© copyright 2015 mandana ehsanipour
TRANSCRIPT

i
© Copyright 2015
Mandana Ehsanipour

ii
Bioconversion of lignocellulosic hydrolysate to acetic acid using Moorella thermoacetica
Mandana Ehsanipour
A thesis
Submitted in partial fulfillment of the
requirements for the degree of
Master of Science
University of Washington
2015
Committee:
Renata Bura
Richard Roy Gustafson
Program Authorized to Offer Degree:
School of Environmental and Forest Resources

iii
University of Washington
Abstract
Bioconversion of lignocellulosic hydrolysate to acetic acid using Moorella thermoacetica
Mandana Ehsanipour
Chair of the Supervisory Committee:
Dr. Renata Bura
School of Environmental and Forest Resources
Abstract
A systematic study of bioconversion of lignocellulosic sugars to acetic acid by Moorella
thermoacetica strain ATCC 39073 was conducted. Four different hydrolysates obtained after
steam pretreatment of lignocellulosic biomass were selected and fermented to acetic acid in batch
fermentations. The bacterial strain (ATCC 39073) can effectively ferment xylose and glucose in
steam exploded hydrolysates from wheat straw, forest residues, switchgrass, and sugarcane straw
to acetic acid. This homoacetogen prefers xylose over glucose. The highest acetic acid yield
obtained was in steam pretreated sugarcane straw hydrolysate and the lowest in forest residues
hydrolysate (71% and 39% of theoretical respectively based on total sugars). This acetogen can
partially consume arabinose, galactose and mannose within 72 h of fermentation. Acetic acid yield
in forest residues was adversely affected by high concentration of arabinose, galactose and
mannose. Glucose to xylose ratio has a direct effect on production yield. It was also observed that
M. thermoacetica can tolerate process derived inhibitory compounds from steam explosion
pretreatment (total phenolics up to 3 g/L and furans up to 0.5 g/L).

iv
Table of Contents
List of Figures ................................................................................................................................ vi
List of Tables ................................................................................................................................ vii
List of abbreviation ...................................................................................................................... viii
Acknowledgements ........................................................................................................................ ix
Chapter 1- Introduction ................................................................................................................... 1
1.1 Bioconversion of lignocellulosic biomass to fuels and chemicals ....................................... 1
1.2 Lignocellulosic biomass....................................................................................................... 1
1.2.1 Cellulose ................................................................................................................... 2
1.2.2 Hemicellulose ........................................................................................................... 2
1.2.3 Lignin structure ......................................................................................................... 3
1.2.4 Extractives and ash ................................................................................................... 3
1.3 Bioconversion process ......................................................................................................... 4
1.3.1 Pretreatment .............................................................................................................. 4
1.3.2 Enzymatic hydrolysis ................................................................................................ 5
1.3.3 Fermentation ............................................................................................................. 6
1.3.4 Inhibitors ................................................................................................................... 6
1.4 Acetic acid ........................................................................................................................... 7
1.5 Moorella thermoacetica ....................................................................................................... 8
1.5.1 Cultivation Media ................................................................................................... 10
1.5.2 Fermentation setups ................................................................................................ 10
1.5.3 Extracted sugars ...................................................................................................... 10
1.6 Research objectives ............................................................................................................ 11
Chapter 2 Fermentation of lignocellulosic sugars to acetic acid by Moorella thermoacetica...... 12
2.1 Introduction ........................................................................................................................ 12
2.2 Materials and methods ....................................................................................................... 15
2.2.1 Moorella Thermoacetica strain (ATCC 39073) ..................................................... 15
2.2.2 Culture media conditions ........................................................................................ 15
2.3 Water-soluble fraction (hydrolysate) ................................................................................. 15
2.4 Overliming ......................................................................................................................... 16
2.5 Fermentation ...................................................................................................................... 16

v
2.5.1 Hydrolysate fermentation........................................................................................ 17
2.5.2 Synthetic sugars fermentation ................................................................................. 17
2.6 HPLC analysis ................................................................................................................... 18
2.6.1 Monomeric sugars ................................................................................................... 18
2.6.2 Acetate, hydroxymethylfurfural and furfural analysis ............................................ 19
2.7 Phenolic measurement ....................................................................................................... 19
2.8 Results and discussion ....................................................................................................... 19
2.9 Conclusion ......................................................................................................................... 25
References ..................................................................................................................................... 26
Chapter 3 – Conclusion and future work ...................................................................................... 32
3.1 Conclusion ......................................................................................................................... 32
3.2 Future work ........................................................................................................................ 33
3.3 Significance........................................................................................................................ 34
References ..................................................................................................................................... 35
Appendices ................................................................................................................................ 41
1 Flask fermentations utilizing inoculant with low cell concentration (0.2 – 4 g/L) ............ 43
2 Flask fermentations utilizing inoculant with high cell concentration (7 g/L) .................... 47
2.1 Flask fermentations - comparison of arabinose, galactose and mannose to glucose ......... 50
2.2 Flask fermentations of synthetic sugars - comparison of CO2 to N2 gases ........................ 51
2.3 Fermentation of CO2 gas as carbon source instead of sugars ............................................ 52
3 pH-controlled batch fermentations of glucose and xylose ................................................. 53

vi
List of Figures
Figure 1. Schematic diagram of fermentation experiments. All lignocellulosic biomass were
steam exploded at 195°C, except for sugarcane straw. Sugarcane straw was steam exploded at
190°C. ........................................................................................................................................... 18
Figure 2. Sugars consumption, acetic acid production and cell concentration during fermentation
of synthetic sugars, including single sugar (plots 1, 2, 3, 4, 5, 6), double sugars (plot 7) and
mixture of sugars (plot 8) .............................................................................................................. 45
Figure 3. Sugars consumption, acetic acid production and cell concentration during fermentation
of synthetic sugars including single sugar (plots 1, 2), mixture of different ratio of xylose and
glucose (plots 3, 4, 5, 6, 7, 8, 9) .................................................................................................... 48
Figure 4. Sugars consumption and acetic acid production during flask fermentation utilizing
synthetic sugars model for sugarcane straw hydrolysate comparison of N2 and CO2 gases ........ 52
Figure 5. Acetic acid production during flask fermentation of synthetic medium without sugar 53
Figure 6. Sugars consumption and acetic acid production during batch fermentation of synthetic
glucose and xylose in pH-controlled fermenter including various ratio of glucose to xylose (plots
1, 2, 3, 4), and different glucose concentration (plots 5, 6), effect of glucose concentration on
sugar uptake and acetate product (plot 7) ..................................................................................... 55

vii
List of Tables
Table 1. Composition of different lignocellulosic feedstock .......................................................... 2
Table 2. Summary of process variables during pH-controlled batch fermentation of hydrolysates
and controls ................................................................................................................................... 30
Table 3. Summary of process variables during flask fermentation of single sugar to acetic acid,
comparing arabinose, galactose and mannose to glucose during 50 h fermentation .................... 30
Table 4. Summary of process variables during flask fermentation of mixed sugars, monitoring
consumed sugars and produced acetic acid .................................................................................. 31
Table 5. Summary of process variables during pH-controlled batch fermentation of glucose and
xylose mixture, with different concentration, over 50 h , 75 h and 96 h of fermentation ............ 31
Table 6 Summary of process variables, from preliminary flask fermentations, utilizing inoculant
with low cell content ..................................................................................................................... 45
Table 7. Summary of process variables during fermentation of synthetic glucose and xylose
utilizing inoculant with high dry cell concentration (7 g/L) ......................................................... 49
Table 8. Summary of process variables during flask fermentation of single sugar-comparing
arabinose, galactose and mannose with glucose ........................................................................... 51
Table 9. Summary of process variables during flask fermentation-comparison of N2 and CO2
gases .............................................................................................................................................. 52
Table 10. Summary of process variables during fermentation of synthetic glucose and xylose in
pH-controlled fermenter................................................................................................................ 55

viii
List of abbreviation
°C degree Celsius
C6 hexoses
C5 pentoses
CO carbon monoxide
CO2 carbon dioxide
C6H12O6 glucose
CH3COOH acetic acid
C5H10O5 xylose
CH4 methane
CCR carbon catabolite repression
CaCl2 calcium chloride
Co(NO3)2-6H2O cobalt nitrate hexahydrate
CSTR continues stirred-tank reactor
DOE department of energy
DP degree of polymerization
FF furfural
Fe(NH4)2SO4-6H2O ammonium ferrous sulfate hexahydrate
g/L gram per liter
h hour
H2 hydrogen
HMF 5-hydroxymethylfurfural
KH2PO4 monopotassium phosphate
K2HPO4 dipotassium phosphate
M Molar
mL milliliter
mL/min Milliliter per minute
µm micrometer
MgCl2-6H2O Magnesium chloride hexahydrate
N2 nitrogen gas
NaHCO3 sodium bicarbonate
NaOH sodium hydroxide
NaMoO4-2H2O sodium molybdate dihydrate
Na2SeO3 sodium selenite
(NH4)2SO4 ammonium sulphate
NiCl2-6H2O nickel chloride hexahydrate
nm nanometer
OD optical density
ODW oven–dried weight
rpm Revolutions per minute
SO2 Sulfur dioxide
USDA United States Department of Agriculture
UV ultraviolet light
v/v volume per volume

ix
Acknowledgements
I would like to express my deepest appreciation to my supervisor and chair Dr. Renata Bura for
her support and motivation. She provided insight and continuously challenged me to become a
better researcher. Her teachings on thoroughness and thoughtfulness will serve me for the rest of
my life.
I would like to thank my committee members Dr. Richard Roy Gustafson, and Dr. William
McKean for allowing me the opportunity to present my research to them.
It has been an honor to work with Biofuel and bioproducts laboratory group. My sincere thanks to
Dr. Shannon Ewanick and Dr. Azra Vajzovic Suko for their valuable mentorship as well as
friendship. I thank my colleagues and friends Dr. Elliott C. Schmitt, Dr. Rodrigo Morales Vera,
Chang Dou, Erik Budsberg, Jordan Crawford, Oliver Jan and Guanqun Luo for their support.
I take this opportunity to express my gratitude to all of the Department faculty members at school
of environmental and forest sciences for their constant assistance. This project would not have
been possible without the help of numerous graduate and undergraduate students. I am thankful to
Ternessa Cao, Erika Green, Yuka Harada, Kelsey Lewis, Adam Ropp, Jake Singer, Anna Song,
Kaila Turner, Conner Ross, Victoria Yuen, Zeyu Song, Vinh Tran, Josie Nguyen, Therese Krieg,
Naila Ribeiro Mori and Wilian Marcondes.
Last but not least, I would like to thank my family and friends for their continuous support
throughout my life. I am grateful to my husband, Ghassem Namazi, and my parents, Mahvash
Farazandeh and Abbas Ehsanipour for inspiring me to meet new challenges.

x
To my parents, Mahvash and Abbas
To my husband, Ghassem
To my sister and brother, Roxana and Mohammad
For their endless love, support and motivation

1
Chapter 1- Introduction
1.1 Bioconversion of lignocellulosic biomass to fuels and chemicals
The world’s consumption of petroleum based energy and chemicals has been increasing
dramatically. In order to satisfy the global energy demand, oil production from current levels of
about 80 million barrels a day needs to rise significantly to 120 million by 2040 [1]. The adverse
environmental impact of such a dramatic increase in energy usage is apparent. Global warming,
water contamination, air pollution, earth quakes, droughts and many other environmental issues
we face today is a direct result of these non-friendly environmental policies [2]. To lessen the
environmental impact of fossil fuels, the global trend in recent years has been geared towards
renewable resources and biorefineries. To support this endeavor, the United States Department of
Energy (DOE) and Department of Agriculture (USDA) have mandated that by year 2022, 5% of
heat and power energy, 20% of liquid transportation fuels, and 25% of chemicals and materials
should come from biomass[3]. The mandates emphasize particular interest in utilizing
lignocellulosic biomass for the production of biofuels and biochemicals [4].
Lignocellulosic biomass, a second generation of feedstock is a promising alternative for the
production of chemicals and fuels. It is the most abundant renewable biomass, with a worldwide
annual production of ~200 billion tons [5, 6]. Bioconversion of lignocellulose to biochemicals and
biofuels is now at the forefront of biorefinery industries.
1.2 Lignocellulosic biomass
Lignocellulosic feedstock is mainly originated from woody biomass such as hardwood (e.g. poplar,
birch, aspen), softwood (e.g. spruce, pine), agricultural residues (e.g. corn stover, wheat straw,
sugarcane straw), herbaceous or energy crops (alfalfa, switchgrass), forest residues, and other
wastes (e.g. municipal and industrial) [7]. Lignocellulose makes up the major structural material

2
of plants and is composed of a complex matrix of polysaccharides including cellulose and
hemicellulose, lignin, extractives and ash [8, 9]. The chemical composition of lignocellulose
depends on plant species, age and growth conditions. Table 1 presents the content of cellulose,
hemicelluloses, lignin, extractives and ash which varies significantly between different plants [7,
10-12].
Table 1. Composition of different lignocellulosic feedstock
Lignocellulosic
biomass
Composition (%)
Cellulose Hemicellulose Lignin Extractives Ash
Hardwood 40-55 24-40 18-25 1-5 1-4
Softwood 45-50 25-35 25-35 3-8 0.3-2
Agricultural residues 25-45 10-40 5-15 12-16 1-3
1.2.1 Cellulose
Cellulose is comprised of a long chain of glucose monomers linked to one another by β-1, 4
glycosidic bonds. The number of glucose units repeated in the chain is known as degree of
polymerization (DP) which may be 10,000 or higher [9]. The linear structure of glucose chain
enables intra and intermolecular hydrogen bonds between different layers of homo-polysaccharide.
This results in elementary fibrils that contain crystalline and amorphous regions. Likewise,
intermolecular hydrogen bonds contribute to the resistance of crystalline cellulose towards
degradation. Cellulose is highly crystalline and is resistant to hydrolysis and biological digestion
[9, 13].
1.2.2 Hemicellulose
Hemicellulose is an amorphous polymer which is, highly branched and water-insoluble [14]. This
heteropolysaccharide consists of major building blocks including hexoses (D-glucose, D-galactose
and D-mannose) and pentoses sugars (D-xylose and L-arabinose) with backbone chain composed

3
of xylan β-1, 4 linkages [7]. The degree of branching and functional groups such as acetate and
methoxyl, as well as sugars in hemicelluloses varies depending on the type of plant [13].
Agricultural residues and hardwoods are rich in pentose sugar xylose (O-acetyl-4-O-
methylglucuronoxylan), whereas softwoods are rich in hexose sugar mannose (O-
acetylgalactoglucomannan). Acetyl groups can be cleaved off from hemicellulose and form acetic
acids during pretreatment [9].
1.2.3 Lignin structure
Lignin, derived from Latin lignum, is a carbon-rich renewable resource and one of the most
abundant natural polymers, only second to cellulose [15]. It plays an important role in plant biology
by greatly increasing the strength of the cell, thus giving it the necessary rigidity [16]. It is
covalently linked to hemicellulose via ferulic acid ester linkages and holds cellulose and
hemicellulose [13]. Lignin is a highly branched amorphous and complex random polymer of
hydroxylated and methoxylated phenylpropane units. The biosynthesis of lignin involves free
radical polymerization of phenylpropane building blocks resulting in an extremely variable
structure, even within the same plant. There are three major aromatic units or monolignols: ρ-
coumaryl, coniferyl, and sinapyl alcohols. These monolignols are incorporated into lignin in the
form of phenylpronoid units: ρ-hydroxypheny (H-lignin), guaiacyl (G-lignin), syringyl (S-lignin)
[17].
1.2.4 Extractives and ash
A minor component of lignocellulosic structure known as extractives contains a variety of specific
chemicals. These include phenolics such as tannins, terpene alcohols, ketones as well as resin
components such as fatty acids, alcohols, resin acids, phytosterols, and minerals (calcium,
magnesium and potassium) [18, 19]. These compounds can be extracted using polar and non-polar

