1-160 biologia12 2015 roman6. tord.qxd:1-80 biologia12 · temei. În aceste alineate găsim foarte...
TRANSCRIPT

Această serie de manuale cuprinde conţinutuldisciplinei de biologie pentru gimnaziu în treivolume.
Pentru o înţelegere cât mai uşoară a textelordin manuale, acestea au fost completate cufiguri. Referirile la figuri sunt prezente în text înformatul prezentat mai jos:
notarea figurilor din manual (Fig. 61)
Textele din manual sunt tipărite cu litere deformat normal. Pentru identificarea şi reţinereacât mai uşoară a noţiunilor importante, acesteasunt tipărite cu litere îngroşate sau cursive.
Alineatele cu litere mărunte marcate la început cuun pătrat verde cuprind informaţii suplimentare aletemei prezentate, şi în aceste părţi vom găsi noţiuniaccentuate cu litere îngroşate sau cursive. Acestemateriale suplimentare completează elementele capi-tolului respectiv cu noi noţiuni sau alte abordări ale
temei respective, uneori prezentând elemente practicelegate de subiect sau completări de natură istorică aletemei. În aceste alineate găsim foarte multe informaţiilegate de menţinerea sănătăţii şi de modul de viaţăsănătos.
Cele trei teme majore ale materiei sunt pre-zentate pe parcursul a şase capitole. La subsolulfiecărei pagini din manual este trecută denumi-rea capitolului, ajutând astfel orientarea înconţinutul materiei. Capitolele sunt divizate însubunităţi mai mici. Aceste unităţi didactice setermină cu întrebări şi exerciţii. Încheierea uni-tăţilor mai mari se face printr-un rezumat, careeste util pentru studiul individual prin faptul căoferă o retrospectivă prescurtată a elementelorprincipale prezentate în capitolul respectiv. Lasfârşitul manualului, principalele noţiuni suntcuprinse într-un index, în care sunt marcate cuasterisc (*) noţiunile care sunt incluse în materiade bacalaureat.
4
INTRODUCERE
1-160 biologia12 2015 roman6. tord.qxd:1-80 biologia12.qxd 3/22/16 10:37 AM Page 4

EREDITATEA ÎN LUMEA VIE
1-160 biologia12 2015 roman6. tord.qxd:1-80 biologia12.qxd 3/22/16 10:37 AM Page 5

6 BAZELE EREDITĂŢII
Legile de bază ale geneticii
Organismele vii au o structură ca şi cum ar fifost proiectate în prealabil. În fond, proprietăți-le lor sunt determinate de reacțiile biochimicedesfăşurate în mod programat în proceselemetabolice care se desfăşoară în organism. Unrol important în acest mecanism îl au substanțe-le catalizatoare ale reacțiilor, proteinele enzi-matice şi diferiții compuşi cu rol plastic şi regu-lator, tot de natură proteică. Structura proteine-lor depinde în primul rând de secvența aminoa-cizilor care intră în componența lor. Aceastăsecvență este determinată de molecula ADNprin diferite canale informaționale, prin careinfluențează sinteza de proteine. Informațiilereferitoare la secvența aminoacizilor la diferiteproteine este stocată în anumite segmente alemoleculei ADN. În final, putem spune că aces-te segmente ale moleculei ADN sunt răspunză-toare în mod direct, prin intermediul sintezeiproteice, de proprietățile pe care le au indiviziiunei specii. Informațiile stocate în moleculaADN se moştenesc de la o generație la alta, prinprocesul de reproducție. Deci, materialul eredi-tar este ADN-ul, mai exact acele unități alemoleculei, care vor determina natura a câte uneitrăsături, respectiv genele. De fapt nu trăsături-le se transmit ereditar, ci acele gene care ledetermină pe acestea. Înfățişarea determinată defuncționarea genelor, totalitatea caracterelorstructurale şi funcționale ale individului poartădenumirea de fenotip. Fundalul genetic caredetermină formarea acestuia, totalitatea genelorunui organism formează genotipul organismu-lui respectiv.
În timpul diviziunii celulare, masa de ADN acelulelor se condensează sub formă de cromo-zomi. În celulele diploide, cromozomii cores-punzători, denumiți omologi, formează perechide cromozomi. În acestea se află câte două bucăți din fiecare genă, așezate peacelași loc pe care erau cromozomul patern șimatern. În natură, o anumită genă poate
avea mai multe variante, pe care le denumimalele. Exemple în acest sens sunt cele trei genealele care determină grupele sanguine ABO laom. În populațiile naturale, numărul alelelorunei gene variază de obicei între unu şi două-zeci, în cazuri mai rare de la unu la mai multesute. În populația respectivă, alela cea mai frec-ventă o denumim alela sălbatică. În celulahaploidă întâlnim numai una dintre perechea degene alele care în celula diploidă apare subformă de pereche. Dacă într-o celulă diploidă,locul destinat unei anumite gene – locus – de peambii cromozomi omologi este ocupat de douăalele identice, individul purtător este homozi-got, iar dacă alelele sunt diferite, individul esteheterozigot pentru gena respectivă. Indiferentde numărul total al alelelor existente pentru ogenă, într-un individ se vor găsi maximum douădintre acestea. Dacă luăm în considerare faptulcă numărul genelor la un individ dintr-o speciede vertebrate poate depăşi mai multe zeci demii, şi într-o populație a speciei respective potsă apară nenumărate alele ale unei gene, esteevident că în natură probabilitatea apariției adoi indivizi cu materialul genetic total identiceste foarte redusă. Astfel, putem spune că fiecare individ deține un material genetic indi-vidual şi unic. Această diversitate imensă agenotipurilor stă la baza varietății fenotipice pe care o putem observa la indivizii diferitelorspecii.
Genetica este disciplina care studiază legitățile ere-dității. Fondatorul acestei ştiințe este considerat călu-gărul, profesorul şi botanistul austriac JohannGregor Mendel (1822 – 1884). Prin încrucişarea adiferite soiuri de plante a obținut hibrizi care dacăerau încrucişați mai departe între ei, generau urmaşiîntre care apăreau indivizi cu trăsături foarte diferite.Deoarece a fost interesat să cunoască exact cauzeleacestor fenomene, în 1854 a început să efectueze oamplă serie de cercetări. Pentru efectuarea acestora aconsiderat că mazărea este ca cea mai potrivită specie.Experiențele le efectua pe un număr mare de indivizi,pentru a înlătura efectele derutante ale fenomenelorîntâmplătoare. A proiectat cu migală fiecare experi-
BAZELE EREDITĂŢII
1-160 biologia12 2015 roman6. tord.qxd:1-80 biologia12.qxd 3/22/16 10:37 AM Page 6

7BAZELE EREDITĂŢII
ment şi a asigurat cu rigurozitate toate condițiile nece-sare.
