generalità sui protisti. · generalità sui protisti. eterogeneo , evolutosi dalle monere e dal...
TRANSCRIPT

I protisti e gli altri eucarioti.
In molti casi è difficile tracciare una linea di
demarcazione netta fra piante, funghi e animali, tutti
organismi che si sono evoluti da comuni progenitori
protisti seguendo vie diverse.
Generalmente i protisti con caratteristiche simili a
quelle degli animali, vengono definiti protozoi e distinti
dai protofiti (alghe) che rappresentano protisti con
caratteristiche di tipo vegetale. Esistono inoltre protisti
che possono essere paragonati ai funghi: le muffe
mucillaginose e le muffe d'acqua.
1) La distinzione tra protisti e animali è relativamente
semplice: si definisce animale un organismo
pluricellulare, eterotrofo, che si nutre per ingestione.
2) Le piante differiscono invece dai protisti per la
capacità di sviluppare embrioni protetti dai tessuti
materni.
3) La distinzione tra funghi e protisti è più complicata
ma è comunque basata:
- sull‟eventuale presenza di flagelli (quasi mai presenti
nei funghi)
- sul tipo di gameti (in alcuni protisti esistono gameti
maschili e femminili morfologicamente diversi, mentre
ciò non si verifica nei funghi)
- sulla presenza di uno stadio dicariotico (descritto nei
funghi)
- sulla composizione chimica della parete cellulare (i
funghi possiedono pareti costituite principalmente da
chitina).
Prima di descrivere le modalità di vita e la
classificazione degli eucarioti microbici, consideriamo le
differenze tra le cellule eucariotiche e quelle
procariotiche, cercando di capire come è avvenuto, nella
storia della vita, il passaggio da procarioti a eucarioti.
La perdita della parete, la comparsa del citoscheletro e delle membrane interne.
La cellula eucariotica presenta moltissime differenze
rispetto a quella procariotica. In particolare, nella cellula
eucariotica sono presenti un ampio sistema
di membrane interne e numerosi organuli che,
nell‟insieme, svolgono complesse reazioni metaboliche;
inoltre, il DNA è contenuto all‟interno di un nucleo vero
e proprio, circondato da una membrana. Secondo i biologi, è impossibile che tutte queste novità
siano comparse simultaneamente nel corso
dell‟evoluzione. I resti fossili testimoniano che la cellula
eucariotica comparve più di due miliardi di anni fa, ma
non esistono dati che ci permettano di definire con
certezza come sia avvenuto il passaggio dal progenitore
procariotico. Proviamo allora a ricostruire la sequenza
più logica che possa avere portato al nuovo tipo
cellulare, senza dimenticare i cambiamenti globali
concomitanti, come il passaggio graduale da
un‟atmosfera priva di ossigeno a un‟atmosfera aerobica.
Il primo passo verso la condizione eucariotica fu
probabilmente la perdita della parete cellulare tipica
della cellula procariotica ancestrale (▶figura 1). La
perdita di questo involucro rigido rappresenta una
conquista vantaggiosa per numerose ragioni: senza la
parete, la cellula può aumentare maggiormente di
dimensioni; inoltre, la superficie cellulare diventa
flessibile, favorendo la comparsa di introflessioni ed
estroflessioni della membrana (utili per inglobare cibo)
ed aumentando l‟area destinata agli scambi con
l‟esterno.
Il ripiegamento della membrana plasmatica di una
grossa cellula procariotica avrebbe dato origine al
sistema di membrane interne e al nucleo, mentre un
ulteriore stadio evolutivo sarebbe stato rappresentato
dalla comparsa di un citoscheletro.
1. Generalità sui Protisti.
Con il termine protisti si intende di regola un raggruppamento biologico (regno) piuttosto eterogeneo, evolutosi dalle monere
e dal quale hanno successivamente preso origine tutti gli organismi eucariotici. Secondo una definizione più generica,
possiamo definire protisti tutti quegli organismi eucariotici che non è possibile classificare come piante, funghi o animali.
Alcuni biologi considerano veri protisti soltanto gli organismi unicellulari e includono quelli pluricellulari in uno degli altri tre
regni degli eucarioti. In realtà, vedremo che anche certi organismi a struttura pluricellulare possono essere classificati come
protisti.
I protisti hanno nel complesso caratteristiche metaboliche sorprendentemente varie, e in ciò essi sono secondi soltanto alle
monere. Dal punto di vista della nutrizione, ad esempio, alcuni sono autotrofi mentre altri eterotrofi e con specializzazioni
alimentari assai diverse; in alcuni casi è stata addirittura osservata la loro capacità di passare rapidamente dall'autotrofia alla
eterotrofia.
I protisti sono presenti in ogni ambiente; la maggior parte sono comunque acquatici, diffusi sia in ambiente marino che
d‟acqua dolce, ma anche nei liquidi corporei di altri organismi, nei terreni umidi o nelle cortecce degli alberi marcescenti.

Il citoscheletro è costituito da microfilamenti e
microtubuli formati da particolari proteine; oltre a
sostenere la cellula e a mantenerne la forma, è utile
anche nella divisione cellulare e per il trasporto di
materiali nel citoplasma. In alcune cellule, inoltre, i
microtubuli potrebbero essersi evoluti dando origine a
un flagello. Il primo vero eucariote possedeva quindi un
citoscheletro e un involucro nucleare, alcuni organuli e
forse anche uno o due flagelli.
L’acquisizione per endosimbiosi di mitocondri e cloroplasti.
Mentre tutto questo stava avvenendo, i cianobatteri
producevano ossigeno con la fotosintesi, innescando
disastrose conseguenze per la maggior parte degli altri
viventi, che erano anaerobici. Alcuni procarioti, tuttavia,
riuscirono ad adattarsi, dando origine ai primi eucarioti
capaci di fagocitosi. A questo punto, può essere entrata
in gioco l‟endosimbiosi (▶figura 1), grazie alla quale si
sarebbero originati i primi mitocondri e cloroplasti.
Figura 1. Dalla cellula procariotica alla cellula eucariotica.
Una possibile sequenza degli eventi evolutivi che hanno portato alla comparsa della cellula eucariotica.
Secondo la teoria dell‟endosimbiosi, i mitocondri
avrebbero avuto origine da proteobatteri eterotrofi
capaci di utilizzare l‟ossigeno. Questi proteobatteri
sarebbero stati incorporati in cellule eucariotiche più
grandi, evolvendosi poi in mitocondri.
Inizialmente la funzione principale dei nuovi organuli
era quella di limitare la tossicità dell‟ossigeno
trasformandolo in acqua; più tardi, questo processo
venne accoppiato alla sintesi di molecole ad alto
contenuto energetico (ATP), dando il via
alla respirazione cellulare. L‟endosimbiosi avrebbe
quindi conferito grandi vantaggi sia all‟ospite (la
possibilità di disporre di una maggiore quantità di
energia e la capacità di vivere in ambiente aerobico) sia
ai piccoli proteobatteri inglobati, che potevano
svilupparsi in un ambiente protetto. L‟interdipendenza
tra la cellula ospite e il proteobatterio sarebbe diventata
sempre più stretta, tanto da rendere impossibile la
sopravvivenza dei due sistemi separati. Alla fine di
questo passaggio, la cellula eucariotica moderna risultò
completa. Alcuni eucarioti oggi molto diffusi, le piante,
sono il risultato di un‟ulteriore specializzazione:
l‟incorporazione di un procariote simile ai cianobatteri
attuali, destinato a diventare il cloroplasto. Si sarebbero
formate così le cellule eucariotiche fotosintetiche tipiche
dei vegetali.
Ci sono varie prove a sostegno del modello
endosimbiontico. Per esempio, i mitocondri e i
cloroplasti odierni assomigliano molto ai batteri e infatti
contengono piccole quantità di DNA, RNA e ribosomi
più simili a quelli delle cellule batteriche che a quelli
eucariotici. Inoltre, questa teoria può spiegare anche
perché mitocondri e cloroplasti sono gli unici organuli
cellulari circondati da due membrane: quella interna
corrisponderebbe alla membrana plasmatica del
batterio e quella esterna sarebbe quella della cellula
ospite che si è invaginata per accoglierlo.
La cellula eucariotica possiede caratteristiche molto
funzionali: dal citoscheletro, che regola gli spostamenti
nei comparti interni della cellula e permette varie
modalità di locomozione, a organuli altamente
specializzati. Come vedremo, grazie a questi nuovi
strumenti i protisti hanno imparato a sfruttare molti
ambienti e molte risorse.
Riproduzione.
Sebbene la maggior parte degli eucarioti microbici
mostri entrambi i tipi di riproduzione, asessuata e
sessuata, alcuni gruppi non presentano affatto una
riproduzione di tipo sessuato. Inoltre, come vedremo,
non sempre sessualità e riproduzione sono legate in
modo diretto.
Riproduzione asessuata nei protisti
I quattro processi di riproduzione asessuata che si
riscontrano nei protisti sono i seguenti:
(1) scissione binaria, cioè semplice divisione della
cellula dell‟organismo genitore;
(2) scissione multipla, ovvero suddivisione del genitore
in più di due cellule figlie;
(3) gemmazione, cioè crescita di un nuovo individuo
sulla superficie della cellula originaria;
(4) formazione di spore, cellule specializzate capaci di
svilupparsi in nuovi organismi completi.

