Prelude 732
EPD - 5,6 5 10 20
Precisión publicada
0,87 0,86 0,85 0,78
Precisión( ) 0,992 0,990 0,989 0,975
Rango de confianza al 68%
-5,96 a -5,24 3 a 7 6,8 a 13,2 18,2 a 21,6
Cambio posible ± 0,36 ± 2 ±3,2 ±1,6
RD Madison Ave 6X
EPD 1,1 39 54 8
Precisión publicada
0,67 0,63 0,51 0,40

Introducción a la Producción Animal - FCV - UNNE
2011
Precisión( ) 0,944 0,929 0,872 0,800
Rango de confianza al 68%
-0,77 a 2,97 33,8 a 44,2 43,7 a 64,3 3,7 a 12,3
Cambio posible ± 1,87 ±5,2 ± 10,3 ±4,3
Repetibilidad: (en publicaciones lecheras) precisión de una predicción
Intervalo de confianza: es un rango de valores dentro del cual esperamos –con una
probabilidad dada, un grado determinado de confianza – que se encuentre un valor verdadero
de interés.
Cambio posible (PC) o error estándar de predicción: Una medida de predicción que indica la
cantidad potencial de un futuro cambio en una predicción.
Importancia de los sumarios de padres
Los sumarios de padres son importantes primero y principal por la selección. Aumentan la
efectividad de la selección en dos formas: (1) expandiendo el pool de padres disponibles, así
aumenta la intensidad de la selección, (2) usando grandes cantidades de datos para la
predicción genética, así aumenta la precisión.
¿Por qué los sumarios de padres y no de madres? Como ya fue explicado, la selección de
machos es mucho más importante que la selección de las hembras en términos de su efecto en
la tasa de cambio genético en una población. La selección del padre realmente conduce al
cambio genético. Otra razón es que los padres del rebaño o lote de otra persona son más
accesibles que las madres. Muchos hijos pueden ser producidos por el precio de un padre. Y si
es posible biológica y políticamente dentro de especies o raza usar la inseminación artificial, el
acceso a padres sobresalientes es incluso más fácil.
Los sumarios de padres también pueden ser utilizados para manejar el riesgo de selección, el
riesgo de que los valores de cría verdaderos de un padre sean significantemente más pobres
de lo esperado. Los padres cuyas precisiones publicadas son altas deberían criar como se
aconseja en el sumario. Los padres con precisiones bajas puede que no. La información de
precisión provista por los sumarios de padres permite a los criadores tomar la cantidad de
riesgo que quieran.
Los sumarios de padres, son también herramientas de marketing. Muy parecido a las
exposiciones y carreras, proveen un foro de competencia para los criadores. Si un padre tiene
EPDs particularmente deseables, esos EPDs se vuelven puntos de publicidad para su dueño.
Evaluación genética para individuos que no son padres
La evaluación genética a gran escala no está limitada a la producción de EPDs para padres. Los
análisis modernos BLUP producen predicciones para todos los animales. Por supuesto son
demasiados para ser publicados en un solo documento. Usualmente las asociaciones de
criadores o las agencias del gobierno mandan a los criadores en forma impresa la información
que contiene los EPDs y las precisiones de sus animales.

Introducción a la Producción Animal - FCV - UNNE
2011
Tipos de EPDs
Los criadores pueden recibir varios tipos de EPDs. Los EPD paternos son EPDs para animales
con datos de progenie. Son el producto inmediato del análisis de gran escala y vienen con
medidas de precisión asociadas. Los EPDs publicados en los sumarios de padre son ejemplo de
EPDs paternos.