4
solvents. The extractives are often responsible for certain characteristics in plants including color,
odor, physical and flavor properties [20].
1.3 Bioconversion process
Bioconversion is the key process in a biorefinery when converting lignocellulosic biomass into
low-value high-volume fuels and high-value low-volume chemicals [14]. The bioconversion
process encompasses upstream and downstream processes, from feedstock preparation to product
recovery. Some of these steps include i) pretreatment to overcome recalcitrant nature of
lignocellulosic biomass, and unlock polymeric sugars and improve accessibility of the substrate to
subsequent steps ii) enzymatic hydrolysis to depolymerize cellulose and hemicellulose, and iii)
fermentation of released sugars to different biochemicals and biofuels [21]. Process design criteria
significantly depends on the type of lignocellulosic biomass and desired product that will be
produced.
1.3.1 Pretreatment
Pretreatment is a fractionation technique (physical, chemical, or both) meant to disrupt the
structural bonds, break the lignin sheath, separate hemicelluloses from cellulose, and alter
crystallinity of cellulose. There have been a lot of effort to develop efficient pretreatment methods
to overcome the complex structure of lignocellulosic materials [22]. Amongst pretreatment
methods, steam explosion has proven to have the ability to fractionate a wide variety of
lignocellulosic biomass including woody (softwoods and hardwoods), agricultural and herbaceous
residues, as well as energy crops [23] with high sugar recovery [24]. Steam explosion pretreatment
can be carried out at high temperatures (160-260°C), pressure (0.69-4.83 MPa or 100-700 Psi) and
residence time for several seconds to a few minutes [25]. Fractionation can be improved by sulfur
dioxide impregnation (1-4% w/w substrate) [22]. To maximize hydrolyzability of solids and

5
hemicellulose removal in softwoods and hardwoods, it is necessary to use SO2 gas as a catalyst in
steam pretreatment [9]. Assessment of pretreatment condition is described as “severity factor”, a
correlation between steam temperature and residence time, where t is time in seconds and T is
temperature in degrees Celsius:
Ro=t.e (T-100)/14.75 (Equation 1.1)
One will have to include the pH value when pretreatment is performed with acidic pH, using SO2
gas. The following equation describes the combined severity (CS), where pH is measured after
pretreatment.
CS=log (Ro) - pH (Equation 1.2)
Depending on lignocellulosic biomass, a low severity factor may result in an incomplete
fractionation of biomass, whereas a high severity factor results in a more complete deconstruction
with better hemicellulose solubility and further delignification [24].
1.3.2 Enzymatic hydrolysis
Enzymatic hydrolysis is a process in which biocatalysts facilitate the conversion of physio-
chemically pretreated substrate to fermentable sugars. Enzymatic pretreatment of lignocellulosic
biomass relies on two enzyme complexes including cellulases and hemicellulases, multi-domain
proteins [26]. Thus efficient conversion of lignocellulosic biomass during enzymatic hydrolysis
requires a mixture of different enzymes acting sequentially or in concert.
In a natural fermentation process, a potent cellulolytic organism such as Trichoderma reesei
produces complex mixtures of enzymes composed of three major groups. The groups include
endogluconases (CM-cellulases), exoglucanases or cellobiohydrolases, and ß-glucosidases or
cellobiases. Digestibility of celluloses according to its structure occurs as below [27]:

6
Endoglucanases cleave the internal ß–1, 4–glycosidic bonds of amorphous, swollen
cellulose and release glucose, cellobiose and cello-oligosaccharides.
The exoglucanases cleave cellobiose units from the ends of the polysaccharide chains.
ß–glucosidases cleave cellobiose to release glucose.
1.3.3 Fermentation
Fermentation is a metabolic process that harnesses a microorganism’s ability to digest
lignocellulosic sugars (pentoses and hexoses) to produce products such as ethanol, butanol,
acetone, acetic acid, butyric acid, H2 gas, and CH4 gas [28]. Numerous microbial strains (bacteria
and fungi) have been utilized in fermentation of lignocellulosic substrates [29]. Some of the well
utilized microorganisms include Zymomonas mobilis [30], Saccharomyces cerevisiae, Clostridium
thermocellum as ethanologenic [31]; Moorella thermoacetica, Clostridium ljungdahlii as
acetogenic [32]; Clostridium tyrobutyricum JM, a hydrogen producing [33]; and Clostridium
acetobutylicum for production of acetone, ethanol, and butanol [34].
The versatility of microorganisms plays a significant role in fermentation processes employed in
biorefineries [29]. In all fermentations a high level of production yield is desired which depends
on specific microbial traits including high tolerance to substrate concentration, and end product as
well as inhibitor compounds. Furthermore, it is required to have a broad substrate utilization (C6 and
C5 sugars), with minimal byproduct formation [35].
1.3.4 Inhibitors
A drawback during pretreatment is formation of chemical compounds that may inhibit microbial
activity or increase the lag phase of microbial growth during a fermentation process. Inhibitors
(toxic substances) may also decrease the production yield in a fermentation process [35]. The

7
composition of toxic compounds depends on type of lignocellulosic biomass and the severity of
pretreatment method [36]. Inhibitors are categorized in three different groups i) weak acids: acetic
acid, levulinic acid, glucuronic acid, furoic acid and humic acid, ii) furan derivatives:
hydroxymethylfurfural (HMF), furfural (FF), and iii) phenolic compounds: hydroxybenzoic acid,
vanillic acid, and ferulic acid [37]. Acetic acid forms through acetyl cleavage in hemicellulose,
furfural through degradation of pentose monomers, and HMF from degradation of hexoses. FF and
HMF can also be further degraded to levulinic and humic acids. Furthermore, degradation of lignin
results in phenolic derivatives [37-40]. The concentration of these chemical compounds varies
according to type of lignocellulosic biomass and severity of pretreatment conditions [35, 41].
These fermentation inhibitors are building blocks for other chemicals such as furfural [42] for
tetrahydrofurfuryl alcohol [43] and levulinic acid for valerolactone [42]. The US Department of
Energy (DOE) has created a list of value added chemicals that may be extracted from
lignocellulosic biomass, thus giving credibility to lignocellulosic biorefinery [44]. Acetic acid is a
value added chemical that can also be produced through a fermentation process.
1.4 Acetic acid
The current global interest in renewable resources and biorefineries has provided momentum and
new opportunities in the area of manufacturing biochemicals. In order for current biorefineries to
be competitive with legacy based fossil fuel technologies, a more efficient process for the
production of bio-based chemicals needs to be developed. One such bio-based chemical is acetic
acid, an important carboxylic acid with a broad spectrum of industrial and household applications
including polymers, paints, foods and pharmaceuticals. Acetic acid is a major building block for
chemicals such as polyvinyl and cellulose acetate which is further used in the production of fibers
and fabrics [45].

8
Currently, more than 65% of global acetic acid manufacturing process is based on catalytic
carbonylation of methanol utilizing rhodium complex catalyst [45, 46]. Other procedures involve
oxidation of acetaldehyde, liquid phase oxidation of n-butane or naphtha, and chemical conversion
of syngas produced through gasification of coal [47]. The United States acetic acid production
capacity is 788 MM gal/year with spot market price at $2.5 per gallon [47, 48]. In comparison,
bioethanol production capacity is 15,135 MM gal/year [49] with a price of $1.8 per gallon [50].
Hence, production of acetic acid, a high-value commodity through bioconversion of
lignocellulosic biomass is economically viable [51]. Acetogenic bacteria has shown to be a
promising candidate for the production of acetic acid [52].
1.5 Moorella thermoacetica
Since 1923, over a hundred acetogenic species representing twenty two genera have been isolated
from a variety of habitats [32]. Among acetogens, Moorella thermoacetica (a gram-positive
thermophilic, anaerobic bacterium, previously known as clostridium thermoacetica) is a
homoacetogen in which acetate is the major product of its metabolic pathway [53]. This acetogen
is robust with a wide range of growth temperatures (45-65ºC) and pH levels (5.7-7.7). The
optimum growth temperature range is 55-60ºC and pH level of 6.8 [54]. This bacteria has a diverse
and unique metabolic capability. It is able to produce acetic acid or acetate by metabolizing certain
sugars including xylose, fructose and glucose. Xylose is the preferred sugar and consumed first,
followed by fructose and glucose [55-57].
Metabolic pathway of Moorella thermoacetica has been extensively studied by Wood, Ljungdahl,
Daniel and Drake. Basically, glucose or fructose is oxidized to pyruvate through Embden-
Meyehof-Parnas glycolytic pathway (glycolysis) and is further metabolized to two molecules of
acetate and two molecules of carbon dioxide (CO2). This acetogen is also able to synthesize acetate

9
from carbon dioxide through acetyl-CoA Wood–Ljungdahl pathway [54, 58]. As a result, syngases
may be utilized as a substrate (chemolithoautotrophic growth on H2/CO2 or CO/CO2) [52-54]. This
acetogen is capable of fixing CO2 in which one molecule of acetate is synthesized from carbon
dioxide through Acetyl-CoA pathway (Wood–Ljungdahl pathway) reactions [54, 58]. Conversion
of glucose to acetic acid is presented in the following stoichiometry reactions. Sources of CO2 can
be from outside of cells (exogenous CO2 used to maintain anaerobic condition) or through
decarboxylation of pyruvate (inside of cells) [53].
Oxidation: C6H12O6 + 2H2O → 2CH3COOH + 2CO2 + 4H2 (1)
Reduction: 2CO2 + 4H2 →+ 3COOH + 8H2O (2)
Net reaction: C6H12O6 → 3CH3COOH (3)
Unlike glucose, the pathway for conversion of xylose to acetate in M. thermoacetica has not been
well investigated [54]. However, Pierce and coworkers, 2008, emphasized that M. thermoacetica
encodes xylose to acetate through pentose phosphate pathway in which xylose converts to xylitol.
Xylitol is subsequently oxidized to xylulose followed by another conversion to glyceraldehyde 3-
phosphate. At this point it merges with glycolysis pathway [59]. Stoichiometric conversion of
xylose to acetic acid is summarized in the following reaction [54].
2 C5H10O5 → 5CH3COOH (4)
A typical acetic acid production in a batch fermentation using synthetic glucose (20 g/L) is around
13 g/L [51]. The highest acetic acid production reported was at 31 g/L using CO/CO2 gas mixture
[60]. On average, bacterial tolerance of acetate is between 30-50 g/L in a batch fermentation [61].
Previous efforts regarding production of acetic acid utilizing M. thermoacetica have by and large
concentrated on industrially suitable culture media and fermentation setups using synthetic sugars.

10
1.5.1 Cultivation Media
M. thermoacetica has complex nutrient requirements in which a typical cultivation medium
includes carbon source, trace metals, salts, reducers, yeast extract and buffer solutions [52]. One
of the major issues has been the development of a cost effective growth medium using different
substitutions for yeast extract [62-65]. Corn steep liquor (CSL) pretreated with dolime (calcium
and magnesium oxide, CaO-MgO) and vitamin supplementation has shown to be a promising
replacement since final acetate concentration was improved by 110% (from 19 to 40 g/L in a fed-
batch fermentation) [65].
1.5.2 Fermentation setups
Various fermentation setups have also been evaluated to optimize acetate production [66-69].
Results from fed-batch fermentation and continuous stirred-tank reactor (CSTR), using cell recycle
membrane system were shown to have a higher concentration of acetate (34-38 g/L) than one-step
CSTR (25.5 g/L) [65].
1.5.3 Extracted sugars
A number of commercial and extracted sugars have also been evaluated for the production of acetic
acid. Brownell and coworker utilized extracted xylose from debarked poplar wood chips (Soxhlet
extraction using benzene–ethanol) and commercial oat spelt xylan (average acetate yield of 70%)
[70]. The commercial glucose from -cellulose and pulp mill sludge with high enzyme loading at
50 IFPU/g glucan were also utilized. Overall acetate yield was 60% and 85% respectively. The
authors stipulated that acetate production was limited by toxic materials from the sludge [71].

11
1.6 Research objectives
To the best of the author’s knowledge, sugars obtained from bioconversion of lignocellulosic
biomass have not been investigated for acetic acid production. For the first time this paper presents
an original study about bioconversion of lignocellulosic hydrolysate to acetic acid utilizing M.
thermoacetica. The objective of this research is to examine the ability of M. thermoacetica to
ferment monomeric sugars in four different steam exploded hydrolysates including wheat straw,
forest residues, switchgrass, and sugarcane straw.

12
Chapter 2 Fermentation of lignocellulosic sugars to acetic acid by
Moorella thermoacetica
To be submitted for publication in an international journal.
Abstract
A systematic study of bioconversion of lignocellulosic sugars to acetic acid by Moorella
thermoacetica strain ATCC 39073 was conducted. Four different water-soluble fractions
(hydrolysates) obtained after steam pretreatment of lignocellulosic biomass were selected and
fermented to acetic acid in batch fermentations. The bacterial strain (ATCC 39073) can effectively
ferment xylose and glucose in hydrolysates from wheat straw, forest residues, switchgrass, and
sugarcane straw to acetic acid. This homoacetogen prefers xylose over glucose. The highest acetic
acid yield was obtained from sugarcane straw hydrolysate with 71% of theoretical based on total
sugars. The lowest acetic acid yield was observed in forest residues hydrolysate with 39% of
theoretical based on total sugars. This acetogen consumes arabinose, galactose and mannose within
72 h of fermentation in the mixture of lignocellulosic sugars. Acetic acid yield in forest residues
was adversely affected by high concentration of arabinose, galactose and mannose. It was also
observed that M. thermoacetica can tolerate process derived inhibitory compounds from steam
explosion pretreatment (total phenolics up to 3 g/L and furans up to 0.5 g/L).
2.1 Introduction
The current global interest in renewable resources and biorefineries has provided momentum and
new opportunities in the area of manufacturing biochemicals. In order for current biorefineries to
be competitive with legacy based fossil fuel technologies, a more efficient process for the
production of bio-based chemicals needs to be developed. One such bio-based chemical is acetic
acid, an important carboxylic acid with a broad spectrum of industrial and household applications
including polymers, paints, foods and pharmaceuticals. Acetic acid is a major building block for
chemicals such as polyvinyl and cellulose acetate which is further used in the production of fibers
and fabrics [1].
Currently, more than 65% of global acetic acid manufacturing process is based on methanol
carbonylation utilizing rhodium complex (rhodium iodides) catalyst [1, 2]. Other procedures
involve oxidation of acetaldehyde, liquid phase oxidation of n-butane or naphtha, and chemical

