yersinia enterocolitica: prevalenza, isolamento e ricerca ... · 2.1 epidemiologia di yersinia ......
TRANSCRIPT

Università degli Studi di Pisa
Facoltà di Scienze Matematiche Fisiche e Naturali
Laurea Magistrale in Biologia Applicata alla Biomedicina
Curriculum Fisiopatologico
Yersinia enterocolitica: prevalenza,isolamento e ricerca geni di patogenicità da
matrici alimentari prelevate in Toscana
1
Sede di Tirocinio: Istituto Zooprofilattico Sperimentale delle Regioni Lazio e Toscana, sezione
di Pisa
Candidata: Irene Barsanti
Relatore interno: Prof.ssa Annalaura CarducciRelatore esterno: Dott. Matteo Senese
Anno accademico 2015/2016

Indice Generale1 Introduzione.......................................................................................................................................32 Excursus storico di Yersinia enterocolitica........................................................................................8
2.1 Epidemiologia di Yersinia enterocolitica.................................................................................102.2 Caratteristiche biochimiche, metaboliche, colturali e condizioni ottimali di crescita.............162.3 Geni di patogenicità e patogenesi della yersiniosi...................................................................202.4 Alimenti di origine animale.....................................................................................................37
2.4.1 Latte e prodotti lattiero-caseari........................................................................................372.4.2 Pastorizzazione, Termizzazione e Tecnologia ESL..........................................................442.4.3 I patogeni del latte............................................................................................................482.4.4 Fermentazione prodotti lattiero-caseari e batteri lattici...................................................512.4.5 Presenza di Yersinia spp nel latte e prodotti a base di latte..............................................55
2.5 Prodotti a base di carne............................................................................................................582.5.1 Yersinia spp nei prodotti a base di carne..........................................................................63
2.6 Scopo della Tesi.......................................................................................................................673 Materiali e metodi............................................................................................................................69
3.1 Procedura di analisi qualitativa................................................................................................713.1.1 Arricchimento in terreni liquidi selettivi..........................................................................713.1.2 Semina in piastra ed identificazione................................................................................713.1.3 Conferma..........................................................................................................................73
3.2 Procedura analisi quantitativa: il metodo MPN.......................................................................783.3 Protocollo operativo per la ricerca dei geni AIL, YST A e YST B nei ceppi di campo di Yersinia enterocolitica (Thoener P., et al, 2002- ISS rapporti ISTISAN 97/31)............................83
4 Risultati............................................................................................................................................904.1 Risultati dell'analisi molecolare...............................................................................................924.2 Discussione dei risultati.........................................................................................................100
5 Conclusioni....................................................................................................................................115Bibliografia.......................................................................................................................................117
2

1 Introduzione
Nei paesi industrializzati il 30% della popolazione viene colpita ogni anno da malattie trasmesse
dagli alimenti. Negli USA l’incidenza è stimata a 76 milioni di casi per anno, cui conseguono più di
325.000 ospedalizzazioni e oltre 5.000 decessi. In Italia le stime sono di circa 300.000 casi/anno,
ma le statistiche, a causa di mancate segnalazioni e/o diagnosi inesatte, sottostimano la reale
incidenza di queste patologie (dati WHO, 2008). La gestione della sicurezza è attuata attraverso
l’adozione di misure di controllo preventive finalizzate a minimizzare, per quanto possibile, la
contaminazione e lo sviluppo dei microrganismi patogeni in ogni fase della catena alimentare. Non
si possono, infatti, ottenere alimenti sicuri se non si mantengono sotto controllo tutte le fasi del
processo produttivo: allevamento, trasporto, lavorazione, distribuzione, vendita e non da ultimo, la
preparazione per il consumo.
Nel corso degli ultimi anni le metodologie di gestione della sicurezza alimentare consolidate, si
fondano sull’applicazione integrata e sistematica dei piani di HACCP e delle buone pratiche di
fabbricazione (GMP) e delle procedure di corretta prassi igienica (GHP).
L’applicazione di metodologie a carattere preventivo fondate sul controllo di processo, risulta
efficace se attuata in ciascuna fase della filiera. Gli Operatori del Settore Alimentare nel progettare
ed applicare il proprio sistema di autocontrollo, devono considerare nella misura appropriata le
interazioni che si determinano fra i differenti fattori della produzione (materie prime, igiene degli
ambienti di produzione/lavorazione, operatori, etc.) nelle differenti fasi del processo.
Le malattie di origine alimentare rappresentano un importante e crescente problema di salute
pubblica ed economica in molti paesi.
Le radicali trasformazioni sociali ed economiche hanno cambiato in modo assai rilevante
3

l'approvvigionamento alimentare e hanno al contempo favorito nuovi indirizzi produttivi,
commerciali e consumistici.
Tali innovazioni, se da una parte hanno conseguito risultati positivi, dall'altra hanno finito per
influenzare, in senso negativo, gli attributi qualitativi e la genuinità delle materie prime alimentari
causando pericoli potenziali per la salute umana.
I Servizi sanitari, i relativi organismi nazionali e sopra nazionali e le stesse istituzioni scientifiche,
hanno dovuto rivolgere una sempre maggiore attenzione ai gravi problemi connessi alla
contaminazione ambientale da agenti chimico-fisici ed ai non sempre controllabili meccanismi del
loro passaggio dall'ambiente agli animali e da questi, con gli alimenti prodotti (carne, latte, uova,
ecc), all'uomo, situato all'apice di molte catene trofiche ( Del Bono-Renon 2001).
Le zoonosi sono infezioni e malattie trasmesse direttamente tramite il contatto con animali infetti o
indirettamente, attraverso il consumo di alimenti contaminati.
Al fine di diminuire e/o evitare la trasmissione di queste malattie, è importante l'identificazione
degli animali e dei prodotti derivati, che rappresentano le principali fonti di infezione ( Del Bono,
Renon 2001).
In Italia, le malattie a trasmissione alimentare, sono segnalate dal sistema di notifica obbligatorio
delle malattie infettive, coordinato a livello nazionale dal Ministro della Salute. I dati relativi alle
notifiche obbligatorie, sono di solito fortemente distorti dalla sottonotifica, e la trasmissione delle
informazioni è spesso poco tempestiva. Per questa ragione sono stati messi a punto sistemi di
sorveglianza, basati sulle attività dei laboratori di microbiologia clinica, che permettono la raccolta
dei dati con tempestività e accuratezza maggiori di quelle del sistema sanitario.
Nella nostra nazione, la sorveglianza di laboratorio di alcune infezioni a prevalente trasmissione
alimentare fa capo al sistema ENTER-NET, inserito in un network cui partecipano numerosi paesi
europei e extraeuropei. In Italia, tale sistema è costituito da una rete di laboratori microbiologici
4

coordinati da centri di riferimento regionali rappresentati da laboratori ospedalieri, da Istituti
Zooprofilattici Sperimentali (IZS) e da Agenzie Regionali per la Protezione Ambientale (ARPA)
(Cantoni C. 2011).
L'Autorità Europea sulla Sicurezza Alimentare (EFSA) ha stabilito che tutti gli stati membri sono
tenuti a compiere delle indagini periodiche riguardo ad alcuni microrganismi patogeni per l'uomo,
inclusa Yersinia spp, presenti nelle derrate alimentari (EFSA, 2009).
L'attenzione verso il genere Yersinia nasce dal fatto che, negli ultimi decenni, si è registrato un
incremento nell'incidenza delle yersiniosi in diversi paesi europei, collocandola al terzo posto tra le
zoonosi più frequentemente notificate (EFSA, 2009).
Un totale di 7.017 casi confermati di yersiniosi sono stati segnalati nell' UE nel 2011,
corrispondente ad un incremento del 3,5% rispetto all'anno precedente ( Quaderni di zooprofilassi
N° 12 agosto 2014 ).
Per quanto riguarda la realtà italiana, sebbene le tossinfezioni alimentari rientrino nel sistema di
notifica obbligatoria delle malattie infettive, purtroppo, questo sistema non viene applicato nel caso
della yersiniosi, malattia causata dai ceppi patogeni di Yersinia enterocolitica. Quindi, non sono
disponibili dei reali tassi di incidenza nel nostro paese e, poiché la maggior parte di casi di
yersiniosi si verifica sporadicamente senza un'origine apparente, l'epidemiologia dell'infezione da
Yersinia enterocolitica è complessa e non del tutto compresa (Fredriksson-Ahomaa et. al 2003).
L'emergenza della yersiniosi è probabilmente legata ai cambiamenti apportati negli allevamenti di
bestiame, nelle tecnologie alimentari e nell'industria del cibo. Di grande importanza sono i
cambiamenti nell'industria della carne, dove la produzione si è spostata dai piccoli macelli,
interessando pochi individui, ai grandi impianti che macellano migliaia di animali al giorno,
fornendo i loro prodotti alla nazione e a livello internazionale. Le fattorie si sono ingrandite e le
tecniche di zootecnia sono diventate molto più intensive. Mentre molte tecniche moderne di
5

macellazione riducono il rischio di contaminazione della carne, esistono opportunità di trasmissione
da animale a animale del microrganismo e di cross- contaminazione delle carcasse e dei prodotti a
base di carne che non erano note pochi decenni fa. Inoltre, i progressi nel confezionamento e
refrigerazione consentono ora all'industria e ai consumatori di conservare gli alimenti per periodi
molto più lunghi, questo è un fattore significativo nei confronti di un agente patogeno adattato al
freddo come Yersinia enterocolitica (Bari Md. L. et al 2011).
Yersinia enterocolitica è un patogeno estremamente versatile, in grado di adattarsi a diverse
condizioni ambientali e di contrastare le difese dell'ospite tramite diversi fattori di patogenicità
codificati da geni a localizzazione cromosomica e plasmidica ( Cattabiani, 2002).
Le Yersiniosi, un tempo ritenute tipiche malattie infettive, sono state considerate anche come
malattie alimentari, ricondotte alle modificate abitudini conservative ( refrigerazione domestica) ed
approvvigionali ( ristorazione collettiva ).
E' quindi necessario, da una parte non sottovalutare il potenziale pericolo di contaminazioni
microbiche che comporterebbero conseguenze negative sia per la tutela del consumatore, che per
gli allevatori, dall'altra mantenere un puntuale monitoraggio dell'epidemiologia dei patogeni
potenzialmente presenti negli allevamenti, al fine di mettere in atto tutte le misure precauzionali del
caso senza creare allarmismo ingiustificato.
Yersinia enterocolitica appartiene alla famiglia alle Enterobacteriacee, è un germe che ha modeste
esigenze nutritive e buona adattabilità a svilupparsi in substrati alimentari refrigerati. Il suo
sviluppo massimo avviene a pH neutro, pur potendo proliferare in ambienti acidi e alcalini.
Possiede un temperatura ottimale di sviluppo di 29 ° C, ma ha anche la prerogativa di moltiplicarsi
in un intervallo di temperatura tra 0°C a 45°C; anche una moderata salinità ( fino al 5% di NaCl)
non ne inibisce la duplicazione (Andreis, Ottaviani 2002).
La capacità di replicarsi a temperature di refrigerazione è un aspetto significativo nell'igiene degli
6