A făcut însemnări detaliate până şi despre cele maimici amănunte ale experiențelor şi pentru elaborareaconcluziilor a folosit metode matematice şi statistice,încă nefolosite în biologie în acea vreme. Rezultatelemuncii sale de mai mulți ani le-a publicat în 1866, înlucrarea intitulată „Experiențe asupra hibridizăriiplantelor”, în care relatează observațiile efectuate asu-pra a peste zece mii de hibrizi, la care a urmărit evo-luția a şapte trăsături distincte. Astfel, la mazăre a stu-diat culoarea şi forma boabelor, aşezarea florilor şilungimea vrejului. Concluziile formulate în urmarezultatelor experimentale sunt considerate până azilegile de bază ale geneticii. În lipsa cunoştințelor dehistologie la acea vreme, contemporanii au primit cuneînțelegere activitatea lui Mendel, rolul epocal alacesteia în punerea bazelor biologiei moderne a fostrecunoscut numai după moartea savantului, după1900, când mecanismul diviziunii celulare şi structu-ra cromozomilor au devenit cunoscute.
Pentru prezentarea legilor de bază ale eredi-tăţii am ales câteva din experiențele efectuatede Mendel. Una din proprietățile ereditare tipi-ce ale plantelor de mazăre este suprafața netedăsau rugoasă a boabelor (Fig. 1). Să începemanaliza cu plante din generația parentală homo-zigote pentru această proprietate. Pentru identi-ficarea alelelor folosim diferite litere. În situațiade față, perechea de alele care determină supra-fața netedă a boabelor să le notăm cu AA, întimp ce perechea de alele care determină supra-fața rugoasă a boabelor să fie notate cu aa.Desigur, în gameții haploizi materni sau paterniexistă numai una dintre alelele A sau a. Să ana-lizăm posibilitățile de recombinare dintre game-ții parentali cu ajutorul unui tabel (Fig. 2)!Cuvântul „parental” provine din latinescul„parentes” care înseamnă părinte, din carecauză de multe ori în tabelele genetice generațiaparentală este notată cu litera P. În mod asemă-nător, simbolul pentru notarea primelor douăgenerații descendente F1 şi F2 provine dincuvântul latinesc filiales care înseamnă urmaş.După încrucişarea gameților proveniți de la ceidoi părinți homozigoți, la fiecare individ dingenerația F1 se regăsesc alelele care determinăatât proprietățile materne, cât şi cele paterne.Deci, din punctul de vedere al genotipului,generația F1 este heterozigotă, având compo-nența perechii de alele Aa, iar din punctul de
Fig. 1 Proprietățile boabelor de mazăre
neted AA
rugos aa
galben BB
verde bb
Fig. 2 Legea uniformității hibrizilor în prima generație
P
AA aa
F1
A A
a
a
Aa Aa
Aa Aa
Genotipul la F1: 4 AaFenotipul la F1: 4 neted
1-160 biologia12 2015 roman6. tord.qxd:1-80 biologia12.qxd 3/22/16 10:37 AM Page 7
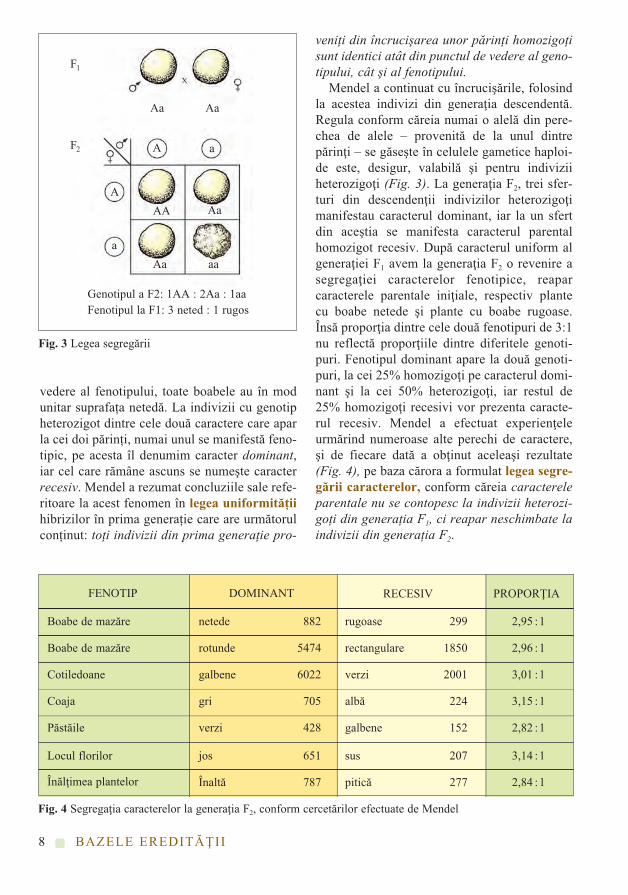
vedere al fenotipului, toate boabele au în modunitar suprafața netedă. La indivizii cu genotipheterozigot dintre cele două caractere care aparla cei doi părinți, numai unul se manifestă feno-tipic, pe acesta îl denumim caracter dominant,iar cel care rămâne ascuns se numeşte caracterrecesiv. Mendel a rezumat concluziile sale refe-ritoare la acest fenomen în legea uniformităţiihibrizilor în prima generație care are următorulconținut: toţi indivizii din prima generaţie pro-
veniţi din încrucişarea unor părinţi homozigoţisunt identici atât din punctul de vedere al geno-tipului, cât şi al fenotipului.
Mendel a continuat cu încrucişările, folosindla acestea indivizi din generaţia descendentă.Regula conform căreia numai o alelă din pere-chea de alele – provenită de la unul dintrepărinţi – se găseşte în celulele gametice haploi-de este, desigur, valabilă şi pentru indiviziiheterozigoţi (Fig. 3). La generaţia F2, trei sfer-turi din descendenţii indivizilor heterozigoţimanifestau caracterul dominant, iar la un sfertdin aceştia se manifesta caracterul parentalhomozigot recesiv. După caracterul uniform algeneraţiei F1 avem la generaţia F2 o revenire asegregaţiei caracterelor fenotipice, reaparcaracterele parentale iniţiale, respectiv plantecu boabe netede şi plante cu boabe rugoase.Însă proporţia dintre cele două fenotipuri de 3:1nu reflectă proporţiile dintre diferitele genoti-puri. Fenotipul dominant apare la două genoti-puri, la cei 25% homozigoţi pe caracterul domi-nant şi la cei 50% heterozigoţi, iar restul de25% homozigoţi recesivi vor prezenta caracte-rul recesiv. Mendel a efectuat experienţeleurmărind numeroase alte perechi de caractere,şi de fiecare dată a obţinut aceleaşi rezultate(Fig. 4), pe baza cărora a formulat legea segre-gării caracterelor, conform căreia caractereleparentale nu se contopesc la indivizii heterozi-goţi din generaţia F1, ci reapar neschimbate laindivizii din generaţia F2.