Questi tipi di riproduzione offrono il vantaggio di
produrre un grande numero di discendenti in tempi
rapidi, e risultano particolarmente efficienti se le
condizioni ambientali sono favorevoli. In tutti e quattro i
casi i figli saranno perfettamente identici al
microrganismo di partenza; una popolazione di
individui geneticamente identici, originati per
riproduzione asessuata, è chiamata clone.
La riproduzione sessuata: dai gameti allo zigote
La riproduzione sessuata è la più complicata e comporta
la formazione di speciali cellule chiamate gameti. I
gameti si originano per meiosi, un tipo di divisione che
dimezza il corredo genetico della cellula originaria. Con
la fecondazione, il gamete proveniente dal padre si
fonde con quello proveniente dalla madre dando origine
a un nuovo organismo chiamato zigote.
Alternanza di generazione.
Alcune alghe e alcuni protisti simili ai funghi sono
caratterizzati da alternanza di generazioni, nella quale
un organismo diploide produttore di spore (sporofito)
da origine a un organismo pluricellulare aploide
produttore di gameti (gametofito); dalla fusione di due
gameti si origina successivamente un nuovo organismo
diploide (▶figura 2). L'organismo diploide, quello
aploide, oppure entrambi, possono anche riprodursi
asessualmente. L'organismo aploide e quello diploide
differiscono dunque geneticamente, ma possono anche
non mostrare differenze a livello morfologico. Se il
gametofito è morfologicamente uguale allo sporofito, si
parla di alternanza di generazione isomorfa, viceversa,
si alternanza di generazioni eteromorfa.
Figura 2. Alternanza di generazione. In alcuni protisti si può osservare un'alternanza tra la generazione diploide, che produce spore, e la generazione aploide, che produce gameti.
Di regola, nei protisti pluricellulari, nei funghi e nelle
piante, i gameti non vengono prodotti direttamente dal
processo meiotico. Alcune cellule specializzate
dell'organismo diploide, definite sporociti, si dividono
infatti per meiosi producendo quattro spore da cui
prenderà origine, in seguito a successive divisioni
mitotiche, un organismo pluricellulare aploide, detto
gametofito. Quest'ultimo produrrà i gameti per mitosi o
per citocinesi. Al contrario delle spore, i gameti
possono invece produrre nuovi organismi soltanto
fondendosi con altri gameti. Dalla fusione di due gameti
si origina uno zigote diploide e da quest'ultimo, per
successive divisioni mitotiche, deriva lo sporofito. Dagli
sporociti prodotti dallo sporofito si formano infine
spore aploidi che ricominciano il ciclo.
Esistono poi casi di processi sessuati ma non
riproduttivi, come la coniugazione che si verifica nei
ciliati.
Particolarità di alcuni protisti.
Vacuoli contrattili.
I protisti che vivono nelle acque dolci si trovano immersi
in un mezzo ipotonico, e poiché possiedono un
potenziale osmotico più negativo di quello
dell'ambiente esterno, essi tendono ad assorbire
costantemente acqua per osmosi. Diversi gruppi di
protisti possiedono tuttavia particolari strutture definite
vacuoli contrattili (▶figura 3), che consentono loro di
vivere in ambienti ipotonici: l'acqua assorbita in eccesso
viene infatti convogliata nei vacuoli ed espulsa.
Figura 3. I vacuoli contrattili. Quando il vacuolo è pieno, un minuscolo poro si apre alla superficie della cellula permettendo la comunicazione con l'esterno. A questo punto il vacuolo si contrae rapidamente espellendo all‟esterno il proprio contenuto.
Un semplice ma interessante esperimento conferma che
l'espulsione dell'acqua costituisce la funzione principale
dei vacuoli contrattili. Osservando dei parameci al
microscopio, si possono facilmente notare i loro vacuoli
che si contraggono con una frequenza caratteristica.
Ponendo successivamente alcuni di questi organismi in
soluzioni con differente potenziale osmotico, si può
osservare come il ritmo di contrazione dei loro vacuoli
pulsanti cambi in relazione alla differenza di potenziale
con l'ambiente esterno: quanto meno negativo sarà il
potenziale della soluzione, tanto più ipertonici saranno i
parameci rispetto a essa e tanto più velocemente i loro
vacuoli assorbiranno acqua e aumenteranno il ritmo di
contrazione. Al contrario, i vacuoli cesseranno di
contrarsi quando la concentrazione del mezzo di
sospensione sarà isotonica rispetto all'ambiente
cellulare dei parameci.

Endosimbiosi.
Il fenomeno dell‟endosimbiosi prevede che organismi
appartenenti a specie diverse vivano insieme, uno
dentro l'altro ed è largamente diffuso fra i protisti, che
possono fungere sia da ospiti che da endosimbionti. Ad
esempio il protista Myxotricha paradoxa, ospita al suo
interno e sulla sua superficie una varietà di batteri
simbionti. Tutti i radiolari (phylum Sarcodina), ad
esempio, ospitano al loro interno protisti fotosintetici
(▶figura 4) e appaiono verdastri o giallastri a seconda
del tipo di endosimbionti. Quest'associazione risulta
vantaggiosa sia per i radiolari, che possono così
sfruttare i prodotti della fotosintesi degli ospiti, sia per
gli endosimbionti, che presumibilmente possono
beneficiare sia dei metaboliti prodotti dai radiolari che
di una certa protezione fisica.
I Flagellati o Mastigofori.
Si tratta del più ampio (oltre 10000 specie descritte) e di
gran lunga più eterogeneo phylum di protisti.
Tutti i membri del phylum Mastigofori possiedono uno
o più flagelli e sono per questo comunemente definiti
Flagellati.
All‟interno dei protozoi flagellati esiste una notevole
diversità morfologica.
In passato venivano inclusi in questo phylum sia
organismi autotrofi che eterotrofi e a causa della forte
diversità nelle specializzazioni alimentari, i
Mastigophora sono spesso considerati cellule con
caratteristiche intermedie tra piante e animali. Per
questo motivo alcuni biologi hanno proposto di
suddividere i flagellati in due sottogruppi:
−i fitoflagellati, le forme autotrofe, più simili a piante;
− gli zooflagellati, le forme eterotrofe più simili agli
animali,.
In realtà, alcune specie di fitoflagellati coloniali, di
colore verde e relativamente grandi ed altre che
ricordano addirittura gli stadi acquatici mobili di alcune
piante terrestri, vengono considerate alghe verdi dalla
maggior parte dei botanici.
Per comodità didattica, considereremo in questo
paragrafo soltanto le forme eterotrofe mentre
tratteremo quelle autotrofe tra le alghe.
Figura 4. L‟endosimbiosi tra alghe e radiolari. Alcune alghe endosimbionti che vivono all'interno di questi radiolari, forniscono loro il cibo e parte della pigmentazione visibile attraverso il rivestimento esterno. Sia i radiolari che le alghe sono protisti.
Una grande quantità di flagellati vivono in simbiosi
all‟interno di altri animali. Ad esempio, nell'apparato
digerente delle termiti che si nutrono di legno è
presente una vasta gamma di flagellati che possiedono
alcune fra le più bizzarre e complicate strutture
corporee descritte fra i protisti.
Altre specie sono invece parassite e tra di esse si
ritrovano anche molti organismi patogeni per l'uomo.
La malattia del sonno è una malattia tropicale diffusa
solo nelle regioni equatoriali dell'Africa provocata dal
protozoo flagellato parassita, Trypanosoma brucei
(▶figura 5).
Figura 5. Trypanosoma brucei gambiense.
E‟ il flagellato parassita che rappresenta la causa della malattia del sonno. Nei tripanosomi il flagello è disposto lungo il margine della cellula, come parte di una struttura detta membrana ondulante.
2. I protozoi: protisti simili agli animali.
I protozoi sono eucarioti unicellulari, eterotrofi per ingestione.
I protozoi sono organismi eucariotici unicellulari, capaci di compiere movimenti, di reagire a stimoli di varia natura, di
ingerire particelle alimentari e di riprodursi.
I protozoi vengono suddivisi in quattro phyla a seconda della capacità di movimento e delle modalità della locomozione:
Flagellati si spostano mediante flagelli
Sarcodini si spostano emettendo pseudopodi
Ciliati si spostano mediante ciglia
Sporozoi non sono dotati di strutture adibite al movimento

Il vettore (ospite intermedio) di questo parassita è una
mosca del genere Glossina, comunemente indicate con
il nome di mosca tse-tse (▶figura 6), che può infettare
sia gli animali selvatici che il bestiame domestico o
l'uomo.
Figura 6. Glossina palpalis, la mosca tse-tse.
Questo genere di mosca è il vettore che trasmette il tripanosoma da una persona malata ad una sana, infettandola.
Il nome "malattia del sonno" è dovuto al fatto che
l'ultimo stadio della malattia è caratterizzata da apatia,
sonnolenza e cachessia. Nell'ultima fase di questo
stadio il malato non è più in grado di alzarsi né di
mangiare. Si può diagnosticare mediante
l'identificazione del parassita nel sangue (nello stadio
iniziale) o nel liquido cefalo-rachidiano (nello stadio
avanzato).
Purtroppo sono disponibili solo farmaci sviluppati a
metà del secolo scorso che presentano importanti
effetti collaterali e scarsa efficacia terapeutica. È
interessante chiedersi come mai per malattie così
importanti come le tripanosomiasi non esistano ancora
farmaci a grande diffusione. Una prima considerazione
è che la sperimentazione «privata» di un nuovo farmaco
richiede cospicui investimenti e molti anni di ricerche:
ciò significa che gli eventuali ricavi derivati dalle future
vendite dovrebbero compensare questi investimenti.
Purtroppo, le tripanosomiasi colpiscono soprattutto le
persone più povere dei Paesi in via di sviluppo, che non
offrono incentivi finanziari ai potenziali investitori
farmaceutici. Come fanno notare le organizzazioni
internazionali che si occupano di salute, si tratta delle
«malattie dimenticate» dai Paesi cosiddetti sviluppati.
Un‟altra considerazione è che anche in Paesi che
possiedono le capacità tecniche e industriali per la
ricerca e la produzione di medicine, malattie di questo
genere sono ugualmente difficili da combattere. I
tripanosomi riescono infatti a ingannare il sistema
immunitario umano, i vaccini stessi e gli eventuali
farmaci perché mutano rapidamente, modificando di
continuo le molecole di riconoscimento della superficie
cellulare propria o della cellula infettata.
Infine, a differenza delle cellule procariotiche dei
batteri, questi microrganismi unicellulari sono
eucariotici; le loro cellule sono molto simili alle nostre,
e i farmaci in grado di ucciderli risultano tossici anche
per il nostro organismo.
Il ciclo vitale di Trypanosoma è descritto in (▶figura 7).
Figura 7. Ciclo vitale di Trypanosoma brucei. Il ciclo prevede due fasi, una all‟interno del corpo della mosca tse-tse, e l‟altro all‟interno del corpo umano.
Un altro flagellato parassita è Leishmania, un genere di
protozoi parassiti responsabili della leishmaniosi umana
e animale. È responsabile delle varie forme di
leishmaniosi, diffuse in Eurasia, Africa e America, che
provocano danni a livello cutaneo (▶figura 8) e viscerale
ed interferiscono con le risposte immunitarie; in
mancanza di trattamenti opportuni essi possono essere
letali.
Figura 8. Leishmaniosi.
Tipica lesione cutanea alla mano provocata dalla leishmania.
In Europa ed in Asia è diffusa da insetti del
genere Phlebotomus (pappataci) (▶figura 9). Gli ospiti
primari di questo parassita sono i vertebrati. Leishmania
comunemente infetta roditori, procavie, canidi e l'uomo.
Circa 12 milioni di persone nel mondo sono colpite da
leishmaniosi.
Figura 9. Pappatacio.
Phlebotomus papatasi, detto anche flebotomo o pappatacio è
un insetto simile a una zanzara di piccole dimensioni. E‟ molto
diffuso nell'area mediterranea.

