zoonotic protozoa: from land to sea
TRANSCRIPT

Zoonotic protozoa: from land to seaRonald Fayer1, Jitender P. Dubey2 and David S. Lindsay3
1Environmental Microbial Safety Laboratory, Animal and Natural Resources Institute, Agricultural Research Service,
United States Department of Agriculture, Beltsville, MD 20705, USA2Animal Parasitic Diseases Laboratory, Animal and Natural Resources Institute, Agricultural Research Service,
United States Department of Agriculture, Beltsville, MD 20705, USA3Center for Molecular Medicine and Infectious Diseases, Department of Biomedical Sciences and Pathobiology,
Virginia-Maryland Regional College of Veterinary Medicine, Virginia Tech, 1410 Prices Fork Rd, Blacksburg, VA 24061-0342, USA
Attention to worldwide pollution of the coastal marine
environment has focused primarily on toxic algal
blooms and pathogenic bacteria that multiply in
nutrient-rich waters. However, massive but unseen
amounts of feces from humans, their pets, and their
domesticated animals are discharged, dumped, or
carried in runoff, bringing encysted zoonotic protozoan
parasites to estuaries and coastal waters. Here, they
contaminate bathing beaches, are filtered and concen-
trated by shellfish eaten by humans and marine
mammals, and infect a wide range of marine animal
hosts, resulting in morbidity and mortality to some
populations. This review addresses the extent of
contamination and the animals affected by three genera
of important zoonoticprotozoa:Giardia,Cryptosporidium
and Toxoplasma.
Giardia, Cryptosporidium and Toxoplasma are foundworldwide and are among the most frequently reportedparasites of humans and animals. All are widely dispersedand have been associated with outbreaks of infectionresulting from drinking contaminated surface water.Giardia cysts and Cryptosporidium oocysts are trans-mitted by the fecal–oral route from humans and animals.Toxoplasma gondii oocysts are transmitted from cats tohundreds of warm-blooded vertebrate species. Althougheach infected person or animal often excretes largenumbers of these encysted parasites, relatively few arerequired to initiate an infection. Furthermore, they aresmall, buoyant, resistant to most commonly used disin-fectants, and can survive in moist environments for weeksor months, even when exposed to a wide range oftemperatures and salinities. These traits can contributeto rapid and widespread dispersal, especially after rainfall[1]. Rainfall-initiated runoff from urban, suburban andrural landscapes, and wastewater outfalls, can carryencysted protozoa into untreated waters used for agricul-ture, recreation and, in some locations, for drinking.Recent evidence indicates that these encysted protozoanpathogens are also in coastal waters, having contaminateda variety of shellfish and infected many species of marinemammals. These findings raise concern for the health ofanimals in coastal waters and for humans who eat rawshellfish and who use these waters for recreation.
Corresponding author: Ronald Fayer ([email protected]).Available online 3 September 2004
www.sciencedirect.com 1471-4922/$ - see front matter Q 2004 Elsevier Ltd. All rights reserved
Species and hosts
Giardia duodenalis, the species that infects humans,consists of seven assemblages (genotypes). Among them,assemblage AI is found in humans, livestock, dogs, cats,beavers and other animals; assemblage AII is found onlyin humans [2]; and assemblage B is found in humans,beavers, dogs, muskrats and other animals [2,3]. Trans-mission from humans to beavers, dogs and muskratssuggests some Giardia are zoonotic, and similar genesequences among isolates support this possibility. Assem-blages C and D are found primarily in canids, andassemblages E, F and G, found primarily in hoofedlivestock, cats and rats, respectively, have not beenfound in human infections.
Several Cryptosporidium species infect humans:Cryptosporidium hominis (formerly Cryptosporidiumparvum genotype 1), C. parvum, Cryptosporidium melea-gridis, Cryptosporidium felis, Cryptosporidium canis,Cryptosporidium muris, cervine genotype and pig geno-type I [4]. Most infections are associated with C. hominisand C. parvum, with other species found predominantly inother hosts. Cryptosporidium hominis has been detectedin a few monkeys, sheep and a marine mammal (dugong),whereas C. parvum has been found in numerous cattle,sheep and deer, as well as in mice and pigs [4].
Three routes of transmission of T. gondii includetransmission through oocysts shed in the feces of wildand domesticated felids, ingestion of tissue cysts frommeat, and congenital transmission. The degree of trans-mission to humans and animals by each of these routes isnot always entirely clear and, although the oocyst mightappear the most likely source of infection for non-carnivores, the extent of vertical transmission in mosthost species is unknown and should not be underesti-mated [5]. All methods of transmission are possible, butoocysts are the only stage that enter the environment andremain infectious for up to a year under moist temperateconditions while being widely dispersed in large numbers.Within the species T. gondii, strains are based onpathogenicity in mice, and specific genetic markers haveconfirmed a clonal population with three dominantgenotypes (types I, II and III) evolved from sexualrecombination in felid definitive hosts [6–8]. Additionalgenotypes are known but appear to be less common. TypesI and II have been identified in human infections but thefull genetic diversity of isolates in humans is unknown.