8 BAZELE EREDITĂŢII
Fig. 3 Legea segregării
F1
Aa Aa
F2 a
a
A
A
Genotipul a F2: 1AA : 2Aa : 1aaFenotipul la F1: 3 neted : 1 rugos
AA Aa
Aa aa
Fig. 4 Segregaţia caracterelor la generaţia F2, conform cercetărilor efectuate de Mendel
FENOTIP DOMINANT RECESIV PROPORŢIA
Boabe de mazăre
Boabe de mazăre
Cotiledoane
Coaja
Păstăile
Locul florilor
Înălţimea plantelor
netede 882
rotunde 5474
galbene 6022
gri 705
verzi 428
jos 651
Înaltă 787
rugoase 299
rectangulare 1850
verzi 2001
albă 224
galbene 152
sus 207
pitică 277
2,95 :1
2,96 :1
3,01 :1
3,15 :1
2,82 :1
3,14 :1
2,84 :1
1-160 biologia12 2015 roman6. tord.qxd:1-80 biologia12.qxd 3/22/16 10:37 AM Page 8

9
Prin cercetările efectuate de Mendel s-au elu-cidat relaţiile dintre transmiterea ereditară adiferitor caractere. La început a studiat alelelecâte unui caracter, apoi a efectuat încrucişăriîntre părinţi ce se deosebeau prin două perechide caractere. La una din aceste experienţe, pelângă caracterul neted sau rugos al boabelor, astudiat şi transmiterea culorii galbene sau verzi acotiledoanelor, iar evaluarea rezultatelor le-afăcut prin metode matematice-statistice. Dintrepărinţii homozigoţi utilizaţi în experienţă, indi-vizii paterni dominanţi aveau alele AA pentruboabe netede şi BB pentru cotiledoane galbene. Indivizii materni recesivi se caracterizau prinalelele aa pentru boabe rugoase şi bb pentrucotiledoane verzi (Fig. 5).
După cum era de aşteptat, toţi indivizii gene-raţiei F1 au fost identici. Fenotipul lor a fostdeterminat de caracterele dominante, deci fieca-re aveau boabe netede cu cotiledoane galbene.
În urma încrucişării indivizilor heterozigoţiîntre ei, în generaţia F2 s-au obţinut nouă feluride genotipuri diferite şi patru fenotipuri. Dupăstudierea genotipurilor se poate constata că pro-porţiile de transmitere ale ambelor caractereluate în parte coincid cu cele constatate la trans-miterea caracterelor determinate de o pereche dealele. Deci, dacă simplificăm rezultatul obţinutde (4AA : 8Aa : 4aa), vom obţine formula dejacunoscută a raporturilor dintre genotipuri de(1AA : 2Aa : 1aa). Dacă împărţim tabelul gene-tic cu datele de mai sus cu o linie transversală înpatru grupe, vom observa că în fiecare grupărepartiţia fenotipurilor este de 3:1. Dacă anali-zăm concomitent toate variantele fenotipice pro-duse în raport de 9:3:3:1, vom observa că apardescendenţi la care sunt alăturate perechi decaractere care nu se găseau alăturate pe cromo-zomii indivizilor din generaţia parentală.
Apariţia perechilor de caractere rugos-galbenşi neted-verde l-a condus pe Mendel la conclu-zia că în timpul încrucişărilor, cele patru aleledin gameţii parentali care determină cele douăcaractere se recombină total independent şi laîntâmplare, dând astfel naştere la combinaţii decaractere complet noi. Pe baza rezultatelor anenumărate asemenea experienţe, Mendelenunţă legea segregării independente a pere-chilor de caractere, care de fapt stabileşte că în
BAZELE EREDITĂŢII
AB AB
ab
ab
AaBb AaBb
AaBb AaBb
AABB aabb
neted, galben x rugos, verde
x
AaBb AaBb
x
AB Ab aB ab
AB
Ab
aB
ab
AABB AABb AaBB AaBb
AABb AAbb AaBb Aabb
AaBB AaBb aaBB aaBb
AaBb Aabb aaBb aabb
P
F1
F1
F2
Fig. 5 Legea segregării independente a perechilor decaractere
1-160 biologia12 2015 roman6. tord.qxd:1-80 biologia12.qxd 3/22/16 10:37 AM Page 9

urma transmiterii ereditare independente a dife-ritelor perechi de caractere, la generaţia F2 potsă apară şi combinaţii care diferă de formele ceapar la generaţia parentală. Acest fenomen semanifestă numai la încrucişări la care participăpărinţi ce se deosebesc prin mai mult de o pere-che de caractere. Mendel a întreprins cercetăriprivitoare la transmiterea ereditară a trei perechide caractere, unde a obţinut 27 genotipuri diferi-te. În cazul perechilor de alele ce se recombinăindependent, la generaţia F2 în urma recombină-rii unei perechi de caractere pot să apară 3, pen-tru două perechi de caractere 9, pentru treiperechi de caractere 27, iar pentru n perechi decaractere apar 3n genotipuri diferite.
Câteva exemple de transmitere ereditară
Experienţele de încrucişări efectuate la începu-tul secolului XX ca urmare a activităţii luiMendel au dovedit fără echivoc că legile de bazăale geneticii sunt general valabile. La început aufost reluate experienţele mendeliene, apoi expe-rienţele cu diferite specii de plante şi animale s-au extins tot mai mult. Un exemplu relevant aleredităţii dominante-recesive la animale estecel al taurinelor având culoarea părului bălţatăcu negru sau bălţată cu roşu (Fig. 6). S-ademonstrat că transmiterea acestui caracter seface monogenic, ca şi la încrucişările soiurilorde mazăre. Alela dominantă a heterozigoţilor(V) determină culoarea bălţată cu negru, iaralela recesivă (v) este gena care determinăculoarea bălţată cu roşu. Generaţia F1, pe bazalegii uniformităţii, va fi formată numai din indi-vizi cu genotipul Vv şi fenotipul bălţat cu negru.În generaţia F2, în urma segregării alelelor aparindivizi dominanţi – bălţat cu negru – şi recesivi– bălţaţi cu roşu – în raport de 3:1.