Un gruppo particolare di flagellati comprende i parassiti
del genere Giardia e Trichomonas. Essi presentano oltre
ai flagelli, membrane ondulate che contribuiscono alla
locomozione della cellula e sono caratterizzati dalla
completa assenza di mitocondri. In passato, l‟assenza di
mitocondri che si riscontra in questi due parassiti aveva
fatto ritenere che questi potessero rappresentare il
gruppo originario degli eucarioti. Tuttavia, oggi si pensa
che questa sia una caratteristica derivata, non primitiva,
dovuta al parassitismo; probabilmente, gli antenati di
questi organismi possedevano mitocondri che, nel corso
dell‟evoluzione, si sono ridotti o sono stati persi.
L‟esistenza attuale di organismi del genere mostra
come la vita degli eucarioti sia possibile anche senza
mitocondri, un fatto che rende particolarmente
interessanti queste forme.
Giardia lamblia (▶figura 10) è un comune parassita
tropicale che contamina le acque potabili provocando la
giardiasi intestinale; questo microrganismo contiene
due nuclei cellulari ed è dotato di flagelli multipli.
Figura 10. Giardia lamblia.
E‟ il flagellato parassita che rappresenta la causa della malattia
intestinale definita giardiasi.
La giardiasi (o lambliasi) si manifesta come diarrea
acuta o cronica debilitante e/o sindrome da
malassorbimento. Il contagio avviene mediante
ingestione di alimenti e bevande contaminate da cisti.
Si può diagnosticare mediante l'identificazione del
parassita o delle sue cisti nelle feci. Si può trattare con
vari farmaci. Il ciclo vitale di Giardia è in (▶figura 11).
Figura 11. Ciclo vitale di Giardia lamblia.
Trichomonas vaginalis (▶figura 12) è invece
responsabile di una malattia a trasmissione sessuale
detta tricomoniasi che colpisce uomini e donne.
Figura 12. Trichomonas vaginalis.
E‟ il flagellato parassita che rappresenta la causa di una malattia venerea diffusa ma non grave.
La tricomoniasi nella donna può provocare forte prurito
locale, presenza di secrezioni anomale (perdite
schiumose, giallastre e maleodoranti) associate o meno
a dolore durante la minzione o i rapporti sessuali con
un'esacerbarsi dei sintomi durante le mestruazioni.
Nell'uomo può causare uretriti e prostatiti, che si
manifestano sottoforma di piccole perdite e dolore
nell'urinare. Spesso però, l‟infezione è asintomatica. Il
contagio avviene quasi sempre per trasmissione
sessuale, o anche attraverso il contatto con asciugamani
o biancheria intima. La diagnosi viene effettuata
ricercando in laboratorio la presenza del protozoo
(Trichomonas) su campioni di essudato vaginale nella
donna o di essudato uretrale o sperma nell'uomo. Si
tratta con farmaci che, somministrati per via orale,
vengono concentrati a livello urogenitale dove espletano
la propria azione. (▶figura 13).
Figura 13. Ciclo vitale di Trichomonas vaginalis.
Un gruppo di flagellati, i coanoflagellati, è infine
considerato come progenitore delle spugne, il più
antico phylum di animali tutt'ora viventi. I poriferi sono
infatti organismi coloniali, piuttosto che veri e propri
pluricellulari, e i coanoflagellati mostrano strette
somiglianze con le tipiche cellule delle spugne.

I Sarcodini.
I Sarcodini, ovvero le amebe e gli altri protisti a loro
affini, sono organismi capaci di formare pseudopodi,
cioè estensioni citoplasmatiche prive di una forma
definita. Nella letteratura popolare, il termine ameba è
spesso usato per indicare una massa informe di
sostanza vivente, ovvero la forma più semplice di vita
«animale» che si possa immaginare. L'esame di una
tipica ameba ci permette di comprendere come possa
essersi originato questo riferimento.
Un'ameba (▶figura 14) consiste in una singola cellula
che si nutre di piccoli organismi e di particelle
organiche per fagocitosi, cioè mediante l'emissione di
processi citoplasmatici definiti pseudopodi. Le particelle
alimentari vengono concentrate in vacuoli alimentari
all'interno del citoplasma e qui lentamente digerite e
assimilate. Gli pseudopodi funzionano anche come
organi di movimento.
Figura 14. Amoeba proteus.
Nella foto si vede un ameba con diversi pseudopodi.
Nonostante le apparenze, l'ameba non è tuttavia un
organismo primitivo. Sempre maggiori indizi
suggeriscono infatti che la semplice struttura di questi
organismi sia in realtà una condizione derivata
secondariamente nel corso dell'evoluzione e che il
phylum dei Sarcodina si sia originato da progenitori
appartenenti al phylum dei Mastigophora.
Figura 15. Un'ameba con caratteristiche intermedie Mastigamoeba aspera possiede sia pseudopodi che flagello; questa specie viene considerata spesso come una forma intermedia tra il phylum dei Mastigophora e quello dei Sarcodina.
Mastigamoeba aspera, (▶figura 15) una specie con
caratteristiche intermedie tra i Sarcodina e i
Mastigophora potrebbe essere classificata a buon diritto
in entrambi i phyla.
Inoltre, le amebe del genere Naegleria, alcune delle
quali possono causare una malattia letale del sistema
nervoso, possiedono un ciclo biologico a due stadi, uno
rappresentato da cellule di tipo ameboide, e l'altro da
cellule flagellate.
Le amebe sono quindi forme piuttosto specializzate di
protisti, adattate alla vita acquatica in ambienti lacustri,
fluviali o di altro genere. Il loro tipico movimento
ameboide e la modalità con cui inglobano particelle
alimentari, le rendono tuttavia adatte soprattutto a una
vita in ambienti ricchi di organismi sedentari o di
particelle organiche non mobili.
Altri Sarcodina mostrano un grado senz'altro maggiore
di specializzazione; la loro organizzazione cellulare è
più simile a quella animale e consente un adattamento
al ruolo di predatori, parassiti o detritivori. Nessuna di
queste forme è invece fotosintetica. Vi sono inoltre
amebe che vivono entro particolari rivestimenti formati
da granelli di sabbia cementati insieme, oppure
all'interno di gusci spinosi o squamosi da loro stessi
prodotti.
Un altro interessante gruppo di Sarcodina è quello dei
foraminiferi (▶figura 16), organismi prevalentemente
marini capaci di secernere involucri di carbonato di
calcio, (CaCO3) I loro pseudopodi sono lunghi, filiformi e
ramificati e sporgono all'esterno del guscio attraverso
minuscoli pori (foramina), formando una sorta di rete
viscosa che viene impiegata per catturare altri
organismi. Dopo la riproduzione, che avviene per
mitosi, le nuove cellule abbandonano il guscio della
cellula madre e ne formano altri.
Figura 16. I foraminiferi sono dotati di rivestimenti calcarei simili a minuscole conchiglie.
I gusci dei foraminiferi sono formati da una matrice proteica indurita da carbonato di calcio. Nell‟arco di milioni di anni, i loro residui hanno dato origine a immensi depositi di calcare e alle spiagge sabbiose. In questa microfotografia sono illustrate numerose specie diverse.

I rivestimenti dei foraminiferi, accumulatisi sui fondi
oceanici nel corso delle ere geologiche, formano in varie
parti del mondo depositi calcarei spessi fino ad alcune
migliaia di metri ed estesi per milioni di chilometri
quadrati. Gli scheletri dei foraminiferi formano talvolta
anche la sabbia di alcune spiagge, e un grammo di
questa sabbia può contenere fino a 50000 gusci. I
rivestimenti delle singole specie di foraminiferi hanno
forme caratteristiche e si conservano facilmente come
fossili nei sedimenti marini (▶figura 17).
Figura 17. Le bianche scogliere di Dover.
I gusci di foraminiferi che dopo la morte si sono accumulati e depositati sui fondali marini hanno permesso nel corso di millenni la formazione di queste famose scogliere di natura calcarea che si affacciano sul canale della Manica e sono state immortalate da poeti e cantanti.
Ogni periodo geologico è, fra l'altro, caratterizzato da
peculiari specie di tali organismi e per questa ragione,
oltre al fatto di essere particolarmente abbondanti, i
resti dei foraminiferi sono preziosi indicatori per la
classificazione e la datazione delle rocce sedimentarie.
Gli eliozoi (▶figura 18) sono Sarcodini caratterizzati da
un rivestimento di pseudopodi rigidi e di forma
allungata. Come i foraminiferi, essi vivono liberi
nell'acqua (sia dolce che salata) e utilizzano gli
pseudopodi per intrappolare organismi di dimensioni
inferiori.
Figura 18. Un eliozoo.
Gli eliozoi sono sarcodini dotati di pseudopodi rigidi disposti a raggiera che conferiscono al protozoo la forma di “sole”.
Un terzo gruppo con caratteristiche biologiche simili
alle precedenti è quello dei radiolari (▶figura 19). Si
tratta di protisti esclusivamente marini, fra i quali si
possono osservare forme particolarmente eleganti.
Quasi tutte le specie di radiolari sono capaci di
secernere involucri trasparenti di solito silicei (SiO2), dai
quali si protendono verso l'esterno pseudopodi
aghiformi. Gli involucri delle diverse specie sono
estremamente vari e risultano spesso caratterizzati da
elaborati disegni geometrici. Alcuni radiolari sono
inoltre fra i protisti di maggiori dimensioni, con
involucri che misurano diversi millimetri di diametro.
Figura 19. I radiolari sono caratterizzati da un involucro vitreo finemente decorato.
I radiolari sono dotati di prolungamenti a forma di spicole (A) e secernono complessi rivestimenti silicei (B).
Grandi ammassi di scheletri di radiolari, risalenti anche
a 700 milioni di anni fa, formano vasti sedimenti sui
fondi degli oceani, soprattutto nelle regioni tropicali e
da questi sedimenti si sono formate rocce dette
radiolariti (▶figura 20).
Figura 20. Le radiolariti sono rocce sedimentarie formatesi grazie all‟accumulo di gusci di radiolari.
Queste curiose rocce rosse sono il prodotto dell‟accumulo, sul fondo di un oceano scomparso, dei resti di piccoli radiolari marini vissuti intorno ai 160 milioni di anni fa. Il nome delle rocce “radiolariti” deriva dagli organismi che le hanno prodotte.