Con algunos modelos BLUP, los EPDs para los individuos que no son padres se calculan en
forma un poco diferente de los EPDs paternos. Los EPDs no paternos de este tipo típicamente
no vienen con medida de precisión publicada. Si un individuo que no es padre no tiene
información de performance que contribuya con su EPD, entonces ese EPD es necesariamente
una estimación de pedigrí o genealógica, y es simplemente el promedio de los EPDs de los
padres del individuo. Las estimaciones de este tipo son siempre de precisión baja, aunque
podrían no serlo si el carácter en consideración es altamente heredable y/o es un carácter
repetido. Por ejemplo, un caballo castrado (claramente no es padre) con una larga historia en
las carreras podría posiblemente tener un EPD altamente preciso para la habilidad de la
carrera. (Por supuesto una diferencia esperada de progenie para un castrado es una idea sin
sentido)
Debido a que los análisis EPD hechos y derechos son realizados en meses hasta un año, a
menudo hay necesidad de actualizar los EPDs entre análisis. Por ejemplo muchos análisis de
información de ganado de carne son realizados luego de que se reporta la información sobre
destete para que los sumarios de padres estén disponibles antes de la próxima temporada de
servicios. Sin embargo, mientras tanto, se recoge información adicional de performance post
destete de los animales jóvenes. Los EPDs interinos - EPDs actualizados, calculados entre
análisis BLUP - incorporan la nueva información. En un sentido los EPDs interinos son
“salidas”. Solo una porción de la información disponible es incluida en los procedimientos
cortos usados para calcularlos. Sin embargo, usualmente son mejores, que los EPDs viejos que
reemplazan.
EPD paternos: son EPDs para animales con datos de progenie y vienen con medidas de
precisión asociadas.
EPDs no paternos: Un EPD para un animal sin datos de progenie, típicamente no vienen con
medida de precisión publicada.
Estimación de pedigrí: Una predicción genética basada solamente en datos de progenie.
EPDs interinos: Un EPD actualizado calculado entre los análisis BLUP e incorpora nueva
información.
COMO INTERPRETAR LA INFORMACIÓN DE LA EVALUACIÓN GENÉTICA
Los productos de la evaluación a gran escala – EPDs y medidas de precisión – pueden ser
confusas y son a menudo malinterpretadas.
1. Las predicciones son para comparar animales

Introducción a la Producción Animal - FCV - UNNE
2011
Los EPDs no predicen performance de por sí. Por ejemplo, no se puede decir que las hijas de
un verraco con un EPD para peso de la camada a los 21 días de + 5lb van a producir camadas
de un promedio de 105 lb. Su producción real va a depender de otras cosas aparte de los
genes heredados del padre. EL ambiente es importante. Si las hijas del verraco experimentan
un mejor o peor ambiente que el promedio (es decir si los efectos de grupo contemporáneo
(Ecg) son mejores o peores que el promedio), entonces la performance estará afectada de
acuerdo a eso. La performance también será afectada si los cerdos son consanguíneos o cruza,
es decir, por el valor combinatorio de los genes (GCV) y por los valores de cría de las madres de
las cerdas y por los valores de cría de los verracos con los cuales se aparean.
El EPD del verraco para peso de la camada a los 21 días es la diferencia esperada entre la
performance promedio de sus hijas y la performance promedio de las hijas de un verraco con
un EPD de cero – cuando todo lo demás es igual. Por supuesto, a menos que se esté
familiarizado con un verraco cuyo EPD para peso de la camada a los 21 días es exactamente
cero, esta no sería la comparación más informativa.
Una comparación más significativa sería entre un verraco con EPD de +5 lb con otro verraco de
interés cuyo EPD es de – 5lb. En este caso, esperamos que las hijas del primer verraco destete
camadas 7 lb (5-(-2) =7) más pesadas que las hijas del segundo verraco. Los EPDs son para
comparar animales.
Los criadores informados generalmente tienen una percepción sobre los niveles de EPDs que
mejor encajan en su ambiente, manejo, sistema de apareamiento y mercado. De la
experiencia, sabe como los hijos de los animales con diferencias de progenie particulares se
desempeñan usualmente en sus establecimientos. Para estos criadores, un EPD por si solo
significa un cierto nivel de performance. Sin embargo, tan pronto como dejamos el contexto
de una granja o rancho específico, la conexión entre EPDs y la performance actual se pierde.
Entonces los EPDs por si solos no son informativos, pero sí en cambio las diferencias en EPDS.