alimenti (Bari Md. L. et al. 2011).
Alla specie Yersinia enterocolitica appartengono microrganismi distribuiti nell'ecosistema acquatico
e terrestre, la maggior parte dei quali sono considerati apatogeni e ubiquitari. Solo alcuni stipiti,
provvisti di meccanismi di virulenza, sono patogeni per l'uomo, che generalmente contrae
l'infezione mediante l'ingestione di acqua e alimenti contaminati ( Bottone E.J. 1999).
Secondo recenti studi, la patogenicità di numerosi sierotipi di Yersinia enterocolitica sarebbe in
parte legata al potere invasivo del germe sulle cellule epiteliali delle mucose intestinali, ed in parte
da ricondursi ad un principio tossico, enterotossina, assai resistente alle alte temperature, tanto da
venire denaturata solo da temperature superiori ad un parametro di 120°C per 30 minuti.( Del
Bono, Renon 2001).
La via di trasmissione più comune è quella oro-fecale tramite cibo contaminato. Il contatto diretto
persona-persona è raro. Occasionalmente Yersinia enterocolitica patogena è stata isolata in vegetali
e acqua quindi, sono anch'esse potenziali fonti di infezione per l'uomo. Tuttavia, il tasso di
isolamento di Yersinia enterocolitica patogena è basso, questo è dovuto anche alla limitata
sensibilità dei metodi di rilevamento (Bari Md. L. et al 2011).
7

2 Excursus storico di Yersinia enterocolitica
Yersinia enterocolitica fu descritta per la prima volta nel 1934 negli Stati Uniti da McIver e Pike
(McIver and Pike 1934).
Alcuni anni dopo, Schleiftein e Coleman (1939) posero l'attenzione su batteri, isolati da contenuto
intestinale di pazienti, aventi caratteristiche simili a quelle descritte da McIver e Pike. Per questi
microrganismi fu proposto il nome di Bacterium enterocoliticum (Schleifstein and Coleman 1939).
Solo nel 1944, Van Loghem, introdusse il genere Yersinia ( in onore a A.J. Yersini, il primo a
descrivere il bacillo della peste nel 1894), consentendo la differenziazione dell'attuale Yersinia
enterocolitica dagli isolati Yersinia enterocolitica-like.
Fu Frederiksen nel 1964 a dimostrare, mediante indagini biochimiche comparative, che tutti questi
stipiti batterici di incerta classificazione, possedevano caratteristiche comuni sovrapponibili a quelle
di Bacterium enterocoliticum: venne quindi proposta la nuova specie microbica Yersinia
enterocolitica ed inserita nella famiglia del Enterobacteriaceae (Frederiksen, W. 1964).
Dalla prima descrizione del genere Yersinia, avvenuta nel 1944 da VanLoghem, la situazione
tassonomica si è evoluta e modificata in continuazione in conseguenza delle diversità emerse a
seguito di indagini effettuate, sia con tecniche classiche, che di biologia molecolare.
Accanto alle 3 specie “storiche “ - Yersinia pestis, Yersinia pseudotubercolosis, Yersinia
enterocolitica- sono oggi descritte altre 8 specie che evidenziano profili biochimici- enzimatici tali
da discostarsi dalla specie enterocolitica e proporsi, prima come ceppi “enterocolitica-like”, poi
come specie a se stanti: Yersinia frederiksenii, Yersinia intermedia, Yersinia kristensenii, Yersinia
mollaretii, Yersinia bercovieri, Yersinia aldovae, Yersinia rhodei, Yersinia ruckeri (Cattabiani F.
2002).
8

Attualmente al genere Yersinia appartengono undici specie: tre specie sono patogene per l'uomo,
mentre otto specie sono apatogene, possono causare infezioni opportunistiche. Le specie apatogene
sono ubiquitarie e di scarso significato clinico e comprendono: Yersinia intermedia, Yersinia
frederiksenii, Yersinia kristensenii, Yersinia rhodei, Yersinia aldoviae, Yersinia mollaretii, Yersinia
bercovieri, Yersinia ruckeri. Le specie patogene sono Yersinia pestis, Yersinia pseudotubercolosis e
Yersinia enterocolitica le quali, pur presentando caratteristiche epidemiologiche e modalità di
trasmissione differenti, condividono importanti determinati di virulenza capaci di eludere le difese
immunitarie aspecifiche dei loro ospiti.
Tra i microrganismi in grado di provocare gastroenteriti, Yersinia enterocolitica va assumendo un
interesse crescente. L'incremento dei casi è anche correlato allo sviluppo delle conoscenze su tale
germe ed al maggior numero di accertamenti condotti per la sua identificazione.
Yersinia enterocolitica è ampiamente distribuita in natura, è stata rilevata nella carne di suino e nei
prodotti derivati, ma anche in altri animali, nel latte, nelle verdure e nel pesce ( Del Bono, Renon
2001).
9
Fig. 1: Yersinia enterocolitica vista al microscopio.

2.1 Epidemiologia di Yersinia enterocolitica
Yersinia enterocolitica viene isolata frequentemente dal contenuto intestinale di suino, che sembra
rappresentare il “serbatoio animale” asintomatico del microrganismo.
Le fonti di infezione per l'uomo sono, principalmente, i prodotti di derivazione suina
(Fredriksson-Ahomaa, et.al.2006; http://www.antropozoonosi.it/Malattie/Yersiniosi/Yersiniosi.php).
Questo microrganismo è stato isolato da torte, carni confezionate sottovuoto, prodotti ittici, ortaggi,
latte e altri prodotti alimentari, oltre che da carne di manzo, agnello e maiale. Sembra che la carne
di maiale sia la principale fonte di ceppi patogeni per l'uomo. Più specificamente si è osservato che
le tonsille di maiale rappresentano la principale fonte di contaminazione di fegato, cuore e reni
(Cantoni C. 2011).
Il legame tra l'animale serbatoio e l'infezione umana è principalmente dovuto all'ingestione di carne
suina cruda o poco cotta (Bottone E.j. 1997).
Yersinia enterocolitica appartenente al biosierotipo 4/O: 3 è stata occasionalmente isolata da cani e
gatti (Yanagawa et al. 1978). Il ritrovamento del batterio in questi animali con diarrea, suggerisce
che il microbo giochi un ruolo nell'infezione (Ahvonen et al. 1973). Yersinia enterocolitica 4/O: 3 è
stata anche isolata nelle feci di cani sani (Yanagawa et al 1978). I cani hanno dimostrato di espellere
il microrganismo con le feci per diverse settimane dopo l'infezione (Ahvonen et al 1973 ; Fenwick
et al 1994 ). Cani e gatti sono una probabile causa di infezione per l'uomo, questo è dovuto al loro
stretto contatto con le persone, specialmente con i bambini. Tuttavia, la trasmissione diretta tra
animali domestici e uomo deve essere ancora dimostrata (Robins-Browne 1997). E' stato dimostrato
però, che l'incidenza di yersiniosi è molto elevata tra i bambini al di sotto dei 6 anni di età
(Kontiainen et al.1994).
E' stato provato che la carne cruda o poco cotta di maiale è una fonte di infezione per Yersinia
10

enterocolitica 4/O: 3 anche per cani e gatti, il batterio può causare enteriti in animali domestici, i
quali possono eliminare all'esterno il patogeno senza manifestare sintomi evidenti per diverse
settimane dopo l'infezione (Fenwick et al. 1994).
Gli animali domestici possono veicolare la trasmissione dell'infezione tra maiali e bambini
(Fredriksson-Ahomaa et.al 2001). Circa il 60-70% dei casi di yersiniosi si verificano nei bambini,
spesso i casi umani si sono evidenziati in concomitanza con focolai di malattia negli animali
domestici “conviventi”, quali cuccioli o gattini (www.antropozoonosi.it).
Dato che nel suino gli stipiti patogeni di Yersinia enterocolitica vengono isolati prevalentemente a
livello tonsillare (Fredriksson-Ahomaa 2003) e solo in misura minore nel contenuto intestinale, le
tonsille rappresentano la principale fonte di contaminazione per le carcasse (Thibodeau V. et al.
1999).
Diversi animali di affezione e domestici, come cani, gatti, bovini, ovini, cavalli e diversi animali
selvatici, come roditori, soprattutto topi, scimmie, cervi, agnelli, pesci, ostriche e volpi sono stati
incriminati come possibili serbatoi di infezione (Cantoni C. 2011).
Gli alimenti ritenuti responsabili più di frequente sono il latte in polvere e derivati lattei e più in
generale, alimenti di origine vegetale ed animale refrigerati o non sufficientemente cotti (Del Bono,
Renon 2011).
Infatti, in Europa i maiali sono portatori sani dei ceppi patogeni di Yersinia enterocolitica,
soprattutto di quelli appartenenti al biotipo 4 (sierotipo O: 3) e al biotipo 2 (sierotipo O: 9 e O: 5,27)
anche se con una frequenza minore. Il batterio è presente nella cavità orale, specialmente nelle
tonsille, nei linfonodi e nell'intestino e nel contenuto intestinale. I ceppi di Yersinia enterocolitica
appartenenti al biotipo 4 (sierotipo O: 3) sono stati frequentemente isolati sulla superficie delle
carcasse macellate come risultato della diffusione del batterio tramite feci, attraverso il contenuto
intestinale e le tonsille durante le operazioni di macellazione. La carne cruda di maiale potrebbe
11

quindi essere una fonte importante di Yersinia enterocolitica appartenente al biosierotipo 4 (O: 3).
In Norvegia, i focolai di infezione da Yersinia enterocolitica in greggi di capre sono stati causati da
batteri facenti parte del biotipo 5 (sierotipo O:5), questo biotipo è stato isolato anche in capre in
Nuova Zelanda e in Australia. Il biotipo 4 è stato isolato in Nuova Zelanda nel contenuto intestinale
di agnelli, inoltre sono stati isolati ceppi di Yersinia enterocolitica del biotipo 2 ( O: 5,27 e O: 9)
nei cervi.
In Germania sono stati isolati ceppi appartenenti al biotipo 4 (O: 3) e al biotipo 2 (O: 9) in pollame,
questa probabilmente è l'unica volta in cui sono stati isolati questi biotipi patogeni in carne di
pollame e non vi è stata alcuna possibilità di contaminazione crociata con suini o altri animali.
I ratti sono serbatoi di infezione per ceppi del biotipo 1B (O: 8, O: 21) in Giappone, in Nord
America e negli Stati Uniti.
Pozzi, fiumi e laghi sono soggetti alla contaminazione fecale da parte di animali selvatici e
domestici, però i ceppi isolati a livello ambientale appartengono al biotipo 1A o ad altre specie di
Yersinia non patogene (EFSA 2007).
12
Tab. 1: Correlazione tra specie animali e biotipi /sierotipi di Yersinia enterocolitica (EFSA 2007).
Specie Biotipo SierotipoAnimale
suini 4-2 O:3 O:9 O:5,27
ovini 4-5 O:2 O:3
cervi 2 O:5,27 O:9
pollame 4-2 O:3 O:9
animali 4 O:3 domestici
roditori 1B O:8 O:21