I Ciliati.
I Ciliati, sono organismi unicellulari caratterizzati dalla
presenza di ciglia sulla loro superficie cellulare. Insieme
ai Mastigophora, questo gruppo spicca per varietà di
forme e importanza ecologica. I ciliati sono tutti
organismi eterotrofi e possiedono strutture corporee
molto più specializzate della maggior parte dei flagellati
e degli altri protisti.
Una caratteristica peculiare dei Ciliati è la presenza di
due diversi tipi di nuclei, un macronucleo e, all'interno
della stessa cellula, fino a 80 micronuclei. I micronuclei,
tipici nuclei eucariotici diploidi, sono essenziali per la
ricombinazione del patrimonio genetico delle cellule,
mentre i macronuclei, che possono essere sia diploidi
che poliploidi, controllano la sintesi proteica e
presiedono alle attività vegetative.
Paramecium (▶figure 21 e 22) un genere
particolarmente studiato, può essere scelto come
esempio per descrivere la complessa struttura e il
comportamento dei ciliati. La superficie della cellula,
dalla tipica forma a fagiolo, è coperta da un'elaborata
pellicola, una struttura composta principalmente da una
membrana esterna e da uno strato interno di strutture
vescicolari strettamente aderenti, definite alveoli. La
pellicola comprende anche uno strato di organuli, detti
tricocisti, con funzione difensiva, che in caso di
stimolazione meccanica o chimica possono essere
emessi in pochi millisecondi sotto forma di filamenti
rigidi.
Figura 21. I parameci hanno una struttura complessa.
Il disegno illustra schematicamente l‟anatomia interna di
un Paramecium.
Le ciglia sono strutture capaci di garantire un tipo di
locomozione generalmente più preciso di quello
generato dai flagelli o dagli pseudopodi. Un paramecio
può infatti dirigere il battito delle ciglia per spostarsi sia
avanti che indietro, con un tipico movimento a spirale.
Esso è inoltre in grado di cambiare rapidamente
direzione qualora incontri uno stimolo negativo. Alcuni
fra i ciliati di maggiori dimensioni detengono il primato
della velocità nel regno dei protisti, superando i due
millimetri al secondo. Alcune ciglia hanno inoltre una
funzione sensoria e sono capaci di trasmettere gli
stimoli al resto del citoplasma, rendendo possibile una
maggiore coordinazione dei rapidi movimenti dell'intero
organismo.
Figura 22. Paramecio.
La fotografia mostra un esemplare di un Paramecium.
I parameci si riproducono generalmente per divisione
cellulare: i micronuclei si dividono mitoticamente,
mentre i macronuclei vengono semplicemente ripartiti
fra le cellule figlie (▶figura 23).
Figura 23. Ciclo vitale di Paramecium. In Paramecium si alternano riproduzione asessuata e coniugazione. Nei parameci è stata descritta anche una forma
piuttosto elaborata di riproduzione sessuale detta
coniugazione (▶figura 24).
Figura 24. Coniugazione in Paramecium. Quando due individui entrano in coniugazione, si fondono e si scambiano i micronuclei, e ciò consente una ricombinazione genetica. Dopo la coniugazione, le cellule si separano e tornano a vivere come individui indipendenti.
In questo processo due individui si portano a stretto
contatto e uniscono le loro regioni anteriori. Nelle ore
successive, in ognuna delle due cellule tutti i
micronuclei degenerano, tranne uno che si divide per
meiosi. Ognuno dei quattro nuclei così formatisi va
successivamente incontro a divisione mitotica; si
ottengono così otto nuclei aploidi, sette dei quali
degenerano e uno si riproduce nuovamente per mitosi.
Il risultato finale è quindi la produzione di due nuclei
aploidi. Nel frattempo il macronucleo degenera e
scompare. Uno dei due nuclei aploidi rimane nella
cellula di origine, mentre l'altro migra nella cellula del
partner dove si fonde con il corrispondente nucleo
aploide: in questo modo si attua un processo di
ricombinazione genetica con formazione di un nuovo
nucleo diploide. Questo scambio è fra l'altro reciproco e
ognuno dei due parameci cede e riceve la stessa
quantità di DNA. Dopo che i due individui si sono
separati, il nucleo diploide di ciascuno si divide
mitoticamente, producendo due nuovi nuclei diploidi. Di
questi, uno è destinato a dare origine al macronucleo e
l'altro, per successive mitosi, ai micronuclei.
La coniugazione di Paramecium è un processo sessuale
di ricombinazione genetica, ma non può essere
considerato un processo riproduttivo poiché comincia e
termina con le stesse due cellule.
Tale processo risulta tuttavia periodicamente necessario
affinché le varie generazioni agamiche di parameci
possano susseguirsi.
Alcuni esperimenti hanno infatti dimostrato che i cloni a
cui non è consentito di compiere la coniugazione
possono sopravvivere per non più di 350 divisioni
cellulari prima di scomparire.
Le strutture descritte per Paramecium sono comuni alla
maggior parte dei ciliati. Alcuni di questi organismi si
caratterizzano tuttavia per la particolare complessità dei
loro organuli. In alcuni ciliati, ad esempio, alcune ciglia
possono fondersi e formare strutture dette cirri, capaci
di muoversi in maniera indipendente ma coordinata,
permettendo così all'organismo di spostarsi quasi
«camminando» sopra una superficie (▶figura 25).
Figura 25. Ciliati ipotrichi.
Questi ciliati si spostano mediante ciuffi di ciglia detti cirri,
La locomozione è inoltre coordinata da strutture
paragonabili a fibre nervose dette neurofibrille. Queste
raggiungono i singoli cirri, ed è stato sperimentalmente
dimostrato che la loro recisione provoca la perdita di
coordinazione dei cirri. In molti casi sono inoltre
presenti nel citoplasma fibre paragonabili a quelle
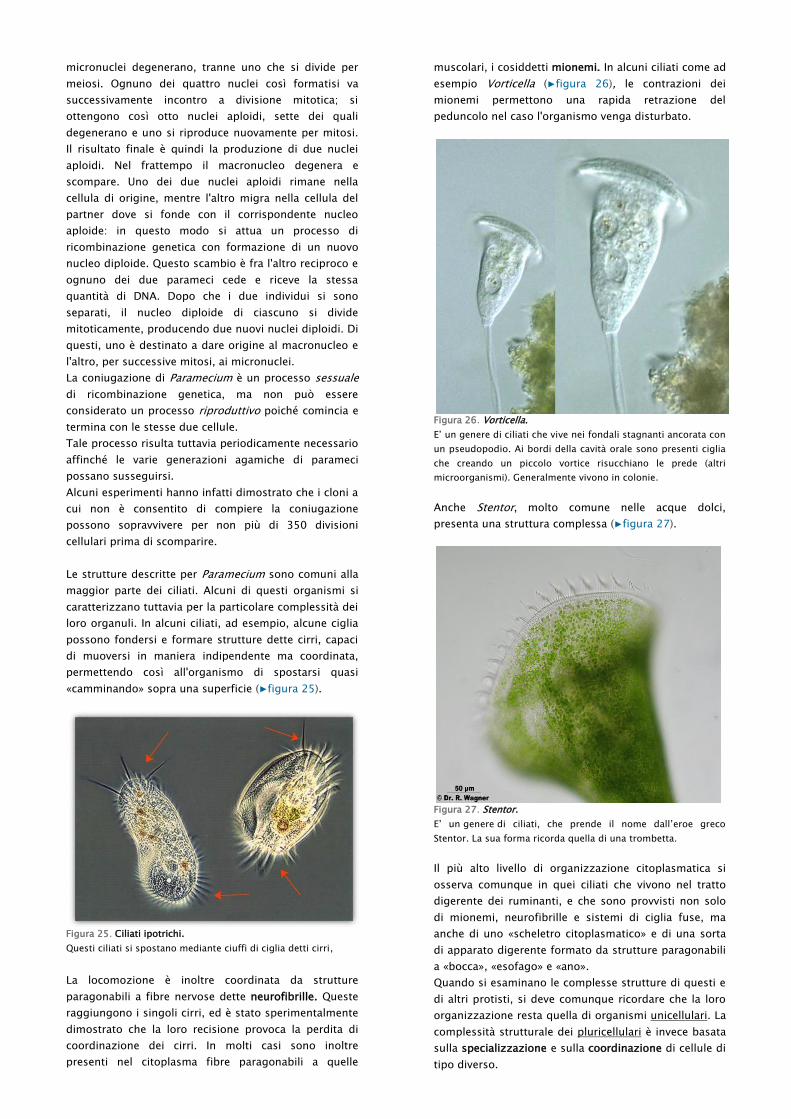
muscolari, i cosiddetti mionemi. In alcuni ciliati come ad
esempio Vorticella (▶figura 26), le contrazioni dei
mionemi permettono una rapida retrazione del
peduncolo nel caso l'organismo venga disturbato.
Figura 26. Vorticella.
E‟ un genere di ciliati che vive nei fondali stagnanti ancorata con
un pseudopodio. Ai bordi della cavità orale sono presenti ciglia
che creando un piccolo vortice risucchiano le prede (altri
microorganismi). Generalmente vivono in colonie.
Anche Stentor, molto comune nelle acque dolci,
presenta una struttura complessa (▶figura 27).
Figura 27. Stentor.
E‟ un genere di ciliati, che prende il nome dall‟eroe greco
Stentor. La sua forma ricorda quella di una trombetta.
Il più alto livello di organizzazione citoplasmatica si
osserva comunque in quei ciliati che vivono nel tratto
digerente dei ruminanti, e che sono provvisti non solo
di mionemi, neurofibrille e sistemi di ciglia fuse, ma
anche di uno «scheletro citoplasmatico» e di una sorta
di apparato digerente formato da strutture paragonabili
a «bocca», «esofago» e «ano».
Quando si esaminano le complesse strutture di questi e
di altri protisti, si deve comunque ricordare che la loro
organizzazione resta quella di organismi unicellulari. La
complessità strutturale dei pluricellulari è invece basata
sulla specializzazione e sulla coordinazione di cellule di
tipo diverso.