Review TRENDS in Parasitology Vol.20 No.11 November 2004
. doi:10.1016/j.pt.2004.08.008

Review TRENDS in Parasitology Vol.20 No.11 November 2004532
Waterborne outbreaks and prevalence in humans
Giardia cysts have been detected in O2350 samples ofsurface water from eight countries at nearly five cysts perliter and in 21–100% of the samples examined [9]. In theUSA, from 1984 to 1994, 18 drinking-water-associatedoutbreaks of giardiasis affected 3994 individuals [10],whereas an estimated 2.8!108 cases occur globally eachyear [11].
Worldwide, Cryptosporidium oocysts have been foundin ground water, lakes, rivers, estuaries and ocean water[12]. In over 3700 surface water samples from 11countries, 0–100% of specimens contained 0–252.7 oocystsper liter [9]. Of 69 cryptosporidiosis outbreaks from 1983to 1999 in North America, the UK and Japan, 45 wererelated to contaminated drinking water. The largestoutbreak, from contaminated lake water, affected anestimated 403 000 individuals [12]. Cryptosporidiosishas been reported in over 50 countries. Seroprevalencerates of 25–35% in industrialized countries increase to64% in Latin America, and 42% and 57.5% in China andBrazil, respectively, where water purification and sani-tation standards are lower [13]. A 44% seroprevalenceamong dairy farmers versus 24% in others suggests azoonotic link [13].
A toxoplasmosis outbreak in 31 military troops from theUSA was associated with drinking water from a junglepond in Panama [14]. A municipal water system that usedunfiltered, chloraminated surface water was the likelysource of a community-wide outbreak of toxoplasmosisinvolving O100 individuals in Canada [15]. The epidemiccurve appeared bimodal, with peaks preceded byincreased rainfall and turbidity in the implicated reser-voir. Toxoplasmosis has been reported worldwide. In theUSA and UK, an estimated 16–40% of people acquireT. gondii infections, whereas, in Central and SouthAmerica and continental Europe, infection estimatesreach 50–80% [16–18].
Fecal production and fecal pathogens in the
environment
Pathogenic protozoa enter the environment in feces fromland-dwellers – both human and non-human hosts. Propermanagement, treatment and dispersal of human feces andanimal manures are essential for clean and safe drinkingand recreational water. Recreational water also includesestuaries and coastal waters. The enormity of the problemof keeping water clean and safe might seem overwhelmingwhen one considers the volume of feces that enters theenvironment. Although, from space, the Earth appears tobe covered in large part by water and is seen as a brightblue sphere, closer inspection would reveal human andanimal feces spread over much of the planet. On average,each of the current 6.35 billion human inhabitantsproduces w200 g of feces daily, bringing the annual globaldeposit of human feces to w232 million metric tons (Mg).Most is disposed with little or no treatment in under-developed countries before disposal into rivers or coastalwaters, whereas most is aerobically digested in developedcountries before such dispersal.
Worldwide domesticated livestock populations, esti-mated at 1.5 billion cattle and buffalo, 878 million pigs,
www.sciencedirect.com
and 1.7 billion sheep and goats, annually produce w3.6billion Mg, 228 million Mg and 408 million Mg of manure,respectively [19]. Most is deposited on fields with little orno treatment except in places that enforce manuremanagement programs. Other estimates of livestockmanure production reach 5.45 billion Mg per year, withbeef cattle accounting for w55%.
Cyst excretion can approach 107 Giardia cysts per gramof feces [9]. The median infectious dose is thought to bebetween 25 and 100 cysts, but volunteers became infectedafter ingesting as few as ten cysts [9]. Zoonoticassemblage A has been found in dairy cattle manure[20] and, in some farms, this constitutes as much as45% of Giardia isolates [20].
Infected persons and neonatal calves can excrete109–1010 C. parvum oocysts over approximately a week.Yet, in one study, as few as 30 oocysts initiated an infectionin a human volunteer, and the median infective dose was132 oocysts [21]. Although Cryptosporidium oocysts arefrequently identified by microscopy in feces from neonatalruminants, including cattle, sheep, goats and farmed deer,gene sequencing of oocysts from nearly 1000 dairy calvesdemonstrated that most pre-weaned calves had thezoonotic species C. parvum, whereas virtually all post-weaned calves had other non-zoonotic genotypes [22].
The worldwide population of felids, the only source ofT. gondii oocysts, is unknown, but it is estimated that w60million pets and 40 million free-ranging cats live in theUSA. These and other felid species such as bobcats,jaguars, lions and tigers are found worldwide. Although adomesticated cat might defecate only 25 g of feces per day,and the shedding period for Toxoplasma oocysts might lastonly a few days during the life of a cat, tens of millions ofoocysts can be excreted per day and as few as oneoocyst has been infective for mice and pigs [23]. Theminimum infectious dose for humans is unknown butthought to be low.