13
conversion of syngas produced through gasification of coal [3]. The United States acetic acid
production is 788 MM gal/year with spot market price at $2.5 per gallon [3, 4]. In comparison,
bioethanol production is 15,135 MM gal/year [5] with a price of $1.8 per gallon [6]. Hence,
production of acetic acid, a high-value commodity through bioconversion of lignocellulosic
biomass is economically viable [7]. Acetogenic bacteria has shown to be a promising candidate
for the production of acetic acid [8].
Amongst acetogens, Moorella thermoacetica (a gram-positive thermophilic, anaerobic bacterium,
previously known as Clostridium thermoacetica) is a homoacetogen in which acetate is the major
product of its metabolic pathway [9]. M. thermoacetica is a robust and versatile heterotroph with
optimum growth temperature range of (55-60) ºC, and pH level of 6.8 [10]. This bacteria has a
diverse and unique metabolic capability. It is able to produce acetic acid / acetate by metabolizing
certain sugars including xylose, fructose and glucose. Xylose is the preferred sugar and consumed
first, followed by fructose and glucose [11-13]. The conversion of glucose and fructose is through
Embden-Meyehof-Parnas glycolytic pathway [10, 14] while xylose is through pentose phosphate
pathway [15]. Furthermore, M. thermoacetica is able to synthesize acetate from carbon dioxide
through acetyl-CoA Wood–Ljungdahl pathway [10, 14]. As a result, syngases may be utilized as
a substrate (chemolithoautotrophic growth on H2/CO2 or CO/CO2) [8-10]. Typical acetic acid
production in a batch fermentation using synthetic glucose (20 g/L) is around 13 g/L [7]. The
highest acetic acid production reported was at 31 g/L using CO/CO2 gas mixture [16]. On average,
bacterial tolerance of acetate is between 30-50 g/L in a batch fermentation [17].
Previous efforts regarding production of acetic acid utilizing M. thermoacetica have by and large
concentrated on developing industrially suitable culture media and fermentation setups using
synthetic sugars. M. thermoacetica has complex nutrient requirements in which a typical

14
cultivation medium includes carbon source, trace metals, salts, reducers, yeast extract and buffer
solution [8]. One of the major issues has been the development of a cost effective growth medium
using different substitutions for yeast extract [18-21]. Corn steep liquor (CSL) pretreated with
dolime (calcium and magnesium oxide, CaO-MgO) and vitamin supplementation has shown to be
a promising replacement since final acetate concentration was improved by 110% (from 19 to 40
g/L in a fed-batch fermentation) [21]. Various fermentation setups have also been evaluated to
optimize acetate production [22-25]. Results from fed-batch fermentation and continuous stirred-
tank reactor (CSTR), using cell recycle membrane system were shown to have a higher
concentration of acetate (34-38 g/L) than one-step CSTR (25.5 g/L) [21].
A number of commercial and solvent extracted sugars have also been evaluated for the production
of acetic acid. Brownell and coworker utilized extracted xylose from debarked poplar wood chips
(Soxhlet extraction using benzene–ethanol) and commercial oat spelt xylan (average acetate yield
of 70%) [26]. The commercial glucose from -cellulose and pulp mill sludge were also utilized and
the overall acetate yield was 60% and 85%, respectively. The authors stipulated that acetate
production was limited by toxic materials from the sludge [27].
To the best of our knowledge, this is the first report on the bioconversion of lignocellulosic sugars
to acetic acid by M. thermoacetica. The objective of this work is to examine M. thermoacetica in
fermentation of four different types of hydrolysates obtained after steam pretreatment of
lignocellulosic biomass including wheat straw, forest residues, switchgrass, and sugarcane straw.
The ultimate goal of our research is to establish bioconversion process of lignocellulosic biomass
to acetic acid using both hexose and pentose fractions after steam pretreatment for production of
acetic acid.

15
2.2 Materials and methods
2.2.1 Moorella Thermoacetica strain (ATCC 39073)
Moorella thermoacetica (ATCC 39073) was acquired from the American Type Culture Collection
(Manassas, VA) grown in a growth medium containing g/L of the following compounds: yeast
extract, 10.0; glucose, 10.0; xylose, 10.0; L-Cysteine Hydrochloride Monohydrate 1.0; KH2PO4
7.0; NaHCO3 16.0; K2HPO4 5.5; (NH4)2SO4 1.0; MgCl2-6H2O, 0.33; CaCl2 0.05; NaMoO4-2H2O
0.0025; Fe(NH4)2SO4-6H2O 0.04; Co(NO3)2-6H2O 0.03; Na2SeO3 0.0002; NiCl2-6H2O 0.0002.
The pH level of the medium was adjusted to 7.3 using 5M NaOH [7, 11].
2.2.2 Culture media conditions
Cells were grown to a high cell density through three sequential passagings, from 50 mL to 200
mL in stoppered and crimp-sealed Wheaton serum bottles. Prior to inoculation, Wheaton serum
bottles containing growth medium were sparged with filter-sterilized 100% CO2 and inoculated
with 10% by volume serum bottles incubated at 58°C for 36 h in dark condition (incubator in
Bactron II anaerobic chamber, Sheldon manufacturing, Inc.). The third passage (200 mL) was used
as the inoculum for fermentation. After 36 h of growth, cell cultures were harvested and
centrifuged at 4000 rpm (Beckman GS-15R, Germany) for 5 minutes. They were subsequently
decanted to yield cell pellets with a concentration of 7 g/L dry cell weight (DCW). The
concentration of dry cell weight was measured based on optical density using a UV
spectrophotometer (Shimadzu UV-1700, Columbia, MD) via standard curves relating 600nm
absorbance to DCW per liter.
2.3 Water-soluble fraction (hydrolysate)
The water-soluble fractions (hydrolysates) used in this study were produced from wheat straw,
forest residues, switchgrass, and sugarcane straw. Each lignocellulosic biomass was soaked to

16
saturation in deionized (DI) water overnight prior to SO2-catalyzed steam explosion. The detailed
procedure of steam explosion experiments has been described previously by Ewanick [28]. Briefly,
samples of 200g oven-dried weight (ODW) lignocellulosic biomass were impregnated overnight
with anhydrous 3% (w/w) SO2. The samples were subsequently loaded in a steam gun and kept
for five minutes at a temperature of 195°C, except for sugarcane straw which was treated at 195°C
(Figure 1). The water-soluble fraction (hydrolysate) from steam explosion of each lignocellulosic
biomass was recovered by filtration and kept at 4°C until use. The initial concentration of sugars
present in the hydrolysates was measured.
2.4 Overliming
Prior to fermentation, each hydrolysate was conditioned by an overliming (OL) process in which
the pH level was increased from 1.9 to 10.0 with Ca(OH)2 (industrial and chemical purposes,
Graymont, USA). The pH level was next readjusted to 5.0 with additional sulfuric acid. The
overliming process was carried out based on procedure described by national renewable energy
laboratory (NREL) [29].
2.5 Fermentation
Four different lignocellulosic hydrolysates as well as their corresponding synthetic sugar models
were fermented. Figure 1 presents a simplified scheme of fermentation experiments. All
fermentation experiments were performed in duplicates using a 1.3 L Bioflo 115 (New Brunswick
Scientific Co., Inc., Edison, NJ) bioreactor with 500 mL of working volume under anaerobic
conditions using filter-sterilized 100% CO2 gas sparged into the bottom of the fermenter vessel
continuously prior to inoculation. Sparging was maintained at headspace after inoculation and
throughout fermentation. Inoculation was performed with 10% v/v of three-stage seeded bacterial
cells grown on glucose and xylose in a 1:1 ratio. All fermentations were maintained at 58°C with

17
continuous agitation (175 rpm), CO2 gas flow rate of 1 standard liter per minute (SLPM) and
monitored for 72 h. A solution of 5 M NaOH was added to sustain pH value of 6.8 throughout
fermentation. Samples were taken at the time of inoculation and at specific intervals thereafter.
One milliliter of aliquots was immediately centrifuged (10,000 rpm) for 5 minutes at 4°C to
separate microbial cell and supernatant. The supernatant was filtered by using a 0.2 µm syringe
filter and then stored at -20°C until analysis. The bacterial growth was monitored during
fermentations. Optical density (OD) of cell was measured at 600nm by spectrophotometer
(Shimadzu UV-1700, Columbia, MD). Conversion of OD to dry cell mass was constructed based
on NREL protocol (N. Dowe and J. McMillan, 2001).
2.5.1 Hydrolysate fermentation
Prior to fermentation, all overlimed hydrolysates were supplemented with nutrients used in growth
medium and initial pH level was adjusted to 7.3. Xylose concentration in all hydrolysates was
spiked to 20 g/L in order to stablish a baseline. All hydrolysates were filter sterilized using a 500
mL rapid flow bottle tope with 0.2 µm sterile filter.
2.5.2 Synthetic sugars fermentation
Synthetic sugars fermentation was performed as an appropriate control for each type of
hydrolysate. The media was prepared similarly to growth medium except for sugars content.
Glucose, arabinose, galactose and mannose were added based on their corresponding concentration
measured in each one of overlimed hydrolysates. For all controls, xylose concentration was kept
at 20 g/L. The fermentation of all synthetic sugars models were performed with the same manner
as fermentation experiments of hydrolysates.

18
Figure 1. Schematic diagram of fermentation experiments. All lignocellulosic biomass were steam
exploded at 195°C, except for sugarcane straw. Sugarcane straw was steam exploded at 190°C.
2.6 HPLC analysis
2.6.1 Monomeric sugars
The concentration of monomeric sugars (arabinose, galactose, glucose, xylose and mannose) was
measured on a Dionex (Sunnyvale, CA) high-performance liquid chromatography (HPLC, ICS-
3000) system equipped with AS (auto sampler), ED (electrochemical detector), dual pumps and
anion exchange column (Dionex, CarboPac PA1). Deionized water at 1.0 mL/min was used as
eluent, post column addition of 0.2 M NaOH at a flow rate of 0.5 mL/min ensured optimization of
baseline stability and detector sensitivity. After each analysis, the column was reconditioned with
0.25 M NaOH. Ten microliters of each sample was injected after filtration through a 0.2 µm
Lignocellulosic biomass
Steam explosion
195°C, 5 min, 3% SO2
190°C*, 5 min, 3% SO2
Solid and liquid
separation
Liquid fraction
(hydrolysate)
Synthetic sugars model in each
hydrolysate
Wheat
straw Switchgrass
Forest
residues
Sugarcane
straw*
Acetate
Control
Fermenter 58 °C, pH 6.8, 175 rpm
Control cabinet (PID controller) for CO2 gas flow, temperature, pH value
and agitation

19
syringe filter (Restek Corp., Bellefonte, PA, USA). Standards were prepared containing sufficient
arabinose, galactose, glucose, xylose and mannose to encompass the same range of concentration
as the samples. Fucose was added to all samples as an internal standard.
2.6.2 Acetate, hydroxymethylfurfural and furfural analysis
Acetic acid, hydroxymethylfurfural and furfural were measured using refractive index detection
on a Shimadzu Prominence LC. The separation of these compounds was achieved by an anion
exchange column [REZEX RHM-Mono saccharide H+(8%); Phenomenex, Inc., Torrance, CA,
USA] with an isocratic mobile phase that consisted of 5mM H2SO4 at a flow rate of 0.6 mL/min.
The column oven was maintained at a constant temperature of 63°C. Ten microliters of each
sample was injected after being appropriately diluted in deionized water and filtered through a 0.2
µm syringe filter. The standards were prepared and used to quantify the unknown samples.
2.7 Phenolic measurement
Total phenolic was measured based on colorimetric reaction using Folin-Ciocalteau (FC) method.
Gallic acid was utilized as an equivalent to the phenolic compounds. Gallic acid (0.500 g) was
dissolved in 10 mL of ethanol and diluted to 100 mL in volumetric flask using distilled/deionized
water. A set of standards (different concentration of gallic acid) was prepared to create a calibration
carve. Sodium carbonate solution was used for colorimetric reaction. Procedure was followed as
described by A. L. Waterhouse, 2012 [30].
2.8 Results and discussion
In order to examine the ability of M. thermoacetica to ferment lignocellulosic sugars to acetic acid,
fermentation of four different types of lignocellulosic hydrolysates were conducted. Hydrolysates
obtained from steam pretreated wheat straw, forest residues, switchgrass, and sugarcane straw
were used as medium for production of acetic acid by M. thermoacetica (ATCC 39073). The

20
synthetic sugars models (controls) consisted of synthetic sugars at the same concentration as
measured in hydrolysates (described in “Methods” section). A typical fermentation profile for
sugar consumption, acetic acid production, and cell growth for each hydrolysate is shown in Figure
2, plots A, B, C and D.
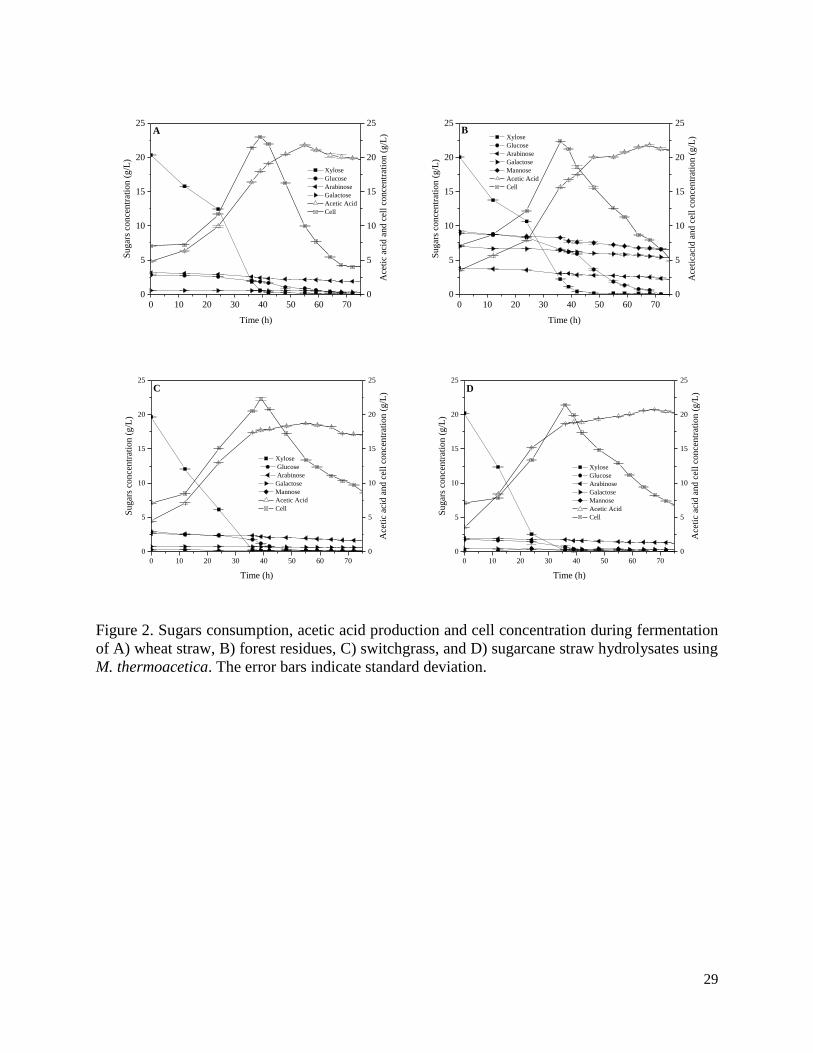
In general, it was shown that xylose was the initial sugar which was completely metabolized by
M. thermoacetica in all hydrolysates. Glucose started to be metabolized after more than half of
xylose was consumed. A minimal amount of arabinose, galactose and mannose was gradually
consumed in a period of 72 h of fermentation. The fastest xylose consumption (35 h) was observed
in sugarcane straw and switchgrass hydrolysates while the slowest (50 h) was observed in forest
residues hydrolysate (Figure 2, plots B, D). Xylose uptake commenced by M. thermoacetica in all
hydrolysates without any lag phase, except for wheat straw hydrolysate which had a lag phase of
12 h. Glucose was completely consumed within a range of 40 h (fastest in sugarcane straw
hydrolysate) to 70 h (slowest in forest residues hydrolysate). Low concentrations of minor sugars
including arabinose, galactose and mannose were detected in all hydrolysates except for forest
residues which had a considerable amount of these sugars (Table 1). The consumption of these
sugars was incomplete and the following was observed: the lowest consumption of arabinose in
switchgrass (37%), galactose and mannose in sugarcane straw (18% and 28% respectively), and
the highest consumption of arabinose and galactose in wheat straw (62% and 49% respectively),
mannose in switchgrass (66%).
In fermentation of controls, a similar trend in sugars consumption was observed (graphs for
controls not shown). Xylose uptake in each control was the same as its corresponding hydrolysate
except for wheat straw and forest residues controls in which it was consumed slower (60 h and 65
h respectively). The pattern for glucose consumption was also comparable to hydrolysates.