Gli Sporozoi.
Gli sporozoi sono protozoi esclusivamente parassiti, il
cui nome deriva dalla presenza, almeno in alcune
specie, di stadi infettivi simili a spore. Gli sporozoi
differiscono dalla maggior parte degli altri protisti per la
mancanza di vacuoli contrattili; in questo caso
l'ingresso di acqua è limitato dalla rigidità della pellicola
esterna, che impedisce al tempo stesso un eccessivo
rigonfiamento della cellula. Gli sporozoi hanno
generalmente una forma ameboide, ma ciò non indica
in alcun modo una relazione con i Sarcodina, quanto
piuttosto un caso di convergenza evolutiva.
Al pari di molti altri parassiti obbligatori, gli sporozoi
possiedono cicli vitali molto complessi, in cui si
alternano stadi di riproduzione sessuale e asessuale,
spesso legati a ospiti diversi.
Fra i più noti sporozoi vi sono senza dubbio i parassiti
del genere Plasmodium (▶figura 28), un gruppo
altamente specializzato che conduce parte del proprio
ciclo vitale all'interno delle cellule ematiche dell'uomo
causando la malaria, una delle malattie più diffuse a
livello mondiale.
Figura 28. Plasmodium, il plasmodio della malaria.
Plasmodium è l‟agente eziologico della malaria.
Il vettore di questo parassita è rappresentato dalle
femmine di zanzara del genere Anopheles (▶figura 29)
che, in seguito alla puntura, immettono i plasmodi allo
stadio di sporozoiti nel circolo sanguigno dell'uomo.
Figura 29. Anopheles, la zanzara anofele.
Questo genere di zanzara è il vettore che trasmette il plasmodio da una persona malata ad una sana, infettandola.
Una volta giunte nel torrente circolatorio, le cellule
parassite si spostano rapidamente nel fegato e nel
sistema linfatico, dove si moltiplicano ripetutamente e
assumono la forma di merozoiti. Successivamente i
merozoiti tornano nel sistema circolatorio infestando i
globuli rossi. All'interno di questi ultimi i parassiti si
moltiplicano per circa due giorni, producendo ognuno
fino a 36 nuovi plasmodi e causando la rottura del
globulo rosso con conseguente rilascio dei nuovi
parassiti, pronti ad attaccare altre cellule.
Se un'altra Anopheles punge l'individuo malarico in
questo stadio, alcune cellule di plasmodio possono
essere risucchiate insieme al sangue e infettare la
zanzara. A questo punto del ciclo i parassiti attaccano le
cellule intestinali della zanzara, si riproducono e
successivamente si portano nelle ghiandole salivari, da
cui possono essere trasmessi a un altro ospite.
Figura 30. Ciclo vitale di Plasmodium.
Le specie di Plasmodium che causano la malaria trascorrono parte del loro ciclo vitale nell'uomo e parte nella zanzara. I gametociti maschili e femminili, provenienti dal sangue umano, si trasformano nell'intestino della zanzara in gameti maschili e femminili. Lo zigote che si origina dalla fusione di due gameti si insinua fra le cellule dell'intestino formando una ciste. Questa può dividersi e dare origine a numerose cellule dette sporozoiti, che invadono le ghiandole salivari della zanzara. Quando la zanzara punge nuovamente una persona, gli sporozoiti vengono immessi nel circolo sanguigno di quest'ultima. Da qui essi raggiungono le cellule del fegato e del sistema linfatico, in cui si dividono e formano un nuovo tipo di cellule, i merozoiti. Questi possono invadere nuove cellule epatiche o del sistema linfatico, formando una nuova generazione di merozoiti, oppure possono penetrare nei globuli rossi. La moltiplicazione e l'accrescimento dei merozoiti causano la rottura dei globuli rossi e la liberazione delle cellule di Plasmodium, che invadono altre emazie con un ciclo che si ripete ogni 48 ore. All'interno dei globuli rossi, alcuni merozoiti si sviluppano poi in gametociti maschili e femminili, capaci di ricominciare il ciclo all'interno di una zanzara.
Il ciclo vitale del Plasmodium (▶figura 30) può essere
interrotto bonificando le aree paludose in cui le zanzare
si riproducono. L'uso di insetticidi può inoltre rivelarsi
utile per ridurre drasticamente le popolazioni di
Anopheles, ma prima di adottare tale metodo occorre
valutare attentamente tutti i possibili rischi ecologici e
sanitari.

La malaria si manifesta con forte febbre intermittente,
brividi e irrigidimento muscolare, epatosplenomegalia
(aumento del volume di fegato e milza); tachicardia
(aumento della frequenza del battito cardiaco) e delirio,
spesso compaiono in concomitanza con eventi febbrili
acuti. La diagnosi si effettua sottoponendo a indagine
emoscopica un campione di sangue prelevato dai
capillari periferici (polpastrello o lobo auricolare) ove i
plasmodi tendono a concentrarsi maggiormente.
Purtroppo non esistono vaccini e i farmaci non sono
molto efficaci. L‟unico rimedio risulta ancora oggi il
chinino (▶figura 31) che si estrae dalla corteccia di una
pianta tropicale, la Cinchona (▶figura 32).
Figura 31. Formula di struttura del chinino.
Questo alcaloide rappresenta il primo farmaco antimalarico ed ancora oggi rappresenta il principale rimedio nella lotta contro la malaria.
Figura 32. Cinchona.
E‟ il genere di piante tropicali originario delle Ande, comunemente nota con il nome di china. Dalla sua corteccia si estrae il chinino.
Un‟altra specie patogena appartenente al gruppo degli
sporozoi è Toxoplasma gondii parassita intracellulare
obbligato diffuso tra i mammiferi e gli uccelli che
provoca la toxoplasmosi (▶figura 33).
Il Toxoplasma vive in genere nel tratto intestinale
del gatto, che si infetta cibandosi di carne di piccoli
roditori. Esso rappresenta l'animale serbatoio per la
riproduzione del protozoo, in quanto nel suo intestino il
toxoplasma svolge il suo ciclo di riproduzione sessuata.
Le oocisti vengono emesse con le feci del gatto e
possono essere ingerite da un altro animale o
rarissimamente dall'uomo, i quali rappresentano quindi
i suoi ospiti intermedi. La coabitazione tra gatti e uomo
non è un fattore di rischio importante per l'infezione di
Toxoplasma, in quanto il gatto elimina le oocisti solo
per 1-2 settimane in seguito all'infezione, e le stesse
non sono ancora infettanti, poiché sporulano nel
terreno nell'arco di 1-5 giorni; quindi con una regolare
pulizia della lettiera e dell'ambiente si annulla il rischio.
Molto più pericoloso, è cibarsi di carni crude o poco
cotte (specie di agnello e suino), di insaccati, di verdure
lavate male o di latticini non pastorizzati. Nell'uomo la
toxoplasmosi può manifestarsi in forma acquisita o
connatale. L'infezione può causare schizofrenia.
Figura 33. Toxoplasma gondii. E‟ lo il genere di piante tropicali originario delle Ande, comunemente
Il ciclo vitale di Toxoplasma gondii è rappresentato
nella (▶figura 34).
Figura 34. Ciclo vitale di Toxoplasma gondii. Il ciclo vitale del Toxoplasma gondii ha due fasi. La prima avviene nell'ospite definitivo, un felino e comprende la riproduzione sessuata:il felide, ad esempio un gatto, si infetta ingerendo carne contenente cisti del parassita oppure oocisti sporulate. La seconda fase, nella quale il parassita si riproduce solo in maniera asessuata può aver luogo in ogni animale a sangue caldo, mammiferi (incluso lo stesso gatto) o uccelli. Anche questi ospiti intermedi si possono infettare o da oocisti sporulate presenti nelle feci o dal consumo di carne cruda o poco cotta di animali parassitati: il parassita passa la barriera e invade per via ematogena cellule di svariati tessuti. Di norma dopo questa prima fase l'ospite acquisisce una certa immunità perché gli anticorpi prodotti limitano l'invasività.

Gimnomiceti.
II phylum Gimnomiceti è costituito da due diversi gruppi
di muffe mucillaginose: quelle acellulari (classe
Mixomiceti), dette anche plasmodiali, e quelle a
struttura cellulare (classe Acrasiomiceti). I due
raggruppamenti mostrano tuttavia alcune caratteristiche
comuni: gli individui sono mobili, ingeriscono per
endocitosi cibo particolato e formano spore su corpi
fruttiferi eretti.
Nel corso del loro ciclo vitale, inoltre, essi vanno
incontro a sorprendenti modifiche strutturali, che
prevedono uno stadio formato da cellule isolate che si
nutrono per assorbimento. Alcune di queste muffe
possono coprire una superficie di oltre un metro di
diametro e raggiungere un peso di oltre 50 grammi.
Individui di entrambe le classi si ritrovano
principalmente in ambienti freschi e umidi come le
foreste. Essi possono inoltre apparire come strutture
incolori oppure di color arancio o giallo brillante.
Mixomiceti (muffe mucillaginose acellulari)
La forma acellulare delle muffe mucillaginose è
costituita da una massa citoplasmatica priva di parete e
contenente numerosi nuclei diploidi, che si estende sul
substrato formando una fitta rete di filamenti e viene
definita plasmodio. Il plasmodio dei mixomiceti è un
esempio di cenocito, ovvero una struttura
comprendente molti nuclei, delimitata da una sola
membrana plasmatica. Il citoplasma esterno del
plasmodio, di regola in uno stato meno fluido di quello
interno, garantisce invece una certa rigidità alla
struttura plasmodiale. Alcuni mixomiceti, come ad
esempio quelli del genere Physarum (▶figura 35),
forniscono un interessante esempio di plasticità
citoplasmatica.
Le regioni citoplasmatiche esterne possono infatti
fluidificarsi, con conseguente flusso di citoplasma verso
le aree periferiche e slittamento della massa
plasmodiale in una determinata direzione.
Questo movimento può talvolta cambiare direzione in
seguito allo spostamento del citoplasma in una diversa
porzione cellulare.
Figura 35. I funghi mucillaginosi plasmodiali
(A) I plasmodi di questo Physarum ricoprono una roccia. (B) I caratteristici corpi fruttiferi di Physarum (ingrandimento maggiore).
Mentre si espande sul substrato, il plasmodio ingloba
particelle di cibo, in prevalenza batteri, lieviti, spore di
funghi e resti di piante e animali in decomposizione.
Al processo di diffusione del plasmodio contribuisce
anche una proteina contrattile, la mixo-miosina,
organizzata in microfilamenti che, insieme a molecole
simili a quelle della miosina, determinano il movimento.
Allo stadio plasmodiale, le muffe possono accrescersi
quasi indefinitamente, almeno finché esistono adeguate
riserve di cibo e finché condizioni ambientali quali
umidità e pH permangono favorevoli.
3. Protisti simili ai funghi.
I protisti simili a funghi sono eucarioti unicellulari o plurinucleati, eterotrofi per assorbimento, generalmente privi di chitina e
mancanti di fase dicariotica. Come è stato precedentemente accennato, questi protisti differiscono dai funghi per:
· la presenza di flagelli (talvolta questi sono presenti solamente in alcuni stadi del loro ciclo vitale);
· per la presenza di gameti maschili e femminili morfologicamente diversi;
· per la mancanza di chitina nella parete cellulare;
· per l„assenza di uno stadio dicariotico (ovvero di uno stadio in cui in una singola cellula sono presenti due nuclei
geneticamente distinti);
I protisti simili ai funghi vengono suddivisi in due phyla :
Gimnomiceti funghi mucillaginosi
Oomiceti muffe d‟acqua

