12_konformacioni_prelazi_proteina.pdf
TRANSCRIPT

Konformacioni prelazi: razvijanje i uvijanje proteina • Denaturacija (razvijanje): reversibilna i ireversibilna
– Sredstva za denaturaciju – Metode za praćenje denaturacije/renaturacije – Mehanizmi denaturacije – Određivanje konformacione stabilnosti
• Uvajanje proteina
– Brzina uvijanja proteina – Kooperativna priroda uvijanja-razvijanja proteina – Mehanizmi uvijanja malih i velikih proteina – Uvijanje proteina in vivo – Konformacione bolesti
Literatura: – PSF: pogl. 8 + Dodatak – Voet: pogl. 8 (9) – Lehninger: pogl. 16 – Pr. Ponašanje proteina

Konformaciona stabilnost: G između nativne i razvijene koformacije u vodi (vodenoj sredini/puferu)
Gtotal = Hniz (TSniz + TSvode)
K = [N]/[D]
K>1
RTlnK = Gtotal
Gtotal < 0
Konformaciona entropija (Sniz<0 za uvijanje):
favorizuje razvijanje proteina!!!!!
Entropija vode (Svode>0 za uvijanje):
favorizuje uvijanje proteina!!!!

Zašto se nativni proteini (lako) denaturišu?
• Nativna konformacija je stabilnija od razvijene, ali je razlika mala:
– Konformaciona stabilnost (G) malih globularnih proteina:
-20 do - 40 kJ/mol.
• Promena spoljašnjih uslova (koji menjaju Hniz i Svode) može da izazove razvijanje (denaturaciju) proteina.
Gtotal = Hniz (TSniz + TSvode)
Hniz < 0 Svode> 0

Denaturacija proteina: ireversibilna i reversibilna!

Denaturanti
• Temperatura
• pH: promena jonizacionog stanja kritičnih ostataka
• Jonski ili polarni denaturanti: urea i guanidinium
• Visoke koncentracije organskih supstanci rastvorljivih u vodi
• Detergenti:
– Vezuju se jako za razvijeni oblik

Denaturacija proteina organskim rastvaračima

Detergenti: vezivanje SDS-a za molekul proteina
CH3 – (CH2)11 – OSO3-

Po kojim osobinama se razlikuju nativni i denaturisani protein? (praćenje razvijanja
proteina)
• Biološka aktivnost
• Rastvorljivost
• Kalorimetrija
• Optička aktivnost (ORD, CD)
• UV, fluorescentni, NMR spektri
• Reaktivnost aminokiselinskih ostataka
• Hidrodinamičke veličine (viskoznost, gel filtracija...)

UV spektri Trp, Tyr i Phe
A = a() b c
Na absorptivnost (a()) utiče okolina/rastvarač!

(A) Apsorpcioni spektar nativnog (crno) i denaturisanog (sivo) papaina.
(B) Diferencijalni spektar izmedju nativnog i razvijenog papaina.
(C) Kriva denaturacije papaina.

Optička aktivnost sekundarnih struktura

UV i fluorescentni spektri

Fluorescentni spektar nativne (a) i razvijene (b) ribonukleaze.

Reversibilna denaturacija malih (do 100 ak) proteina

Razvijanje/uvijanje proteina je kooperativan proces
Kriva je sigmoidna.: Proces je kooperativan: nema stabilnih intermedijera. “Sve ili ništa”! U uskom intervalu eksp. uslova nalaze se u ravnoteži nativna i razvijena konformacija proteina.

Određivanje (relativne) konformacione stabilnosti proteina
Tm: temperatura na kojoj je 50% proteina denaturisano!

Efekat soli na stabilnost RNA-ze
• Soli utiču na stabilnost proteina!
• Kalijum fosfat i amonijum sulfat stabilizuju proteine, zato se ove soli često koriste u radu sa proteinima.

Sposobnost jona da stabilizuju protein je u korelaciji sa Hofmeisterovom serijom.

Kako deluju urea i GuHCl?