All other wild and domesticated animals contributeadditional trillions of kg to the global accumulation offeces. This results in a sum accumulation of an incalcul-able number of pathogens.
Contamination of food from polluted fresh and sea
water
Of waters used to irrigate vegetables in the USA andseveral Central American countries, 60% tested positivefor Giardia cysts and 36% tested positive for Crypto-sporidium oocysts [24]. Giardia cysts have been found oncoriander, carrots, mint, radishes and potatoes irrigatedwith wastewater [11], and outbreaks related to contami-nated fruits and vegetables have been reported. Crypto-sporidium oocysts were found on cilantro, lettuce,radishes, tomatoes, cucumbers and carrots in CostaRica, with the highest contamination during the rainyseason when irrigation water from rivers received indus-trial, domestic and agricultural wastewater [25]. Simi-larly, in Peru, Cryptosporidium oocysts were detected on14.5% of 110 fresh vegetables collected at 13 markets, andlater at 16.5% of 62 vegetables from 15 markets [26]. Anoutbreak of cryptosporidiosis was related to eating freshgreen onions. Surface water from many agricultural sites

Review TRENDS in Parasitology Vol.20 No.11 November 2004 533
eventually enters estuaries and coastal waters. Giardiacysts and Cryptosporidium oocysts have been found inshellfish in fresh, estuarine and coastal waters worldwide(Table 1). However, despite widespread and heavy con-tamination of shellfish in some areas, no related outbreaksor individual cases have been reported.
Toxoplasma gondii provides the most direct evidencethat encysted protozoa enter the marine environmentfrom terrestrial sources. Toxoplasmosis can be acquiredfrom ingestion of oocysts or from tissue cysts in raw orundercooked meat. The link between T. gondii oocystscontaminating the marine environment and humantoxoplasmosis acquired as a food-borne pathogen isfound in Inuit women of child-bearing age in northernCanada [27]. Nearly 50% of the women tested wereserologically positive for T. gondii. Of 22 responders to aquestionnaire, those seropositive to T. gondii were fourtimes more likely to have eaten dried seal meat and sixtimes more likely to have eaten seal liver than seronega-tive women. Toxoplasma gondii has been detected in sealsfrom several geographic areas (Table 2).
Giardia, Cryptosporidium and Toxoplasma in marine
waters
Encysted protozoans originating in human and animalfeces are transported in runoff from agricultural, sub-urban and urban land surfaces, wastewater dischargesand other sources to rivers and streams, which carrycontaminated sediments to estuaries and eventually tocoastal waters. Despite the fact that many countriesdispose of raw sewage and dredged sediments fromshipping lanes in coastal waters, there are no mandatedlocal or national monitoring programs for protozoan
Table 1. Cryptosporidium detected in contaminated shellfish
Location Species (common name)
Sligo area, Ireland Mytilus edulis (Common mussel)
Chesapeake Bay, MD, USA Crassostrea virginica (Virginia or Eastern oy
Chesapeake Bay, USA Ischadium recurvum (Bent mussel)
Chesapeake Bay, USA C. virginica
Galicia, Spain Dosinia exoleta (Rayed artemis)
Venerupis (Ruditapes) rhomboideus
(Carpet shell clam)
Venerupis (Ruditapes) pullastra (Venus clam
Venus verrucosa (Mediterranean common c
Mytilus galloprovincialis (Mediterranean m
Ostrea edulis (Native, common or flat oyste
Italy Venerupis (Ruditapes) phillipinarum
(Japanese littleneck)
UK O. edulis
Galicia, Spain M. galloprovincialis
Cerastoderma edule (Common cockle)
St Lawrence River, Canada Dreissena polymorpha (Zebra mussel)
Chesapeake Bay, USA C. virginica
Atlantic coast, USA C. virginica
Portugal C. edule
Scrobicularia plana (Soft shell clam)
M.edulis
Venerupis (Tapes) decussates (Carpet shell
Donax sp. (Donax clam)aAbbreviations: B, bioassay in mice; IFA, immunofluorescence microscopy; M, molecul
www.sciencedirect.com
parasites in estuaries and coastal waters where they caninfect marine wildlife, enter the food chain and potentiallyinfect humans during recreational activities. However, afew studies indicate they are present.
Marine waters near sewage outfalls in Mamala Bay afew kilometers from Waikiki bathing beach in Hawaii [28]and off the coast of Panama [9] were found to containnumerous Giardia cysts and Cryptosporidium oocysts.High numbers of Cryptosporidium oocysts were detectedin marine waters off the coast of San Juan, Puerto Rico[29]. Tidal regions of the Georges River, which isimportant for recreation and shellfish production forSydney, Australia, had elevated concentrations of Giardiaand Cryptosporidium from rainfall and sewage overflowsafter wet weather events [30]. Some areas of Hong Kongdischarge large quantities of sewage into surroundingmarine waters mostly after preliminary or primarytreatment [31]. Of the Hong Kong beaches found suitablefor bathing, 41% of water samples contained Giardia cystsand, of beaches found unsuitable for bathing, 64% hadGiardia cysts, all of which is significantly linked to sewagepollution [31]. Toxoplasma gondii oocysts have not beenreported in coastal waters.