Quando invece le condizioni esterne cambiano, possono
verificarsi due diverse situazioni. Il plasmodio può
formare una struttura di resistenza, lo sclerozio,
consistente in una massa irregolare di strutture indurite
simili a cellule, capace di trasformarsi nuovamente in un
plasmodio qualora le condizioni tornino a essere
favorevoli. In alternativa, il plasmodio può trasformarsi
in una struttura fruttifera capace di produrre spore, lo
sporangioforo. La rigidità di queste strutture
peduncolate o ramificate è dovuta alla deposizione di
cellulosa o chitina alla superficie delle cellule;
componenti che sono invece assenti nelle altre fasi del
ciclo vitale.
I nuclei dei plasmodi sono diploidi e si dividono per
meiosi durante lo sviluppo degli sporangiofori. Nella
parte apicale di questi ultimi si sviluppano in seguito
una o più protuberanze di varia forma e colore, dette
sporangi, all'interno dei quali i nuclei aploidi vengono
rivestiti da una parete rigida e si trasformano in spore.
Se lo sporangioforo si dissecca, le spore vengono
liberate nell'ambiente esterno e, germinando, danno
origine a cellule aploidi, flagellate e prive di parete,
capaci di moltiplicarsi mitoticamente o fungere da
gameti. Se le condizioni ambientali non sono favorevoli,
queste cellule possono incistarsi formando una nuova
parete e funzionare così da forme di resistenza.
Al manifestarsi di condizioni favorevoli, le cisti si
aprono e rilasciano le cellule che, fondendosi due a due,
formano uno zigote diploide. Da quest'ultimo, per
successive divisioni mitotiche del nucleo non seguite da
divisioni del citoplasma, può infine formarsi un nuovo
plasmodio (▶figura 36).
Figura 36. Ciclo vitale di un mixomicete.
Il ciclo vitale dei mixomiceti è molto complesso e prevede straordinarie trasformazioni.
Acrasiomiceti (muffe mucillaginose cellulari)
L'altro gruppo di Gymnomycota è composto da muffe
con struttura cellulare, la cui forma vegetativa principale
è rappresentata da una cellula ameboide (detta
mixoameba) (▶figura 37).
Essa è dotata di un singolo nucleo aploide, si ciba
inglobando batteri e altre particelle alimentari e si
riproduce per mitosi o scissione. Gruppi di cellule
isolate e indipendenti possono sopravvivere finché le
condizioni esterne di umidità e disponibilità di cibo
permangono favorevoli.
Figura 37. I funghi mucillaginosi cellulari.
Il ciclo vitale del fungo Dictyostelium illustrato in una composizione fotografica (lo stesso esemplare è stato fotografato nei vari stadi di maturazione, e poi gli scatti sono stati montati in sequenza).
Quando le condizioni diventano sfavorevoli, queste
cellule sono in grado di aggregarsi e formare una
struttura fruttifera detta pseudoplasmodio. Quest'ultimo
differisce dalla vera struttura plasmodiale per il fatto
che le singole cellule mantengono le loro membrane
plasmatiche e quindi la loro identità; esso non può
quindi essere considerato un cenocito. Il segnale
chimico che stimola l'aggregazione delle mixoamebe è
il 3',5'-adenosin monofosfato ciclico (cAMP), un
«messaggero» che esercita un ruolo importante anche
in certi processi metabolici degli animali.
Uno pseudoplasmodio può spostarsi sul substrato per
diverse ore prima di dare origine a una delicata
struttura fruttifera peduncolata. Le cellule poste
all'estremità del corpo fruttifero si sviluppano in spore,
dotate di una parete sottile, che in seguito si staccano.
Quando le condizioni ambientali tornano favorevoli, le
spore germinano e danno origine a nuove mixoamebe.
A questo ciclo di riproduzione asessuale se ne affianca
talora uno sessuale, in cui due mixoamebe
(possibilmente di due tipi sessuali diversi) si fondono
formando una struttura sferica che successivamente
germina, rilasciando nuove mixoamebe.
Il ciclo vitale di un tipico acrasiomicete è descritto nella
(▶figura 38).
Figura 38. Ciclo vitale di un acrasiomicete.

Oomiceti.
Il phylum degli Oomiceti è costituito da diversi protisti
eterotrofi e comprende le muffe d‟acqua ed altri
organismi terrestri.
Le muffe d‟acqua sono filamentose e immobili e si
nutrono per assorbimento, cioè secernono sostanze che
digeriscono le grandi molecole di nutrienti scindendole
in molecole più piccole, facili da assorbire.
Una caratteristica distintiva di tutti gli Oomiceti è la
presenza di flagelli nelle cellule riproduttive e di pareti
cellulari contenenti cellulosa. Alcune specie presentano
più nuclei racchiusi in una sola membrana plasmatica: i
filamenti che costituiscono l‟oomicete, quindi, non sono
separati internamente da pareti che confinino ciascun
nucleo in una singola cellula. In questo modo, il
citoplasma è un fluido continuo che fluisce nell‟intero
corpo dell‟organismo. Gli Oomiceti sono generalmente
diploidi per gran parte del loro ciclo vitale.
Muffe d‟acqua comuni sono quelle del
genere Saprolegnia (▶figura 39), acquatiche e
saprofaghe, che rivestono di un feltro biancastro pesci o
insetti morti nell‟acqua (▶figura 40). Il termine comune
di «muffa», che deriva dall‟iniziale classificazione di tali
forme nel regno dei funghi, non deve portare a
confondere questi organismi con i veri funghi che
portano questo nome.
Figura 39. Saprolegnia parasitica.
Un particolare che evidenzia le spore sessuali (oospore) prodotte all‟interno di strutture dette oogoni.
Figura 40. Saprolegnia, un oomicete saprofita.
Micelio della muffa d'acqua Saprolegnia che cresce sui resti di un pesce.
Gli Oomiceti terrestri sono in genere decompositori
innocui o utili, ma alcuni provocano malattie nelle
piante coltivate. Ad esempio Plasmopara viticola è
l‟agente eziologico della peronospora della vite, una
delle principali malattie di questa pianta (▶figura 41).
Figura 41. Peronospora della vite.
La peronospora della vite, Plasmopara viticola, è un microrganismo originario dell‟America e importato accidentalmente in Francia intorno al 1878, da cui si è poi diffuso in tutta Europa. Esso provoca una tipica malattia della vite, con un ciclo fortemente condizionato dalle condizioni climatiche. Si tratta della causa di una delle più diffuse e pericolose malattie della vite in molte regioni europee ed italiane.
La muffa Phytophthora infestans, è invece la causa della
peronospora della patata, e fu responsabile della
distruzione dell'intero raccolto di patate, con
conseguente carestia, nel 1845-1847 in Irlanda,
carestia che causò la morte per fame di almeno un
milione di persone e ne costrinse all'emigrazione
almeno altri due (▶figura 42).
Figura 42. Peronospora della patata e del pomodoro.
La peronospora della patata e del pomodoro, Phytophthora infestans, è un microrganismo che colpisce le piante della famiglia delle Solanacecee.

Euglenofite.
Per molti zoologi questa divisione comprende forme che
fanno parte del phylum dei Flagellati. I pigmenti
fotosintetici sono clorofilla a e clorofilla b, proprio come
nelle alghe verdi (Clorofite) e in tutte le piante.
Il più tipico rappresentante di questo gruppo di
organismi è Euglena (▶figura 43)
Figura 43. Euglena. Euglena, una specie che può essere considerata sia protozoo, sia alga. Nella fotografia si notano i numerosi cloroplasti di Euglena.
Euglena è una specie che mostra quanto sia difficile
separare in modo netto le forme autotrofe da quelle
eterotrofe.
Esso si sposta grazie al movimento di uno dei due
flagelli di cui è provvisto, e che fornisce la spinta
necessaria tramite un movimento ondulatorio. Euglena
si riproduce per mitosi e citodieresi, ovvero tramite la
modalità più semplice. Da un punto di vista trofico,
Euglena mostra un'estrema flessibilità. In piena luce si
comporta da organismo autotrofo, e impiega i propri
cloroplasti per sintetizzare composti organici tramite la
fotosintesi. Se invece viene mantenuto nell'oscurità,
esso perde i pigmenti fotosintetici e inizia a nutrirsi
esclusivamente di materia organica morta.
Tale processo è tuttavia completamente reversibile
poiché, se riportato alla luce, Euglena è capace di
ripristinare i propri pigmenti e di tornare autotrofo.
È stato inoltre osservato che cellule di Euglena trattate
con certi antibiotici o sostanze mutagene perdono
completamente i pigmenti fotosintetici divenendo
incapaci, anche nelle generazioni successive, di
effettuare la fotosintesi. Le cellule che ne derivano
continuano in ogni caso a sopravvivere come eterotrofi.
4. Le alghe: protisti simili alle piante.
Le alghe sono eucarioti unicellulari, plurinucleati o pluricellulari, fotosintetici. Le forme pluricellulari hanno embrioni privi di
tessuti materni.
Le alghe sono degli organismi fotosintetici dai quali probabilmente dipende il 50-60% della fotosintesi che si compie sull'intero
pianeta. La rimanente parte può essere attribuita alle piante, mentre il contributo dei cianobatteri e di altri batteri fotosintetici
è trascurabile, sebbene esso possa risultare importante in alcuni ecosistemi acquatici.
La principale differenza tra piante e alghe consiste nel fatto che nelle prime lo zigote da origine a un embrione pluricellulare
protetto dai tessuti dell'individuo in cui si origina, mentre nelle seconde ciò non si verifica.
Le alghe mostrano un'interessante varietà di forme. Alcune sono organismi unicellulari relativamente semplici, altre invece
formano filamenti costituiti da cellule distinte o da strutture multinucleate (cenociti), e altre ancora sono pluricellulari,
caratterizzate da complesse ramificazioni oppure a forma di foglia e con tessuti e organi ben distinti. Questa varietà di forme
può essere presente anche all'interno dei singoli raggruppamenti di alghe, come ad esempio in quello delle alghe verdi
(phylum Chlorophyta). Anche i cicli vitali delle alghe sono estremamente vari, e tutte, tranne le alghe rosse (phylum
Rhodophyta), possiedono uno stadio con cellule flagellate mobili. In alcuni casi, ad esempio nei dinoflagellati (phylum
Pyrrophyta), questa è addirittura la forma prevalente. Eccetto che per il mannitolo, i grassi e gli olii, tutti i composti di riserva
delle alghe sono polimeri del glucosio, che differiscono gli uni dagli altri per il tipo di legame fra le molecole di glucosio
adiacenti, per il grado di ramificazione delle catene polisaccaridiche e per le dimensioni della catena. Le sostanze pectiche che
si trovano nelle pareti cellulari sono tutte polimeri di uno zucchero acido, l'acido galatturonico.
Le alghe vengono suddivise in sei phyla :
Euglenofite euglene e forme affini
Pirrofite dinoflagellati
Crisofite diatomee
Feofite alghe brune
Rodofite alghe rosse
Clorofite alghe verdi