Yersinia enterocolitica è stata anche isolata nelle mosche trovate nelle porcilaie e nelle cucine,
questo suggerisce che questi artropodi, vettori/insetti, possono giocare un ruolo importante nella
trasmissione delle yersinie enteriche tra animale e uomo. Le mosche possono anche facilitare la
diffusione di malattie nosocomiali, questo è di particolare interesse perché, è stato segnalato almeno
un caso di mosche che trasportano ceppi resistenti agli antibiotici di batteri appartenenti al gruppo
delle Enterobacteriacee in ospedali libici (Rahuma N., et al 2005).
Le specie di Yersinia enterocolitica sono ampiamente distribuite nell'ambiente terrestre e nelle
acque di laghi, pozzi e ruscelli, che costituiscono le fonti di contagio per gli animali a sangue caldo.
Nel 1976 il latte al cioccolato fu confermato come il veicolo di infezione in un focolaio di malattia
intestinale nei bambini in età scolare. L' agente patogeno, identificato come Yersinia enterocolitica
sierotipo O:8, fu isolato da uno dei quattro cartoni di latte al cioccolato testate (Francis D.W., et al
1980).
L'epidemiologia di una malattia alimentare è strettamente legata ai sierotipi. Yersinia
enterocolitica è la terza causa di diarrea batterica in Francia e Europa ( Lahuerta A. et al 2011).
Episodi assai significativi di intossicazione alimentare sono stati segnalati in Sud Africa, Canada,
Giappone ed USA; sporadiche segnalazioni nei Paesi Europei (Del Bono, Renon 2001 ).
La distribuzione geografica di Yersinia enterocolitica è diversa. Yersinia ha più di 50 sierotipi
distinti ( sulla base delle variazioni antigeniche sulla membrana lipopolisaccaridica delle cellule) e
pochi di essi sono patogeni. Il sierotipo O: 8 è il patogeno principale negli USA seguito da O: 3, O:
5, 27, O: 13 a, 13 b, O: 20, O: 9.
Il biosierotipo patogeno 4/O: 3 è quello più comune nei paesi industrializzati (Bottone E. J. 1999) e
più comunemente isolato nell'uomo in Europa, Cina seguito da O: 9 e O: 8.
Inoltre, vari sierotipi mostrano una specificità geografica, per esempio, il sierotipo predominante in
13

Australia, Europa e Canada è O : 3, in Giappone, Scandinavia e Olanda è predominante il sierotipo
O:9 ( Grahek-Ogden D. et al 2007).
In Polonia nel 2012, furono riportati circa 10.054 casi di infezioni batteriche intestinali, il tasso di
incidenza era del 26,1 per 100 mila abitanti. Nonostante l'infezione di Yersinia rappresentasse
soltanto il 2% dei casi totali (l'incidenza era del 0,6 per 100 mila abitanti), rimaneva un fattore
eziologico importante nelle infezioni gastrointestinali. Furono individuati 231 casi di yersiniosi
inclusi 201 di yersiniosi intestinale e 30 casi di yersiniosi extraintestinale. Il numero di casi fu
minore rispetto ai casi riscontrati nel 2011 ( 257 casi, con un' incidenza del 0,67 per 100 mila
abitanti). La percentuale di ospedalizzazione risultò minore rispetto ai cinque anni precedenti, ma
rimase comunque elevata, nei bambini tra 0-4 anni di età si ridusse significativamente rispetto agli
anni precedenti (2011-76%, 2012- 52%).
L'infezione con il sierotipo O: 3 risultò predominante e rappresentava il 96,7% di tutti i ceppi
isolati in Polonia nel 2012 (Kaminska S. - Sadkowska-Todys M. 2014).
14
Tab. 2: Correlazione tra distribuzione geografica,sierotipi e biotipi di Yersinia enterocolitica(Bottone1997).
Distribuzione Sierotipo Biotipo
Geografica
O:8 1B
Stati uniti O:3 4
O:3 4
Canada O:3 4
Europa O:3 4
O:9 2
Giappone O:3 4
Sud Africa O:3 4

Il numero dei casi è largamente sottostimato perché' l'isolamento di questo batterio dalle feci è
difficoltoso. In Francia ed in Europa i casi di yersiniosi si manifestano sporadicamente, anche se
sono stati descritti alcuni focolai di infezione.
L'aumento del tasso di yersiniosi in Francia negli anni 2000 è stato transitorio, e probabilmente
causato da condizioni ambientali locali che favorirono l'esposizione/trasmissione di Yersinia
enterocolitica patogena (Martin. L. et al 2015).
In Toscana, tramite i dati a noi pervenuti dal Centro di Riferimento Regionale sulle Tossinfezioni
Alimentari (CeRRTA), che fanno riferimento al periodo tra l'anno 2002 e il 2012, si sono verificati
focolai sporadici di yersiniosi. In ogni caso, non si è evidenziata una stretta correlazione tra il
focolaio d'infezione e la presenza di Yersinia enterocolitica negli alimenti. Soltanto durante l'anno
2007, in un bambino di 5 anni, che presentava la sintomatologia clinica della yersiniosi, si è potuta
individuare anche la categoria di alimenti incriminata. L'infezione, infatti, avvenne tramite
l'ingestione di carne, o prodotti derivati, inadeguatamente cotti in ambiente domestico.
Tuttavia, l'epidemiologia dell'infezione è complessa e in parte ancora non del tutto compresa
(Fredriksson -Ahoma et al 2010).
15

2.2 Caratteristiche biochimiche, metaboliche, colturali e condizioni ottimali di crescita
Yersinia enterocolitica condivide molte delle caratteristiche dei germi appartenenti alla famiglia
delle Enterobacteriaceae: forma bastoncellare, gram negativo, ossidasi negativo, anaerobio
facoltativo e asporigeno. Fermenta il glucosio con produzione di acidi, ma non di gas e il lattosio è
fermentato molto lentamente. Questa specie è generalmente mobile tramite flagelli peritrichi a
25-30°C, mentre la mobilità è assente o molto ridotta se incubata a 37°C.
Una caratteristica del batterio, rispetto agli altri membri appartenenti alla famiglia, è quella di
replicarsi a temperature comprese tra 0°C e 44°C: il tempo minimo di replicazione è di 34 minuti a
una temperatura di 20-25°C.
Yersinia enterocolitica è psicrofila, quindi cresce bene a basse temperature ( intorno ai 4°C), questo
permette la sua moltiplicazione anche in alimenti refrigerati. Inoltre, il batterio è in grado di
resistere al congelamento e sopravvivere in cibi congelati per lunghi periodi; perfino dopo ripetuti
cicli di congelamento e scongelamento (Toora et al.1992). Anche un basso numero iniziale di germi
è in grado di proliferare fino a raggiungere i livelli di pericolosità senza alterare i caratteri
organolettici dell'alimento stesso.
Il congelamento non risulta essere stressante per il microrganismo, mentre una buona inibizione
della crescita avviene nei prodotti lattiero-caseari sottoposti a pastorizzazione e maturazione
(Andreis, Ottaviani 2002).
Yersinia enterocolitica può proliferare in cibi come carne e latte a temperature prossime ma anche
al di sotto dello 0° C (Bari Md. L. et al 2011).
Il batterio è sensibile al riscaldamento essendo distrutto dalle tecniche di pastorizzazione a 71,8°C
16

per 18 secondi ( Toora et.al 1992).
Studi dimostrano, che in cibi con pH neutro conservati a una temperatura di 5°C, Yersinia
enterocolitica subisce un aumento da 10 UFC/ml a 2.8x107 UFC/ml in 5 giorni. La produzione di
tossina da parte di questo patogeno è influenzata dalla temperatura di crescita e dalla composizione
dei prodotti alimentari. Yersinia enterocolitica produce enterotossina termo-stabile nel latte a 25°C
ma non a 5°C ( Francis D.W. et al 1980 ). I ceppi che crescono bene a un temperatura di 4°C non
producono livelli significativi di tossina tali da essere rilevata nei topi durante studi in vivo
( Kapperud G. 1987).
La maggior parte delle cellule di Yersinia enterocolitica vengono uccise o alterate quando
conservate durante il deposito a -20 °C. Quando la carne macinata viene inoculata con cellule di
Yersinia enterocolitica e essa viene conservata a -20°C per 30 giorni, circa il 83% delle cellule
inoculate vengono distrutte e il restante 24 % vengono irreversibilmente alterate (Swaminathan B.
et al 1982).
Il minimo valore di pH che viene riportato per Yersinia enterocolitica è compreso tra i valori 4.2 e
4.4 ( Stern N.J. et al 1980).
17
Fig. 2: Caratteristiche di Yersinia enterocolitica.
Bacillo/coccobacillo (pleomorfismo). Gram negativo delle dimensioni di 0,5-0,8 micron per una lunghezza di 1-3micron;Anaerobio facoltativo asporigeno;Catalasi +;Ossidasi-;Glucosio fermentante con produzione di acidi ma non di gas;Lattosio non fermentante (alcuni ceppi sono dei lenti fermentatori);Non produce H2S;Mobile a 25°C, ma non a temperature superiori a 30°C;Temperatura ottimale di crescita 28-30°C;Psicrofilo;pH ottimale di crescita 7.6, tollera fino al 5% di NaCl.