21
However, glucose in control for forest residues was not completely utilized over the fermentation
period of 72 h (1 g/L remained, plots not shown). The incomplete consumption of arabinose,
galactose and mannose in controls were different from hydrolysates. The lowest arabinose and
galactose consumption was observed in forest residues control (42% and 13% respectively) and
mannose in controls for switchgrass (12%). The highest consumption of these sugars was observed
as follows: arabinose in control for sugarcane straw (54%), galactose in control for switchgrass
(36%), and mannose in control for sugarcane straw (55%).
The highest cell biomass was observed in wheat straw hydrolysate (23 g/L) while the lowest was
observed in sugarcane straw hydrolysate (21.4 g/L) (Table 1; Figure 2). In synthetic sugar models,
wheat straw control had the highest cell concentration (24.5 g/L) while forest residues control had
the lowest (19 g/L). In general, cell concentration was higher (0.2-1.5 g/L) in each control
compared to its hydrolysate except for synthetic sugar model for forest residue (Table 1).
Acetic acid production yield was calculated based on total initial sugars (Table 1). The highest
acetic acid yield was observed in steam pretreated sugarcane straw hydrolysate with 71% of
theoretical yield (17.2 g/L) while the lowest yield was observed in forest residues hydrolysate with
38% of theoretical (18.2 g/L). Acetic acid yield was higher for all hydrolysates compared to their
corresponding controls except for forest residues control (39%) and switchgrass control (55%).
The conversion of sugars to acetic acid in fermentation of hydrolysates were as good as or higher
than their corresponding controls.
It was observed that M. thermoacetica was not able to consume arabinose, galactose and mannose
in a fermentation of single sugar which resulted in a cluster of dead cells (Table 2). The effects of
minor sugars on acetic acid yield was investigated in two separate flask fermentations using mixed
sugars with equal concentrations (Table 3). In one fermentation, inoculant was pre-grown on the

22
same mixture of sugars while in the second fermentation inoculant was pre-grown on a 1:1 ratio
of glucose to xylose. Table 3 presents that M thermoacetica could completely metabolize xylose
and glucose while the consumption of other sugars was incomplete (<40%) which resulted in a
very low acetic acid yield.
There are not any references regarding fermentation of mixed lignocellulosic sugars by M.
thermoacetica. In a study by Balasubramanian and coworkers, fermentation of mixed synthetic
sugars (4.5 g/L xylose, 5.5 g/L glucose, 5 g/L fructose, 5.5 g/L arabinose, 4.5 g/L galactose and 5
g/L mannose) using M. thermoacetica’s inoculant with a very low cell concentration (~2%, v/v)
was investigated[13]. The consumption of minor sugars for a fermentation period of 135 h was
estimated as follows: arabinose 36%, galactose 11%, and mannose 20%. Additionally, acetic acid
yield based on total sugars was 45% [13]. The authors utilized an average concentration of 5 g/L
for each sugar and inoculant with ~2% (v/v) pre-grown on glucose and subsequent transfer in
xylose. By contrast, in our experiments the consumption of arabinose, galactose and mannose was
less than 50%. This difference in partial consumption of minor sugars can be attributed to the
following: i) presence of fructose in their fermentation medium, ii) the initial concentrations of
sugars and iii) inoculant’s cell concentration. It is worth noting that the presence of these minor
sugars lowered the acetic acid yield which is comparable to fermentation results of forest residues
hydrolysate.
Another possible explanation for low acetic acid yield could be the glucose to xylose ratio. Table
4 presents pH-controlled batch fermentation results from the mixtures of glucose and xylose with
different concentrations. The concentration of glucose and xylose in fermentation number (no.) 1
corresponds to wheat straw, switchgrass and sugarcane straw hydrolysates. Glucose and xylose
concentration in fermentation no. 3 is similar to forest residues hydrolysates. In this set of

23
experiments, glucose and xylose consumption as well as acetic acid yield were monitored over a
period of 50 h, 75 h and 96 h of fermentation. In a 50 h period of fermentation, the highest acetic
acid yield based on total sugars (75% of theoretical) was observed in mixtures with high
concentration of glucose to xylose (20 g/L to 3 g/L and 20g/L to 9 g/L). In these mixtures, xylose
and glucose were completely metabolized except for fermentation no. 4 which had a residual
glucose less than 1 g/L. By contrast, low acetic acid yield was observed in fermentation no. 3 and
4 (50% and 59% of theoretical respectively) with considerable residual of glucose and xylose
(Table 4, fermentation no. 1 and 3). After 75 h of fermentation, media with high glucose
concentration (Table 4, fermentation no. 2 and 4) presented higher acetic acid yield (75% and 77%
of theoretical respectively) than fermentation no. 1 and 3 with high xylose concentration (67% and
70% of theoretical respectively). A Similar trend in acetic acid yield was also observed after 96 h.
It was noted that glucose to xylose ratio had a significant influence on acetic acid yield and sugars
uptake.
In fermentation of hydrolysates and their controls, as well as mixtures of glucose and xylose,
glucose uptake was clearly repressed by xylose which emphasizes the presence of carbon
catabolite repression (CCR) mechanism in M. thermoacetica. The CCR mechanism in M.
thermoacetica appeared to influence acetic acid yield. Glucose to xylose ratio as well as the
presence of arabinose, galactose and mannose intensify CCR behavior in M. thermoacetica.
Specifically, in forest residues hydrolysate, acetate yield was adversely affected by high
concentration and incomplete consumption of minor sugars as well as glucose to xylose ratio
(Figure 2, plot B; Table 1). Furthermore, bacterial pre-growth on a 1:1 ratio of glucose to xylose
may have a direct influence on CCR and subsequently on acetate yield. Brumm observed a 20%

24
lower acetate yield in fermentation of xylose utilizing culture which was grown on an equivalent
amount of fructose or glucose compared to culture pre-grown on xylose [12].
Based on our research results, in order to obtain an optimum acetic acid yield, it is imperative to
have lignocellulosic hydrolysate with a high xylose content and zero or a very minimal amount of
other lignocellulosic sugars. Hence, steam explosion pretreatment design is a key process that
needs to be customized to extract xylose and glucose in a step-wise manner. In the initial step, with
low severity factor, collect as much xylose in liquid stream (hydrolysate) as possible. In the second
step, steam explode the solid stream to collect the remaining xylose which is in water-soluble
stream. The choice of lignocellulosic feedstock also improves acetic acid yield. Hardwoods and
agricultural residues have a great potential for the production of acetic acid. Softwoods would not
be a good option due to their high content of galactoglucomannan [31].
M. thermoacetica tolerated process derived inhibitory compounds from steam explosion
pretreatment including hydroxymethylfurfural (HMF) and furfural (FF) (up to 0.28 g/L and 0.9
g/L, respectively, data are not shown) and total phenolics of up to 3 g/L (Table 1). This bacterial
strain did not metabolize phenolic compounds during fermentations. However, it did consume
HMF and FF in switchgrass (0.44 g/L and 0.1 g/L respectively) and sugarcane straw (0.01 g/L of
each) hydrolysates in the first 12 h of fermentation (Table 1). The consumption of HMF and FF
may also intensify CCR mechanism. Total phenolics and furans did not inhibit microbial growth
or acetic acid production. This characteristic has also been noted in a prior study utilizing yeast for
production of xylitol and ethanol from lignocellulosic hydrolysate. This study used a mixture of
hybrid poplar and Douglas fir collected after steam explosion pretreatments [32].

25
2.9 Conclusion
It was presented for the first time that M. thermoacetica (ATCC 39073) can effectively ferment
xylose and glucose content in water-soluble fraction of steam exploded lignocellulosic biomass
including wheat straw, forest residues, switchgrass, and sugarcane straw to acetic acid. In all
fermentations xylose was the major and the first sugar being consumed. Xylose consumption was
faster in switchgrass and sugarcane straw hydrolysates (35 h) than wheat straw and forest residues
hydrolysates (40 and 45 h respectively). Xylose repressed glucose uptake in which glucose was
metabolized when the majority of xylose was consumed. M. thermoacetica consumed less than
50% of arabinose, galactose and mannose within 72 h of fermentation in a mixture of sugars. The
highest cell biomass was observed in wheat straw hydrolysate (23 g/L) and the lowest in sugarcane
straw hydrolysate (21.4 g/L). The highest acetic acid production based on total sugars was obtained
from sugarcane straw with 71% of theoretical yield while it was 64% in its control. The lowest
acetic acid yield based on total sugars was observed in forest residues with 39% of theoretical. The
incomplete consumption of arabinose, galactose and mannose adversely affected acetic acid yield.
M. thermoacetica demonstrated that it tolerates process derived inhibitory compounds from steam
explosion pretreatment including HMF (0.44 g/L), FF (0.1 g/L) and total phenolics (3 g/L). The
variations observed in acetic acid yields, repressed glucose uptake by xylose, and incomplete
consumption of minor sugars confirmed CCR behavior in M. thermoacetica.
Acknowledgements
This research was supported by U. S. Department of Agriculture with funding from Agriculture
and Food Research Initiative (USDA AFRI grant 2011-68005-30407). We would like to thank the
University of Washington-Denman Professorship funds in Bioresource Engineering as well as

26
Biofuels and Bioproducts Laboratory Research Group. Our special thanks to Dr. Shannon
Ewanick, Dr. Rodrigo Morales, and Naila Ribeiro Mori for providing hydrolysates.
References
1. Grand View Research I (2014) Acetic acid market analysis and Segment forecasts to 2020. In:.
http://www.grandviewresearch.com/industry-analysis/acetic-acid-market
2. Yoneda N, Kusano S, Yasui M, Pujado P, Wilcher S (2001) Recent advances in processes and
catalysts for the production of acetic acid. Applied Catalysis A: General Published by Elsevier
Science B.V. 221:253-265
3. Clark B (2015) Chemical profile, US acetic acid. ICIS Chemical Business 287, no.9:34-34
4. Bloomberg. In:. http://www.bloomberg.com/
5. BBI International (2015) U.S. Ethanol Plants. In: .
http://www.ethanolproducer.com/plants/listplants/US/Existing/All
6. DTN/The Progressive Farmer (2015) Daily ethanol rack price. In:.
http://www.dtnprogressivefarmer.com/dtnag/renewable-fuels
7. Sugaya K, Tuse D, Jones JL (1986) Production of acetic acid by Clostridium thermoaceticum
in batch and continuous ferrmentation. Biotechnology and bioengineering 28:678-683
8. Drake HL (1994) Introduction to acetogenesis. In: Drake HL (ed) Acetogenesis. Chapman and
Hall, London, pp 3-60
9. Ljungdahl LG (1986) The autotrophic pathway in acetate synthesis in acetogenic bacteria.
Annual Review of Microbiology 40:415-450
10. Drake HL, Daniel SL (2004) Physiology of the thermophilic acetogen Moorella
thermoacetica. Research in Microbiology 155:422-436
11. Andreesen JR, Schaupp A, Neurauter C, Brown A, Ljungdahl LG (1973) Fermentation of
glucose, fructose, and xylose by Clostridium thermoaceticum: effect of metals on growth yield,
enzymes, and the synthesis of acetate from CO2. Journal of Bacteriology 114:743-751
12. Brumm PJ (1988) Fermentation of single and mixed substrates by the parent and an acid-
tolerant, mutant strain of Clostridium thermoaceticum. Biotechnology and Bioengineering
32:444-450
13. Balasubramanian N, Kim JS, Lee YY (2001) Fermentation of xylose into acetic acid by
Clostridium thermoaceticum. Journal of Applied Biochemistry and Biotechnology 91-93:367-
376
14. Ljungdah LG (2009) A life with acetogens, thermophiles, and cellulolytic anaerobes. Annual
Review of Microbiology 63:1-25

27
15. Pierce E, Xie G, Barabote RD, Saunders E, Han CS, Detter JC, Richardson P, Brettin TS,
Das A, Ljungdahl. Lars G., Ragsdale SW (2008) The complete genome sequence of Moorella
thermoacetica (f. Clostridium thermoacetica). Environmental Microbiology 10:2550-2573
16. Hu P, Rismani-Yazdi H, Stephanopoulos G (2013) Anaerobic CO2 fixation by the acetogenic
bacterium Moorella thermoacetica. American Institute of Chemical Engineers 0:1-8
17. Wang G, Wang DI (1984) Elucidation of growth inhibition and acetic acid production by
Clostridium thermoaceticum. Applied and Environmental Microbiology 47:294-298
18. Lundie Jr. L, Drake HL (1984) Development of a Minimally Defined medium for the
Acetogen Clostridium thermoaceticum. Journal of Bacteriology 159:700-703
19. Bock SA, Fox SL, Gibbons WR (1997) Development of low cost, industrially suitable
medium for the production of acetic acid from Clostridium thermoaceticum. Biotechnology and
Applied Biochemistry 25:117-125
20. Witjitra K, Shah M, Cheryan M (1996) Effect of nutrient sources on growth and acetate
production by Clostridium thermoaceticum. Enzyme and Microbial Technology 19:322-327
21. Shah MM, Cheryan M (1995) Acetate production by Clostridium thermoaceticum in corn
steep liquor media. Journal of Industrial Microbiology 15:424-428
22. Schwartz RD, Keller FA (1982) Acetic acid production by Clostridium thermoaceticum in
pH-controlled batch fermentation at acidic pH. Applied and Environmental Microbiology
43:1385-1392
23. Wang G, Wang DIC (1983) Production of acetic acid by immobilized whole cells of
Clostridium thermoacetica. Applied Biochemistry and Biotechnology 8:491-503
24. Parekh SR, Cheryan M (1994) Continuous production of acetate by Clostridium
thermoaceticum in a cell-recycle membrane bioreactor. Enzyme and Microbial Technology
16:104-109
25. Shah MM, Cheryan M Improvement of productivity in acetic acid fermentation with
clostridium thermoaceticum. 1995 51/52:413-422
26. Brownell JE, Nakas JP (1991) Bioconversion of acid-hydrolyzed poplar hemicellulose to
acetic acid by Clostridium thermoaceticum. Journal of Industrial Microbiology 7:1-6
27. Borden JR, Lee YY, Yoon H (2000) Simultaneous Saccharification and Fermentation of
Cellulosic Biomass to Acetic Acid. Applied Biochemistry and Biotechnology 84-86:963-970
28. Ewanick SM, Bura R, Saddler JN (2007) Acid-catalyzed steam pretreatment of lodgepole
pine and subsequent enzymatic hydrolysis and fermentation to ethanol. Biotechnology and
bioengineering 98:737-746
29. Mohagheghi A, Ruth M, Schell DJ (2006) Conditioning hemicellulose hydrolysates for
fermentation: effect of overliming pH on sugar and ethanol yields. Process Biochemistry
41:1806-1811
30. Waterhous A (2012) Folin-Ciocalteau Micro Method for Total Phenol in Wine. In: .
http://waterhouse.ucdavis.edu/faqs/folin-ciocalteau-micro-method-for-total-phenol-in-wine