Longevity of encysted protozoa in sea water
Giardia cysts have not been experimentally examined todetermine their longevity in sea water, but infections inmarine mammals suggest they are somewhat resistant atleast to exposure at low salinities (Table 3). Cryptospor-idium oocysts can survive in sea water under a variety oflaboratory conditions for long periods of time. Bioassays inmice indicated that oocysts remained infectious afterbeing held at 10 parts per thousand (ppt) salinity at
No. positive/no. examined Detection
methoda
Ref.
1/3 sites IFA [57]
ster) 6/6 noncommercial sites;
5-18/30 oysters per site
IFA [34]
2/2 sites IFA [58]
5/7 commercial sites;
0-29/30 oysters/site
IFA [59]
15/29 shellfish IFA [60]
)
lam)
ussel)
r)
5/8 clams IFA [60]
1/1 oyster IFA [60]
2/9 sites IFA, B, M [61]
1/9 sites IFA, B, M [61]
514 pooled mussels;
positive finding
IFA, M [62]
7 sites, 8 collections/site;
all sites positive at each collection
IFA, B, M [1]
24/37 sites IFA, B, M [38]
4/26 cockles IFA [63]
1/10 clams IFA [63]
3/32 mussels IFA [63]
clam) 3/18 clams IFA [63]
2/5 clams IFA [63]
ar methods (detected Cryptosporidium parvum).
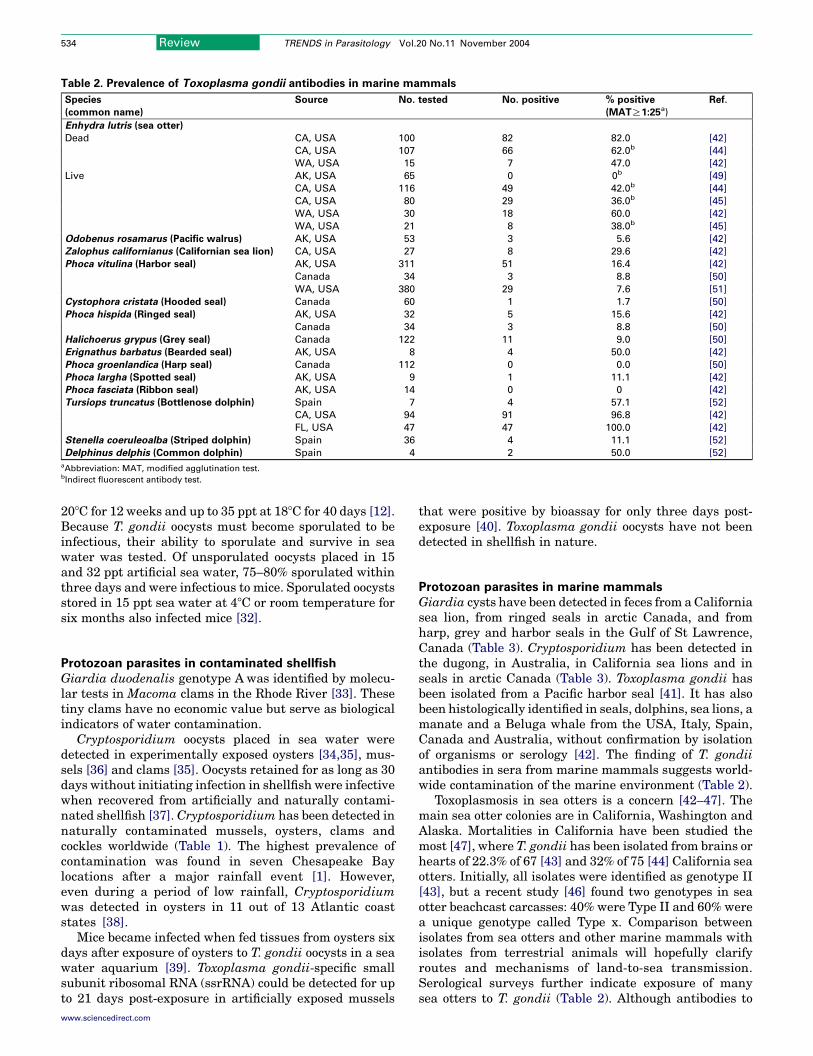
Table 2. Prevalence of Toxoplasma gondii antibodies in marine mammals
Species
(common name)
Source No. tested No. positive % positive
(MATR1:25a)
Ref.