La presenza di acidi organici riduce la capacità di Yersinia enterocolitica di moltiplicarsi a bassi
livelli di pH. L'acido acetico ha un'attività inibitoria maggiore rispetto all'acido lattico e acido
citrico allo stesso valore di pH. Bhaduri nel 2011 eseguì un esperimento variando il pH di prodotti
alimentari a pH 4,5 portandolo a pH 6. Il numero di cellule vitali diminuii, ma il 95 % delle cellule
sopravvissute mantenne il plasmide di virulenza. Tuttavia, a un pH di 3, le cellule che avevano
mantenuto le caratteristiche di virulenza, non riuscirono comunque a sopravvivere.
Un altro fattore di crescita che influenza lo sviluppo del microrganismo è l'attività dell'acqua (Aw),
il valore minimo di Aw che permette la crescita del microrganismo è 0,96. Il batterio è in grado di
sopravvivere ad un ambiente salino al 5% ma non con una salinità al 7%. Con una salinità del 9%
ad una temperatura di 25C° tutte le cellule batteriche muoiono. Bhaduri nel suo esperimento, variò
anche il grado di salinità di alcuni prodotti alimentari (0,5%, 2% e 5%). Il numero di cellule vitale
diminuì anche in questo caso, ma il 96% delle cellule sopravvissute continuò a mantenere le
caratteristiche di virulenza conservando il plasmide, questo indicò che il grado di salinità non aveva
effetto sulla stabilità del plasmide (Bhaduri S. 2011- Bari Md. L. et al, 2011).
Yersinia enterocolitica viene suddivisa in base agli antigeni O, in 6 biotipi e in oltre 60 sierotipi,
molti dei quali non patogeni per l'uomo. I biotipi possono essere distinti in base al loro livello di
patogenicità, soltanto uno di loro non è patogeno (Biotipo 1A). I biotipi 2-5 sono debolmente
patogeni, mentre il biotipo 1B è quello che ha il livello maggiore di patogenicità per l'uomo ed è
letale nei topi e in modelli di infezione. Dei 60 sierotipi conosciuti soltanto undici sono associati
con malattie nell'uomo e la maggior parte di esse possono essere ricondotte a tre sierotipi
comunemente virulenti: 4/O: 3, 1B/O: 8, e 2/O: 9. Questi tre sierotipi sono considerati gli agenti
18

causa del maggior numero di yersiniosi, variabili in base alla geografia (Robins- Browne R.M., et
al 1989). I sierotipi riscontrati più frequentemente nei pazienti affetti da yersiniosi sono: 4/O:3, 2/O:
9, 2/O: 5, 27 e 1B/ O: 8 (Galindo C.L., et al 2011). Il sierotipo 4/O: 3 rappresenta il principale
responsabile delle infezioni. Recentemente i sierotipi: O: 1, O: 2, O: 3, O: 5, O: 8 e O: 9, sono stati
associati a manifestazioni cliniche sia in animali che nell'uomo ( Gyles 2004).
Oltre l'antigene somatico O di natura lipopolisaccaridica, Yersinia enterocolitica possiede un
antigene flagellare H ed un antigene capsulare K. Sono stati individuati 6 diversi antigeni capsulari
(da K1 a K6) e numerosi antigeni flagellari H ( da “a” a “t” ).
La capacità di propagarsi a temperatura di refrigerazione, in alimenti sottovuoto con una prolungata
shelf-life, è di notevole importanza in materia di igiene alimentare. Yersinia riesce a sopravvivere in
cibi surgelati per lunghi periodi di tempo. Yersinia enterocolitica non è in grado di crescere con un
pH minore di 4,2 o maggiore di 9,0 o con una concentrazione salina maggiore di 7% ( Aw< 0,945).
Yersinia enterocolitica non è un microrganismo resistente al calore, quindi non sopravvive a
processi di pastorizzazione o a trattamenti termici come la normale procedura di cottura dei cibi in
ambiente casalingo. I trattamenti termici di carne o latte a 60°C per 1-3minuti inattivano
efficacemente Yersinia enterocolitica. Però, se il livello iniziale del microrganismo è molto elevato,
la completa distruzione può non verificarsi durante la pastorizzazione. Può avvenire solo quando le
cellule vengono trattate a 47°C per 12-70 minuti( Swaminathan B., et al 1982).
L'effetto dell'acido lattico sulla crescita di Yersinia O: 9 è maggiore in condizioni di anaerobiosi
piuttosto che in condizioni di aerobiosi, anche se il batterio ha dimostrato di essere più tollerante a
condizioni di pH bassi in atmosfera anaerobica in assenza di acido lattico (Bari Md. L, et al 2011).
19

2.3 Geni di patogenicità e patogenesi della yersiniosi
Tra i microrganismi capaci di provocare gastroenteriti, Yersinia enterocolitica va assumendo un
interesse crescente. L'incremento della casistica è correlato anche allo sviluppo delle conoscenze sul
germe e a un maggior numero di accertamenti condotti per la sua evidenziazione.
La virulenza di Yersinia enterocolitica dipende dalla sua marcata capacità di penetrare e
sopravvivere e diffondersi nel tessuto intestinale (Cantoni. C. 2011).
Quale via di contagio, si può pensare a una trasmissione diretta di germi da uomo malato a uomo
sano o da animali ( anche se molto rara) a uomo, come pure, per via indiretta, tramite l'acqua, il latte
e altri alimenti contaminati (Cantoni C. 2011).
La via di trasmissione più comune di Yersinia patogena si suppone sia la via oro-fecale, tramite
ingestione di cibo contaminato. La trasmissione diretta persona-persona è rara. Sono stati riportati
casi di neonati esposti all'infezione tramite altre persone, questo avviene quando le basilari norme
20
Tab. 3: Differenze fenotipiche nell'ambito dei biotipi di Y Yersinia enterocolitica (Bottone et al., 1999).
Test Biotipi
1A 1B 2 3 4 5
Salicina (24) + - - - - -Xilosio + + + + - v
Sorbosio + + + + + -Trealosio + + + + + -Sorbitolo + + + + + -
Lipasi + + - - - -Esculina (24) +/- - - - - -
Indolo + + v - - -V-P + + + + +
Nitrati + + + + + -Ornitina + + + + + +(+)
Pyrazimidasi + - - - - -
+(+)

igieniche vengono meno o sono inadeguate. Trasmissione indiretta da persona a persona si è
verificata appartenente in molti casi tramite la trasfusione di sangue infetto. Un rischio comune di
trasmissione per gli allevatori è il loro stretto contatto con i suini. Tuttavia, la trasmissione di
Yersinia enterocolitica tra maiale e uomo non è ancora stata provata. Una delle più importanti fonti
di contaminazione per l'uomo è l'ingestione di carne di maiale cruda o poco cotta, anche se questo
gioca un ruolo limitato nella trasmissione e nello sviluppo della yersiniosi ( Bari Md. L., et al 2011).
Dopo l'ingresso attraverso la via orale, Yersinia enterocolitica migra attraverso lo stomaco ed il
piccolo intestino fino alla porzione terminale dell'ileo. I batteri penetrano la lamina propria della
mucosa passando attraverso le cellule epiteliali, si moltiplicano nelle placche del Peyer e nei
follicoli, causando una tipica lesione detta ileite terminale. Generalmente l'infezione è limitata a
questo distretto, ma alcune volte i microrganismi raggiungono i linfonodi mesenterici e danno
origine ad un'infezione sistemica.
La patogenicità di molti sierotipi di Yersinia enterocolitica (60 circa quelli conosciuti oggi), sarebbe
in parte legata al potere invasivo del germe sulle cellule epiteliali delle mucose intestinali, ed in
21
Tab. 4: Bio-sierotipi patogeni isolati con maggior frequenza (Cattabiani, 2002).
Patogenicità
1A Ambientali*
O:8, O:9, O:4, O:13 a O:13b, O:18, O:20, O:21, 1B Patogeni O:5,27
2 O:9, O:5,27
3 Bassa O:1,2,3 O:5,27Patogenicità
4 O:3
5 O:2,3
* Oggi, le evidenze portano a considerarli come ceppi “opportunistici”.
Biotipo Sierotipo

parte riconducibile alla presenza di enterotossina termoresistente (Andreis, Ottaviani 2002).
Le prime osservazioni sulle basi genetiche della patogenicità di Yersinia enterocolitica risalgono
agli anni '80 e sono relative alla scoperta di un plasmide (Gemski P. et al 1980) e successivamente,
allo studio dei geni a localizzazione cromosomica (Miller V.L., Falkow S. 1988). Il plasmide di
70-75 Kb viene denominato da Portnoy e Falkow pYV, mentre i loci cromosomici sono nominati
INV, AIL, YST (Portnoy D.A., Falkow S. 1981).
Il plasmide pYV (plasmide di virulenza di Yersinia enterocolitica) è presente soltanto nei ceppi
virulenti, codifica per l'espressione di numerose proteine (da 16 a 20 ) localizzate sulla membrana
esterna della parete (Portnoy D.A et al 1984) (Skurnik M. 1985). Il plasmide è un prerequisito per la
piena attività di virulenza di Yersinia enterocolitica, ma può essere facilmente perso durante le
manipolazioni in laboratorio e a seconda delle condizioni di coltura (viene facilmente perso quando
le condizioni di temperatura nel metodo di coltura sono superiori ai 30°C). Quindi è per questo
motivo che la differenziazione dei ceppi patogeni non deve essere fatta soltanto in base alla
presenza del plasmide di virulenza, ma anche in relazione alla presenza dei geni cromosomici
(Haoxuan Z., et al 2008). Una di queste proteine codificate dal plasmide e' YadA, proteina di
membrana esterna, espressa a 37°C ma non a 25°C, di natura fibrillare che ricopre la superficie
batterica e media l'adesione del patogeno alle cellule della mucosa intestinale, specialmente a
livello della zona ileo-cecale ( Kapperud G. et al 1987). Inoltre la sua presenza induce un
incremento dell'idrofobicità di superficie con conseguente aumento della resistenza alla fagocitosi,
conferisce attitudine di agglutinazione ed inibisce l'attivazione del complemento. Recentemente è
stata individuata un'ulteriore funzione di protezione del microrganismo, eventualmente fagocitato,
dai processi litici ossigeno- dipendenti, con la conseguenza che macrofagi e neutrofili divengono
essi stessi dei veicoli di disseminazione (Cattabiani F. 2002). La presenza di YadA promuove
l'adesione del batterio al muco, in questo modo lo strato di muco, solitamente posto a difesa, diventa
22

un elemento facilitante la colonizzazione del batterio; il microrganismo intrappolato nel muco si
moltiplica più rapidamente, metabolizzando sia la mucina stessa, sia gli altri componenti, inoltre il
rivestimento di muco trasforma l'idrofobicità di superficie in idrofilia, condizione che facilita l'
adesione all'epitelio ( Paerregaard A. et al 1991).
I geni cromosomici, AIL e INV, codificano per proteine di membrana, denominate proteine
d'ingresso, che coadiuvano l'adesione alla mucosa e ne condizionano l'invasione. L'ingresso nelle
cellule mucosali, possibile solo nei ceppi ail+ e inv+, consente il raggiungimento del sito di elezione
del microrganismo rappresentato dalle cellule M delle placche di Peyer; successivamente, viene
trasportato dalle cellule M nella lamina propria della sottomucosa, dove si moltiplica, e da dove,
per via linfo- ematogena, può disseminarsi in tutto l'organismo ( Cattabiani F. 2002).
La presenza del gene AIL coadiuva il plasmidio pYV nell'aumentare la resistenza all'attività
battericida del siero ( Bliska J.B. 1992), caratteristica che si esprime al massimo grado nella fase
23
Fig. 3: Meccanismo di colonizzazione dell' epitelio intestinale (Fabrega A., Vila J.2012).