28
31. Lundqvist J, Jacobs A, Palm M, Zacchi G, Dahlman O, Stalbrand H (2003) Characterization
of galactoglucomannan extracted from spurce (Picea abies) by heat-fractionation at different
conditions. Carbohydrate Polymers 51:203-211
32. Bura R, Vajzovic A, Doty SL (2012) Novel endophytic yeast Rhodotorula mucilaginosa
strain PTD3 I: production of xylitol and ethanol. Journal of Industrial Microbiology &
Biotechnology 39:1003-1011
29
0 10 20 30 40 50 60 700
5
10
15
20
25
Xylose
Glucose
Arabinose
Galactose
Acetic Acid
Cell
Time (h)
Sugar
s co
nce
ntr
atio
n (
g/L
)
0
5
10
15
20
25A
Ace
tic
acid
and c
ell
conce
ntr
atio
n (
g/L
)
0 10 20 30 40 50 60 700
5
10
15
20
25
Time (h)
Sugar
s co
nce
ntr
atio
n (
g/L
)
Xylose
Glucose
Arabinose
Galactose
Mannose
Acetic Acid
Cell
0
5
10
15
20
25
Ace
tica
cid a
nd c
ell
conce
ntr
atio
n (
g/L
)
B
0 10 20 30 40 50 60 70
0
5
10
15
20
25
Xylose
Glucose
Arabinose
Galactose
Mannose
Acetic Acid
Cell
Time (h)
Su
gar
s co
nce
ntr
atio
n (
g/L
)
0
5
10
15
20
25A
ceti
c ac
id a
nd
cel
l co
nce
ntr
atio
n (
g/L
)C
0 10 20 30 40 50 60 70
0
5
10
15
20
25
Time (h)
Sugar
s co
nce
ntr
atio
n (
g/L
)
Xylose
Glucose
Arabinose
Galactose
Mannose
Acetic Acid
Cell
D
0
5
10
15
20
25
Ace
tic
acid
and c
ell
conce
ntr
atio
n (
g/L
)
Figure 2. Sugars consumption, acetic acid production and cell concentration during fermentation
of A) wheat straw, B) forest residues, C) switchgrass, and D) sugarcane straw hydrolysates using
M. thermoacetica. The error bars indicate standard deviation.

30
Table 2. Summary of process variables during pH-controlled batch fermentation of hydrolysates and controls
Hydrolysate (H),
Synthetic (S)
Furfural
(g/L),
STD
HMF
(g/L), STD
Total
phenolics
(g/L), STD
Initial sugars5
(g/L), STD
Max Cell
Concentration6
(g/L), STD
Acid
produced
(g/L), STD
Yield (%) ,
STD Xyl Glu Ara Gala Man
WS1 – H 0.01(0.01) 0.01(0.01) 2.77 (0.01) 20.34 (0.01) 2.82 (0.01) 3.13 (0.01) 0.56 (0.02) 0.00 23.01 (0.03) 16.90 (0.01) 63.06 (0.53)
WS – S 0.00 0.00 0.00 20.02 (0.07) 3.02 (0.04) 2.61 (0.05) 0.50 (0.05) 0.00 24.50 (0.05) 14.20 (0.01) 54.38 (0.15)
FR2 – H 0.01(0.01) 0.01(0.01) 2.52 (0.01) 20.01 (0.05) 8.87 (0.03) 3.79 (0.02) 7.06 (0.10) 9.17 (0.09) 22.30 (0.28) 18.22 (0.14) 37.59 (0.04)
FR – S 0.00 00 0.00 19.70 (0.07) 9.12 (0.06) 3.89 (0.02) 6.94 (0.06) 9.33 (0.01) 19.30 (0.01) 19.01 (0.02) 38.82 (0.06)
SG3 – H 0.44 (0.01) 0.067 (0.01) 2.92 (0.01) 19.67 (0.03) 2.91 (0.04) 2.62 (0.03) 0.75 (0.01) 0.29 (0.04) 22.30 (0.2) 14.11 (0.01) 53.79 (0.04)
SG – S 0.00 0.00 0.00 20.46 (0.02) 2.11 (0.01) 1.63 (0.01) 0.39 (0.01) 0.04 (0.01) 22.40 (0.3) 13.54 (0.01) 55.02 (0.35)
SCS4 – H 0.01(0.01) 0.01(0.01) 2.80 (0.1) 20.19 (0.23) 1.74 (0.01) 1.96 (0.03) 0.39 (0.01) 0.09 (0.01) 21.40 (0.01) 17.20 (0.02) 70.66 (0.66)
SCS – S 0.00 0.00 0 .00 20.05 (0.01) 1.77 (0.01) 1.59 (0.01) 0.31 (0.01) 0.01 (0.01) 22.60 (0.01) 15.20 (0.01) 64.12 (0.01) 1WS, wheat straw; 2FS, forest residues; 3SG, switchgrass; 4SCS, sugarcane straw; 5Xyl, xylose; Glu, glucose, Ara, arabinose; Man,
mannose 6max cell concentration calculated based on dry cell
Table 3. Summary of process variables during flask fermentation of single sugar to acetic acid, comparing arabinose, galactose and
mannose to glucose during 50 h fermentation
Sugar Concentration (g/L) Acetic acid yield (%) Glucose 22 (0.08) 66 (0.02)
Arabinose 20 (0.08) 0 (0.01)
Galactose 20 (0.06) 0 (0.01)
Mannose 20 (0.08) 0 (0.01)

31
Table 4. Summary of process variables during flask fermentation of mixed sugars, monitoring consumed sugars and produced acetic
acid
Fermentation no. Initial sugar (g/L) / Consumed (%) Cell growth / initial cell
concentration
(g/L)
Max cell
concentration
(g/L), STD
Acid
produced
(g/L), STD
Yield (%)
based on
total sugars,
STD
Ara* Gal* Glu* Xyl* Man*
STD STD STD STD STD
1 3.40 / 39.70 3.50 / 32.90 3.10 / 97.70 3.20 / 100 3.80 / 36.80
mixture of five sugars / 0.2 6.60 (0.01) 4.70 (0.01) 27.00 (0.02) 0.02 / 0.01 0.01 / 0,01 0.01 /0.01 0.01 / 0.01 0.01 / 0.01
2 4.20 / 25.70 4.30 / 14.50 4.10 / 100 3.60 / 100 3.00 / 29.70
1:1 / 7 13.20 (0.01) 6.00 (0.01) 32.00 (0.01) 0.01 / 0.01 0.01 / 0.01 0.01 / 0.01 0.01 / 0.01 0.02 /0.01
*Ara, arabinose; Gal, galactose; Glu, glucose; Xyl, xylose; Man, mannose
Table 5. Summary of process variables during pH-controlled batch fermentation of glucose and xylose mixture, with different
concentration, over 50 h , 75 h and 96 h of fermentation
Fermentation
no.
Initial sugars
(g/L) Total
sugars
(g/L)
Max
cell
conc.*
(g/L)
50 h 75 h 96 h
Acid
produced
(g/L)
Residual sugars
(g/L) Yield
(%)
Acid
produced
(g/L)
Residual sugars
(g/L) Yield
(%)
Acid
produced
(g/L)
Residual sugars
(g/L)
Yield
(%)
Glucose Xylose Glucose Xylose Glucose Xylose Glucose Xylose
1 3.2 19.54 22.7 20.8 13.6 2.8 2.1 59.9 15.8 0 0 69.4 15.8 0 0 69.4
2 20.6 3.1 23.7 20 17.6 0 0 74.6 17.6 0 0 74.6 17.6 0 0 74.6
3 8.7 19.4 28.1 24.5 14.0 6.5 1.3 50.0 18.7 3 0 66.7 20.6 0 0 73.3
4 19.4 9.5 28.9 21.9 21.7 0.8 0 75.1 22.3 0 0 77.1 22.3 0 0 77.1
*Conc., concentration; the reported results are the average of triplicate studies with a standard deviation of ≤ 0.02

32
Chapter 3 – Conclusion and future work
3.1 Conclusion
It was presented for the first time that M. thermoacetica (ATCC 39073) can effectively ferment
xylose and glucose content in water-soluble fraction of steam exploded lignocellulosic biomass
including wheat straw, forest residues, switchgrass, and sugarcane straw to acetic acid. In all
fermentations xylose was the major and the first sugar being consumed. Xylose consumption was
faster in switchgrass and sugarcane straw hydrolysates (35 h) than wheat straw and forest residues
hydrolysates (40 and 45 h respectively). Xylose repressed glucose uptake in which glucose was
metabolized when the majority of xylose was consumed. M. thermoacetica consumed less than
50% of arabinose, galactose and mannose within 72 h of fermentation in a mixture of sugars. The
highest cell biomass was observed in wheat straw hydrolysate (23 g/L) and the lowest in sugarcane
straw hydrolysate (21.4 g/L). The highest acetic acid production based on total sugars was obtained
from sugarcane straw with 71% of theoretical yield while it was 64% in its control. The lowest
acetic acid yield based on total sugars was observed in forest residues with 39% of theoretical. The
incomplete consumption of arabinose, galactose and mannose adversely affected acetic acid yield.
M. thermoacetica demonstrated that it tolerates process derived inhibitory compounds from steam
explosion pretreatment including HMF (0.44 g/L), FF (0.1 g/L) and total phenolics (3 g/L). The
variations observed in acetic acid yields, repressed glucose uptake by xylose, and incomplete
consumption of minor sugars confirmed CCR behavior in M. thermoacetica.

33
3.2 Future work
This research has focused on bioconversion of lignocellulosic biomass to acetate for the production
of acetic acid. The ultimate goal is to establish a viable bioconversion process that is technically
and economically feasible in an industrial setting. Several areas are recommended for future
investigations.
Optimization of fermentation
One area that will improve acetic acid yield is fermentation condition. In this study, all
fermentations were conducted by utilizing CO2 gas and mechanical agitation at 175 RPM.
Additionally, inoculant was grown in a 1:1 ratio of synthetic glucose and xylose. Further
optimizations are essential in the areas of medium composition, stirring and utilization of N2 gas
instead of CO2. Usage of inoculant pre-grown on hydrolysate, lower agitation rate and N2 gas will
provide additional information on how the fermentation process may be optimized.
Detoxifying hydrolysates
There are various detoxification techniques to remove fermentation inhibitors (e.g. furans and
phenolics) from lignocellulosic hydrolysates prior to fermentation. In this study, overliming
method was utilized to detoxify lignocellulosic hydrolysates. Negative side effects of overliming
include sugar loss in hydrolysate and presence of excess salts (Ca2+ and Na+) on microbial growth.
Detoxification of lignocellulosic hydrolysates by activated charcoal is recommended. This is due
to the fact that adsorption of activated charcoal is widely used to remove inhibitors such as furan
and phenolic compounds without significant sugar loss. Thus, higher acetate yield and a more
efficient inhibitors removal may be achieved by hydrolysate treated with activated charcoal.

34
Continuous fermentation process
Various fermentation setups utilizing CSTR and synthetic sugar substrates were investigated in
order to improve productivity in acetic acid fermentation. However, challenges around process
design utilizing lignocellulosic hydrolysate in continuous fermentation still remains. One of the
major difficulties is the accumulation of acetic acid product and inhibitory compounds which
hinders the fermentation process and lowers the yield. In order to address this issue, reverse
electro-enhanced dialysis can be applied to improve the production yield.
3.3 Significance
Lignocellulosic acetic acid is a value added and promising sugar-derived chemical that serves as
a major economic driver for a biorefinery. The ultimate goal is to develop poplar-based biofuels
and biochemicals through a sustainable and economical process. This study has provided
significant insight regarding the bioconversion of lignocellulosic biomass to acetic acid utilizing
Moorella thermoacetica. The following details the significance of the main findings:
The research has demonstrated for the first time the ability of M. thermoacetica to ferment
monomeric sugars in lignocellulosic hydrolysates obtained after steam explosion
pretreatment to acetate.
M. thermoacetica partially consumes arabinose, galactose and mannose in a mixture of
sugars.
In order to obtain an optimum acetic acid yield, it is imperative to have lignocellulosic
hydrolysate with the highest xylose content and the least or no amount of other
lignocellulosic sugars.

35
Steam explosion pretreatment design is a key process that needs to be customized and
applied in two or three steps. First, with low severity factor, collect as much xylose in liquid
stream (hydrolysate) as possible. The next step is to provide a solid stream with glucose
content only. Liquid and solid phases need to be refined to separate arabinose, galactose
and mannose.
Choice of lignocellulosic feedstock also improves acetate yield. Hardwoods and
agricultural residues have a great potential for the production of acetic acid. Softwoods
would not be a good option due to their high content of galactoglucomannan.
References
1. U.S. Energy Information Administration (2014) International energy outlook 2014 DOE/EIA-
0484(2014)
2. Philbrook A, Alissandratos A, Easton CJ (2013) Biochemical processes for generating fuels
and commodity chemicals from lignocellulosic biomass In: Petre M (ed) Environmental
biotechnology - New approaches and prospective applications. InTech, pp 39-64
3. Balan V (2014) Current challenges in commercially producing biofuels from lignocellulosic
biomass. ISRN Biotechnology 2014:1-31
4. US DOE (2015) Lignocellulosic biomass for advanced biofuels and bioproducts: workshop
report, DOE/SC-0170. U.S. Department of Energy Office of Science. In:.
http://genomicscience.energy.gov/biofuels/lignocellulose/.
5. Chandel AK, Singh OV (2011) Weedy lignocellulosic feedstock and microbial metabolic
engineering: advancing the generation of 'Biofuel'. Applied Microbiology and Biotechnology
89:1289-1303
6. Schuth F, Rinaldi R, Meine N, Kaldstrom M, Hilgert J, Rechulski MDK (2014)
Mechanocatalytic depolymerization of cellulose and raw biomass and downstream processing of
the products. Catalysis Today 234:24-30
7. Limayem A, Ricke SC (2012) Lignocellulosic biomass for bioethanol production: current
perspective, potential issues and future prospects. Progress in Energy and Combustion Science
38:449-467
8. Kato DM, Elía N, Flythe M, Lynn BC (2014) Pretreatment of lignocellulosic biomass using
Fenton chemistry. Bioresource Technology 162:273-278
9. Jorgensen H, Kristensen JB, Felby C (2007). Biofuels, Bioproducts and Biorefining 1:119-134