Pirrofite.
Le Pirrofite sono tutte alghe unicellulari con una
caratteristica colorazione giallo-bruna, data dai
pigmenti fotosintetici e dai pigmenti accessori presenti
nei cloroplasti. Possiedono clorofilla a e clorofilla c.
I dinoflagellati, il gruppo più rappresentativo di questo
phylum, sono di grande interesse ecologico, evolutivo e
morfologico, e sono probabilmente secondi soltanto
alle diatomee (appartenenti al phylum delle
Chrysophyta) come produttori fotosintetici di materia
organica negli oceani.
Molti dinoflagellati sono endosimbionti di altri protisti e
invertebrati marini, come ad esempio i coralli, alla cui
crescita contribuiscono con i loro prodotti fotosintetici.
Altri, invece, sono incapaci di fotosintetizzare e vivono
come parassiti di altri organismi marini.
La struttura cellulare dei dinoflagellati è particolarmente
interessante. Questi organismi possiedono due flagelli,
di cui uno si trova alloggiato in un solco equatoriale
intorno alla cellula e l'altro passa attraverso un solco
longitudinale prima di protendersi libero verso l'esterno
(▶figura 44).
Figura 44. Dinoflagellati.
Ceratium e Peridinium, due tipici dinoflagellati in cui è possibile notare il flagello assiale e quello equatoriale..
Molti dinoflagellati sono organismi marini e alcuni di
questi, che si riproducono abbondantemente nelle
acque calde e tranquille, possono dare origine alle
«maree rosse» (▶figura 45), così dette per la
colorazione rossastra assunta dalle acque superficiali a
causa della fluorescenza della clorofilla presente nei
cloroplasti.
Figura 45. Maree rosse.
Alcuni dinoflagellati sono responsabili del fenomeno delle maree rosse.
Alcune di queste specie producono inoltre sostanze
neurotossiche, che sono in grado di uccidere migliaia di
pesci. Anche il genere Gonyaulax emette una potente
tossina, che può accumularsi senza alcun effetto nei
molluschi e nei crostacei, ma può uccidere un uomo che
se ne cibi.
Molti dinoflagellati sono inoltre bioluminescenti
(▶figura 46); colture di questi organismi mantenute in
completa oscurità emettono un debole scintillio. Se poi
la coltura viene disturbata, agitando il mezzo di
sospensione o facendovi gorgogliare dell'aria, ogni
individuo può emettere lampi luminosi, anche un
migliaio di volte più potenti della debole luminosità
emessa inizialmente.
Figura 46. Bioluminescenza.
Molti dinoflagellati sono in grado di emettere luce grazie allo stesso meccanismo che viene utilizzato da altri organismi (come ad esempio, le lucciole): un enzima, definito luciferasi, ossida una sostanza detta luciferina e tale reazione chimica libera energia sotto forma luce visibile all‟occhio umano.

Crisofite (Alghe giallo-dorate).
II phylum delle Crisofite comprende diatomee e altri
organismi a esse simili. Gli individui di alcune specie
sono unicellulari, mentre altri possono formare
aggregati filamentosi.
Possiedono sia clorofilla a che clorofilla c. Molte hanno
un colore bruno o giallo per la presenza di abbondanti
carotenoidi nei loro cloroplasti e tutte utilizzano
crisolaminarina (un carboidrato) od olii come prodotti di
riserva.
Molte diatomee depositano silice (diossido di silicio
SiO2) nelle loro pareti cellulari, che in alcune specie
sono formate da due parti distinte (valve), con quella
superiore che si sovrappone all'inferiore come un
coperchio su una scatola. Le diatomee possono avere
sia simmetria radiale che bilaterale.
Questi organismi, marini o d'acqua dolce, sono
caratterizzati da un'estrema varietà di forme e la loro
tassonomia è interamente basata sulle caratteristiche
delle pareti esterne (▶figura 47).
Figura 47. Diatomee. Le diatomee mostrano una grande varietà di forme, di regola specie-specifiche.
Le diatomee possono riprodursi sia sessualmente che
asessualmente (▶figura 48).
Nel caso della riproduzione asessuale, si verifica una
divisione cellulare in cui ciascuna delle due cellule figlie
eredita una delle due valve della cellula madre. È
interessante notare che la parte mancante viene sempre
riformata all'interno di quella già esistente. Questo
processo è quindi caratterizzato da una progressiva
diminuzione delle dimensioni dei nuovi individui a ogni
evento riproduttivo, e se il processo si ripetesse sempre
allo stesso modo, si arriverebbe alla scomparsa di
questi organismi.
La riproduzione sessuale risolve adeguatamente il
problema: i gameti che ogni individuo produce sono
privi di parete e si fondono formando uno zigote che,
prima di costituire una nuova parete cellulare, aumenta
notevolmente le proprie dimensioni e per questo esso
viene anche definito auxospora, dal greco auxesis =
aumento.
Figura 48. La riproduzione delle diatomee. Nella riproduzione asessuale per mitosi e citodieresi (sequenza verticale sulla destra nella figura), ogni cellula figlia eredita una delle due valve; ogni cellula dovrà quindi produrre una nuova valva e quest'ultima si forma sempre all'interno di quella già esistente. In seguito, si verifica una progressiva riduzione dimensionale degli individui, interrotta dalla riproduzione sessuale: lo zigote (ciclo sulla sinistra), si accresce prima di ricostituire il proprio rivestimento.
Le diatomee sono diffuse ovunque nell„ambiente marino
e di acqua dolce.
I resti delle loro pareti contenenti silicio, molto
resistenti alla decomposizione, possono accumularsi sui
fondi oceanici e formare estesi giacimenti di rocce
sedimentarie (diatomiti). La così detta terra di diatomee,
ottenuta da queste rocce, trova numerosi impieghi
industriali, come materiale per l'isolamento o la
filtrazione e per la pulitura dei metalli.
I dinoflagellati (Pirrofite) e le diatomee (crisofite) sono i
principali responsabili del fenomeno di
“eutrofizzazione” delle acque dei mari, dei laghi e dei
fiumi che si verifica quando le concentrazione di sali
minerali (contenenti N, P e K) raggiunge valori elevati. In
questo caso, la proliferazione di queste alghe
unicellulari, non seguita dallo smaltimento dei
consumatori primari, determina una maggiore attività
batterica, questo fatto induce un aumento del consumo
di ossigeno, e la mancanza di quest‟ultimo provoca a
sua volta la moria di animali marini (ad esempio dei
pesci).

Feofite (Alghe brune).
Le feofite, o alghe brune, sono organismi pluricellulari,
composti da filamenti ramificati o da strutture fogliose
definite talli . Il caratteristico colore, da cui deriva il loro
nome, è determinato dalla presenza del carotenoide
fucoxantina, presente in grandi quantità nei plastidi. La
combinazione di questo pigmento giallo-arancio con il
verde della clorofilla a e c determina il colore.
Le feofite (▶figura 49) includono alcune fra le alghe di
maggiori dimensioni (come ad esempio quelle del
genere Macrocystis che può raggiungere la lunghezza di
60 m). Si tratta di forme quasi esclusivamente marine,
alcune delle quali galleggiano in mare aperto (l'esempio
più noto è costituito dal Sargassum, che può costituire
densi banchi nel mar dei Sargassi), mentre altre vivono
attaccate alle rocce nelle regioni costiere. In queste
ultime forme è presente, nella parte a contatto con il
substrato, una struttura espansa a ventosa detta aptere,
che consente una salda adesione.
Figura 49. Alghe brune.
(A) La specie Pelvetia canaliculata mostra un esempio della struttura filamentosa delle alghe brune. (B) I filamenti dell‟alga microscopica Ectocarpus osservati al microscopio ottico. (C) Le palme di mare sono caratterizzate dalla presenza di fronde simili a «foglie». (D) Le palme di mare e molte altre specie di alghe brune aderiscono al substrato per mezzo di robuste strutture di ancoraggio.
Alcune alghe brune possono formare steli simili a
tronchi oppure lamine simili a foglie, mentre altre sono
caratterizzate da cavità o vesciche piene di gas.
Per ragioni non ancora ben chiare, queste cavità
contengono spesso monossido di carbonio (circa il 5%
del contenuto) in quantità sufficiente a uccidere un
uomo.
Oltre a questa organizzazione in organi, le specie di
maggiori dimensioni mostrano anche un notevole grado
di differenziazione dei loro tessuti. Spesso i filamenti
fotosintetici sono localizzati nelle regioni più esterne
dell'alga, mentre all'interno del tessuto fotosintetico
sono localizzati filamenti di cellule allungate simili ai
tessuti di conduzione delle piante superiori. Questi
filamenti, formati da cellule svasate all'apice, formano
strutture canalicolari che trasportano rapidamente i
prodotti della fotosintesi (principalmente mannitolo)
attraverso il corpo dell'alga.
Il phylum Phaeophyta testimonia lo straordinario grado
di diversità che caratterizza le alghe. Fra le alghe brune
a struttura più semplice possiamo citare quelle del
genere Ectocarpus, che formano filamenti ramificati,
lunghi pochi centimetri, che crescono comunemente
sulle conchiglie e sulle pietre sommerse. In questo caso
il gametofito e lo sporofito possono essere distinti
esclusivamente in base al numero di cromosomi o in
base ai loro prodotti (rispettivamente zoospore o
gameti); si hanno cioè generazioni isomorfe.
Altre alghe brune di maggiore complessità, come ad
esempio le Laminaria (▶figura 50), sono caratterizzate
da un'alternanza di generazioni di tipo eteromorfico, in
cui la fase predominante è lo sporofito. Il gametofito,
una struttura filamentosa e sottile, si origina invece da
zoospore formatesi per meiosi in particolari regioni
dello sporofito. Dal gametofito deriveranno poi gameti
maschili e femminili.
Figura 50. Ciclo vitale di un‟alga bruna. Laminaria è un‟alga bruna che presenta un ciclo vitale aplo-diplobionte con alternanza di generazioni eteromorfiche.
Nel genere Fucus (▶figura 51) si assiste infine a
un'ulteriore riduzione del gametofito: non esiste una
fase aploide pluricellulare, ma soltanto una fase aploide
multinucleata, in cui i gameti sono prodotti
direttamente per meiosi.
Figura 51. Un‟alga bruna. Fucus, una comune alga bruna.
Le pareti cellulari delle alghe brune possono contenere
fino al 25% di acido alginico, un polimero di uno
zucchero acido, piuttosto viscoso, capace di cementare
cellule e filamenti e dunque in grado di funzionare
come aptere. Questa sostanza viene impiegata
commercialmente come emulsionante in gelati,
cosmetici e altri prodotti.