stazionaria di crescita, piuttosto che nella fase logaritmica. AIL è altamente correlato con la
virulenza del batterio, viene ritrovato esclusivamente nei ceppi patogeni di Yersinia enterocolitica, a
differenza del gene INV che invece è presente in tutti i ceppi di Yersinia isolati. AIL è coinvolto
nell'adesione e nel processo di invasione delle specifiche colture cellulari, così come nella
sopravvivenza contro gli effetti battericidi del siero ( Fabrega A., Vila J. 2012).
Il gene INV codifica per una proteina d' ingresso denominata anche “ invasina”, che dà inizio
all'ingresso nelle cellule epiteliali legandosi a recettori specifici ( Cattabiani F. 2002).
Ceppi mutanti Inv- aderiscono alle cellule M, ma in misura minore rispetto ai ceppi wild-type e il
loro livello potenziale di colonizzazione delle placche del Peyer è notevolmente ridotto (Bari Md.
L., et al 2011).
Prima che il batterio stabilisca uno stretto contatto con l'epitelio intestinale, è stato riportato che i
flagelli e, quindi, la motilità, svolgono un ruolo importante nell'iniziare l' invasione della cellula
ospite. L'inattivazione dei geni che codificano per i flagelli, richiesti per l'espressione della motilità,
è associata con la diminuzione del grado di invasività paragonabile a quella riscontrata nei ceppi in
cui il gene INV è stato inattivato (Fabrega A., Vila J. 2012).
Infine, il gene YST, codifica per l'elaborazione di un' enterotossina termostabile denominata Yst
(Pai C.H. 1978). Il significato patogenetico dell'enterotossina, che si forma soltanto a una
temperatura inferiore ai 30°C, è ancora poco chiara. Dopo essere stata riscaldata a 100°C per 20
minuti la tossina è ancora attiva (Cantoni C. 2011). L'enterotossina è resistente alla cottura, ma
sembra essere coinvolta solo nelle fasi iniziali e nelle forme lievi della malattia ( Ist. Zooprofilattico
Sperimentale della Lombardia e Emilia Romagna 2004).
La specie Yersinia è suddivisa in 6 biotipi, 1A,1B,2,3,4,e 5. I biotipi patogeni per eccellenza sono il
1B e 2-5, grazie alla presenza del plasmidio pYV, dei geni cromosomici AIL e INV e del gene
codificante per l'enterotossina YST A. Il biotipo 1A è stato in passato considerato non patogeno.
24

Tuttavia, i ceppi appartenenti al biotipo 1A sono stati associati con focolai nosocomiali e di origine
alimentare di diarrea. Studi hanno dimostrato, che gastroenteriti causate dai ceppi appartenenti al
biotipo 1A sono indistinguibili da quella causate dai biotipi patogeni primari. Gli studi di Grant et
al, nel 1998-1999, hanno provato che alcuni ceppi appartenenti al biotipo 1A di Yersinia
enterocolitica, invadevano le cellule epiteliali e resistevano all'uccisione da parte dei macrofagi,
seppur in misura minore rispetto ai patogeni primari. I biotipi patogeni primari hanno la capacità di
produrre un'enterotossina termostabile YST A, la quale contribuisce alla virulenza del batterio. Il
ruolo delle altre varianti della proteina YST ( la forma b e c), soprattutto per il biotipo 1A, non è
ancora conosciuta. Questo principalmente perché, la tossina viene prodotta in vitro solo a una
temperatura sotto ai 30°C. Inoltre, la presenza del gene YST non è necessariamente correlata con la
produzione della tossina in vitro (Grant T., et al 1998- Grant T. et al 1999).
La produzione di enterotossina è comunque un fattore di virulenza molto importante nell'instaurarsi
dell'infezione, inoltre, è stato dimostrato che i ceppi appartenenti al biotipo 1A, hanno la capacità
anch'essi di produrre una forma di enterotossina termostabile a una temperatura di 28°C,
denominata YST B. Studi recenti hanno dimostrato che il gene YSTB è ampiamente distribuito nei
ceppi appartenenti al biotipo 1A, e che la produzione della tossina può essere indotta dalle
condizioni di pH presenti nell'ileo. Questo suggerisce come YST B sia un importante fattore di
virulenza nei batteri facenti parte del biotipo 1A di Yersinia enterocolitica (Singh I., Virdi J.S.
2004). Thoerner et al nel 2003 individuò che il genotipo predominante nell' 80% dei ceppi isolati di
Yersinia enterocolitica appartenenti al biotipo 1A, presentava il gene YST B, dimostrando che
questi ceppi erano privi del plasmide di virulenza, ma per esempio, il gene INV era presente ma non
funzionale in molti dei ceppi (Peruzy M.F. 2014). L'assenza di un marcatore per la virulenza in
ceppi del biotipo 1A isolati da casi clinici umani, può essere illustrativa della patogenicità
intrinseca debole di questo biotipo. Il risultato clinico è probabilmente determinato più da fattori
25

dell'ospite, che da fattori di virulenza batterica (Thoener P.,et al 2003).
In “vitro” i ceppi yst+, enterotossigeni, elaborano enterotossina solo a temperatura ambiente o
comunque non superiore a 30°C, ma non a 37°C: il gene, cioè, viene represso a questa temperatura.
Nonostante questa evidenza, alcune indagini effettuate in vivo su suini, topi e conigli hanno
evidenziato risultati contrastanti (Cattabiani F. 2002). Ciò nonostante, il biotipo 1A, considerato
inizialmente non patogeno, può causare diarrea e recenti studi dimostrano, che frequentemente,
presenta una variante della tossina yst. Si pensa che questo sia il suo unico mezzo di virulenza che
gli permette di indurre una sintomatologia diarroica (Frabrega A., Vila J. 2012).
Dal momento che la via usuale di infezione è rappresentata dal consumo di alimenti contaminati, il
patogeno deve in primo luogo adattarsi al diverso livello di temperatura. L'adattamento è mediato
dal plasmide pYV. Lian C.J. e altri ricercatori, in uno studio del 1987, hanno evidenziato come dopo
già 6 ore dalla somministrazione orale al coniglio di ceppi pYV+, coltivati a 25°C, le proteine di
superficie codificate dal plasmide venivano espresse nell'ambiente del piccolo intestino. Si presume
che le cellule, incubate a 25°C, abbiano un limitato numero di copie di queste proteine o, esse siano
presenti a livello citoplasmatico e inizino ad essere trasportate sulla membrana esterna quando la
temperatura raggiunge i 37°C ( Portnoy D.A. et al 1985- Lian C. J. et al, 1987). Con quanto detto, si
pensa che Yersinia enterocolitica sia sottoposta ad un ciclo di trasmissione “caldo-freddo”, durante
il quale il microrganismo, provvisto di tutti gli attributi di virulenza, viene eliminato da un ospite a
sangue caldo, contamina acqua o alimenti, regolando o abolendo il suo corredo di fattori di
patogenicità, e provvede di nuovo ad aumentare rapidamente il numero delle copie delle sue
proteine di membrana, non appena torna a trovarsi a temperature di 35-37°C. Sembrerebbe, quindi,
che esista, nell'ambito dei numerosi fattori di patogenicità di Yersinia enterocolitica, una
26

cooperazione armonica in funzione della temperatura ambientale. In un primo momento, in seguito
ad ingestione di alimenti contaminati, e quindi, in condizioni termiche inferiori a 35-37°C, i ceppi
virulenti farebbero ricorso a determinanti di patogenicità sotto il controllo cromosomico,
successivamente, in seguito all' acclimatamento alla temperatura dell'ospite, esprimerebbero i fattori
di patogenicità sotto il controllo plasmidico, comprendenti i meccanismi atti a contrastare le difese
dell'organismo, la fagocitosi, la devitalizzazione ossigeno-dipendente e l'attivazione del
complemento. In molti ceppi patogeni, molti dei fattori di virulenza sono associati alla temperatura,
la quale è molto importante nello stabilire l'infezione. A temperatura ambiente ( meno di 28°C) e in
condizioni di acidità a 37°C, Yersinia patogena esprime ottimamente la proteina invasina,
codificata dal locus cromosomico INV. Dopo l'ingestione l'invasina si lega all'integrina β1 sulle
cellule dell'ospite facilitando la penetrazione del batterio negli strati epiteliali. Il graduale aumento
di temperatura all'interno dell'ospite induce l'espressione di fattori di virulenza necessari a creare
una roccaforte all'interno dei tessuti linfatici, ed eludere il rilevamento da parte del sistema
immunitario. L'espressione del locus cromosomico AIL, per esempio, avviene a 37°C, e la risultante
proteina ail migliora l'invasione a livello delle cellule epiteliali.
La proteina di adesione A (YadA), oltre a mediare l'adesione al muco e alle cellule dell'epitelio, in
concerto con l'invasina, promuove l'invasione dell'ospite. YadA, come già detto in precedenza, è una
proteina multifunzionale, fattore di virulenza di superficie codificata dal plasmide pYV, che
conferisce al batterio la capacità di aderire alle proteine della matrice extracellulare. L'induzione
dell'espressione di questa proteina è coordinata con l'up-regulation delle proteine Yop ( proteine di
membrana esterna di Yersinia). Similmente all'invasina, YadA avvia l'internalizzazione tramite il
legame con una fibronectina che è legata a l'integrina 5β1. YadA lega la fibronectina, il collagene
di tipo I,II e IV e la laminina, sebbene con affinità differenti, favorendo in tal modo proprietà di
virulenza variabili. YadA sollecita una risposta infiammatoria nelle cellule epiteliali, tramite la
27