El significado del cero
Sería mucho más fácil interpretar EPDs si supiéramos que significa un EPD de cero. Pero así
como no podemos predecir la performance (en el sentido absoluto o no relativo) de las hijas
de un verraco con un EPD para peso de la camada a los 21 días de +5 lb, tampoco podemos
predecir la performance real del hijo de un animal con un EPD de cero.
Es un error asumir que cualquier nivel particular de EPD resultará en un cierto nivel de
performance en todos los ambientes y rebaños o lotes. De acuerdo con el modelo genético
para caracteres cuantitativos presentado anteriormente, la performance asociada con un EPD
de cero debe ser equivalente a µ, la performance promedio en una población. Tan práctica
como la definición puede ser, no se aplica al caso de evaluación genética a gran escala de
poblaciones que contienen grupos contemporáneos diversos genéticamente y han cambiado a
través del tiempo por la selección.
En la realidad, la definición del punto cero o base depende del modelo estadístico usado para
la predicción genética y en algunas características propias de los datos. Para propósitos
prácticos, el punto cero no tiene significado. Para darle un significado a la base, los que

Introducción a la Producción Animal - FCV - UNNE
2011
realizan las evaluaciones genéticas a gran escaña a veces ajustan los EPDs de maneras que la
base represente el EPD promedio de todos los animales nacidos en un año específico. Por
ejemplo, en la evaluación de cerdos, los EPDs podrían ser ajustados de manera que un EPD de
cero para peso de la camada a los 21 días represente el promedio de la raza para ese carácter,
digamos en 1985.
Los ajustes de EPD de este tipo no afectan las comparaciones de EPDs de ningún modo. Si la
diferencia entre los EPDs de dos animales era de 7 lb antes del ajuste, se mantendrá en 7 lb
después del mismo. Relacionar un EPD a un año base simplemente nos permite tener una
definición, aunque inadecuada, de cero.
Las diferentes razas en las especies tienen diferentes bases. Por esta razón (y por otras), los
EPDs no deberían ser comparados entre razas. Incluso si los EPDs de dos razas esta atados al
mismo año base, sus bases solo serían iguales si ambas razas estuvieran al mismo nivel
genético ese año. Encontrar formas confiables de comparar los EPDs a entre razas es uno de
los retos más importantes en mejoramiento animal.
Base: (en la evaluación genética a gran escala) El nivel de mérito genético asociado con un EPD
de cero.
Año base: (en la evaluación genética a gran escala): El año elegido para representar la base. El
EPD de todos los animales nacidos en el año base es cero.
Uso de las medidas de precisión
Las medidas de precisión proveen información sobre el riesgo de selección, el riesgo de que los
valores de cría verdaderos o la diferencia de progenie de un animal sean significativamente
peor o más pobre que lo esperado. EL uso de individuos con alta precisión de EPDs conlleva
pequeño riesgo de selección. El uso de individuos con baja precisión de EPDs es mucho más
riesgoso.
Las precisiones clásicas y publicadas no siempre pintan una imagen clara del riesgo de
selección asociado con el uso de un animal en particular. Los rangos de confianza son mejores
en este aspecto. Suponga, por ejemplo, que un criador de ganado de carne está interesado en
vender semen de uno o dos padres listados en la tabla 12.2: RCN Crescendo 538 y RD Madison
Ave 6X. Debido a su preocupación por la dificultad al parto, se quiere asegurar de que el toro
que elija no tiene una diferencia de progenie verdadera para peso al nacimiento mayor a +2 lb.
Los EPDs para peso al nacimiento de ambos padres son menores a 2. El EPD de RCN Crescendo
538 es + 9, y el EPD de RD Madison Ave 6X es de + 1,1. Sin embargo, las precisiones publicadas
difieren –0,87 y 0,67 respectivamente. A pesar del hecho que sus EPDs para peso al nacimiento
son casi idénticos, la diferencia en precisión le dice que RD Madison Ave 6X representa mayor
riesgo de selección. La pregunta es ¿Cuánto riesgo?