Problem uvijanja proteina (The Protein Folding Problem)
• Data je određena aminokiselinska sekvenca (primarna struktura) kakva će biti tercijarna/kvaternarna struktura?
• Input: AAVIKYGCAL… Output: 11, 22…

Kako se razvijeni polipeptidni niz uvija u
nativnu konformaciju?
• Pokušajima i greškama isprobavajući sve konformacije dok ne dostigne nativnu najstabilniju konformaciju (najniže G)
• ili
• isprobava samo manji deo konformacija jer aminokiselinska sekvenca određuje i mehanizam (put) uvijanja!

Brzina uvijanja proteina
• Cyrus Levinthal:
• Kada bi protein isprobavao sve konformacije uvijanje proteina od 100 aminokiselina bi trajalo 1050 godina!!!! Levinthalov paradoks!
• Brzina uvijanja in vitro je brz proces: milisekunde - minuti
• Brzina uvijanja in vivo je još veća!

Kako aminokiselinska sekvenca određuje (kodira) 3-D strukturu (uvijanje)
proteina.......
a kako određuje proces (put ) uvijanja?
Isti faktori koji određuju način uvijanja i stabilnost globule određuju i put uvijanja!

Kako se ispituje mehanizam uvijanje proteina?
• Biofizičke tehnike (“stopped flow” tehnike)
– CD u dalekoj UV oblasti (detektovanje sekundarnih struktura)
– CD u bliskoj UV oblasti, fluorescenca (nepolarni ostaci/tercijarne interakcije)
– NMR
• Proteinski inženjering.
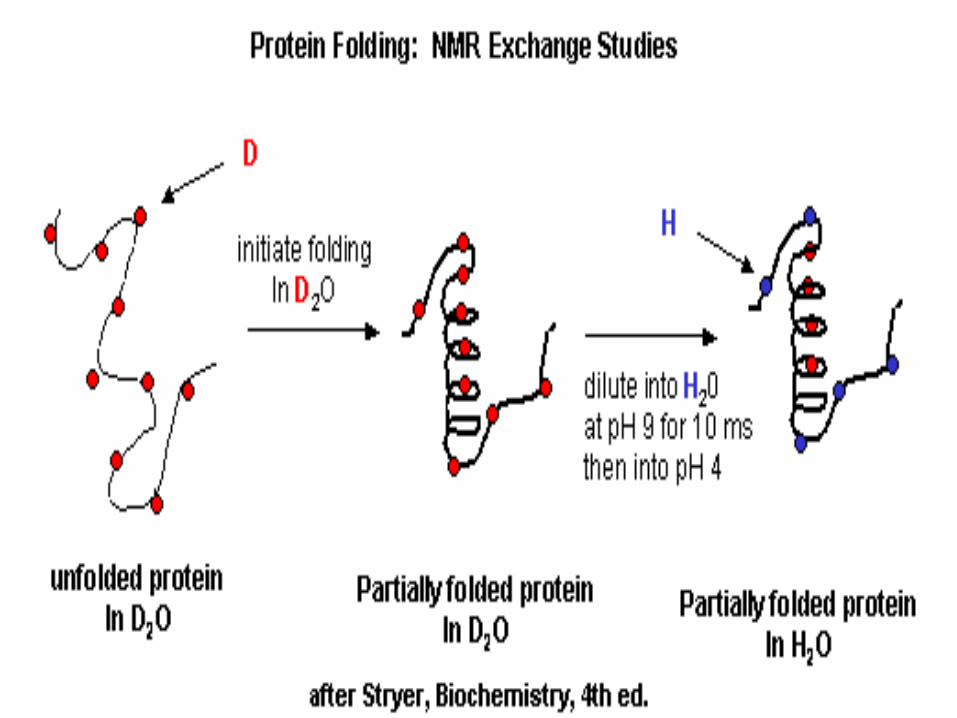


Mehanizam uvijanja malih globularnih proteina:
• brzo, 5 ms, nastajanje sekundarnih
struktura (“burst”)
• “hidrofobni kolaps”: 5 - 1000 ms