produzione dell'interleuchina 8 (IL-8) attraverso la proteina MAPK, contribuendo, così, alla cascata
infiammatoria. L'interazione di YadA con le fibre di collagene contribuisce all'instaurarsi della
yersiniosi cronica, con l'insorgenza di artrite, come è stato dimostrato nei modelli animali. In
aggiunta all'inibizione e invasione delle cellule dell'ospite, entrambe AIL e YadA, giocano un ruolo
importante nella resistenza al complemento e nell'evasione del sistema immunitario. Infatti,
inibiscono la via alternativa del complemento legandosi al fattore H, usurpando la sua funzione
naturale per prevenire la lisi delle cellule dell'ospite. AIL e YadA, similmente, sovvertiscono anche
la via classica del complemento e la via della lecitina, legandosi alla C4b-binding protein,
promuovendo così la degradazione del fattore C4b, prevenendo la formazione della C3 convertasi,
che altrimenti porterebbe alla lisi delle cellule batteriche (Galindo C.L., et al 2011).
Una volta all'interno dell' ospite, il microrganismo sopravvive all'attacco dei macrofagi esprimendo
proteine coinvolte in una strategia antifagocitaria tramite l'iniezione, grazie alla codifica di un
sistema di secrezione di tipo III, di tre proteine effettrici: YopH, T ed E, le quali determinano la
distruzione del citoscheletro del macrofago. YopH è una tirosina-fosfatasi, defosoforila la paxillina
e la proteina FAK, che sono coinvolte nell'assemblaggio del complesso citoscheletrico necessario
nel processo di fagocitosi. YopT provoca la depolimerizzazione dei filamenti di actina tramite una
ridistribuzione delle RhoA GTPasi. YopE determina l'inibizione delle GTPasi associate alle
proteine Rho coinvolte nella fagocitosi (Bari Md. L., et al 2011).
Le Yersinie virulente attraversano il lume intestinale, si attaccano e penetrano la barriera mucosa
sovrastante le cellule epiteliali della mucosa, ed eventualmente aderiscono alle cellule intestinali. Le
cellule batteriche hanno dimostrato di legarsi preferenzialmente e penetrare nelle cellule M delle
placche del Peyer. Una volta internalizzati, i batteri vengono quindi trasportati attraverso la barriera
epiteliale ed espulsi dal lato basolaterale delle cellule M. L'evidenza suggerisce che nelle primi fasi
28

di infezione, i fagociti internalizzino i batteri.
E' stato dimostrato che i batteri internalizzati si replicano nei macrofagi murini nativi, quindi, si
presume, per essere trasportati all'interno della migrazione dei fagociti ai linfonodi mesenterici,
provocando una risposta infiammatoria che causa dolore addominale.
Inoltre, fagociti che hanno internalizzato i batteri possono diffondere, attraverso il flusso
sanguigno, al fegato e della milza. Una volta che si è localizzata a livello delle placche del Peyer,
dei linfonodi mesenterici, del fegato e della milza, Yersinia enterocolitica si replica in una forma
extracellulare tramite la formazione di micro-ascessi. All'interno di queste lesioni le cellule
batteriche formano microcolonie che appaiono resistenti alla fagocitosi da parte di macrofagi e
neutrofili ( Fabrega A., Vila J. 2012).
Il complesso T3SS (sistema di secrezione di tipo III), che è codificato dal plasmide, gioca un ruolo
sostanziale sia nello stabilire che nella riuscita dell'infezione. Il T3SS, ingettosoma, fa da ponte tra
la membrana interna e quella esterna del batterio, e le proteine Yop effettrici vengono traslocate
nella cellula ospite tramite la proteina di secrezione F (YscF). Le proteine YopB e YopD formano
29
Fig. 4: Meccanismi di evasione dell'attività macrofagica di Yersinia enterocolitica( Bari Md. L., et al 2011).

un poro attraverso la membrana della cellula ospite, consentendo l'aggancio e l'eventuale
traslocazione delle proteine effettrici. Per il corretto assemblaggio dell'ingettosoma, sono richieste
anche le proteine chaperone citoplasmatiche YscE e YscG.
Esistono 6 proteine Yop effettrici (YopE, YopH, YopP/J, YopO/ YpkA e YopM), che mediano
l'evasione del sistema immunitario, interferendo con il normale pathway di trasduzione del segnale,
distruggendo i filamenti di actina del citoscheletro, e inducendo il processo di apoptosi nella cellula
ospite.
La consegna delle proteine Yop richiede un contatto stretto tra la cellula batterica e la cellula ospite,
questo è mediato dalla presenza di YadA e dall'invasina, le quali legano le β1 integrine, le quali
stimolate, determinano l'attivazione delle chinasi Src e RhoA, che facilitano la traslocazione delle
proteine Yop modulando la polimerizzazione dei filamenti di actina.
La formazione dell'igettosoma, in vitro, è determinata dalla temperatura, 37°C, e da una bassa
concentrazione di calcio. Le proteine Yop evadono la risposta immune bloccando le funzioni di
fagocitosi dell'ospite.
La morte della cellula ospite è mediata dalla proteina YopP/J, la quale è una serina-treonina
acetiltrasferasi, che induce l'attivazione di meccanismi apoptotici. Recentemente, si è individuato
che YopP/J induce apoptosi tramite attivazione delle caspasi, indipendentemente dall'attivazione dei
recettori di morte.
Una volta all'interno della cellula bersaglio, YopE,H,P e T cooperando distruggono il citoscheletro
delle cellule epiteliali, dei macrofagi e delle cellule dendritiche, diminuendo la loro capacità di
fagocitare il batterio.
YopP/J può anche facilitare l'evasione del sistema immunitario adattativo, inibendo la capacità
della cellule dendritiche di presentare l'antigene ai linfociti T CD8+ direttamente o eventualmente,
riducendo la popolazione delle cellule dendritiche tramite induzione di apoptosi (Galindo C.L., et al
30

2011). Haoxuan Zheng et al, nel 2008, ha scoperto che non tutti i ceppi patogeni trasportano tutti i
geni di virulenza tradizionali in entrambi i cromosomi e plasmidi causando la malattia. Forse alcuni
di loro mancano di alcuni geni di virulenza tradizionali, contenendo altri marcatori di virulenza
sconosciuti, che interagiscono tra loro e svolgono un ruolo importante nella patogenesi dei diversi
ceppi patogeni di Yersinia enterocolitica ( Haoxaun Z. et al 2008).
L'infezione da Yersinia enterocolitica è bifasica: ha inizio con una fase di quiescenza di circa 36-48
ore, in cui il batterio si replica senza una risposta misurabile da parte dell'ospite. Questa fase iniziale
è seguita da un afflusso di fagociti attivati nei tessuti infetti e nei linfonodi, che inducono una
riposta infiammatoria acuta caratterizzata da formazione di citochine e formazione di tessuto
necrotico. Il sistema di secrezione di tipo III è responsabile della fase iniziale dell' inibizione della
fagocitosi, ma i meccanismi che stanno dietro tale improvviso sistema “off-on” nella risposta
infiammatoria, sono attualmente non pienamente compresi. Il complesso T3SS è richiesto per
l'effettiva colonizzazione a livello sistemico, e la sua inattivazione conduce a una rapida
31
Fig. 5: Eventi molecolari nella cellula macrofagica indotta da Yersinia enterocolitica (Galindo L.C., et al 2011).

eliminazione del batterio dall'organismo umano.
Recenti studi hanno dimostrato, che i magrofagi riescono a compensare l'azione inibitrice delle
proteine Yop sul pathway di presentazione dell'antigene, tramite un meccanismo di autofagia
dipendente. Studi sui topi hanno provato, che sono necessari i linfociti CD4+ e CD8+ per
controllare l'infezione di Yersinia. L'inibizione della proliferazione delle cellule T e delle funzioni
delle cellule dendritiche, è il meccanismo primario tramite cui il batterio evade sia la risposta
innata che adattativa del sistema immunitario. Yersinia induce apoptosi sia in macrofagi naive sia in
macrofagi attivati, questo è coerente con il processo di infezione bifasico.
L' aumento dell' infiammazione associata alla morte delle cellule dell'organismo ospite, potrebbe
inizialmente essere un beneficio per Yersinia, ma in seguito potrebbe contribuire ad una risposta
immunitaria generalizzata ed a un'eventuale eliminazione del batterio (Galindo C.L., et al 2011).
Yersinia enterocolitica è un patogeno zoonotico che causa gastroenteriti acute e occasionalmente
malattie di grave entità nell'uomo. La yersiniosi umana è un'infezione del tratto gastroenterico
autolimitante (spesso non necessità di trattamento antibiotico), caratterizzata da enterite o
enterocolite, talvolta complicata da linfoadenite acuta mesenterica o da ileite terminale, che in
alcuni soggetti può evolvere in forma setticemica ( Bottone E.J. 1999).
Provoca una varia gamma di sintomi che dipendono dall'età del soggetto infettato. Spesso
l'infezione si verifica nei bambini al di sotto dei 5 anni (Stefanelli S., et al, 1986).
La maggior parte delle yersiniosi si verifica sporadicamente nei bambini. I sintomi predominanti
sono febbre, dolore addominale, diarrea spesso con presenza di sangue.
Nei bambini più grandi e negli adulti, le conseguenze della yersiniosi sono gravi e comprendono
infezioni acute, pseudoappendiciti e una serie di eventi extraintestinali per un lungo periodo, come
artrite e eritema nodoso.
32