Enhydra lutris (sea otter)
Dead CA, USA 100 82 82.0 [42]
CA, USA 107 66 62.0b [44]
WA, USA 15 7 47.0 [42]
Live AK, USA 65 0 0b [49]
CA, USA 116 49 42.0b [44]
CA, USA 80 29 36.0b [45]
WA, USA 30 18 60.0 [42]
WA, USA 21 8 38.0b [45]
Odobenus rosamarus (Pacific walrus) AK, USA 53 3 5.6 [42]
Zalophus californianus (Californian sea lion) CA, USA 27 8 29.6 [42]
Phoca vitulina (Harbor seal) AK, USA 311 51 16.4 [42]
Canada 34 3 8.8 [50]
WA, USA 380 29 7.6 [51]
Cystophora cristata (Hooded seal) Canada 60 1 1.7 [50]
Phoca hispida (Ringed seal) AK, USA 32 5 15.6 [42]
Canada 34 3 8.8 [50]
Halichoerus grypus (Grey seal) Canada 122 11 9.0 [50]
Erignathus barbatus (Bearded seal) AK, USA 8 4 50.0 [42]
Phoca groenlandica (Harp seal) Canada 112 0 0.0 [50]
Phoca largha (Spotted seal) AK, USA 9 1 11.1 [42]
Phoca fasciata (Ribbon seal) AK, USA 14 0 0 [42]
Tursiops truncatus (Bottlenose dolphin) Spain 7 4 57.1 [52]
CA, USA 94 91 96.8 [42]
FL, USA 47 47 100.0 [42]
Stenella coeruleoalba (Striped dolphin) Spain 36 4 11.1 [52]
Delphinus delphis (Common dolphin) Spain 4 2 50.0 [52]
aAbbreviation: MAT, modified agglutination test.bIndirect fluorescent antibody test.
Review TRENDS in Parasitology Vol.20 No.11 November 2004534
208C for 12 weeks and up to 35 ppt at 188C for 40 days [12].Because T. gondii oocysts must become sporulated to beinfectious, their ability to sporulate and survive in seawater was tested. Of unsporulated oocysts placed in 15and 32 ppt artificial sea water, 75–80% sporulated withinthree days and were infectious to mice. Sporulated oocystsstored in 15 ppt sea water at 48C or room temperature forsix months also infected mice [32].
Protozoan parasites in contaminated shellfish
Giardia duodenalis genotype A was identified by molecu-lar tests in Macoma clams in the Rhode River [33]. Thesetiny clams have no economic value but serve as biologicalindicators of water contamination.
Cryptosporidium oocysts placed in sea water weredetected in experimentally exposed oysters [34,35], mus-sels [36] and clams [35]. Oocysts retained for as long as 30days without initiating infection in shellfish were infectivewhen recovered from artificially and naturally contami-nated shellfish [37]. Cryptosporidium has been detected innaturally contaminated mussels, oysters, clams andcockles worldwide (Table 1). The highest prevalence ofcontamination was found in seven Chesapeake Baylocations after a major rainfall event [1]. However,even during a period of low rainfall, Cryptosporidiumwas detected in oysters in 11 out of 13 Atlantic coaststates [38].
Mice became infected when fed tissues from oysters sixdays after exposure of oysters to T. gondii oocysts in a seawater aquarium [39]. Toxoplasma gondii-specific smallsubunit ribosomal RNA (ssrRNA) could be detected for upto 21 days post-exposure in artificially exposed mussels
www.sciencedirect.com
that were positive by bioassay for only three days post-exposure [40]. Toxoplasma gondii oocysts have not beendetected in shellfish in nature.
Protozoan parasites in marine mammals
Giardia cysts have been detected in feces from a Californiasea lion, from ringed seals in arctic Canada, and fromharp, grey and harbor seals in the Gulf of St Lawrence,Canada (Table 3). Cryptosporidium has been detected inthe dugong, in Australia, in California sea lions and inseals in arctic Canada (Table 3). Toxoplasma gondii hasbeen isolated from a Pacific harbor seal [41]. It has alsobeen histologically identified in seals, dolphins, sea lions, amanate and a Beluga whale from the USA, Italy, Spain,Canada and Australia, without confirmation by isolationof organisms or serology [42]. The finding of T. gondiiantibodies in sera from marine mammals suggests world-wide contamination of the marine environment (Table 2).
Toxoplasmosis in sea otters is a concern [42–47]. Themain sea otter colonies are in California, Washington andAlaska. Mortalities in California have been studied themost [47], where T. gondii has been isolated from brains orhearts of 22.3% of 67 [43] and 32% of 75 [44] California seaotters. Initially, all isolates were identified as genotype II[43], but a recent study [46] found two genotypes in seaotter beachcast carcasses: 40% were Type II and 60% werea unique genotype called Type x. Comparison betweenisolates from sea otters and other marine mammals withisolates from terrestrial animals will hopefully clarifyroutes and mechanisms of land-to-sea transmission.Serological surveys further indicate exposure of manysea otters to T. gondii (Table 2). Although antibodies to

Table 3. Giardia and Cryptosporidium detected in marine mammals
Parasite Host (common name) Location No.
infected
Detection method Ref.