Los rangos de confianza proveen una pista. El rango de confianza del 68% para el EPD de RCN
Crescendo 538 es de +0,54 a +1,26. Eso significa que la probabilidad de que su diferencia de
progenie verdadera para peso al año esté dentro de este intervalo es de 68%. La probabilidad
de que su diferencia de progenie verdadera sea mayor a +1,26 es relativamente pequeña. Por
el otro lado el rango de confianza de 68% de RD Madison Ave 6X es de -7,7 a +2.97. Su

Introducción a la Producción Animal - FCV - UNNE
2011
diferencia de progenie verdadera para peso al nacimiento podría fácilmente estar por encima
de las + 2lb.
Si parecen insuficientes los rangos de confianza del 68%, se podría convertirlos en rangos de
confianza del 95% sumando un cambio posible a cada extremo de los rangos de 68%. Los
posibles cambios para EPDs de peso al nacimiento de RCN Crescendo 538 y RD Madison Ave 6X
son de 0,36 y 1,87 respectivamente, entonces sus rangos de confianza del 95% serían de +0,18
a 1,62 y – 2,64 a +4,84. Claramente con un rango de confianza del 95% tan amplio como de –
2,64 a +4,84 RD Madison Ave 6X no está muy bien evaluado para peso al nacimiento. Usarlo
sería invitar a un riesgo mayor para dificultad al parto.
Si los rangos de confianza o los posibles cambios no son publicados, a veces es posible
determinarlos por precisiones publicadas. La mayoría de los sumarios proveen tablas de
conversión con este propósito.
Comúnmente se comete el error de considerar diferencias pequeñas en EPDs como
significativas. Por ejemplo, las diferencias en EPDs de peso al nacimiento de RCN Crescendo
538 y RCN Intonation 338 es de 0,4 lb (tabla 12.2). La diferencia entre las diferencias de
progenie verdaderas es probablemente algo más que 0,4 lb, pero incluso si fuera 0,4 lb, tan
pequeña diferencia sería indetectable en todos excepto los rebaños más grandes. En la
mayoría de los casos, es mejor relegar los EPDs a categorías generales (ej. muy alta, alta,
mediana, baja, muy baja) que vincular significancia particular a los números.
Los rangos de confianza proveen los recursos para determinar si los EPDs de dos animales son
realmente diferentes. Los rangos de confianza para los EPDs de peso al nacimiento de los
padres listados en la tabla 12.2 son mostrados gráficamente en la figura 12.2. Los rangos de
confianza para RCN Crescendo 538, RCN Intonation 338 y RD Madison Ave 6X todos
superpuestos. Esto indica que las diferencias en estos EPDs no son significantes en el sentido
estadístico. Sería un error asumir que estos EPDS son realmente diferentes entre ellos. En
contraste los rangos de confianza para los EPDs de peso al nacimiento de RAB George
Washington y RCN Prelude 732 (+8,2 a +8,6 y -5,96 a -5,24, respectivamente) no están ni
remotamente cerca a sobreponerse. Se puede estar seguro de que los pesos al nacimiento
promedio producidos por estos dos padres serán diferentes entre sí y diferentes del promedio
de los pesos al nacimiento producidos por los otros tres padres también.
A veces se confunde el concepto de precisión de predicción con variabilidad en la performance
de los hijos. Por ejemplo se podría concluir de la tabla 12.2 que debido a que las precisiones de
RAB George Washington son altas y los rangos de confianza estrechos, este será padre de
terneros particularmente uniformes.
Igualmente se podría inferir que debido a que las precisiones de RD Madison Ave 6X son bajas
y los rangos de confianza amplia, será padre de terneros particularmente no uniformes. Esto,
por supuesto no tiene sentido. No tenemos razón para creer que los terneros del primer padre
serán más o menos variables en performance que los terneros del segundo padre. La precisión
de un EPD nos dice algo acerca de la confianza del EPD como predictor de la performance
media de los hijos de un individuo. No dice nada acerca de la variación de la performance de
los hijos.

Introducción a la Producción Animal - FCV - UNNE
2011
¿Evaluación o caracterización?