Il tasso di mortalità è pari al 0,02% ( Del Bono, Renon 2001 ).
Il genere Yersinia comprende ceppi in grado di causare malattia nell'uomo e negli animali. E' un
batterio potenzialmente enteropatogeno dotato di una spiccata capacità invasiva, che si trasmette
per ingestione di acqua o alimenti contaminati. E' un germe ubiquitario, di cui molti ceppi non sono
patogeni, solo gli stipiti provvisti di meccanismi di virulenza causano malattia nell'uomo.
Nel suino, portatore asintomatico, il patogeno si localizza a livello dell'amigdala, della lingua, del
palato molle, dei linfonodi e dell' intestino, pertanto anche le feci dell'animale possono costituire
un importante fattore di contaminazione ambientale.
La principale forma clinica della yersiniosi alimentare è quella enterocolitica, contrassegnata dai
seguenti sintomi clinici principali (Andreis, Ottaviani 2002):
• febbre generalmente sempre presente superiore ai 38°C;
• diarrea caratterizzata da frequenti scariche con feci acquose e sanguinolente;
• dolore addominale, frequente e generalmente lieve, localizzato nel quadrante inferiore
destro;
• nausea;
• vomito.
Le complicazioni sono rare, ma molto gravi. Includono ulcerazioni diffuse del piccolo intestino e
del colon, peritonite, perforazione intestinale, megacolon tossico e trombosi delle vene
mesenteriche con cancrena del piccolo intestino.
Il digiuno e l'ileo sono i tratti intestinali più colpiti, sebbene anche il cieco e il colon prossimale
possano essere coinvolti. Inizialmente, la colonizzazione di Yersinia della mucosa intestinale
comporta la formazione di microascessi nella lamina propria. Questi contengono aggregati di
33

coccobacilli e neutrofili che formano masse necrotiche, che possono eventualmente unirsi
all'interno delle erosioni della mucosa. I linfonodi mesenterici sono solitamente edematosi. Nel
corso di molte infezioni acute, le placche del Peyer sono notevolmente alterate, con colonie di
batteri e ascessi necrotici. La formazione dell'ascesso e l'infiltrazione delle cellule infiammatorie,
causano una perdita dell'integrità della mucosa, atrofia dei villi intestinali e ipertrofia delle cripte.
Come risultato del danno alla mucosa, la perdita di liquidi porta a diarrea da malassorbimento. La
diarrea tende a non essere emorragica, a meno di una forma fulminante.
Yersinia enterocolitica può diffondere nella circolazione sanguigna e giungere ai linfonodi
mesenterici ed alle vie linfatiche dove può determinare microascessi epatici, splenici, che possono
avere esito fatale. Se è già presente un danno mucosale, conseguenza di parassiti ad esempio, gli
animali possono essere predisposti a una yersiniosi settica.
Le manifestazioni secondarie sono rappresentate da eritema nodoso e artropatia reattiva che si
manifestano a poche settimane dalla fase acuta.
La disseminazione sistemica, a causa delle sua gravità e del suo quadro clinico, si verifica
prevalentemente in anziani e soggetti immunocompromessi. Spesso l'infezione si presenta come una
sindrome pseudoappendicolare, con dolori addominali. Questa sintomatologia è accompagnata da
febbre, con o senza diarrea. L'importanza di questa forma di malattia è la sua somiglianza con
un'appendicite. Infatti, spesso questa sintomatologia, porta ad un intervento di appendicectomia con
appendice normale. In alcuni pazienti possono manifestarsi fenomeni immunoreattivi, quali una
fastidiosa poliartrite che interessa in rapida successione ginocchia, anche, piedi, dita e gomiti e può
persistere per alcuni mesi, oppure un eritema nodoso. Sono state descritte anche faringiti
(http://www.antropozoonosi.it/Malattie/Yersiniosi/Yersiniosi.php)
L'età dell'ospite, il suo stato di salute e il sierotipo coinvolto nell'infezione sono importanti nel
definire l'evoluzione della malattia.
34

La diagnosi di certezza è basata sull’isolamento del microrganismo da escreti, secrezioni o tessuti
(feci, essudato faringeo, liquido peritoneale, sangue, linfonodi) e con la dimostrazione della risposta
anticorpale specifica mediante test di sieroagglutinazione. La terapia è antibiotica (gentamicina,
cloramfenicolo, chinoloni, doxiciclina, cotrimossazolo) (www.antropozoonosi.it).
L'incidenza della sindrome, dovuta a Yersinia, è stagionale, con un numero minore di epidemie in
primavera e maggiore in Ottobre e Novembre. L'incidenza è massima nei soggetti molto giovani e
negli anziani (Cantoni C. 2011).
La prevenzione della tossinfezione alimentare da Yersinia enterocolitica prevede un aumento delle
norme di igiene generale, l'accurato lavaggio delle verdure e il rafforzamento della catena del
freddo (Andreis, Ottaviani 2002), inoltre, il miglioramento delle condizioni igieniche durante il
trasporto, la macellazione e la lavorazione delle carni. Fondamentale è l'istruzione e
l'aggiornamento di tutto il personale addetto alla produzione e alla preparazione finale dei prodotti
alimentari.
La prevenzione si basa sull'igiene della macellazione, del consumo e della conservazione:
• preparare e manipolare in modo igienico la carne e gli altri cibi, specialmente se questi
devono essere mangiati crudi, il latte deve essere sempre pastorizzato;
• lavarsi accuratamente le mani prima di maneggiare il cibo o di mangiare e dopo aver
maneggiato carne di maiale cruda e dopo contatti con animali;
• purificare in modo appropriato le acque e proteggere le risorse idriche da feci umane e
animali;
• allontanare in modo igienico feci umane, di cane e gatto;
• durante la macellazione dei maiali, la testa ed il collo devono essere trattate con cura per
evitare di contaminare le carni con la faringe molto colonizzata
35

(http://www.antropozoonosi.it/Malattie/Yersiniosi/Yersiniosi.php).
36

2.4 Alimenti di origine animale
2.4.1 Latte e prodotti lattiero-caseari
Si definisce “latte crudo”, il latte prodotto mediante secrezione della ghiandola mammaria di
animali da allevamento, che non è stato riscaldato a più di 40°C e non è stato sottoposto ad alcun
trattamento avente effetto equivalente, questo in base al Regolamento CE n° 853/2004.
La qualità microbiologica costituisce un requisito base del latte, sia di quello destinato a latte
alimentare sia quello avviato alla trasformazione. La carica batterica del latte è soggetta a precisi
limiti di legge per il latte commercializzato, oltre ad essere uno dei parametri su cui si basa il
pagamento differenziato del latte in funzione della qualità. In Europa, il limite legale per il
contenuto di germi nel latte bovino è di 100.000 UFC/ml (REG. CE n. 853/2004).
L'eccessiva contaminazione batterica costituisce un fattore indesiderato nel caso di latte destinato al
consumo, per la riduzione della conservabilità, per il possibile sviluppo di sapori sgradevoli e per la
necessità di ricorrere a trattamenti termici intensi. La carica batterica del latte può costituire,
inoltre, un segnale indiretto di eventuali contaminazioni da microrganismi patogeni. Nel caso del
latte crudo destinato alla vendita diretta, il grado e il tipo di contaminazione batterica diventano
parametri di grande importanza. Anche nel processo di caseificazione la contaminazione batterica
può costituire un problema, sia per la competizione operata nei confronti dei microrganismi
filocaseari, che per eventuali alterazioni, anche gravi, delle caratteristiche dei prodotti indotte dallo
sviluppo di batteri anomali o dalla presenza dei loro enzimi.
Dai dati pubblicati dall' Istituto Zooprofilattico Sperimentale della Lombardia e dell'Emilia
Romagna (IZSLER, 2010), emerge come la qualità del latte in queste regioni, nel corso dell'ultimo
trentennio, abbia fatto registrare importanti miglioramenti sotto il profilo igienico-sanitario. In
37

particolare, la carica batterica totale negli ultimi dieci anni si è abbassata sensibilmente, con un
progresso inizialmente più rapido e successivamente più lento. A fronte di valori medi annuali
assolutamente rassicuranti, l'andamento mensile della carica batterica nel corso degli ultimi quattro
anni ha continuato a mostrare incrementi sensibili in corrispondenza del periodo caldo. Inoltre, è
esperienza comune che molti allevamenti sperimentino periodicamente innalzamenti improvvisi e
anormali della carica batterica del latte, le cui cause non sono sempre facilmente identificabili
(Quaderni di ricerca n.119- ottobre 2010. Lombardia).
La carica batterica del latte dipende in massima parte dalle condizioni igieniche in cui si svolgono i
diversi processi che costituiscono la filiera del latte. Per quanto riguarda specificamente la fase
produttiva, che si svolge presso la stalla, la contaminazione batterica è influenzata da una
molteplicità di fattori tra cui: le condizioni igieniche in allevamento, il grado di pulizia degli
animali e delle mammelle, la sanità degli animali, la preparazione della mammella in premungitura,
la modalità di lavaggio dell'impianto di mungitura e le condizioni del latte fino al momento del
ritiro (Kelly et al. 2009). Inoltre, le condizioni ambientali di temperatura e umidità possono influire
notevolmente sulla contaminazione batterica del latte, sia in termini qualitativi, che quantitativi
( Quaderni di ricerca n 119-2010. Lombardia).
Generalmente, il latte presente nelle mammelle delle bovine sane è esente da germi, ma già durante
la mungitura, passando attraverso la parte inferiore del condotto lattifero, viene contaminato dai
microrganismi della flora mammaria (micrococchi, stafilococchi coagulasi- negativi, streptococchi,
corinebatteri ecc.). Nella mungitura a mano, attraverso le attrezzature (secchi), l'aria della stalla,
l'epidermide mammaria, il pelo della bovina, le mani e i vestiti del mungitore, vengono trasmessi al
latte microrganismi della flora degli escrementi e dei mangimi (streptococchi, specie di
Lactobacillus, micrococchi, specie di Bacillus, clostridi, batteri propionici, lieviti, muffe ecc.)
(Cantoni C. 2011).
38