10 BAZELE EREDITĂŢII
1. Explicaţi pe baza celor învăţate anul trecut, careeste legătura dintre ordinea bazelor azotate dinmolecula de ADN şi caracterele fenotipice! 2. Ce este gena şi ce este alela? Care este diferenţa
dintre ele? 3. Ce înseamnă noţiunile de genotip şi fenotip?4. Stabiliţi fenotipul bobului de mazăre din Fig. 1
şi menţionaţi ce genotip posibil poate avea!5. Documentaţi-vă şi ţineţi o scurtă prezentare
despre viaţa şi activitatea lui Johann GregorMendel!6. Stabiliţi cu ajutorul figurii 4, ce fenotip a avut
planta homozigotă recesivă studiată de Mendel dinpunctul de vedere al caracterului urmărit?7. Utilizând Fig. 5, explicați cele trei legi ale ere-
dităţii enunţate de Mendel!8. De ce a fost nevoie de urmărirea a două perechi
de caractere pentru demonstrarea legii segregăriiindependente? 9. Ce descendenţi se obţin din încrucişarea unor
plante de mazăre homozigote înalte cu plantehomozigote pitice? (Folosiţi Fig. 4!)10. Ce descendenţi se obţin din încrucişarea a douăplante de mazăre heterozigote cu florile plasate înjos? (Folosiţi Fig. 4!)11. Ce descendenţi se obţin din încrucişarea a douăplante de mazăre heterozigote pentru ambele pro-prietăţi, cu vrejul înalt şi florile plasate în jos?(Folosiţi Fig. 4!)12. Cum şi prin ce încrucişări am putea stabili des-pre o fiinţă cu fenotip dominant dacă are genotiphomozigot sau heterozigot? Ce fel de fenotip trebu-ie să apară la descendent, ca să ne asigure că a fostheterozigot? Fig. 6 Transmiterea ereditară a culorilor bălţate cu
negru şi cu roşu la taurine
P
F1
F2
vvVV
Vv
V v
V VV Vv
Vv vvV
×
1-160 biologia12 2015 roman6. tord.qxd:1-80 biologia12.qxd 3/22/16 10:37 AM Page 10

Numeroase caractere şi câteva simptome aleunor boli ale omului sunt transmise tot prin ere-ditate dominantă –recesivă. O boală cunoscutăeste albinismul, în care conţinutul de pigment alpielii, părului, ochilor este aproape zero, deci laindividul afectat apare o lipsă patologică de pig-ment. În urma studierii unor arbori genealogici,s-a ajuns la concluzia că albinismul este uncaracter recesiv al omului, indivizii albinoşi augenotip homozigot aa. În medie, la 25 – 30.000copii se naşte un albino. Deci, din punctul devedere al pigmentării, majoritatea covârşitoare aoamenilor sunt homozigoţi dominanţi AA, şinumai câţiva sunt heterozigoţi dominanţi, caredeţin alelele Aa. Din punctul de vedere al pig-mentării, copiii născuţi din căsnicia unei persoa-ne homozigote dominante cu una recesivă albi-no vor fi pigmentaţi normal (Fig. 7a).Probabilitatea naşterii unor descendenţi albinodin căsătoria unor persoane heterozigote este de25% (Fig. 7b), iar la căsătoria dintre o persoanăheterozigotă şi una recesivă albino, probabilita-tea creşte la 50%. De la sine se înţelege că dincăsătoria a doi părinţi albino recesivi, toţi copiivor fi albino.
În unele situaţii, dezvoltarea fenotipică a ale-lelor dominante se produce mai puţin accentuatla heterozigoţi decât la homozigoţi. Fenotipulheterozigot este intermediar între cele douăfenotipuri homozigote. Aceasta este transmite-
rea ereditară intermediară. O plantă binecunoscută şi din grădinile din Ungaria sunt flo-rile de barba împăratului din familiaNyctaginaceae (Fig. 8). La indivizii cu flori dediferite culori ale acestei specii, transmiterea
11BAZELE EREDITĂŢII
Fig. 7b Copil albino
Fig. 7a Transmiterea ereditară a albinismului la om
AA aa Aa Aa AA aa
Aa aa
Aa aa
AA Aa
Aa aa
Aa Aa
Aa Aa
A A A a A a
a
a
A
a
a
a
0%fenotip albino 25% 50%
1-160 biologia12 2015 roman6. tord.qxd:1-80 biologia12.qxd 3/22/16 10:38 AM Page 11

ereditară a culorii florilor este determinată de opereche de alele. Dacă se încrucişează plantehomozigote cu flori roşii şi albe, în F1 toţi augenotipul şi fenotipul identic. Însă, spre deosebi-re de cazurile pe care le-am descris până acum,fenotipul dominant la heterozigoţi se manifestămai puţin intens, astfel că descendenţii în loc săaibă flori roşii, vor avea flori roz. În urmaîncrucişării a doi heterozigoţi din aceştia varezulta generaţia F2, segregarea dintre fenotipuldominant şi recesiv se va face în proporţie de 1 : 2 : 1, din care cei doi heterozigoţi vor pre-zenta un fenotip dominant incomplet, având flo-rile roz în loc de roşu (Fig. 8b).
O variantă a transmiterii ereditare dominante-recesive o reprezintă fenomenul de codomi-nanţă. Un exemplu foarte cunoscut pentru acestfenomen este transmiterea ereditară a grupelorsanguine umane ABO. Formarea grupelor san-guine ABO este determinată genetic de trei genealele, dintre care două, IA şi IB sunt dominantefaţă de gena recesivă i. Este cunoscut faptul că laorganismele diploide pe un anumit locus se potafla numai două gene alele, iar relaţia dintreacestea va determina grupa sanguină a descen-denţilor (Fig. 9). Genotipul indivizilor cu grupasanguină A- şi B- avea în componenţă aleledominante. Grupa sanguină A- apare la indiviziicu genotipul homozigot IAIA sau heterozigot IAi,iar grupa sanguină B- este determinată de pere-chea de alele IBIB homozigote sau IBi heterozigo-te. Grupa sanguină O este determinată geneticde perechea homozigotă de alele recesive ii.Alelele IA şi IB sunt dominante în măsură egalăfaţă de alela recesivă i. În cazul prezenţei lorconcomitente sub forma perechii de alele cucomponenţa IAIB, apare fenomenul de codomi-nanţă, efectul ambelor alele se manifestă, iarurmaşii cu acest genotip vor avea grupa sangui-nă AB.
Până acum am analizat exemple în care mani-festarea unui caracter era determinată genetic deo pereche de alele. Însă în numeroase cazuri,manifestarea unui caracter este determinatăde mai multe perechi de alele. Acestea, de fapt,se influenţează reciproc, fără să acţioneze directasupra genelor, ci indirect, prin intermediuluiproteinelor produse de gene. Acestea pot să inte-racţioneze în cadrul proceselor metabolice celu-
12 BAZELE EREDITĂŢII
Fig. 8a Flori roşii, albe şi roz de barba împăratului
Fig. 8b Transmiterea ereditară intermediară
F2
F
F
P
P
PF FF
PP PF
PF PF
PF PF
P P
F
F
F1
P
PP FF×
1-160 biologia12 2015 roman6. tord.qxd:1-80 biologia12.qxd 3/22/16 10:38 AM Page 12

13BAZELE EREDITĂŢII
lare şi determină modificarea raportului desegregare a fenotipurilor, fără să contravină legi-lor de bază ale geneticii. Prin acest mecanism setransmite ereditar forma crestei la încrucişăriledintre diferite rase de găini (Fig. 10). Formacrestei la găini este determinată de două perechide alele. În urma încrucişării unei rase de găini
cu creasta în formă de trandafir cu una avândcreasta tip mazăre, în F1 apare un nou fenotip,creasta tip nucă. Acest fenotip apare numai dacăambele gene alele dominante R şi B sunt pre-zente concomitent la un individ. În urmaîncrucişării heterozigoţilor F1 cu creasta tipnucă, în generaţia F2 segregarea fenotipurilor ar
Fig. 9 Transmiterea ereditară a grupelor sanguine AB0
Fig. 10 Transmiterea ereditară a formei crestei la rasele de găini domestice
creastă trandafir creastă tip mazăre creastă tip nucă creastă simplă
RR RR Rr RrBB Bb BB BbRR RR Rr RrBb bb Bb bbRr Rr rr rrBB Bb BB BbRr Rr rr rrBb bb Bb bb
RB
RB
Rb rB rb
Rb
rB
rb
RbRb
rB
rB
Rr Bb Rr Bb
Rr Bb Rr Bb
P: RR bb × rr BBcreastă trandafir creastă tip mazăre
F1: Rr Bb × Rr Bbcreastă tip nucă creastă tip nucă
A, BA, B, AB, 0fenotipul F1 AB
A × B A × B AB × 0
IB
IB
IA IAIA IA IB
IB
i
i
ii iIA i IA i IB i
IA i IB iIB iIA IB
IA IB IA IB
IA IBIA IB
i
fenotipul P
1-160 biologia12 2015 roman6. tord.qxd:1-80 biologia12.qxd 3/22/16 10:38 AM Page 13
14
trebui să se producă în proporţiile prevăzute delegile mendeliene, adică 9 : 3 : 3 : 1. Cu toateacestea, rezultatul este altul, pentru că în afarăde fenotipurile prenatale – creastă tip trandafir şimazăre – la descendenţi au apărut două fenoti-puri noi, cel dublu dominant – creastă în formăde nucă – şi cel dublu recesiv – creastă simplă,care nu existau la părinţi. Fenotipurile noi auapărut în ontogeneza anumitor indivizi ca rezul-tat al acţiunii reciproce a proteinelor produse decele două tipuri de gene.