Simulacija uvijanja

Uvijanje karbonske anhidraze: intermedijer

Mehanizam uvijanja velikih proteina
• Veliki proteini (>100 ak) imaju i veći sadržaj nepolarnih aminokiselina!
• Mehanizam:
– hidrofobni kolaps u kompaktne intermedijere, “stopljenu globulu” (zapremina 10% veća od nativnog molekula!)
– Zatim, fino podešavanje konformacije !
• Intermedijeri mogu da se nagomilavaju što može dovesti do “pogrešnog uvijanja”

Levci uvijanja:“landscape” teorija uvijanja proteina
Idealizovani levak
Levinthal: ”teren za golf”
Klasični put:
Protein traži “kanjon”
Zgužvana površina sa lokalnim minimumima

a) b)
c)
d)

Uvijanje proteina in vivo
• Živi organizmi mogu da sadrže i do 100 000 različitih proteina.
• Posle sinteze na ribozomima svaki molekul mora da se uvije u nativnu konformaciju određenu njegovom sekvencom.
• U ćeliji se uvijanje dešava u vrlo ispunjenom (“crowded”) okruženju.

Uvijanje proteina
in vivo
30% novo- sintetisanih proteina su DRiP (Defective Ribosomal Products) koji nikada ne mogu da se uviju u nativnu strukturu.

Uvijanje proteina in vivo
U ćeliji postoje:
– pomagači (molekulski “šaperoni”) i
– katalizatori koji ubrzavaju uvijanje:
protein-disulfid izomeraza (PDI)
prolin-izomeraza

Katalizator: protein-disulfid izomeraza

Katalizator: prolin-izomeraza
Peptidil prolil cis-trans izomeraza

Zašto molekulski “šaperoni”?
• Razvijeni proteini in vivo imaju tendenciju da se agregiraju jer se:
– u novo-sintetisanim polipeptidnom nizu nepolarni ostaci nalaze u dodiru sa vodom;
– proteini in vivo uvijaju u prisustvu visoke koncentracije drugih proteina (300 g/L).
• Molekulski šaperoni su proteini koji sprečavaju neproduktivne interakcije delimično uvijenog proteina, posebno one koje dovode do njegovog agregiranja.

Molekulski šaperoni
• “Šaperonini”
• Nukleplazmini:
– neophodni za asocijaciju nukleozoma
• Hsp90:
– uvijanje proteina uključenih u transdukciju signala
• “Heat shock” proteini 70 (Hsp 70):
– sintetizuju se na povišenim temperaturama; visoko konzervirani u prokariotama i eukariotama.
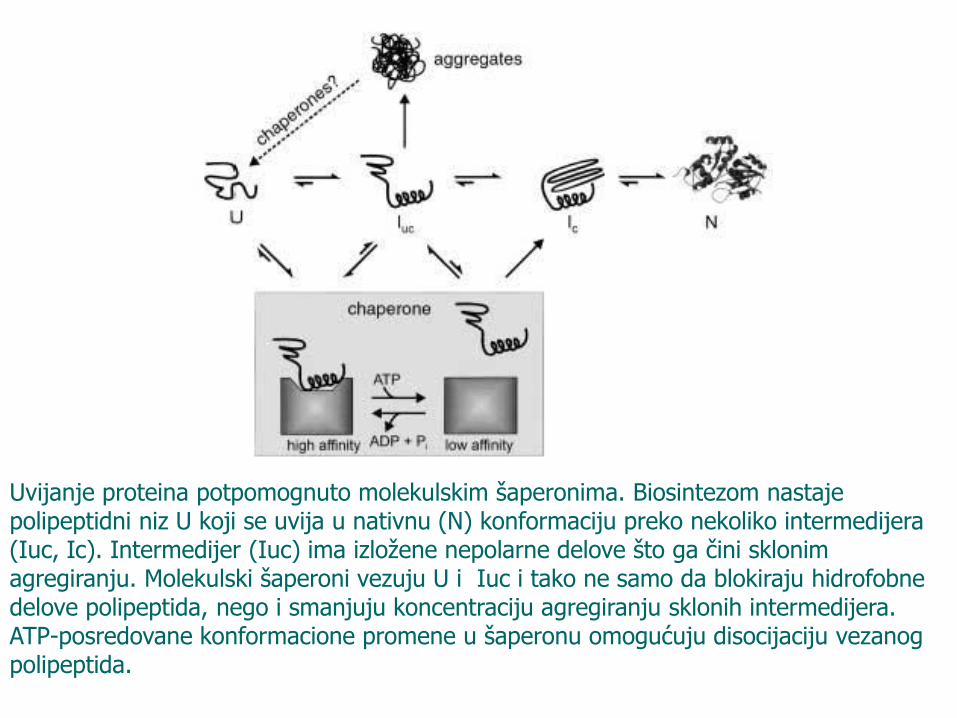
Uvijanje proteina potpomognuto molekulskim šaperonima. Biosintezom nastaje polipeptidni niz U koji se uvija u nativnu (N) konformaciju preko nekoliko intermedijera (Iuc, Ic). Intermedijer (Iuc) ima izložene nepolarne delove što ga čini sklonim agregiranju. Molekulski šaperoni vezuju U i Iuc i tako ne samo da blokiraju hidrofobne delove polipeptida, nego i smanjuju koncentraciju agregiranju sklonih intermedijera. ATP-posredovane konformacione promene u šaperonu omogućuju disocijaciju vezanog polipeptida.