El uso de la palabra “evaluación” en el término “evaluación a gran escala” implica alguna
manera, juzgar los valores de los animales calculando sus predicciones genéticas. En un
sentido, lo estamos haciendo, pero debemos ser cuidadosos de no tomar los resultados de la
evaluación genética muy literalmente y confundir EPDs con valores. El animal con los EPDs más
altos, no es necesariamente el animal más valorable. Para muchos caracteres hay niveles
óptimos de EPDs que no son los más altos ni los más bajos, sino algún punto intermedio entre
los dos. El animal más valorable a menudo representa un balance de los EPDs para varios
caracteres. Los EPDs revelan las habilidades genéticas de un animal, pero no su valor. Tal vez
un mejor termino que “evaluación genética” sería “caracterización genética”.
TRAMPAS EN LA EVALUCACION GENETICA A GRAN ESCALA
El advenimiento de la evaluación genética a gran escala ha revolucionado el mejoramiento
animal para algunas especies. Ha cambiado la forma en la que se opera y acelera el cambio
genético. Sin embargo, eso no significa que la evaluación genética a gran escala no tenga
problemas. La siguiente discusión describe algunas de estas trampas.
Datos defectuosos
La evaluación genética no puede ser mejor que la información que contribuye con ella. Los
datos de campo son reportados por muchos criadores, y esos criadores varían mucho en su
conocimiento y en el interés en los principios del mejoramiento animal. Entonces por su propia
naturaleza, este tipo de información está sujeta a error, intencional o no.
Pedigree
En la evaluación genética moderna, las predicciones genéticas están basadas en gran parte en
la performance de los parientes. Por lo tanto, es importante que las relaciones genealógicas
sean usadas correctamente en la evaluación genética. Por ejemplo, un pedigrí estimado para
animales jóvenes es inútil si los padres del animal no han sido identificados.
La identificación de los padres no es usual en muchas especies, y ello causa errores en la
predicción. Afortunadamente, los errores de este tipo son generalmente de corta vida. Tan
pronto como un individuo tiene suficiente información de progenie, la información
proveniente del pedigrí es de poca relevancia. Las tecnologías que mejoran la identificación
animal seguramente beneficiarán las predicciones genéticas.
Registros de performance
A veces los criadores falsifican los registros de performance. Sucumben ante la tentación de
hacer ver a los animales mejores de lo que realmente son. Sin embargo, la falsificación
deliberada probablemente sea rara. Son más comunes los “cálculos estimados a ojo”. A veces
simplemente es más fácil suponer la performance que medirla cuidadosamente. Por ejemplo,
muchos pesos al nacimiento de ganado de carne no son medidos con balanzas. Son
simplemente “medidos a ojo”. Atajos como éstos reducen la confianza de las predicciones.

Introducción a la Producción Animal - FCV - UNNE
2011
Una fuente común de sesgo en datos de campo es el reporte incompleto. El reporte
incompleto se refiere al reporte de solo los buenos registros de performance. Para ver el
efecto del reporte incompleto, imagine que un criador solo reporta registros del 20% mejor de
la progenie de un padre. Seleccionando la información, el criador ha hecho parecer al padre
mejor de lo que realmente es.
Al contrario de los sesgos causados por el refugo para baja performance, el sesgo causado por
el reporte incompleto no puede ser explicado con procedimientos BLUP avanzados. Mientras
que los primeros registros de los animales refugados por performance pobre son accesibles,
los registros de los animales no reportados no lo son. Es como si estos animales nunca
existieron. Todos los registros deberían ser reportados, los buenos y los malos.
Reporte incompleto: se refiere al reporte de solo algunos registros de performance a una
asociación de cría o agencia de gobierno.
Información utilizada para ajustar los registros de performance
Los registros de performance pueden ser reportados precisamente, pero si la información
utilizada para ajustar esos registros para efectos ambientales conocidos es errónea, los
registros de performance ajustados también serán erróneos. Por ejemplo, si la fecha de
nacimiento para un conjunto de corderos mellizos es registrada un mes después de la
verdadera fecha de nacimiento, los ajustes de los pesos subsecuentes de los corderos para su
edad serán defectuosos. Parecerá que crecieron más rápido de lo que realmente lo hicieron.