Giardia Phoca hispida (Ringed seal) Arctic Canada 3 Microscopy [53]
Giardia Phoca groenlandica (Harp seal) Gulf of St Lawrence 15 Microscopy [54]
Halichoerus grypus (Grey seal) 4
Phoca vitulina (Harbor seal) 1
Giardia P. hispida Ungava Bay, Canada 43 Flow cytometrya b
Giardia Zalophus californianus (California sea lion) Humboldt Bay, USA 1 Microscopy [55]
Cryptosporidium P. hispida Ungava Bay, Canada 2 Flow cytometrya b
Cryptosporidium Z. californianus Humboldt Bay, USA 1 Microscopy [55]
Cryptosporidium Dugong dugon (Dugong) Queensland, Australia 1 Histology [56]
aFluorescence labeling followed by flow cytometry.bB. Dixon et al., unpublished.
Review TRENDS in Parasitology Vol.20 No.11 November 2004 535
T. gondii were not found in Alaskan sea otters, antibodieswere found in other Alaskan marine mammals.
Grey seals that were fed T. gondii oocysts developedinfection [48], but how marine mammals become infectedin nature has not been well studied. Although mostmarine mammals do not prey on warm-blooded animals,which are potential sources of tissue cysts of T. gondii,congenital transmission has been reported in dolphins.Fresh water runoff into coastal waters poses risk for seaotters [45]. Ingesting oocysts from sea water or eatinganimals that have ingested oocysts are likely sources.Although sea otters eat shellfish, their role as paratenichosts for T. gondii is suspected but unproven. Dolphinsbecome infected (Table 2), but they drink little or no waterand derive their nutrition from fish, squid and other cold-blooded sea animals that do not develop tissue cysts ofT. gondii.
Conclusions
There are many indicators of widespread fecal contami-nation from land to sea, including: (i) recorded outbreaksof giardiasis, cryptosporidiosis and toxoplasmosis associ-ated with drinking water; (ii) findings of Giardia andCryptosporidium in surface waters used to irrigate freshvegetables; (iii) detection of Giardia and Cryptosporidiumin marine mammals and shellfish in fresh, estuarine andcoastal waters worldwide, and in marine waters used forrecreation; and (iv) the global presence of antibodies toT. gondii in many species of marine mammals. Someinfections might be indirect, through vertical trans-mission or transport hosts, and wild terrestrial animalsmight be sources of infectious organisms; however, manyorganisms originate from humans, their companionanimals and domesticated animals. Most shellfish con-taminated with Cryptosporidium have been found tocontain oocysts of C. parvum, the zoonotic speciesprevalent in young domesticated ruminants and inhumans. Cryptosporidium hominis, the only speciesidentified in a marine mammal (a dugong), is the mostprevalent species in human infections. Infection of marinemammals with Giardia has not been identified bymolecular methods and molecular data for T. gondii hasbeen reported only for sea otters. Hopefully, moremolecular data will become available from parasitesisolated from marine mammals and will be comparedwith data from terrestrial animals. Such data will clarifyour understanding of the complex relationships among
www.sciencedirect.com
hosts, and will help to identify the routes and mechanismsof land to sea transmission. Nevertheless, practices ofwastewater disposal, drinking water purification, runoffcontrol and farm manure management must improve.Otherwise, as human and animal populations grow, so willthe negative impact of fecal contamination on publichealth and ultimately on marine life.
References
1 Fayer, R. et al. (2002) Temporal variability of Cryptosporidium in theChesapeake Bay. Parasitol. Res. 88, 998–1003
2 Thompson, R.C.A. (2000) Giardiasis as a re-emerging infectiousdisease and its zoonotic potential. Int. J. Parasitol. 30, 1259–1267
3 Sulaiman, I. et al. (2003) Triosephosphate isomerase gene character-ization and potential zoonotic transmission of Giardia duodenalis.Emerg. Infect. Dis. 9, 1444–1452
4 Xiao, L. et al. (2004) Cryptosporidium taxonomy: recent advances andimplications for public health. Clin. Microbiol. Rev. 17, 72–97
5 Duncanson, P. et al. (2001) High levels of congenital transmission ofToxoplasma gondii in a commercial sheep flock. Int. J. Parasitol. 31,1699–1703
6 Grigg, M.E. et al. (2001) Success and virulence in Toxoplasma as theresult of sexual recombination between two distinct ancestries.Science 294, 161–165