“Šaperonini”
Uvlače delimično/pogrešno uvijeni protein u nepolarnu šupljinu (i pomoću ATP-a) obezbeđuju pravilno uvijanje!

Konformacione bolesti: amiloidne bolesti i prioni
• Proteini zadržavaju nativnu konformaciju ili se, ako
postanu delimično razvijeni, degradiraju!
• Amiloidne bolesti: do sada je otkriveno najmanje 18 bolesti kod kojih se agregiraju inače rastvorni proteini u obliku amiloidnih (sličnih skrobu!) fibrilnih agregata.
• Prion (proteinacceous infections particle that lacks nucleic acid): bolest “ludih krava”


Primeri amiloidnih bolesti

Zašto dolazi do agregiranja proteina: energetska površina za uvijanje i agregaciju proteina

Zajedničko za sve proteine koji grade amiloidna vlakna:

Model amiloidnih vlakana: (a) interakcije nizova (b) model amiloidnog vlakna sačinjenog iz
četiri β pločice: pločice su paralelne sa osom heliksa dok je svaki β niz normalan na osu heliksa.
.

In vitro ispitivanja: svaki protein može da nagradi amiloidna vlakna
SH3 domain
SH3 domain
insulin
A-beta fibril

Mali broj proteina gradi amiloidna vlakna in vivo

Neke mutacije favorizuju nastajanje vlakana
Mutacije koje destabilizuju lizozim dovode do toga da intermedijeri duže opstaju, te da se agregiraju!

Šta pogoduje nastajanju amiloida?
• Smanjenje stabilnosti ili kooperativnosti nativnog proteina zbog mutacija.
• Agregiranje zavisi od koncentracije datog proteina, sposobnosti sekvence za agregiranje i brizine agregiranja.
• Amiloidne strukture mogu da nastanu “pelcovanjem” (kao kod kristalizacije!): kada jednom agregiranje započne ide mnogo brže.
• Gubitak kontrolnih i regulatornih mehanizama za uvijanje proteina (zato se ove bolesti javljaju u starosti!)

Šta sprečava agregiranje proteina in vivo?
• Agregiranje (intermedijera) je minimizirano
– kooperativnom prirodom procesa uvijanja
– prisustvom šaperona
– kompaktnom prirodom globule
• Evolucija:
– sekvenci sa specifičnom sposobnošću da nagrade kooperativno globularne strukture
– kontrolni mehanizmi koji održavaju molekule pod pogodnim uslovima