36
10. Sun Y, Cheng J (2002) Hydrolysis of lignocellulosic materials for ethanol production: a
review. Bioresource Technology 83:1-11
11. Christopher L (2012) Adding value prior to pulping: bioproducts from hemicellulose. In:
Akais OC (ed) Global perspectives on sustainable forest management. InTech, pp 225-246
12. Sorke N, Yeats TH, Szemenyei H, Youngs H, Somerville CR (2014) The implication of
lignocellulosic biomass chemical composition for the production of advanced biofuels.
BioScience 64:192-201
13. Gray KA, Zhao L, Emptage M (2006) Bioethanol. Current Opinion in Chemical Biology
10:141-146
14. Magalhaes da Silva SP, Da Costa Lopes AM, Roseiro LB, Bogel-Lukasik R (2013) Novel
pre-treatment and fractionation method for lignocellulosic biomass using ionic liquids . Royal
Society of Chemistry Advances 3:16040-16050
15. Henriksson G (2009) Lignin. In: Ek M, Gellerstedt G, Henriksson G (eds) Pulp and paper
chemistry and technology, 1st edn. Walter de Gruyter, Berlin, pp 121-146
16. Zakzeski J, Bruijnincx P, Jongerius A, Weckhuysen B (2010) The catalytic valorization of
lignin for the production of renewable chemicals. American Chemical Society-Chemical
Reviews 110:3552-3599
17. Sjostrom E (1993) Lignin. In: Sjostrom E (ed) Wood chemistry: fundamentals and
applications, 2nd edn. Academic press, San Diego, pp 71-88
18. Gandolfi S, Ottolina G, Riva S, Fantoni GP, Patel I (2013) Complete chemical analysis of
carmagnola hemp hurds and structural features of its components. BioResources 8:2641-2656
19. Beatson RP (2011). In: Zhu J, Zhang X, Pan X (eds) Sustainable production of fuels,
chemicals, and fibers from forest biomass, Volume 1067 edn. American Chemical Society, pp
279-297
20. Imamura H (1989) Contribution of Extractives to Wood Characteristics. In: Rowe JW (ed)
Natural products of woody plants: chemicals extraneous to the lignocellulosic cell wall. Springer
Berlin Heidelberg, Berlin, pp 843-860
21. Bura R, Chandra R, Saddler J (2009) Influence of xylan on the enzymatic hydrolysis of
steam-pretreated corn stover and hybrid poplar. American Institute of Chemical Engineers
25:315-322
22. Taherzadeh MJ, Karimi K (2008) Pretreatment of lignocellulosic wastes to improve ethanol
and biogas production: a review. International Journal of Molecular Sciences 9:1621-1651
23. Brodeur G, Yau E, Badal K, Collier J, Ramachandran KB, Ramakrishnan S (2011) Chemical
and physicochemical pretreatment of lignocellulosic biomass: a review. Enzyme Research
2011:1-17
24. Gable M, Zacchi G (2007) Pretreatment of lignocellulosic materials for efficient bioethanol
production. Advances in Biochemical Engineering/Biotechnology 108:41-65

37
25. Liu S, Wang Y, Buyo JP, Wang Y, Garver M (2013) Biochemical conversion. In: Stuart PR,
El-Halwagi MM (eds) Integrated Biorefineries: Design, Analysis, and Optimization (Green
Chemistry and Chemical Engineering), 1st edn. CRC Press, Boca Raton, FL, United states of
America, pp 591-650
26. Howard R, Abotsi E, Jansen van Rensburg E, Howard S (2003) Lignocellulose
biotechnology: issues of bioconversion and enzyme production. African Journal of
Biotechnology 2:602-619
27. Szakacs G, Tengerday RP, Nagy V (2006) Cellulases. In: Pandey A, Webb C, Soccol CR,
Larroche C (eds) Enzyme technology. Springer, Delhi, pp 252-272
28. Mussatto SI, Teixeira JA (2010) Lignocellulose as raw material in fermentation processes.
In: Mendez-Vilas A (ed) Current research, technology and education topics in applied
microbiology and microbial biotechnology, Volume 2 edn. Formatex Research Center, Badajoz,
Spain, pp 897-907
29. Zeikus JG (1980) Chemical and fuel production by anaerobic bacteria. Annual Review of
Microbiology 34:423-464
30. Zhang M, Eddy C, Deanda K, Finkelstein M, Picataggio S (1995) Metabolic engineering of a
pentose metabolism pathway in ethanologenic Zymomonas mobilis. American Association for
the Advancement of Science 267:240-243
31. Shin HS, Zeikus JG, Jain MK (2002) Electrically enhanced ethanol fermentation by
Clostridium thermochellum and Saccharomyces cerevisiae. Applied Microbiology and
Biotechnology 58:476-481
32. Drake HL, Daniel SL, Gossner AS (2008) Old Acetogens, New Light. Annals of the New
York Academy of Sciences 1125:100-128
33. Jo JH, Lee DS, Park D, Park JM (2008) Biological hydrogen production by immobilized
cells of Clostridium tyrobutyricum JM1 isolated from a food waste treatment process.
Bioresource Technology 99:6666-6672
34. Lee J, Jang Y, Choi SJ, Im JA, Song H, Cho JH, Seung DY, Papoutsakis ET, Bennett GN,
Lee SY (2012) Metabolic engineering of Clostridium acetobutylicum ATCC 824 for
isoproponal-butanol-ethanol fermentation. Applied and Environmental Microbiology 78:1416-
1423
35. Paes BG, Almeida JRM (2014) Genetic improvement of microorganisms for application in
biorefineries. Chemical and Biological Technologies in Agriculture 1:1-10
36. Buhner J, Agblevor FA (2004) Effect of detoxification of dilute acid corn fiber hydrolysate
on xylitol production. Applied Biochemistry and Biotechnology 119:13-29
37. Almeida JRM, Modig T, Petersson A, Hahn-Hagerdal B, Liden G, Gorwa-Grauslund MF
(2007) Increased tolerance and conversion of inhibitors in lignocellulosic hydrolysates by
Saccharomyces cerevisiae. Chemical Technology and Biotechnology 82:340-349

38
38. Baral NR, Shah A (2014) Microbial inhibitors: formation and effects on aceton-butanol-
ethanol fermentation of lignocellulosic biomass. Applied Microbiology and Biotechnology
98:9151-9172
39. Ballesteros M, Negro M, Manzanares P, Ballesteros I, Saez F, Oliva JM (2007) Fractionation
of Cynara Cardunculus (Cardoon) Biomass by Dilute Acid Pretreatment. Applied Biochemistry
and Biotechnology, 137-140:239-252
40. Jönsson LJ, Alriksson B, Nilvebrant N (2013) Bioconversion of lignocellulose: inhibitors and
detoxification. Biotechnology for Biofuels 6:1-10
41. Behera S, Arora R, Nandhagopal N, Kumar S (2014) Importance of chemical pretreatment
for bioconversion of lignocellulosic biomass. Renewable and Sustainable Energy Reviews
36:91-106
42. Werpy T, Petersen G (2004) Top value added chemicals from biomass, volume I-results of
screening for potential candidates from sugars and synthesis gas DOE/GO-102004-1992
43. Merat N, Godawa C, Gaset A (1990) High selective production of tetrahydrofurfuryl alcohol:
Catalytic hydrogenation of furfural and furfuryl alcohol. Chemical Technology and
Biotechnology, 48:145-159
44. Bozell JJ, Holladay JE, Johnson D, White JF (2007) Top value-added chemicals from
biomass, volume II-results of screening for potential candidates from biorefinery lignin PNNL-
16983
45. Grand View Research I (2014) Acetic acid market analysis and segment forecasts to 2020.
In:. http://www.grandviewresearch.com/industry-analysis/acetic-acid-market
46. Yoneda N, Kusano S, Yasui M, Pujado P, Wilcher S (2001) Recent advances in processes
and catalysts for the production of acetic acid. Applied Catalysis A: General Published by
Elsevier Science B.V. 221:253-265
47. Clark B (2015) Chemical profile, US acetic acid. ICIS Chemical Business 287, no.9:34-34
48. Bloomberg. In:. http://www.bloomberg.com/
49. BBI International (2015) U.S. Ethanol Plants. In:.
http://www.ethanolproducer.com/plants/listplants/US/Existing/All
50. DTN/The Progressive Farmer (2015) Daily ethanol rack price. In:.
http://www.dtnprogressivefarmer.com/dtnag/renewable-fuels
51. Sugaya K, Tuse D, Jones JL (1986) Production of acetic acid by Clostridium thermoaceticum
in batch and continuous fermentation. Biotechnology and bioengineering 28:678-683
52. Drake HL (1994) Introduction to acetogenesis. In: Drake HL (ed) Acetogenesis. Chapman
and Hall, London, pp 3-60
53. Ljungdahl LG (1986) The autotrophic pathway in acetate synthesis in acetogenic bacteria.
Annual Review of Microbiology 40:415-450

39
54. Drake HL, Daniel SL (2004) Physiology of the thermophilic acetogen Moorella
thermoacetica. Research in Microbiology 155:422-436
55. Andreesen JR, Schaupp A, Neurauter C, Brown A, Ljungdahl LG (1973) Fermentation of
glucose, fructose, and xylose by Clostridium thermoaceticum: effect of metals on growth yield,
enzymes, and the synthesis of acetate from CO2. Journal of Bacteriology 114:743-751
56. Brumm PJ (1988) Fermentation of single and mixed substrates by the parent and an acid-
tolerant, mutant strain of Clostridium thermoaceticum. Biotechnology and Bioengineering
32:444-450
57. Balasubramanian N, Kim JS, Lee YY (2001) Fermentation of xylose into acetic acid by
Clostridium thermoaceticum. Journal of Applied Biochemistry and Biotechnology 91-93:367-
376
58. Ljungdah LG (2009) A life with acetogens, thermophiles, and cellulolytic anaerobes. Annual
Review of Microbiology 63:1-25
59. Pierce E, Xie G, Barabote RD, Saunders E, Han CS, Detter JC, Richardson P, Brettin TS,
Das A, Ljungdahl. Lars G., Ragsdale SW (2008) The complete genome sequence of Moorella
thermoacetica (f. Clostridium thermoacetica). Environmental Microbiology 10:2550-2573
60. Hu P, Rismani-Yazdi H, Stephanopoulos G (2013) Anaerobic CO2 fixation by the acetogenic
bacterium Moorella thermoacetica
. American Institute of Chemical Engineers 0:1-8
61. Wang G, Wang DI (1984) Elucidation of growth inhibition and acetic acid production by
Clostridium thermoaceticum. Applied and Environmental Microbiology 47:294-298
62. Lundie Jr. L, Drake HL (1984) Development of a Minimally Defined medium for the
Acetogen Clostridium thermoaceticum. Journal of Bacteriology 159:700-703
63. Bock SA, Fox SL, Gibbons WR (1997) Development of low cost, industrially suitable
medium for the production of acetic acid from Clostridium thermoaceticum. Biotechnology and
Applied Biochemistry 25:117-125
64. Witjitra K, Shah M, Cheryan M (1996) Effect of nutrient sources on growth and acetate
production by Clostridium thermoaceticum. Enzyme and Microbial Technology 19:322-327
65. Shah MM, Cheryan M (1995) Acetate production by Clostridium thermoaceticum in corn
steep liquor media. Journal of Industrial Microbiology 15:424-428
66. Schwartz RD, Keller FA (1982) Acetic acid production by Clostridium thermoaceticum in
pH-controlled batch fermentation at acidic pH. Applied and Environmental Microbiology
43:1385-1392
67. Wang G, Wang DIC (1983) Production of acetic acid by immobilized whole cells of
Clostridium thermoacetica. Applied Biochemistry and Biotechnology 8:491-503
68. Parekh SR, Cheryan M (1994) Continuous production of acetate by Clostridium
thermoaceticum in a cell-recycle membrane bioreactor. Enzyme and Microbial Technology
16:104-109

40
69. Shah MM, Cheryan M Improvement of productivity in acetic acid fermentation with
Clostridium thermoaceticum. 1995 51/52:413-422
70. Brownell JE, Nakas JP (1991) Bioconversion of acid-hydrolyzed poplar hemicellulose to
acetic acid by Clostridium thermoaceticum. Journal of Industrial Microbiology 7:1-6
71. Borden JR, Lee YY, Yoon H (2000) Simultaneous Saccharification and Fermentation of
Cellulosic Biomass to Acetic Acid. Applied Biochemistry and Biotechnology 84-86:963-970

41
Appendices

42
Additional fermentations using synthetic media were conducted at different conditions in
triplicates. Except for sugars content, ingredients used in growth and fermentation media were the
same as described in chapter 2, materials and methods page 24. The following is a general
description of how fermentations were prepared.
Inoculants preparation:
Pre-grown on 1:1 ratio of xylose and glucose for 36 hr at 58°C without agitation, incubated
in dark condition (incubator in Bactron II anaerobic chamber, Sheldon manufacturing,
Inc.).
Low dry cell concentration included 0.2 to 4 g/L of bacterial cell (without harvesting cell
by centrifugation, culture was transferred by 10% (v/v) to fermenter).
High dry cell concentration included 7 g/L of bacterial cell (cells were harvested and
centrifuged at 4000 rpm for 5 minutes, decanted to yield cell pellets concentration of 7
g/L).
Flask fermentations conditions:
Synthetic fermentation media prepared with pH level at 7.5.
Serum vials with 100 mL working volume filled with synthetic fermentation medium and
sparged with filter-sterilized 100% CO2.
Inoculation performed with 10% (v/v) with varied concentration of dry cell (low and high).
Fermentation performed with agitation at 175 rpm (shaking incubator) or no agitation
(static incubator), temperature at 58°C for a period of 48 to 50 h (longer in some cases
from 119 to 168 h).

43
pH-controlled batch fermentations in fermenter 1.3L Bioflo 115 (New Brunswick Scientific
Co., Inc., Edison, NJ) bioreactor:
Synthetic fermentation media prepared with pH level at 7.5.
Fermenter with 500 mL working volume filled with synthetic fermentation medium.
Anaerobic conditions using filter-sterilized 100% CO2 gas sparged into the bottom of the
fermenter vessel continuously prior to inoculation.
Anaerobic condition maintained by sparging 100% CO2 gas at headspace after inoculation
and throughout fermentation.
Inoculation performed with 10% (v/v) and dry cell concentration of 7 g/L.
Fermentations performed with agitation at 175 rpm, temperature at 58°C, pH level
controlled at 6.8 for a period of 72 h or longer (up to 168 h).
1 Flask fermentations utilizing inoculant with low cell concentration (0.2 – 4 g/L)
Preliminary fermentations were conducted to characterize M. thermoacetica with the following
specifications:
Inoculants pre-grown on glucose, xylose, 1:1 ratio of glucose to xylose, mixture of sugars
(xylose, glucose, arabinose, galactose and mannose).
Flask fermentations kept in static incubator (without agitation) and in a dark setting.