Grupos contemporáneos
Los animales son a veces reportados como miembros de un grupo contemporáneo
equivocado. Un animal criado en el grupo contemporáneo A es raramente reportado en un
grupo B, pero los criadores a veces reportan animales como miembros de un solo grupo
contemporáneo cuando, para ser correcto, deberían dividirlos en dos o más grupos
contemporáneos. El mal agrupamiento de los animales causa problemas especialmente
severos cuando los animales que recibieron trato preferencial (animales de competencia por
ejemplo) son reportados en el mismo grupo contemporáneo con animales que no han recibido
tratamiento preferencial. Los animales que reciben tratamiento preferencial parecen
desmesuradamente buenos en comparación con otros.
Falta de conexión entre grupos contemporáneos
Las técnicas BLUP resuelven los efectos de grupos contemporáneos así como también las
predicciones genéticas. Es esta habilidad de separar los efectos ambientales comunes a todos
los miembros del grupo contemporáneo, de los efectos genéticos, lo que hace a BLUP tan útil.
Pero con el objetivo de obtener buenas estimativas de efectos de grupos contemporáneos (y
por lo tanto buenas predicciones genéticas), es necesario que los animales de diferentes
grupos contemporáneos estén relacionados o conectados. La mejor situación sería si todos los
padres en una raza tuvieran progenie en todos los grupos contemporáneos. Entonces cada
grupo contemporáneo estaría conectado con todos los otros grupos contemporáneos por
familias de medio hermanos. Por supuesto, en las poblaciones reales las conexiones de este
tipo no existen. Las relaciones genealógicas entre animales en diferentes grupos

Introducción a la Producción Animal - FCV - UNNE
2011
contemporáneos son a menudo bastante lejanas. Cuanto más cercanas sean las relaciones
mejor la conexión entre grupos, y mejores las predicciones genéticas resultantes.
La estructura de una población afecta la conexión. Las poblaciones en las cuales los padres
tienen progenie en muchos rebaños o lotes tienden a tener buena conexión. Las poblaciones
en la cuales hay poco intercambio de germoplasma entre rebaños o lotes tienden a tener
conexión pobre. La inseminación artificial es una herramienta extremadamente importante
para incrementar la conexión en una población.
Conexión: El grado en que los datos de diferentes grupos contemporáneos dentro de una
población pueden ser comparados como resultado de relaciones genealógicas entre animales
de diferentes grupos.
Interacción genotipo ambiente
Los datos para la evaluación genética a gran escala a menudo provienen de diferentes
ambientes. Esto no es un problema siempre y cuando no haya una interacción genotipo
ambiente (GxE) significativa. Recuerde que las interacciones GxE ocurren cuando la diferencia
de performance entre dos (o más) genotipos cambia de ambiente en ambiente.
Si la progenie de un padre se desempeña bien en relación a la progenie de otros padres en un
ambiente, pero se desempeña pobremente en relación a la progenie de otros padres en otro
ambiente, entonces existe una interacción GxE seria. Los EPDs publicados de un padre, que
están basados en la performance de su progenie en ambos ambientes, podría no reflejar su
diferencia de progenie verdadera porque es expresado en cualquiera de los dos ambientes.
La interacción GxE causa la reordenación de los individuos (la mayoría padres notables) en
diferentes aéreas geográficas y sistemas de manejo. La extensión del reordenamiento depende
de las especies, el carácter y la cantidad de variación en ambiente y manejo. Como regla, los
caracteres de baja heredabilidad como la fertilidad y la supervivencia son más susceptibles a la
interacción GxE que los caracteres altamente heredables.
Interacción genotipo ambiente (GxE): La relación dependiente entre genotipos y ambientes en
las cuales la diferencia de performance entre dos (o más) genotipos cambia de ambiente en
ambiente
ALTERNATIVAS A LA EVALUACIÓN GENÉTICA A GRAN ESCALA
La evaluación genética a gran escala es común, donde la infraestructura necesaria
(computadoras, asociaciones de cría, experiencia técnica, etc.) existen. Sin embargo, no es el
único enfoque capaz de acelerar el cambio genético. Hay alternativas.