Există şi forme de ereditate care sunt tocmaiopusul modelului de transmitere prezentat ante-rior, la care mai multe caractere sunt determi-nate de o singură pereche de alele. Exemplu înacest sens reprezintă una din genele plantelor demazăre, care determină atât culoarea florilor, câtşi culoarea cojii seminţelor. Acest lucru înseam-
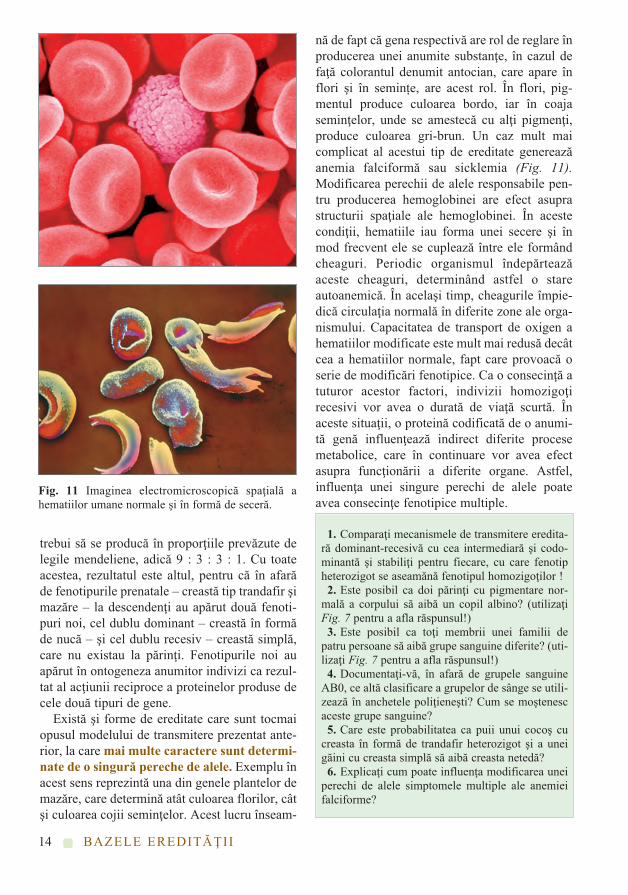
nă de fapt că gena respectivă are rol de reglare înproducerea unei anumite substanţe, în cazul defaţă colorantul denumit antocian, care apare înflori şi în semințe, are acest rol. În flori, pig-mentul produce culoarea bordo, iar în coajaseminţelor, unde se amestecă cu alţi pigmenţi,produce culoarea gri-brun. Un caz mult maicomplicat al acestui tip de ereditate genereazăanemia falciformă sau sicklemia (Fig. 11).Modificarea perechii de alele responsabile pen-tru producerea hemoglobinei are efect asuprastructurii spaţiale ale hemoglobinei. În acestecondiţii, hematiile iau forma unei secere şi înmod frecvent ele se cuplează între ele formândcheaguri. Periodic organismul îndepărteazăaceste cheaguri, determinând astfel o stareautoanemică. În acelaşi timp, cheagurile împie-dică circulaţia normală în diferite zone ale orga-nismului. Capacitatea de transport de oxigen ahematiilor modificate este mult mai redusă decâtcea a hematiilor normale, fapt care provoacă oserie de modificări fenotipice. Ca o consecinţă atuturor acestor factori, indivizii homozigoţirecesivi vor avea o durată de viaţă scurtă. Înaceste situaţii, o proteină codificată de o anumi-tă genă influenţează indirect diferite procesemetabolice, care în continuare vor avea efectasupra funcţionării a diferite organe. Astfel,influenţa unei singure perechi de alele poateavea consecinţe fenotipice multiple.
BAZELE EREDITĂŢII
Fig. 11 Imaginea electromicroscopică spaţială ahematiilor umane normale şi în formă de seceră.
1. Comparaţi mecanismele de transmitere eredita-ră dominant-recesivă cu cea intermediară şi codo-minantă şi stabiliţi pentru fiecare, cu care fenotipheterozigot se aseamănă fenotipul homozigoţilor !2. Este posibil ca doi părinţi cu pigmentare nor-
mală a corpului să aibă un copil albino? (utilizaţiFig. 7 pentru a afla răspunsul!)3. Este posibil ca toţi membrii unei familii de
patru persoane să aibă grupe sanguine diferite? (uti-lizaţi Fig. 7 pentru a afla răspunsul!)4. Documentaţi-vă, în afară de grupele sanguine
AB0, ce altă clasificare a grupelor de sânge se utili-zează în anchetele poliţieneşti? Cum se moştenescaceste grupe sanguine?5. Care este probabilitatea ca puii unui cocoş cu
creasta în formă de trandafir heterozigot şi a uneigăini cu creasta simplă să aibă creasta netedă? 6. Explicaţi cum poate influenţa modificarea unei
perechi de alele simptomele multiple ale anemieifalciforme?
1-160 biologia12 2015 roman6. tord.qxd:1-80 biologia12.qxd 3/22/16 10:38 AM Page 14

15
Ereditatea legată de sex
În momentul fecundării, seturile haploide decromozomi ale gameţilor sunt unite sub formădiploidă în zigot. În zigot, cromozomii seturilorhaploide formează perechi omoloage. Un cro-mozom din perechea de omologi provine de latată, celălalt de la mamă. La plantele dioice şi laanimalele monosexuate, ca şi la om, una dinperechile de cromozomi omologi are rol majorîn determinismul sexelor. Aceştia se numescheterocromozomi sau heterozomi. La perecheade heterozomi, unul din sexe participă cu doicromozomi simetrici, în timp ce la celălalt sex,heterozomii sunt diferiţi (Fig.12). La om şi lamajoritatea animalelor monosexuate şi plantedioice, perechea heterozomală asimetrică XYapare mai ales la masculi, în timp ce la femelesunt caracteristice perechile heterozomale sime-trice de tip XX. La păsări, fluturi şi câteva spe-cii de plante apare determinismul sexual geneticinvers, la acestea femelele poartă perechea deheterozomi asimetrici WZ, iar la masculi apareperechea heterozomală simetrică ZZ. Deoareceîn timpul fecundaţiei întâlnirea celor două tipuride heterozomi au aceeaşi probabilitate, repartiţiapotenţială a sexelor la descendenţi este în mediede 50-50%. Deci, caracterele sexuale sunt pro-prietăţi ce se transmit prin heterozomi, raportulde apariţie al acestora la nivel de generaţie nuvariază prea mult.