44
0 10 20 30 40 50 60 70 80 90 100 1100
5
10
15
20
25
30
35
40
Glucose
Acetic acid
Cell
Time (h)
Su
gar
co
nce
ntr
atio
n (
g/L
)
0
5
10
15
20
25
30
Ace
tic
acid
an
d c
ell
con
cen
trat
ion
(g
/L)
1
0 5 10 15 20 25 30 35 40 450
5
10
15
20 Glucose
Acetic acid
Cell
Time (h)
Su
gar
co
nce
ntr
atio
n (
g/L
)
0
5
10
15
20
25
30
35
Ace
tic
acid
an
d c
ell
con
cen
trat
ion
(g
/L)
2
0 5 10 15 20 25 30 35 40 450
5
10
15
20
25
30
35
40
Glucose
Acetic acid
Cell
Time (h)
Sugar
conce
ntr
atio
n (
g/L
)
0
5
10
15
20
25
30
35A
ceti
c ac
id a
nd c
ell
conce
ntr
atio
n (
g/L
)
3
0 5 10 15 20 25 30 35 40 450
5
10
15
20
25
30
35
40 Glucose
Acetic acid
Cell
Time (h)
Su
gar
co
nce
ntr
atio
n (
g/L
)
0
5
10
15
20
25
30
35
Ace
tic
acid
an
d c
ell
con
cen
trat
ion
(g
/L)
4
0 5 10 15 20 25 30 35 40 45
0
5
10
15
20
25
30
35
40
45
50
55
Glucose
Acetic acid
Cell
Time (h)
Su
gar
co
nce
ntr
atio
n (
g/L
)
0
5
10
15
20
25
30
35
Ace
tic
acid
an
d c
on
cen
trat
ion
(g
/L)5
0 5 10 15 20 25 30 35 40 450
5
10
15
20 Xylose
Acetic acid
Cell
Time (h)
Sugar
conce
ntr
atio
n (
g/L
)
0
5
10
15
20
25
30
35
Ace
tic
acid
and c
ell
conce
ntr
atio
n (
g/L
)6

45
0 5 10 15 20 25 30 35 40 450
2
4
6
8
10 Xylose
Glucose
Acetic acid
Cell
Time (h)
Sugar
conce
ntr
atio
n (
g/L
)
0
2
4
6
8
10
12
14
16
18
20
22
Ace
tic
acid
and c
ell
conce
ntr
atio
n (
g/L
)7
0 5 10 15 20 25 30 35 40 450.0
0.5
1.0
1.5
2.0
2.5
3.0
3.5
4.0 Xylose
Glucose
Arabinose
Galactose
Mannose
Acetic Acid
Cell
Time (h)
Sugar
conce
ntr
atio
n (
g/L
)
0
5
10
15
20
25
Ace
tic
acid
and c
ell
conce
ntr
atio
n (
g/L
)
8
Figure 2. Sugars consumption, acetic acid production and cell concentration during fermentation
of synthetic sugars, including single sugar (plots 1, 2, 3, 4, 5, 6), double sugars (plot 7) and mixture
of sugars (plot 8)
Table 6 Summary of process variables, from preliminary flask fermentations, utilizing inoculant
with low cell content
Graph
Initial sugars
(g/L), STD Fermentation
time (h)
Inoculant
pre-growth
/cell conc.4
(g/L)
Max Cell
Concentration5
(g/L), STD
Acid
produced
(g/L), STD
Yield (%) ,
STD
pH @ 25 ºC
Glucose Xylose Based on
total sugars Before After
1 38.41 (0.01) - 119 glucose / 1 12.94 (0.06) 13.25 (0.01) 34.49 (0.01) 7.5 5.3
1 38.41 (0.01) - 48 glucose / 1 12.94 (0.06) 12.39 (0.01) 32.28 (0.01) 7.5 5.3
2 20.13 (0.02) - 48 glucose / 2 19.11 (0.08) 11.94 (0.02) 53.89 (0.01) 7.5 5.3
3 37.76 (0.08) - 48 glucose / 2 16.80 (0.02) 9.47 (0.02) 25.01 (0.02) 7.5 5.5
4 36.71 (0.08)1 - 48 glucose / 4 16.97 (0.04) 9.06 (0.01) 24.68 (0.01) 7.5 5.5
5 53.93 (0.01) - 48 glucose / 2 14.50 (0.01) 8.60 (0.04) 15.95 (0.02) 7.5 5.6
- - 19.75 (0.02) 48 glucose / 2 2.40 (0.02) < 1 (0.01) < 4 (0.01) 7.5 6.7
6 - 16.26 (0.01) 48 xylose / 1.5 20.33 (0.03) 10.44 (0.01) 64.20 (0.01) 7.5 5.3
7 8.25 (0.01) 7.74 (0.01) 48 1:1 / 1.5 21.17 (0.03) 11.03 (0.04) 68.94 (0.01) 7.5 5.3
8 Mixture of sugars2 48 mixture3 / 0.2 6.64 (0.04) 4.66 (0.03) 27.22 (0.02) 7.5 6.5 1 Inoculation was 20% (v/v); 2 Mixture of glucose 3.31 g/L (+0.10), xylose 3.18 g/L (+0.10), arabinose 3.38 g/L (+0.10), galactose 3.49
g/L (+0.01), and mannose 3.77 g/L (+0.10); 3 Inoculant was pre-grown on mixture of glucose, xylose, arabinose, galactose, and mannose
4 Cell concentration. 5 Maximum cell concentration calculated based on dry cell
Conclusions:
Acetate production in a fermentation period of 48 h was inhibited by low pH level (< 5.7)
and an increase in glucose substrate. M. thermoacetica grows in a pH range of 5.7 to 7.7.
Acetate concentration was decreased from 12 g/L to 8.6 g/L as concentration of glucose

46
was increased from 20 g/L to 60 g/L within 48 h of fermentation. A similar trend was
reported by Balasubramanian and his coworkers based on the effect of initial sugar
concentration on acetate production over a batch fermentation period of 120 h. It was also
confirmed that as unconsumed sugar increased in medium, acetate production yield was
decreased [57].
In fermentation of 40 g/L of glucose, higher cell concentration caused faster sugars uptake
at the beginning of fermentation (24 h). Figure 1, plots 3 and 4 indicate cell concentration
of 4 g/L consumed glucose at a more rapid rate when compared with cell concentration of
2 g/L (1.125 g/L.h and 0.8 g/L.h respectively).
Microbial cells adaptation to xylose and glucose was required to improve acetate yield
(xylose was not metabolized with inoculant pre-grown on glucose only).
Presence of arabinose, galactose and mannose increased duration of sugars uptake and had
negative influence on acetate yield based on total sugars. Figure 1, plots 7 and 8 present
fermentations with average sugar concentration of 16.5 g/L. In a 1:1 ratio of two sugars,
xylose was consumed in 27 h while in 1:1 ratio of five sugars it took 48h. This was an early
indicator of CCR mechanism in M. thermoacetica.
Cell concentration of inoculant pre-grown on a mixture of five lignocellulosic sugars was
very low (0.2 g/L) while it was higher when grown on single sugar or a mixture of xylose
and glucose (2 and 1.5 g/L respectively). This was also indicator of CCR mechanism in M.
thermoacetica.
M. thermoacetica metabolized arabinose, galactose and mannose (23% of total sugars) at
the presence of glucose and xylose with the lowest maximum cell concentration (6.6 g/L).

47
Based on the results of this experiment, it was decided to utilize inoculant pre-grown on a
1:1 ratio of glucose to xylose and with a higher dry cell concentration of 7 g/L for future
fermentations.
2 Flask fermentations utilizing inoculant with high cell concentration (7 g/L)
The following fermentations were conducted to evaluate the effects of different glucose to xylose
ratios on acetate production:
Inoculants were pre-grown on a 1:1 ratio of glucose to xylose.
Inoculation was performed with 10% (v/v) and dry cell concentration of 7 g/L.
All flask fermentations were kept in a shaking incubator at 175 rpm for 50 h.
0 5 10 15 20 25 30 35 40 45 500
5
10
15
20
25
Glucose
Acetic acid
Cell
Time (h)
Sugar
conce
ntr
atio
n (
g/L
)
0
5
10
15
20
25
30
35
Ace
tic
acid
and c
ell
conce
ntr
atio
n (
g/L
)1
0 5 10 15 20 25 30 35 40 45 500
5
10
15
20
25
Xylose
Glucose
Acetic acid
Cell
Time (h)
Sugar
conce
ntr
atio
n (
g/L
)
0
5
10
15
20
25
Ace
tic
acid
and c
ell
conce
ntr
atio
n (
g/L
)
2
0 5 10 15 20 25 30 35 40 45 500
5
10
15
20
25
Xylose
Glucose
Acetic acid
Cell
Time (h)
Sugar
conce
ntr
atio
n (
g/L
)
0
5
10
15
20
25
Ace
tic
acid
and c
ell
conce
ntr
atio
n (
g/L
)3
0 5 10 15 20 25 30 35 40 45 500
5
10
15
20
25
Xylose
Glucose
Acetic acid
Cell
Time (h)
Sugar
conce
ntr
atio
n (
g/L
)
0
5
10
15
20
25
Ace
tic
acid
and c
ell
conce
ntr
atio
n (
g/L
)
4
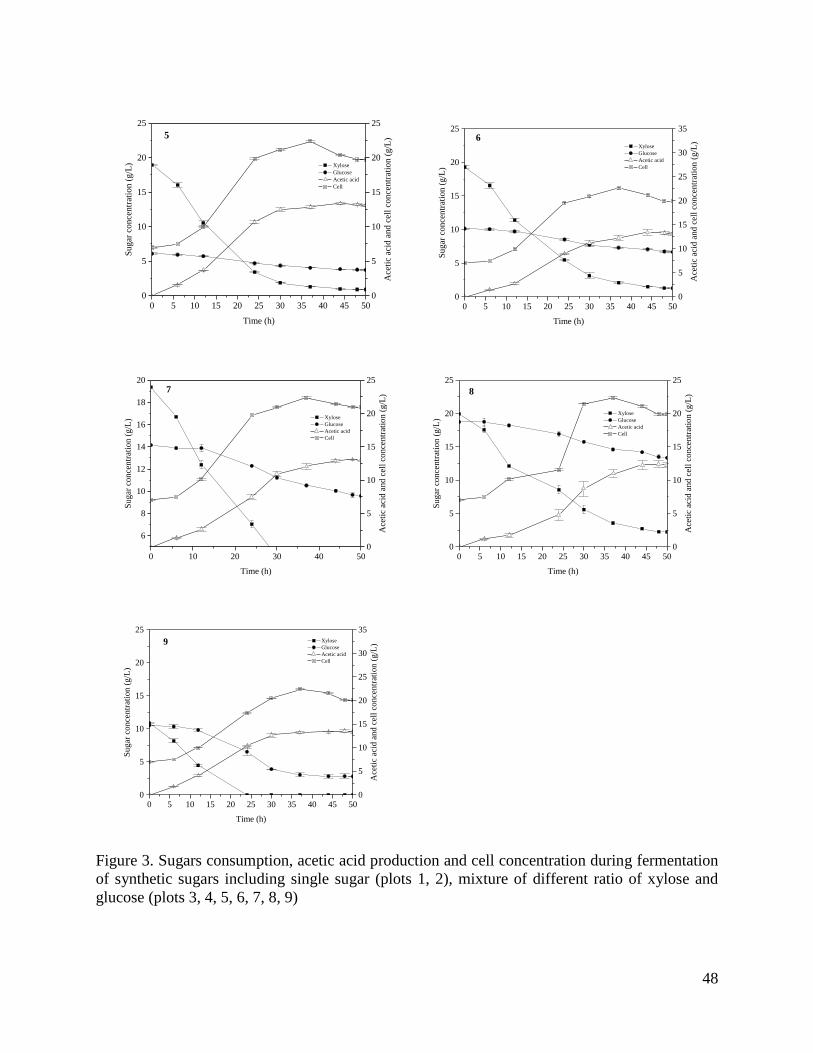
48
0 5 10 15 20 25 30 35 40 45 500
5
10
15
20
25
Xylose
Glucose
Acetic acid
Cell
Time (h)
Sugar
conce
ntr
atio
n (
g/L
)
0
5
10
15
20
25
Ace
tic
acid
and c
ell
conce
ntr
atio
n (
g/L
)5
0 5 10 15 20 25 30 35 40 45 500
5
10
15
20
25
Xylose
Glucose
Acetic acid
Cell
Time (h)
Su
gar
co
nce
ntr
atio
n (
g/L
)
0
5
10
15
20
25
30
35
Ace
tic
acid
an
d c
ell
con
cen
trat
ion
(g
/L)
6
0 10 20 30 40 50
6
8
10
12
14
16
18
20
Xylose
Glucose
Acetic acid
Cell
Time (h)
Sugar
conce
ntr
atio
n (
g/L
)
0
5
10
15
20
25A
ceti
c ac
id a
nd c
ell
conce
ntr
atio
n (
g/L
)7
0 5 10 15 20 25 30 35 40 45 500
5
10
15
20
25
Xylose
Glucose
Acetic acid
Cell
Time (h)
Sugar
conce
ntr
atio
n (
g/L
)
0
5
10
15
20
25
Ace
tic
acid
and c
ell
conce
ntr
atio
n (
g/L
)8
0 5 10 15 20 25 30 35 40 45 500
5
10
15
20
25
Xylose
Glucose
Acetic acid
Cell
Time (h)
Sugar
conce
ntr
atio
n (
g/L
)
0
5
10
15
20
25
30
35
Ace
tic
acid
and c
ell
conce
ntr
atio
n (
g/L
)9
Figure 3. Sugars consumption, acetic acid production and cell concentration during fermentation
of synthetic sugars including single sugar (plots 1, 2), mixture of different ratio of xylose and
glucose (plots 3, 4, 5, 6, 7, 8, 9)

49
Table 7. Summary of process variables during fermentation of synthetic glucose and xylose utilizing inoculant with high dry cell
concentration (7 g/L)
Graph Initial sugars
(g/L)
Sugars
ratio
(G:X)2
Fermentation
Time
(h)
Total sugars
(g/L), STD
Max Cell
conc.4
(g/L), STD
Acid produced
(g/L), STD
Residual sugars
(g/L)
Yield (%),
STD
Glucose
Xylose Glucose Xylose Based on total
sugars
Fig2-1 21.63 (0.08) 0 - 50 21.63 (0.08) 22.56 (0.08) 14.20 (0.20) 3.04 (0.08) 0 65.65 (0.02)
Fig2-2 1.27 (0.10)1 18.12 (0.10) 1:14 50 19.37 (0.02) 22.68 (0.08) 12.91 (0.20) 0.90 (0.05) 0.43 (0.01) 66.64 (0.03)
Fig2-3 4.21 (0.06) 11.85 (0.06) 1:3 50 16.05 (0.06) 22.40 (0.03) 13.20 (0.07) 0.18 (0.05) 0 82.24 (0.02)
Fig2-4 4.17 (0.04) 19.37 (0.01) 1:5 50 23.54 (0.01) 22.55 (0.07) 13.46 (0.24) 2.54 (0.05) 0.68 (0.02) 57.18 (0.02)
Fig 5-23 3.20 (0.01) 19.54 (0.01) 1:6 75 22.74 (0.05) 20.77 (0.02) 15.78 (0.01) 0 0 69.41 (0.01)
Fig 5-23 3.20 (0.01) 19.54 (0.01) 1:6 50 22.74 (0.05) 20.53 (0.02) 13.61 (0.01) 2.84 (0.01) 2.14 (0.01) 59.85 (0.01)
Fig 5-13 20.56 (0.01) 3.09 (0.01) 7:1 75 23.65 (0.01) 20.01 (0.01) 17.63 (0.02) 0 0 74.55 (0.01)
Fig 5-13 20.56 (0.01) 3.09 (0.01) 7:1 50 23.65 (0.01) 20.01 (0.01) 17.63 (0.01) 0 0 74.55 (0.01)
Fig2-5 6.08 (0.10) 18.94 (0.02) 1:3 50 25.02 (0.02) 22.37 (0.08) 13.36 (0.10) 3.73 (0.04) 0.90 (0.02) 53.40 (0.02)
Fig2-6 10.12 (0.10) 19.28 (0.10) 1:2 50 29.40 (0.1) 22.65 (0.10) 13.38 (0.1) 6.66 (0.10) 1.27 (0.10) 45.51 (0.10)
Fig 5-43 8.73 (0.01) 19.36 (0.01) 1.2 75 28.08 (0.01) 24.45 (0.01) 18.72 (0.01) 2.97 (0.01) 0 66.67 (0.01)
Fig 5-43 8.73 (0.01) 19.36 (0.01) 1:2 50 28.08 (0.01) 24.45 (0.01) 14.03 (0.01) 6.53 (0.01) 1.25 (0.01) 49.96 (0.01)
Fig 5-33 19.42 (0;01) 9.47 (0.01) 2:1 75 28.89 (0.01) 21.92 (0.01) 22.27 (0.01) 0 0 77.07 (0.02)
Fig 5-33 19.42 (0.01) 9.47 (0.01) 2:1 50 28.89 (0.01) 21.92 (0.01) 21.70 (0.01) 0.84 (0.01) 0 75.11 (0.02)
Fig2-7 14.16 (0.06) 19.37 (0.01) 1:4 50 33.52 (0.01) 22.40 (0.20) 13.10 (0.03) 9.58 (0.03) 1.78 (0.02) 39.08 (0.01)
Fig2-8 18.74 (0.04) 19.94 (0.01) 1:1 50 38.67 (0.01) 22.38 (0.10) 12.40 (0.05) 13.35 (0.0.2) 2.25 (0.04) 32.07 (0.02)
Fig2-9 10.52 (0.01) 10. 84 (0.06) 1:1 50 21.35 (0.05) 22.38 (0.08) 13.50 (0.03) 2.80 (0.01) 0 63.23 (0.01) 1 Presence of glucose is due to inoculant growing on 1:1 ratio of glucose and xylose 2 Glucose to xylose ratio 3 Highlighted rows: batch fermentations conducted in pH- controlled fermenter 4 Maximum cell concentration, calculated based on dry cell