Núcleos de mejoramiento
Los núcleos de mejoramiento son programas de mejoramiento corporativos que involucran a
varios criadores. La información de los núcleos de mejoramiento, los mejores machos y
hembras que son propiedad de criadores cooperativos son combinados en un núcleo.
Usualmente estos animales son reunidos físicamente para formar núcleos de rebaños. Esto es
necesariamente así, si todos los apareamientos son por servicio natural, pero no tiene que ser

Introducción a la Producción Animal - FCV - UNNE
2011
el caso si se usa inseminación artificial. Una vez que el núcleo es formado, todos (o casi todos)
los padres usados en el esquema son producidos en el núcleo donde se llevan registros
meticulosos de pedigrí y performance, las técnicas de predicción son más sofisticadas y se lleva
a cabo la selección mas intensiva.
Un núcleo de mejoramiento simplificado es diagramado en la figura 12.3. En los núcleos de
mejoramiento cerrados el germoplasma fluye en una sola dirección. Los animales
(particularmente los padres) producidos en el núcleo son usados en los rebaños o lotes
cooperadores. En un núcleo de mejoramiento abierto el flujo es bidireccional. Las hembras
que se han probado son superiores en los rebaños o lotes cooperadores son transferidas a el
núcleo.
Muchas de las ventajas de los núcleos de mejoramiento derivan de la ganancia genética
lograda cuando se forma el núcleo de población de elite. Si los animales del núcleo son bien
elegidos, el mérito genético promedio debería ser mucho mayor que el mérito genético
promedio de los rebaños o lote contribuyentes. Los logros adicionales resultan de una
precisión de selección incrementada e (especialmente en los núcleos de mejoramiento
abiertos) intensidad de selección aumentada. Los núcleos de mejoramientos combinados con
ovulación múltiple y transferencia embrionaria (MOET) han sido propuestos como una forma
de mayor incremento de la tasa de cambio genético. El uso de la transferencia embrionaria
permite a las hembras de elite tener mucha progenie, así se incrementa la intensidad y la
precisión de la selección entre las hembras. Si MOET puede ser satisfactoriamente utilizado
con hembras muy jóvenes, entonces el intervalo generacional en hembras puede ser reducido
sustancialmente. La precisión podría sufrir pero la tasa de cambio genético se podría
incrementar significativamente. La efectividad de los esquemas que involucran MOET depende
mucho de la tasa de éxito de la tecnología MOET.
Los núcleos de mejoramiento puede que no produzcan individuos con las precisiones
extremadamente altas que vemos con la evaluación genética a gran escala, pero lo compensan
con la ganancia genética inicial, lograda en la formación de los núcleos y en la ventaja sumada
(dependiendo de su perspectiva) de un grado de disciplina y consistencia impuesta en los
criadores cooperadores. Y no hay razón para que el núcleo de mejoramiento y la evaluación
genética a gran escala no puedan coexistir. Los rebaños o lotes que cooperan con el núcleo de
mejoramiento pueden ser parte de un esfuerzo de evaluación genética mayor.

Introducción a la Producción Animal - FCV - UNNE
2011
Núcleos de mejoramiento: Un programa de mejoramiento cooperativo en el cual los animales
de elite son concentrados en un rebaño o lote núcleo y el germoplasma superior es luego
distribuido entre los rebaños o lotes cooperativos
Servicio natural: Apareamiento natural (al contrario de la inseminación artificial).
Núcleos de mejoramiento cerrados: Un núcleo de mejoramiento en el cual el germoplasma
fluye en una sola dirección – del núcleo a rebaños o lotes cooperativos.
Núcleo de mejoramiento abierto: Un núcleo de mejoramiento en el cual el flujo del
germoplasma es bidireccional – del núcleo a rebaños o lotes cooperativos y de los rebaños
cooperativos o lotes al núcleo.
Ovulación múltiple y transferencia embrionaria (MOET): Ovulación inducida hormonalmente
de múltiples óvulos, seguido por transferencia embrionaria a madres receptoras. El término es
utilizado en conjunción con estrategias diseñadas para incrementar la tasa de cambio genético
usando la transferencia embrionaria