Din perechea de heterozomi XX de la femelelemamiferelor diploide, numai una este funcţională.Cealaltă devine inactivă încă în stadiile incipiente aledezvoltării embrionare, deoarece în molecula ei deADN survin modificări care inhibă replicarea infor-maţiei. La om, heterozomul X inactiv rămâne intenscondensat şi după diviziunea celulară, astfel încâtpoate fi vizualizat şi observat chiar cu un microscopobişnuit în nucleul celulelor feminine sub forma uneimici formaţiuni intens colorate (testul Barr). Aceastătehnică este folosită ca metodă cromozomală pentrudeterminarea sexului, de exemplu la stabilirea sexuluila sportive în marile competiţii internaţionale.Introducerea testului a fost necesară deoarece existătulburări ce apar în etapele timpurii ale ontogenezeiembrionului mascul, în care producţia de hormonimasculini este inhibată. Ca urmare, organele genitaleexterne ale descendentului şi constituţia corporală aacestuia vor purta caracterele feminine, în timp ce
gonadele acestuia vor fi masculine – testicule -, la felşi structura heterozomală XY. Pentru examinarea cro-mozomală a sexelor – testul Barr-Bertram – se potfolosi fie celule ale epiteliului bucal, fie celule dinrădăcina părului, sau chiar limfocitele dintr-un frotiusanguin.
La sfârşitul verii, în locurile unde se găsescfructe răscoapte, în fermentaţie, apar şi roiuri demusculiţe. Acestea sunt musculiţele de oţet, caresunt de fapt unele din cele mai studiate animaledin punct de vedere genetic. În urma expe-rienţelor efectuate pe aceste insecte s-a observatcă transmiterea anumitor caractere are legăturăcu heterozomii. Culoarea ochilor de la mus-culiţele de oţet la tipul sălbatic este roşie. Acestcaracter este dominant faţă de alelele care deter-mină celelalte culori ale ochilor. Între musculiţeapar foarte rar şi indivizi cu ochii albi. Tocmaiaceste exemplare au fost folosite pentru studie-rea transmiterii ereditare a culorii ochilor (Fig.13). Un mascul cu ochii albi a fost încrucişat larând cu mai multe femele homozigote cu ochiiroşii. Conform legilor fundamentale ale geneti-cii, generaţia F1 a fost uniformă cu ochii roşii.Segregarea caracterelor în F2 s-a produs tot con-form acestor legi 3 : 1. După studierea amă-nunţită a unui grup de peste patru mii de indivizidescendenţi, s-a remarcat un fenomen particular.Toate musculiţele cu ochii albi erau de sex mas-culin. Studiind indivizi cu ochii roşii din F2, s-aconstatat că distribuţia între sexe, femele şi mas-culi este de 2 : 1 . De aici s-a tras concluzia căexistă o legătură între transmiterea culorii ochi-lor şi heterozomi.
Apoi s-au efectuat noi experienţe unde carac-terele parentale au fost inversate. Astfel, masculi
BAZELE EREDITĂŢII
Fig. 12 Transmiterea caracterelor sexuale
X
X
X Y
XX XY WZ WZ
XX XY ZZ ZZ
Z Z
W
Z
1-160 biologia12 2015 roman6. tord.qxd:1-80 biologia12.qxd 3/22/16 10:38 AM Page 15

16
cu ochii roşii au fost încrucişaţi cu femele homo-zigote cu ochii albi. În această situaţie, deja în F1
se constată o segregare asemănătoare cu cea dela transmiterea caracterelor sexuale. O parte adescendenţilor au ochii albi, cealaltă parte ochiiroşii. Toţi indivizii cu ochii albi erau masculi, iarcei cu ochii roşii erau femele. Deci, caracterulpatern a fost moştenit de descendenţii femeli, iarcaracterul matern de către indivizii masculi. Înurma încrucişării indivizilor F1, la generaţia F2
atât la indivizii descendenţi cu ochii roşii, cât şi la cei cu ochii albi, distribuţia pe sexe era de50 : 50.
Toate aceste rezultate se pot explica numaiprin faptul că gena care determină culoareaochilor la musculiţa de oţet este legată de cro-mozomul X. După aceste concluzii, numai lamusculiţa de oţet s-au descoperit mai multe sutede gene, care se leagă de cromozomii sexuali. Şila om se cunosc peste o sută de gene de naturăheterozomală. Există numeroase caractere acăror transmitere ereditară este legată desexul organismului.
Introducerea în genetică a experienţelor cu efecti-ve mari de animale şi folosirea musculiţei de oţet
(Drosophyla melanogaster) la experienţe este legatăde activitatea geneticianului american ThomasMorgan (1866 – 1945).
Această specie de insecte are mai multe trăsăturiavantajoase din punctul de vedere al efectuării expe-rienţelor. Este o specie nepretenţioasă, se poate ţine încondiţii simple şi se înmulţeşte rapid, iar din ouălefecundate în 10-12 zile se dezvoltă indivizi ajunşi lamaturitate sexuală, astfel că în timp scurt se pot exa-mina serii întregi de generaţii. Are în total patruperechi de cromozomi, care din cauza dimensiunilorlor relativ mari pot fi ţinuţi sub observaţie uşor. Esteuna din speciile frecvent utilizate în experienţelegeneticii aplicate până în ziua de azi.
Utilizând experienţe genetice efectuate sistema-tic, Morgan a demonstrat că la musculiţa de oţet celemai mărunte detalii corporale au legătură cu structura cromozomilor. În lucrarea sa intitulatăTeoria cromozomială a eredităţii, publicată în 1926,dezvoltă ideea că fiecărui caracter ereditar al unuiindivid îi corespunde o particulă materială: gena afla-tă pe unul din cromozomii proveniţi de la părinţi.Deoarece unul din cromozomii pereche provine de lamamă, iar celălalt de la tată, fiecare caracter ereditartrebuie să aparţină unei perechi de gene. Pentru stu-diul cromozomilor ca purtători ai informaţiei eredita-re, lui Thomas Morgan i s-a decernat premiul Nobelîn anul 1933.
O altă trăsătură a cărei transmitere este legată
BAZELE EREDITĂŢII
Fig. 13 Transmiterea genelor heterozomale
Xf Y XP XP
XP Y Xf Xf
Xf Y XP Y
XP XP XP YXP Xf XP Y
XP Xf Xf YXP Xf XP Y
XP Xf Xf Y
XP Xf Xf Y Xf Xf Xf Y
XP Xf XP YXP
Xf Xf
Xf
XPXP
XP
F1
F1
F2
F2XP Y Xf Y
XP
1-160 biologia12 2015 roman6. tord.qxd:1-80 biologia12.qxd 3/22/16 10:38 AM Page 16

17
de apartenenţa sexuală este moştenirea maladieidenumită hemofilie care afectează capacitateade coagulare a sângelui. Această boală este con-tactată în primul rând de băieţi. Deoarece sânge-le celor afectaţi nu se coagulează în prezenţaaerului, plăgi relativ mici pot produce hemoragiiînsemnate. Părinţii băieţilor cu hemofilie aufenotipul sănătos, dar gena recesivă responsabi-lă de producerea bolii este purtată de mamă, pecromozomul sexual, şi o transmite la descen-denţii băieţi (Fig. 14). Dacă băiatul afectat ajun-ge la maturitate şi se căsătoreşte cu o femeiecare nu este purtătoare a acestei gene a hemofi-liei, băieţii lor nu vor fi hemofilici, deoarece aumoştenit cromozomul X de la mamă. În schimbfetele, care au moştenit cromozomul X al tatălui,chiar având fenotipul sănătos, vor fi purtătoareale maladiei.