50
Conclusions:
In a 50 h fermentation period, sugar ratios did not have any effect on cell growth.
Acetate yield based on total sugars was negatively affected by an increase in unconsumed
sugars over a 50 h fermentation period. As an example, using a 1:3 glucose to xylose ratio,
fermentation with 16 g/L of initial sugars resulted in a 29% higher acetate yield (based on
total sugars) than another fermentation with 25 g/L of total sugars (Figure 2: plots 3 and
5).
Sugars ratio had a significant influence on acetic acid yield based on total sugars. Higher
glucose to xylose ratios of 2:1 and 7:1 (Figure 5: plots 1and 3) resulted in a higher acetic
acid yield (25% and 15% respectively over 50 h fermentation) than similar ratios of xylose
to glucose (Figure 5: plots 2 and 4). This is an indicator of CCR mechanism in M.
thermoacetica.
Results from pH-controlled batch fermentations during a 50 h fermentation period are
comparable with their corresponding flask fermentations:
i. Figure 5, plot 2 to Figure 2, plot 4 (acetic acid yield based on total sugars 60% and
57 % respectively).
ii. Figure 5, plot 4 to Figure 2, plot 6 (acetic acid yield based on total sugars 50% and
45.5 % respectively).
2.1 Flask fermentations - comparison of arabinose, galactose and mannose to
glucose
The following fermentations were conducted to compare fermentation of arabinose, galactose and
mannose with glucose:
Inoculants were pre-grown on a 1:1 ratio of glucose to xylose.

51
Inoculation was performed with 10% (v/v) and dry cell concentration of 7 g/L.
All flask fermentations were kept in a shaking incubator at 175 rpm for 50 h.
Table 8. Summary of process variables during flask fermentation of single sugar-comparing
arabinose, galactose and mannose with glucose
Type of sugar Initial sugar
(g/L), STD
Max Cell
conc.1
(g/L), STD
Acid produced
(g/L), STD
Acetic acid yield
based on total sugar
(%), STD
Glucose 21.63 (0.08) 22.56 (0.08) 14.20 (0.20) 65.64 (0.02)
Arabinose 20 (0.01) 0 0 0
Galactose 20 (0.01) 0 0 0
Mannose 20 (0.01) 0 0 0 1 Maximum cell concentration, calculated based on dry cell
Conclusions:
In a flask fermentation of single sugar (20g/L), M. thermoacetica was not able to
metabolize arabinose, galactose and mannose. Clusters of dead cells were formed during
fermentation of these three synthetic sugars.
2.2 Flask fermentations of synthetic sugars - comparison of CO2 to N2 gases
Fermentations with the following specifications were conducted to evaluate the effects of
anaerobic gases (CO2 and N2) on acetate production:
Prior to fermentation, flasks were sparged with filter-sterilized 100% CO2 and 100% N2.
Inoculants were pre-grown on a 1:1 ratio of xylose to glucose.
Inoculation was performed with 10% (v/v) and dry cell concentration of 7 g/L.
All flask fermentations were kept in a static incubator and a dark setting space for 159 h.

52
0 15 30 45 60 75 90 105 120 135 150
0
5
10
15
20
25
Xylose
Glucose
Arabinose
Galactose
Mannose
Acetic Acid
Cell
Time (h)
Su
gar
co
nce
ntr
atio
n (
g/L
)
0
5
10
15
20
25
30
35
Ace
tic
Aci
d ,
Bio
mas
s C
ell
(g/L
)
N2 gas
0 15 30 45 60 75 90 105 120 135 150
0
5
10
15
20
25
Xylose
Glucose
Arabinose
Galactose
Mannose
Acetic Acid
Cell
Time (h)
Su
gar
co
nce
ntr
atio
n (
g/L
)
0
5
10
15
20
25
30
35
Ace
tic
Aci
d ,
Bio
mas
s C
ell
(g/L
)
CO2 gas
Figure 4. Sugars consumption and acetic acid production during flask fermentation utilizing
synthetic sugars model for sugarcane straw hydrolysate comparison of N2 and CO2 gases
Table 9. Summary of process variables during flask fermentation-comparison of N2 and CO2
gases
Flask
fermentation
Initial sugar
(g/L), STD
Max Cell
Concentration1
(g/L), STD
Acid
produced
(g/L), STD
Yield (%) , STD
Based on
total sugar
Based on
consumed
sugar
Under N2 25.30(0.04) 30.75 (0.03) 12.21 (0.40) 48.29 (0.1) 59.13 (0.50)
Under CO2 25.30(0.04) 30.50 (0.05) 11.76 (0.07) 46.56 (0.21) 59.27 (0.40) 1 Maximum cell concentration, calculated based on dry cell
Conclusion:
Results from flask fermentations confirmed that N2 gas can be a good substitution for CO2
gas in order to maintain anaerobic condition.
2.3 Fermentation of CO2 gas as carbon source instead of sugars
Fermentations with the following specifications were conducted to evaluate the effect of CO2 as a
carbon source:
Fermentation medium was prepared without sugar.
Inoculants were pre-grown on a 1:1 ratio of glucose to xylose.
Inoculation was performed with 10% (v/v) and dry cell concentration of 7 g/L.

53
Fermentation was conducted in fermenter with agitation at 175 rpm, temperature at 58°C,
pH level controlled at 6.8 for a period of 168 h.
0 20 40 60 80 100 120 140 160
0
1
2
3
4
5
6
7
8
Acetic acid
Cell
Time (h)
Ace
tic
acid
(g
/L)
No sugar
0
1
2
3
4
5
6
7
8
Bio
mas
s C
ell
(g/L
)
Figure 5. Acetic acid production during flask fermentation of synthetic medium without sugar
Conclusion:
M. thermoacetica metabolized CO2 gas in which maximum acetic acid production was 0.4
g/L within168 h of fermentation.
3 pH-controlled batch fermentations of glucose and xylose
Effect of different ratios of glucose to xylose was investigated. Fermentations were conducted with
the following specifications:
Inoculants were pre-grown on a 1:1 ratio of glucose to xylose.
Inoculations were performed with 10% (v/v) and dry cell concentration of 7 g/L.
Fermentations were conducted in fermenter with agitation at 175 rpm, temperature at 58°C,
pH level controlled at 6.8 for different periods of time (75h, 96h, and 166 h).

54
0 10 20 30 40 50 60 70
0
5
10
15
20
Glucose
Xylose
Acetic acid
Cell
Time (hr)
Su
gar
co
nce
ntr
atio
n (
g/L
)1
0
5
10
15
20
Ace
tic
acid
an
d c
ell
con
cen
trat
ion
(g
/L)
0 20 40 600
5
10
15
20
Glucose
Xylose
Acetic acid
Cell
Time (h)
Sugar
conce
ntr
atio
n (
g/L
)
0
5
10
15
20
Ace
tic
acid
and c
ell
conce
ntr
atio
n (
g/L
)
2
0 10 20 30 40 50 60 70 80 90
0
5
10
15
20
Glucose
Xylose
Acetic acid
Cell
Time (hr)
Su
gar
co
nce
ntr
atio
n (
g/L
)
3
0
5
10
15
20A
ceti
c ac
id a
nd
cel
l co
nce
ntr
atio
n (
g/L
)
0 10 20 30 40 50 60 70 80 90
0
5
10
15
20
25
Glucose
Xylose
Acetic acid
Cell
Time (h)
Su
gar
co
nce
ntr
atio
n (
g/L
)
4
0
5
10
15
20
25
Ace
tic
acid
an
d c
ell
con
cen
trat
ion
(g
/L)
0 20 40 60 80 100 120 140 160
0
5
10
15
20
25
30
Glucose
Acetic acid
Cell
Time (h)
Su
gar
co
nce
ntr
atio
n (
g/L
)
5
0
5
10
15
20
25
30
Ace
tic
acid
an
d c
ell
con
cen
trat
ion
(g
/L)
0 20 40 60 80 100 120 140 160
0
5
10
15
20
25
30
35
40
45
50
Glucose
Acetic acid
Cell
Time (hr)
Su
gar
con
cen
tra
tion
(g
/L)
0
5
10
15
20
25
30
35
40
Ace
tic a
cid
an
d c
ell
con
cen
tra
tion
(g
/L)6
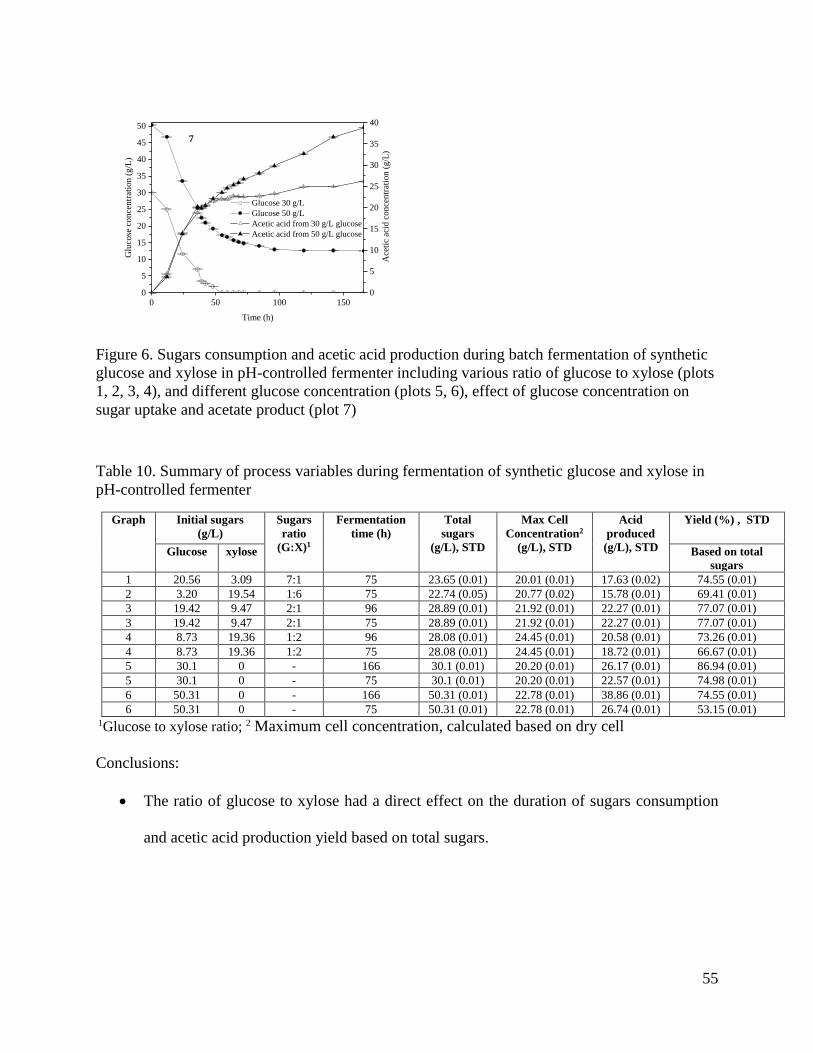
55
0 50 100 1500
5
10
15
20
25
30
35
40
45
50
Glucose 30 g/L
Glucose 50 g/L
Acetic acid from 30 g/L glucose
Acetic acid from 50 g/L glucose
Time (h)
Glu
cose
co
nce
ntr
atio
n (
g/L
)
7
0
5
10
15
20
25
30
35
40
Ace
tic
acid
co
nce
ntr
atio
n (
g/L
)
Figure 6. Sugars consumption and acetic acid production during batch fermentation of synthetic
glucose and xylose in pH-controlled fermenter including various ratio of glucose to xylose (plots
1, 2, 3, 4), and different glucose concentration (plots 5, 6), effect of glucose concentration on
sugar uptake and acetate product (plot 7)
Table 10. Summary of process variables during fermentation of synthetic glucose and xylose in
pH-controlled fermenter
Graph Initial sugars
(g/L)
Sugars
ratio
(G:X)1
Fermentation
time (h)
Total
sugars
(g/L), STD
Max Cell
Concentration2
(g/L), STD
Acid
produced
(g/L), STD
Yield (%) , STD
Glucose xylose Based on total
sugars
1 20.56 3.09 7:1 75 23.65 (0.01) 20.01 (0.01) 17.63 (0.02) 74.55 (0.01)
2 3.20 19.54 1:6 75 22.74 (0.05) 20.77 (0.02) 15.78 (0.01) 69.41 (0.01)
3 19.42 9.47 2:1 96 28.89 (0.01) 21.92 (0.01) 22.27 (0.01) 77.07 (0.01)
3 19.42 9.47 2:1 75 28.89 (0.01) 21.92 (0.01) 22.27 (0.01) 77.07 (0.01)
4 8.73 19.36 1:2 96 28.08 (0.01) 24.45 (0.01) 20.58 (0.01) 73.26 (0.01)
4 8.73 19.36 1:2 75 28.08 (0.01) 24.45 (0.01) 18.72 (0.01) 66.67 (0.01)
5 30.1 0 - 166 30.1 (0.01) 20.20 (0.01) 26.17 (0.01) 86.94 (0.01)
5 30.1 0 - 75 30.1 (0.01) 20.20 (0.01) 22.57 (0.01) 74.98 (0.01)
6 50.31 0 - 166 50.31 (0.01) 22.78 (0.01) 38.86 (0.01) 74.55 (0.01)
6 50.31 0 - 75 50.31 (0.01) 22.78 (0.01) 26.74 (0.01) 53.15 (0.01) 1Glucose to xylose ratio; 2 Maximum cell concentration, calculated based on dry cell
Conclusions:
The ratio of glucose to xylose had a direct effect on the duration of sugars consumption
and acetic acid production yield based on total sugars.

56
Total sugar consumption was considerably quicker with higher glucose to xylose ratio (7:1
and 2:1 sugars ratios took 35 and 55 h respectively) than lower glucose to xylose ratio (1:6
and 1:2 sugars ratios, took 65 and 96 h respectively).
In a fermentation period of 166 h, initial glucose concentration of 30 g/L resulted in 12.4%
higher acetate yield based on total sugar when compared to an initial glucose of 50 g/L.
However, produced acetate was 13 g/L lower. Figure 5, plot 7 shows this is due to the
unconsumed glucose in fermentation with 50 g/L of glucose (after 100 h of fermentation,
12.5 g/L of glucose not touched by the bacterial strain). Lower glucose concentration
should be employed to avoid inhibition of substrate consumption.
In a 60 h fermentation period, glucose uptake is similar between initial sugar concentrations
of 30 g/L and 50 g/L (Figure 5, plot 7)