Rodofite (Alghe rosse).
Quasi tutte le alghe rosse o Rodofite sono organismi
pluricellulari. Il loro caratteristico colore rosso deriva
dalla presenza del pigmento ficoeritrina nei cloroplasti.
In aggiunta a tale sostanza, nelle alghe rosse sono
presenti anche ficocianina, carotenoidi e clorofilla a.
La maggior parte delle alghe rosse vive in ambiente
marino e colonizza tutti gli habitat, dalle pozze di
marea fino a profondità notevoli (anche 170 m se
esistono nutrienti in quantità sufficiente e l'acqua è
limpida abbastanza da consentire la penetrazione della
luce). Gran parte delle specie vivono attaccate al
substrato e sono provviste di aptere (▶figura 52).
Figura 52. Alghe rosse. Due diverse specie di Rodofite che presentano la loro tipica colorazione dovuta a specifici pigmenti fotosintetici.
Le alghe rosse hanno la capacità di cambiare la
composizione relativa dei loro pigmenti fotosintetici in
relazione alle condizioni di illuminazione, con
conseguente cambiamento di colorazione: la stessa
specie può apparire verde brillante se cresciuta in
superficie, oppure rosso scuro se cresciuta in acque più
profonde. In acque profonde, infatti, la maggior parte
della luce che riesce a penetrare, e che può quindi
essere impiegata per la fotosintesi, appartiene alla
banda blu-verde dello spettro luminoso e quindi le
alghe accumulano grandi quantità di ficoeritrina, il
pigmento capace di assorbire tali lunghezze d'onda.
Così, anche se la quantità di clorofilla è uguale a quella
contenuta nelle alghe che vivono in superficie, la
quantità di ficoeritrina accumulata fa apparire queste
alghe di colore rosso anziché verde. La variazione della
pigmentazione in dipendenza della lunghezza d'onda
della luce viene indicata come adattamento cromatico
(metacromatismo) (▶figura 53).
Figura 53. L'assorbimento della luce da parte di un'alga rossa.
Quando l'alga rossa Chondrus crispus cresce al di sotto dei 15 m di profondità, essa accumula ficoeritrina con cui può assorbire la luce blu-verde. In superficie, invece, l'accumulo di ficoeritrina è molto minore e l'alga assorbe principalmente nel rosso, tramite la clorofilla.
Le alghe rosse accumulano i prodotti della fotosintesi
sotto forma di amido delle floridee (composto da
piccole catene ramificate di 15 unità di glucosio) e non
possiedono cellule flagellate in alcuno stadio del loro
ciclo vitale. I gameti maschili sono privi di parete e
leggermente ameboidi, mentre quelli femminili sono
completamente immobili.
Alcune specie di alghe rosse, capaci di depositare
carbonato di calcio all'interno e all'esterno delle loro
pareti cellulari, contribuiscono alla formazione delle
barriere coralline, poiché, dopo la morte delle cellule, il
carbonato si accumula sui fondali formando talvolta
cospicui ammassi.
Alcune Rhodophyta possono produrre grandi quantità di
sostanze polisaccaridiche mucillaginose, costituite
principalmente da galattosio cui è legato un gruppo
solfato. Questo materiale forma gelatine solide ed è la
materia prima dell'agar, una sostanza assai usata per
produrre substrati solubili in acqua, sui quali far
crescere in laboratorio colture di tessuti oppure
microrganismi.
Alcune alghe rosse marine, infine, sono parassite di
altre alghe. In questo caso l'ospite è un'alga
normalmente fotosintetica mentre il parassita è un'alga
incolore e non fotosintetica, che ottiene le sostanze
nutritive dall'ospite. È stato scoperto che l'alga parassita
Choreocolax è capace di inserire i propri nuclei
all'interno delle cellule dell'alga rossa ospite
Polysiphonia, modificandone il metabolismo. Questo è il
primo esempio di un processo di trasferimento di nuclei
dal parassita alle cellule viventi dell'ospite.

Clorofite (Alghe verdi).
Le Clorofite, conosciute comunemente come alghe
verdi, e le Euglenofite sono gli unici protisti che
possiedono l'intero corredo di pigmenti fotosintetici che
caratterizzano le piante superiori. Il pigmento principale
è la clorofilla a, ma al loro interno è presente anche la
clorofilla b. I carotenoidi presenti in questi gruppi sono
prevalentemente 3-carotene e alcune xantofille
(carotenoidi con uno o più gruppi ossidrilici). Il
principale prodotto di accumulo della fotosintesi è,
come nelle piante superiori, l'amido, costituito da
lunghe catene, talora ramificate, di glucosio. Queste
somiglianze, insieme a considerazioni di altro tipo,
hanno indotto a ipotizzare che le piante si siano evolute
dalle alghe verdi.
Questa uniformità dei pigmenti e dei prodotti di
accumulo della fotosintesi è contrapposta a un'estrema
varietà di forme (da ▶figura 54 a ▶figura 63):
All'interno della divisione Clorofite si possono osservare
tutti i termini di passaggio tra organismi unicellulari e
pluricellulari.
Lo stadio tipicamente unicellulare è rappresentato da
Chlamydomonas e da Chlorella specie che potrebbero
essere anche considerate come protozoi flagellati.
Figura 54. Chlamydomonas. Una tipica alga unicellulare e flagellata, che vive nelle acque dolci.
Figura 55. Chlorella. Un altro organismo unicellulare e flagellato, che presenta un‟organizzazione molto semplice.
Gli stadi intermedi tra lo stato unicellulare di e quello
coloniale sono rappresentati da forme coloniali meno
organizzate, come quelle di Gonium e di Pandorina
(▶figura 56).
Figura 56. Gonium e Pandorina sono forme coloniali meno organizzate. Nel genere Gonium (in alto) le cellule sono tutte delle stesse dimensioni. Nel genere Pandorina (in basso) si assiste ad un inizio di specializzazione e di differenziazione in quanto le cellule anteriori della colonia mostrano macchie oculari di maggiori dimensioni.
Lo stadio tipicamente coloniale è rappresentato da
Volvox, (▶figura 57) che forma colonie
sorprendentemente grandi e ben organizzate. In questo
caso le cellule non sono ancora differenziate in tessuti e
organi, come invece accade nelle piante e negli animali,
ma le colonie mostrano quale potrebbe essere stato il
primo gradino evolutivo verso l'organizzazione
pluricellulare.
Figura 57. Colonie di Volvox. Si osservi la regolare distanza delle cellule e la presenza di co-lonie figlie all'interno della colonia principale. Questi organismi fotosintetici vengono talvolta classificati come alghe verdi, un altro gruppo di protisti.

Altri rappresentanti tipici di alghe verdi che
testimoniano la grande varietà di forme sono
considerate nelle seguenti figure.
Figura 58. Oedogonium. Un un'alga filamentosa con un solo nucleo in ogni cellula.
Figura 59. Cladophora. Un‟alga che mostra un'organizzazione pluricellulare con cellule multinucleate.
Figura 60. Bryopsis Un‟alga tubulare e cenocitica, e forma setti che separano una cellula dall'altra soltanto durante la formazione delle strutture riproduttive.
Figura 61. Acetabularia. Un „alga costituita da una singola cellula gigante mononucleata di forma caratteristica, che diviene plurinucleata soltanto alla fine del processo riproduttivo.
Figura 62. Ulva lactuca. Alga nota comunemente come lattuga di mare, è costituita da una struttura laminare dello spessore di due cellule.
Figura 63. Desmidiacee. Alghe unicellulari caratterizzate da pareti cellulari scolpite in maniera elaborata.

I cicli vitali delle alghe verdi sono caratterizzati da
un'estrema variabilità. Esamineremo a titolo di esempio
due di questi cicli, quello di Ulva lactuca (▶figura 64), e
quello di Ulothrix.
Figura 64. Ciclo vitale di Ulva lactuca. Questa alga verde presenta un ciclo vitale aplo-diplobionte con alternanza di generazioni isomorfiche.
Lo sporofito diploide di Ulva lactuca è costituito da una
«foglia» di alcuni centimetri di diametro, in cui si
differenziano alcune cellule specializzate (sporociti).
Attraverso il processo di meiosi e citocinesi, queste
producono spore aploidi mobili (zoospore), capaci di
nuotare grazie alla presenza di quattro flagelli. Quando
una spora trova un ambiente adatto, essa si fissa, perde
i flagelli, e inizia a dividersi mitoticamente, producendo
un filamento sottile che si sviluppa in un'ampia lamina
dello spessore di due cellule: il gametofito,
morfologicamente identico allo sporofito diploide. Ogni
spora contiene l'informazione genetica per un solo tipo
sessuale, e un gametofito è capace di produrre
esclusivamente gameti maschili o femminili, ma mai
entrambi. I gameti si formano per mitosi all'interno di
una singola cellula (gametangio) e non all'interno di una
struttura pluricellulare specializzata come avviene nei
muschi e nelle piante vascolari. Entrambi i tipi di gameti
possiedono due flagelli (a differenza dei quattro delle
zoospore) e sono mobili. I gameti femminili, tuttavia,
appaiono decisamente più grandi di quelli maschili:
questa condizione in cui i gameti sono
morfologicamente distinguibili è detta anisogamia, e
contrapposta all'isogamia di altre alghe e protisti, in cui
i due tipi di gameti sono morfologicamente simili.
Quando due gameti di tipo diverso si incontrano, essi si
fondono, perdono i flagelli e dallo zigote che ne deriva
si forma, per successive mitosi, un nuovo sporofito. I
gameti che non riescono a incontrarne uno
complementare con cui unirsi possono fissarsi al
substrato, perdere i flagelli e, per mitosi, produrre
nuovi gametofiti; in altre parole i gameti possono
funzionare come zoospore, condizione questa molto
rara. Un ciclo vitale come quello di Ulva, in cui sporofito
e gametofito sono morfologicamente identici, è detto
isomorfo.
In altri protisti, invece, esistono generazioni
eteromorfiche, con uno sporofito diverso dal
gametofito. In una variante del ciclo eteromorfico, il
ciclo aplonte, gametofiti pluricellulari producono gameti
da cui deriva uno zigote con funzioni di sporocito. In
altre parole, questo si divide immediatamente per
meiosi, producendo spore che daranno origine a nuovi
gametofiti. In questo tipo di ciclo, dunque, l'unico
stadio diploide è lo zigote.
Un esempio di ciclo aplonte è fornito dalle alghe del
genere Ulothrix (▶figura 65).
Figura 65. Ciclo vitale di Ulothrix. Questa alga verde presenta un ciclo vitale aplobionte.
Altre alghe possiedono un ciclo diplonte come quello
della maggior parte degli animali. In questo caso gli
sporociti producono direttamente per meiosi i gameti, e
dall'unione di questi si origina uno zigote che formerà
un nuovo sporofito. Negli organismi che hanno un ciclo
vitale di questo tipo tutte le cellule tranne i gameti sono
diploidi. Tra i due estremi rappresentati dal ciclo
aplonte e quello diplonte esistono casi in cui sia lo
sporofito che il gametofito sono pluricellulari, ma una
fase (generalmente lo sporofito) risulta nettamente
predominante sull'altra.
Un caso particolare di anisogamia, conosciuto come
oogamia, è quello di Oedogonium, in cui il gamete
femminile è grande e immobile, mentre quello maschile
è piccolo e flagellato.