7 Su, C. et al. (2003) Recent expansion of Toxoplasma through enhancedoral transmission. Science 299, 414–416
8 Volkman, S.K. and Hartl, D.L. (2003) A game of cat and mouth.Science 299, 353–354
9 Smith, H.V. and Grimason, A.M. (2003) Giardia and Cryptosporidium.In TheHandbook of Water andWastewaterMicrobiology (Mara, D. andHoran, N.J. eds), pp. 696–755, Academic Press
10 Marshall, M.M. et al. (1997) Waterborne protozoan pathogens. Clin.Microbiol. Rev. 10, 67–85
11 Lane, S. and Lloyd, D. (2002) Current trends in research into thewaterborne parasite Giardia. Crit. Rev. Microbiol. 28, 123–147
12 Fayer, R. Cryptosporidium: a waterborne zoonotic parasite. Vet.Parasitol. (in press)
13 Casemore, D.P. et al. (1997) Cryptosporidiosis in human and animalepidemiology. In Cryptosporidium and Cryptosporidiosis (Fayer, R.ed.), pp. 66–92, CRC Press
14 Benenson, M.W. et al. (1982) Oocyst transmitted toxoplasmosisassociated with ingestion of contaminated water. N. Engl. J. Med.307, 666–669
15 Bowie, W.R. et al. (1997) Outbreak of toxoplasmosis associated withmunicipal drinking.water. The BC Toxoplasma Investigation Team.Lancet 350, 173–177
16 Dubey, J.P. and Beattie, C.P. (1988) Toxoplasmosis of Animals andMan, CRC Press
17 Tenter, A.M. et al. (2000) Toxoplasma gondii: from animals to humans.Int. J. Parasitol. 30, 1217–1258
18 Jones, J.L. et al. (2001) Toxoplasma gondii infection in the UnitedStates: seroprevalence and risk factors. Am. J. Epidemiol. 154,357–365
19 Wright, R.J. et al. (1998) Agricultural uses of municipal, animal, and

Review TRENDS in Parasitology Vol.20 No.11 November 2004536
industrial byproducts. USDA, Agricultural Research Service, Con-servation Research Report No. 44, US Government Printing Office,Washington, DC
20 Trout, J.M. et al. Prevalence of Giardia genotypes in pre-weaned dairycalves. Vet. Parasitol. (in press)
21 Dupont, H.L. et al. (1995) The infectivity of Cryptosporidium parvumin healthy volunteers. N. Engl. J. Med. 332, 855–859
22 Santin, M. et al. (2004) Prevalence and age-related variation ofCryptosporidium species and genotypes in dairy calves. Vet. Parasitol.122, 103–117
23 Dubey, J.P. et al. (1996) Infectivity of low numbers of Toxoplasmagondii oocysts to pigs. J. Parasitol. 82, 438–443
24 Thurston-Enriquez, J.A. et al. (2002) Detection of protozoan parasitesand microsporidia in irrigation waters used for crop production.J. Food Prot. 65, 378–382
25 Monge, R. and Chinchilla, M. (1996) Presence of Cryptosporidiumoocysts in fresh vegetables. J. Food Prot. 59, 202–203
26 Ortega, Y.R. et al. (1997) Isolation of Cryptosporidium parvum andCyclospora cayetanensis from vegetables collected in markets of anendemic region in Peru. Am. J. Trop. Med. Hyg. 57, 683–686
27 McDonald, J.C. et al. (1990) An outbreak of toxoplasmosis in pregnantwomen in northern Quebec. J. Infect. Dis. 161, 769–774
28 Johnson, D.C. et al. (1995) Detection of Giardia and Cryptosporidiumin marine waters. Water Sci. Technol. 31, 439–442
29 Correa, I.E. et al. (1990) Removal of Giardia spp. by conventionalwater and sewage treatment processes and presence of cysts in surfacewaters. In Tropical Hydrology and Caribbean Water Resources(Krishna, J.H. et al., eds), pp. 535–541, American Water ResourcesAssociation Technical Publication Series
30 Ferguson, C.M. et al. (1996) Relationships between indicators,pathogens and water quality in an estuarine system. Water Res. 30,2045–2054
31 Ho, B.S.W. and Tam, T-Y. (1998) Occurrences of Giardia cysts in beachwater. Water Sci. Technol. 38, 73–76
32 Lindsay, D.S. et al. (2003) Sporulation and survival of Toxoplasmagondii oocysts in sea water. J. Eukaryot. Microbiol. 50, S687–S688
33 Graczyk, T.K. et al. (1999) Giardia duodenalis cysts of genotype Arecovered from clams in the Chesapeake Bay subestuary, Rhode River.Am. J. Trop. Med. Hyg. 61, 526–529
34 Fayer, R. (1998) Survival of infectious Cryptosporidium parvumoocysts in seawater and eastern oysters (Crassostrea virginica) in theChesapeake Bay. Appl. Environ. Microbiol. 64, 1070–1074
35 Freire-Santos, F. et al. (2002) Survival of Cryptosporidium parvumoocysts recovered from experimentally contaminated oysters (Ostreaedulis) and clams (Tapes decussates). Parasitol. Res. 88, 130–133
36 Tamburrini, A. and Pozio, E. (1999) Long term survival of Cryptos-poridium oocysts in seawater and in experimentally infected mussels(Mytilus galloprovincialis). Int. J. Parasitol. 29, 711–715