Transmiterea înlănţuită a genelor şi recombinarea
Prima dovadă a faptului că fiecare genă are unloc bine stabilit pe cromozom s-a obţinut în urmastudierii eredităţii legate de sex. Deoarece pe cro-mozomii indivizilor eucarioţi se evaluează a fimai multe zeci de mii de gene, este puţin proba-bil ca segregarea fiecărei gene să se producă înmod independent. În experienţele efectuate pemusculiţele de oţet, Thomas Morgan a observatcă numeroase caractere se transmit în mod aso-ciat. Un exemplu îl constituie perechea de carac-tere formată din forma aripilor şi forma abdome-nului. La indivizi de tipul sălbatic s-au notat cu Scei cu aripi normale şi cu P cei cu abdomen demărime normală, acestea fiind caracterele domi-nante. S-au identificat însă şi câţiva indiviziavând proprietăţile recesive de aripi rudimentares şi abdomen îngust p . Indivizii din cele douăgrupe – tip sălbatic, dominant şi cei cu proprie-tăţi recesive – au fost încrucişaţi, apoi s-au stu-diat descendenţii F1. Conform aşteptărilor, ladescendenţi s-au manifestat proprietăţile domi-nante. În urma încrucişărilor următoare dintredescendenţi, aşteptările erau ca în F2 cele douăperechi de caractere să producă segregarea inde-pendentă şi să se producă patru fenotipuri în pro-porţii de 9 : 3 : 3 :1 . Însă în experiment, în F2 aureapărut caracterele parentale în proporţie de 3 :1 , deci 75% din indivizi aveau aripi şi abdomennormal, iar 25% din descendenţi aveau aripirudimentare şi abdomenul îngust (Fig. 15). Acestlucru este posibil numai dacă cele două perechide alele se transmit înlănţuit, asociate între ele laurmaşi. Acest lucru însă presupune ca şi pe cro-mozom să fie plasate alăturat ca gene înlănţuite.Numeroase cercetări ulterioare au confirmat con-cluziile lui Morgan referitoare la faptul că anu-mite caractere care se transmit înlănţuit suntdeterminate de anumite segmente cromozomalecare sunt poziţionate apropiat. Astfel, la organis-mele vii, pe lângă caracterele ce se transmit inde-pendent, se observă anumite trăsături care setransmit înlănţuit şi sunt controlate de geneînlănţuite.
Fenomenul înlănţuirii îl întâlnim sistematic laselectarea plantelor de cultură sau la selecţia raselor
BAZELE EREDITĂŢII
Fig. 14 Transmiterea ereditară a hemofiliei
1. Comparaţi determinismul genetic la om şi lavrabie folosind Fig. 12.2. Căutaţi un articol în care este prezentată desca-
lificarea unei sportive în urma efectuării testuluiBarr-Bertram pentru determinarea sexului. Daţiexplicaţia biologică a cazului.3. De ce musculiţa de oţet este o specie avantajoa-
să pentru studierea eredităţii? Prezentaţi argumenteşi contraargumente. 4. Observaţi fenotipul părinţilor şi descendenţilor
din încrucişările din Fig. 13. Ce ne duce la conclu-zia că trăsătura studiată poate avea legătură cu sexulindivizilor?5. Daltonismul şi hemofilia se transmit genetic în
mod asemănător. Este posibil ca doi părinţi cuvedere normală să aibă băiat sau fată afectaţi de dal-tonism? De ce?
XV
XV
XX XY XY
XY
X
X
X
X Y
XXV
XXV
XXV
Y
XVY
1-160 biologia12 2015 roman6. tord.qxd:1-80 biologia12.qxd 3/22/16 10:38 AM Page 17
18
de animale. Înlănţuirea strânsă a proprietăţilor utileale plantelor de cultură sau animalelor domesticeuşurează mult munca agricultorilor, deoarece printr-osingură încrucişare, indivizii speciei cu care se ocupăvor transmite descendenţilor lor mai multe caractereutile. Pe de altă parte, înlănţuirea – altfel spus linka-ge-ul – dintre trăsături utile şi altele categoric deza-vantajoase produce probleme serioase în practică laselecţia speciilor de cultură. Gena care determinăculoarea galbenă a seminţelor de in este strâns legatăde gena care determină conţinutul ridicat al seminţeiîn ulei de bună calitate. Din păcate şi genele caredetermină predispoziţia la boli şi producţii reduse laseminţe sunt înlănţuite cu cele anterioare. Din aceas-tă cauză, în practică nu se poate selecţiona un soi dein care să aibă coaja seminţelor galbenă, sămânţa săconţină mult ulei, să fie rezistentă la boli şi să aibăproducţie ridicată.
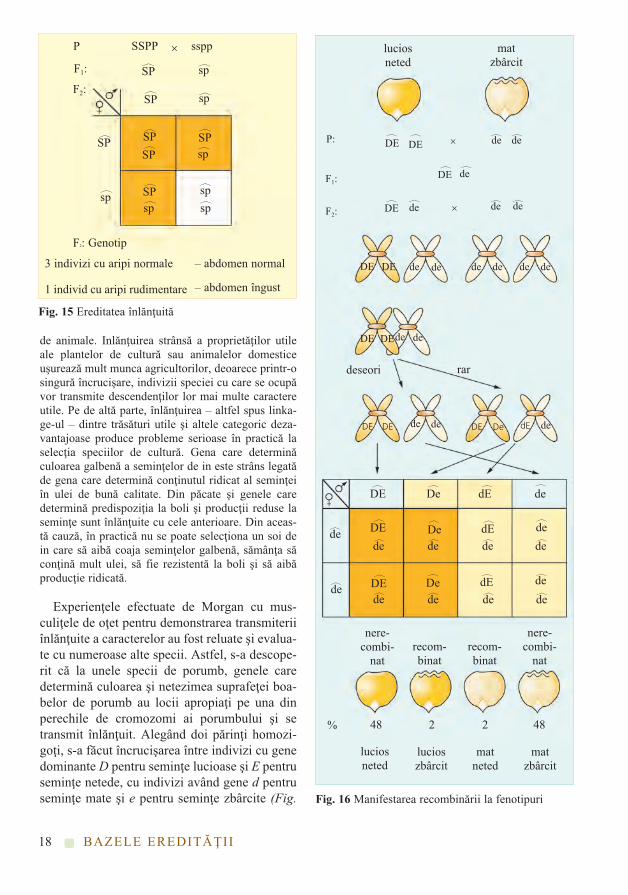
Experienţele efectuate de Morgan cu mus-culiţele de oţet pentru demonstrarea transmiteriiînlănţuite a caracterelor au fost reluate şi evalua-te cu numeroase alte specii. Astfel, s-a descope-rit că la unele specii de porumb, genele caredetermină culoarea şi netezimea suprafeţei boa-belor de porumb au locii apropiaţi pe una dinperechile de cromozomi ai porumbului şi setransmit înlănţuit. Alegând doi părinţi homozi-goţi, s-a făcut încrucişarea între indivizi cu genedominante D pentru seminţe lucioase şi E pentruseminţe netede, cu indivizi având gene d pentruseminţe mate şi e pentru seminţe zbârcite (Fig.