37 Fayer, R. et al. (1997) Potential role of the eastern oyster, Cassostreavirginica, in the epidemiology of Cryptosporidium parvum. Appl.Environ. Microbiol. 63, 2086–2088
38 Fayer, R. et al. (2003) Contamination of Atlantic coast commercialshellfish with Cryptosporidium. Parasitol. Res. 89, 141–145
39 Lindsay, D.S. et al. (2001) Removal of Toxoplasma gondii oocysts fromsea water by eastern oysters (Crassostrea virginica). J. Eukaryot.Microbiol. 48, S197–S198
40 Arkush, K.D. et al. (2003) Molecular and bioassay-based detection ofToxoplasma gondii oocyst uptake by mussels (Mytilus galloprovincia-lis). Int. J. Parasitol. 33, 1087–1097
41 Miller, M.A. et al. (2001) Isolation and characterization of twoparasitic protozoa from a pacific harbor seal (Phoca Vitulinarichardsi) with meningoencephalomyelitis. J. Parasitol. 87, 816–822
www.sciencedirect.com
42 Dubey, J.P. et al. (2003) Toxoplasma gondii, Neospora caninum,Sarcocystis neurona, and Sarcocystis canis-like infections in marinemammals. Vet. Parasitol. 116, 275–296
43 Cole, R.A. et al. (2000) Biological and molecular characterizations ofToxoplasma gondii strains obtained from southern sea otters(Enhydra lutris nereis). J. Parasitol. 86, 526–530
44 Miller, M.A. et al. (2002) Evaluation of an indirect fluorescentantibody test (IFAT) for demonstration of antibodies to Toxoplasmagondii in the sea otter (Enhydra lutris). J. Parasitol. 88, 594–599
45 Miller, M.A. et al. (2002) Coastal freshwater runoff is a risk factor forToxoplasma gondii infection of southern sea otters (Enhydra lutrisnereis). Int. J. Parasitol. 32, 997–1006
46 Miller, M.A. et al. (2004) An unusual genotype of Toxoplasma gondii iscommon in California sea otters (Enhydra lutris nereis) and is a causeof mortality. Int. J. Parasitol. 34, 275–284
47 Kreuder, C. et al. (2003) Patterns of mortality in southern sea otters(Enhydra lutris nesris) from 1998–2001. J. Wildl. Dis. 39, 495–509
48 Gajadhar, A.A. et al. (2004) Experimental Toxoplasma gondii infectionin grey seals (Halichoerus grypus). J. Parasitol. 90, 255–259
49 Hanni, K.D. et al. (2003) Clinical pathology and assessment ofpathogen exposure in southern and Alaskan sea otters. J. Wildl.Dis. 39, 837–850
50 Measures, L.N. et al. (2004) Seroprevalence of Toxoplasma gondii inCanadian pinnipeds – an example of pathogen pollution. J. Wildl. Dis.40, 294–300
51 Lambourn, D.M. et al. (2001) Seroprevalence of Toxoplasma gondii inharbor seals (Phoca vitulina) in southern Puget Sound, Washington.J. Parasitol. 87, 1196–1197
52 Cabezon, O. et al. (2004) Seroprevalence of Toxoplasma gondiiantibodies in wild dolphins from the Spanish Mediterranean coast.J. Parasitol. 90, 643–644
53 Olson, M.E. et al. (1997) Giardiasis in ringed seals from the westernarctic. J. Wildl. Dis. 33, 646–648
54 Measures, L.N. and Olson, M.E. (1999) Giardiasis in pinnipeds fromeastern Canada. J. Wildl. Dis. 35, 779–782
55 Deng, M.Q. et al. (2000) First findings of Cryptosporidium andGiardia in California sea lions (Zalophus californianus). J. Parasitol.86, 490–494
56 Hill, B.D. et al. (1997) Cryptosporidium infection in a dugong (Dugongdugon). Aust Vet. J. 75, 670–671
57 Chalmers, R.M. et al. (1997) Cryptosporidium parvum in environ-mental samples in the Sligo area, Republic of Ireland: a preliminaryreport. Lett. Appl. Microbiol. 25, 380–384
58 Graczyk, T.K. et al. (1999) Cryptosporidium oocysts in Bentmussels (Ischadium recurvum) in the Chesapeake Bay. Parasitol.Res. 85, 518–521
59 Fayer, R. et al. (1999) Cryptosporidium parvum in oysters fromcommercial harvesting sites in the Chesapeake Bay. Emerg. Infect.Dis. 5, 706–710
60 Freire-Santos, F. et al. (2000) Detection of Cryptosporidium oocysts inbivalve molluscs destined for human consumption. J. Parasitol. 86,853–854
61 Gomez-Bautista, M. et al. (2000) Detection of infectious Cryptos-poridium parvum oocysts in mussels (Mytilus galloprovincialis)and cockles (Cerastoderma edule). Appl. Environ. Microbiol. 66,1866–1870
62 Graczyk, T.K. et al. (2001) Cryptosporidium parvum oocysts in zebramussels (Dreissena polymorpha): evidence from the St. LawrenceRiver. Parasitol. Res. 87, 231–234
63 Alonso, I.L. et al. (2003) Evaluation of diagnosis techniques for thedetection of Cryptosporidium oocysts and Giardia cysts in bivalves forhuman consumption. Acta Parasitol. Port. 10, 33