BAZELE EREDITĂŢII
Fig. 15 Ereditatea înlănţuită
P
F1:
F2:
�SP
�SP
�
SP�
SP
�
SP�
SP
sp
�
sp
�
sp�
sp
�sp
�sp
�sp
��
SP
SSPP sspp
F2: Genotip
3 indivizi cu aripi normale
1 individ cu aripi rudimentare
– abdomen normal
– abdomen îngust
×
Fig. 16 Manifestarea recombinării la fenotipuri
luciosneted
matzbârcit
�
DE
�
DE
�
DE
de
�
de
�
de
�
de
�
de
��
DE de
�
de
de
��
DE
�
DE
�
DE
de
�
de
�
de
�
de
�
de�
de�
de
�
de
�
luciosneted
lucioszbârcit
matneted
matzbârcit
% 48 2 2 48
P:
F1:
F2:
×
×
DE DE de de de de de
deDE DE de
deDE DE de dEDE De de�
De�
dE
de
�
de
�
de
�
de
�
�
De
�
dE
�
dE
�
De
deseori rar
nere-combi-
natrecom-binat
recom-binat
nere-combi-
nat
1-160 biologia12 2015 roman6. tord.qxd:1-80 biologia12.qxd 3/22/16 10:38 AM Page 18
19
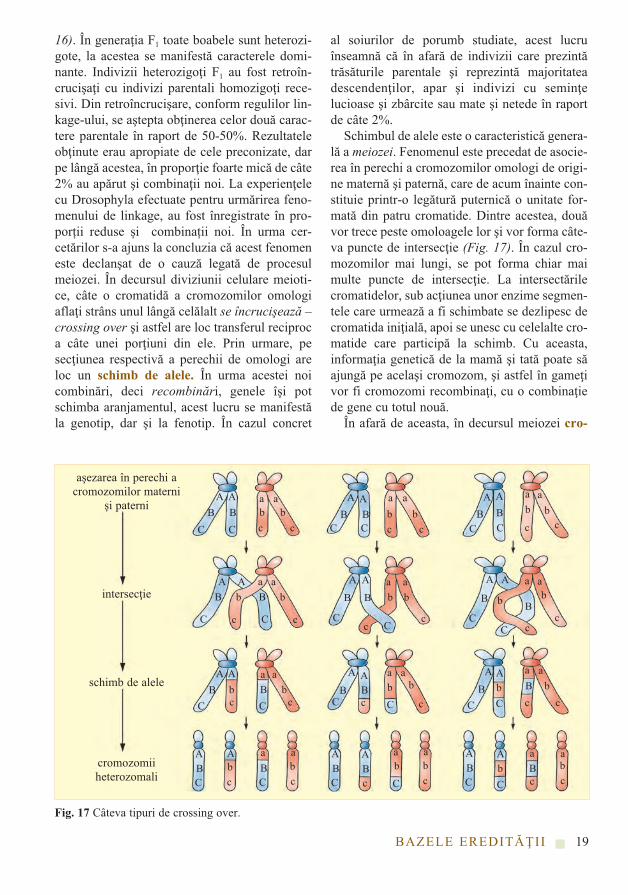
16). În generaţia F1 toate boabele sunt heterozi-gote, la acestea se manifestă caracterele domi-nante. Indivizii heterozigoţi F1 au fost retroîn-crucişaţi cu indivizi parentali homozigoţi rece-sivi. Din retroîncrucişare, conform regulilor lin-kage-ului, se aştepta obţinerea celor două carac-tere parentale în raport de 50-50%. Rezultateleobţinute erau apropiate de cele preconizate, darpe lângă acestea, în proporţie foarte mică de câte2% au apărut şi combinaţii noi. La experienţelecu Drosophyla efectuate pentru urmărirea feno-menului de linkage, au fost înregistrate în pro-porții reduse și combinații noi. În urma cer -cetărilor s-a ajuns la concluzia că acest fenomeneste declanşat de o cauză legată de procesulmeiozei. În decursul diviziunii celulare meioti-ce, câte o cromatidă a cromozomilor omologiaflaţi strâns unul lângă celălalt se încrucişează –crossing over şi astfel are loc transferul reciproca câte unei porţiuni din ele. Prin urmare, pesecţiunea respectivă a perechii de omologi areloc un schimb de alele. În urma acestei noicombinări, deci recombinări, genele îşi potschimba aranjamentul, acest lucru se manifestăla genotip, dar şi la fenotip. În cazul concret
al soiurilor de porumb studiate, acest lucruînseamnă că în afară de indivizii care prezintătrăsăturile parentale şi reprezintă majoritateadescendenţilor, apar şi indivizi cu seminţelucioase şi zbârcite sau mate şi netede în raportde câte 2%.
Schimbul de alele este o caracteristică genera-lă a meiozei. Fenomenul este precedat de asocie-rea în perechi a cromozomilor omologi de origi-ne maternă şi paternă, care de acum înainte con-stituie printr-o legătură puternică o unitate for-mată din patru cromatide. Dintre acestea, douăvor trece peste omoloagele lor şi vor forma câte-va puncte de intersecţie (Fig. 17). În cazul cro-mozomilor mai lungi, se pot forma chiar maimulte puncte de intersecţie. La intersectărilecromatidelor, sub acţiunea unor enzime segmen-tele care urmează a fi schimbate se dezlipesc decromatida iniţială, apoi se unesc cu celelalte cro-matide care participă la schimb. Cu aceasta,informaţia genetică de la mamă şi tată poate săajungă pe acelaşi cromozom, şi astfel în gameţivor fi cromozomi recombinaţi, cu o combinaţiede gene cu totul nouă.
În afară de aceasta, în decursul meiozei cro-
BAZELE EREDITĂŢII
Fig. 17 Câteva tipuri de crossing over.
A A
B B
C C
A A
B B
C C
a a
b b
c c
A A
B BC C
a a
b b
c c
a ab b
c c
A A
B B
CC
a a
b b
cc
A A
B
C C
a a
b B bc c
A
BC
AB
Cc c
a a
b b
A
B
C
A
B
C c
a
b
c
ab
c
a
b
c
a
bA
BC
A
B
C
A
B
C
AB
C c
a
b
c
a
b
ABC c
abA
BC c
ba A
BC
ABc c
ba
C
ba A
BC
Ab
C c
ba
cB
a
aşezarea în perechi acromozomilor materni
şi paterni
intersecţie
schimb de alele
cromozomii heterozomali
1-160 biologia12 2015 roman6. tord.qxd:1-80 biologia12.qxd 3/22/16 10:38 AM Page 19