Le tre principali fonti di contaminazione microbica del latte sono rappresentate dalla mammella
stessa, successivamente alla sintesi, in particolare quando siano presenti infezioni mastistiche, dalla
cute dei capezzoli e dalle superfici dell'impianto di mungitura e dal tank di stoccaggio.
In una bovina sana la contaminazione derivante dalla mammella è molto contenuta; al momento
dell'estrazione dalla mammella, il numero di microrganismi presenti nel latte risulterebbe infatti
molto basso e la carica batterica difficilmente supererebbe il valore di 1000 UFC/ml (Kurweii-
Busse 1973). Una bovina affetta da mastite clinica o subclinica può invece rilasciare un gran
numero di microrganismi nel latte, questo latte solitamente viene scartato.
Da un'esperienza danese (Holm et al, 2004), condotta su campioni di latte di massa provenienti da
75 aziende, caratterizzate da alte conte batteriche nel latte, emerse che, nella maggior parte dei casi
(64%), l'elevata carica batterica del latte era dovuta a batteri caratteristici delle situazioni di scarsa
igiene (della stalla, dell'impianto di mungitura ecc.), nel 28% dei casi l'elevata carica batterica
risultò ascrivibile alla proliferazione di batteri psicrotrofi, caratteristici delle situazioni di
inadeguata conservazione del latte in stalla, e solo nell'8% dei casi vi era una prevalenza di batteri
tipici della mastite.
La mammella e i capezzoli delle bovine risultano inevitabilmente sporchi di deiezioni e di
materiale di lettiera. La carica batterica della lettiera è compresa tra 108 e 1010 UFC/g e il latte di
bovine con mammelle sporche e non sottoposte a pulizia dei capezzoli prima della mungitura, può
contenere una carica batterica più di 105 UFC/ml (Bramley- McKinnon 1990). Si è osservata una
relazione stretta tra pulizia della mammella e la conta batterica del latte (Elmoslemany et al 2009).
I batteri prevalenti sulla superficie dei capezzoli e della mammella includono: Micrococcus spp,
Staphylococcus spp, Enterococcus spp, Bacillus spp, coryneformi, coliformi e altri batteri gram
negativi. Inoltre, sono comunemente presenti batteri termodurici e psicrotrofi. In particolare la
presenza di coliformi nel latte è considerata indice di contaminazione ambientale e fecale e quindi,
39

di scarsa igiene in stalla e in mungitura (Reinemann et al 1997). L' influenza dei batteri presenti
sulla superficie dei capezzoli, sulla conta batterica del latte, dipende dal grado di imbrattamento
delle mammelle, dalla percentuale di bovine con mammelle sporche e dalla procedura di pulitura
dei capezzoli prima delle fasi di mungitura. L'imbrattamento della mammella e dei capezzoli
dipende da molti fattori connessi con la gestione della stalla: il tipo di stabulazione ( fissa, libera
con cuccette, libera con lettiera permanente), la frequenza di rinnovo della lettiera, il tipo di lettiera
(paglia, segatura, sabbia, materassini di gomma ecc.), la frequenza di pulizia delle aree di esercizio
e di alimentazione. Anche il dimensionamento delle cuccette e il loro numero rispetto agli animali
accolti, possono influenzare la frequenza di decubito lungo le corsie, con effetti importanti sulla
pulizia delle bovine e delle mammelle.
La pulizia dei capezzoli con una soluzione disinfettante (pre-dipping), seguita da accurata
asciugatura, sembra rappresentare una delle migliori soluzioni per ridurre la contaminazione del
latte derivante dalla presenza dei batteri presenti sui capezzoli. Tale operazione, consentirebbe di
ridurre la carica batterica presente sulla superficie dei capezzoli del 75%. I prodotti impiegati per il
pre-dipping possono contenere principi attivi con attività battericida differente (Quaderni di Ricerca
n-119 2010 Lombardia ).
Anche l'eliminazione dei primi getti di latte, prima dell'attacco del gruppo di mungitura, consente di
ridurre la contaminazione batterica del latte nel tank, in quanto allontana la porzione di latte
contaminata perché più vicina all'orifizio capezzolare.
Nella mungitura meccanica il maggior pericolo di contaminazione è dovuto a insufficiente pulizia e
disinfezione delle superfici interne delle macchine da mungitura, delle condutture e dei bidoni. Alle
superfici interne possono aderire microrganismi formando biofilm viscosi, nei quali si possono
insediare altri microrganismi patogeni. Nel latte crudo munto meccanicamente può, quindi, trovarsi
40

una miscela di micrococchi, stafilococchi, streptococchi (streptococchi lattici, enterococchi), bacilli
gram negativi (Pseudomonas, Enterobacteriaceae ecc. ), specie di Lactobacillus e di Bacillus,
clostridi, batteri propionici, lieviti e muffe (Cantoni C. 2011).
Inoltre, il latte raccolto in contenitori aperti, durante le operazioni di mungitura, può essere
contaminato da individui infetti (o portatori sani) attraverso colpi di tosse o, semplicemente,
attraverso il contatto con le mani (Jay J. M. 2009).
L'igiene dell'impianto di mungitura contribuisce in maniera importante alla contaminazione
batterica del latte di massa. Se il lavaggio dell' impianto non è effettuato correttamente, i depositi
di latte e i residui di acqua di risciacquo che rimangono nelle tubazioni possono rappresentare
ottimi terreni di crescita per molti batteri, tra cui, in particolare, i batteri temodurici, in grado di
resistere all'azione dell'acqua calda impiegata per il lavaggio (Bramley- McKinnon 1990; Holm et
al 2004). Per contro, se il lavaggio è effettuato a basse temperature, si può avere moltiplicazione di
batteri a rapida crescita quali Lactococcus spp, Pseudomonas spp e coliformi (Murphy- Boor 2000).
I punti critici del processo di lavaggio sono molti, tra cui, temperatura, quantità, qualità e
turbolenza del getto di acqua di lavaggio, concentrazione e tipologia di detergente impiegato e
durata delle fasi. L'efficienza del lavaggio è il risultato della combinazione dei processi termici
(temperatura dell'acqua), fisici ( turbolenza e pressione dell'acqua) e chimici ( tipo e concentrazione
dei prodotti detergenti) in un determinato intervallo di tempo (Reinemann et al 2000). Inoltre, è
influenzata dalle caratteristiche dell' impianto; le diverse parti dell' impianto di mungitura sono
costruite in materiali diversi ( gomma, metallo, vetro o plastica) e hanno una diversa
conformazione, caratteristiche che influenzano la facilità di pulizia e di rimozione di residui di latte
o di sali. In particolare le parti in gomma (guaine e tubi), se fessurate o screpolate, possono essere
zone di difficile pulizia, favorevoli alla crescita microbica (Murphy- Boor 2000).
Nel caso in cui venga venduto lo stesso giorno, il latte crudo deve essere raffreddato almeno a 8°C,
41

e nel caso in cui non venga venduto subito, a 6°C al massimo. La refrigerazione favorisce la
conservabilità del latte in quanto riduce la proliferazione microbica, e conseguentemente rallenta la
caduta del pH (legata alla fermentazione del lattosio) e la comparsa di sapori anomali legati al
deterioramento delle componenti grasse e proteiche. La conservazione nel tank porta, tuttavia,
inevitabilmente ad una modificazione della microflora presente nel latte: i microrganismi riescono a
moltiplicarsi anche a basse temperature, gli psicrofili, tendono a prendere il sopravvento. Il latte
prodotto in buone condizioni igieniche, possiede una carica di batteri psicotrofi inferiore al 10%
della conta batterica totale, ma dopo 2 giorni a 4°C la popolazione psicrotrofa diviene dominante
(Gehringer 1980).
Durante il raffreddamento e la conservazione refrigerata del latte, in seguito a un rallentamento
della crescita dei batteri lattici e a una contemporanea crescita dei batteri psicrotrofi o psicrofili,
come Yersinia spp, si verifica una spostamento della carica microbica a favore dei bacilli gram
negativi spesso proteolitici o lipolitici (Cantoni C. 2011).
Gli psicrotrofi sono facilmente distrutti dai trattamenti termici, ma producono enzimi extracellulari
resistenti anche alle temperature di sterilizzazione, aventi azione lipolitica e proteolitica, che
possono alterare fortemente l'aroma e le caratteristiche dei prodotti (sapore amaro, irrancidimento
ecc. ) (Mucchetti- Neviani 2006). Un ulteriore aspetto di cui tener conto è che la refrigerazione
limita la proliferazione dei batteri lattici, batteri fondamentali per l' avvio alla trasformazione
casearia.
Anche Yersinia enterocolitica è stata spesso isolata in diversi studi condotti su campioni di latte
crudo. La prima epidemia documentata causata da questo microrganismo si è registrata negli Stati
Uniti, ed è stata ricondotta al consumo di latte al cioccolato. Yersinia enterocolitica sembra essere
presente maggiormente nel latte crudo (41,5%), mentre Yersinia frederiksenii risulta essere presente
nel latte pastorizzato (56,1%) (Jay J. M. 2009).
42

La contaminazione batterica del latte crudo può essere ridotta scartando il primo getto di latte
munto, avendo cura di lavare bene la mammella della bovina, pulendo accuratamente gli apparecchi
e le macchine, mediante disinfezione con disinfettanti legalmente ammessi e successiva
risciacquatura con acqua di rubinetto. E' molto importante eliminare i residui di caseina, nei quali
possono concentrarsi le Enterobacteriacee proteolitiche e altri bacilli gram negativi.
Il latte controllato viene sottoposto mensilmente ad analisi microbiologica e non deve superare i
seguenti valori massimi (M), secondo gli standard imposti, in questo caso, dalla regione
Lombardia:
• numero di germi aerobi (30°C) 30.000 UFC/ml;
• germi coliformi (30°C) 100 UFC/ml;
• Staphylococcus aureus 100 UFC/ml;
• Streptococcus agalactiae 0 UFC/ 0,1 ml;
• Salmonelle non riscontrabili in 25 ml;
• cellule somatiche 250.000/ml.
Anche le condizioni climatiche e la stagione, svolgono un ruolo significativo nei confronti della
contaminazione del latte, in quanto, temperature e umidità ambientali influiscono sulla
proliferazione microbica a livello della lettiera, degli animali e delle superfici degli impianti
(Quaderni di ricerca n 119-2010 Lombardia).
Il latte non deve costituire un pericolo per la salute dei consumatori, a causa di tossine o di
microrganismi patogeni (Yersinia enterocolitica) (Cantoni C. 2011). Il ritrovamento di elevate
cariche di batteri psicrotrofi nel latte può essere segno di temperature di raffreddamento inadeguate
e/o di tempi di conservazione troppo lunghi. La presenza di coliformi, è in genere, un indicatore di
43

contaminazione ambientale e fecale del latte per scarsa pulizia degli animali e scarsa igiene durante
le operazioni di mungitura (Reinemann et al 1997).
Prima della diffusione dell'impiego di dispositivi di mungitura meccanica, il latte crudo è stato
veicolo sia di malattie respiratorie (la difterite) sia di infezioni enteriche ( per esempio, la febbre
tifoide) (Jay J. M. 2009).
2.4.2 Pastorizzazione, Termizzazione e Tecnologia ESL
La pastorizzazione è un processo di risanamento termico applicato ad alcuni alimenti, allo scopo di
minimizzare i rischi per la salute, dovuti a microrganismi patogeni sensibili al calore, quali batteri
in forma vegetativa, funghi e lieviti, con un'alterazione minima delle caratteristiche chimiche,
fisiche ed organolettiche dell'alimento (Tortora G. J. et al 2008).
Si differenzia, quindi, dalla sterilizzazione, ottenuta con temperature (e tempi di applicazioni delle
stesse) notevolmente maggiori, che garantisce un marcato aumento dei tempi di conservazione al
prezzo di un'alterazione spesso pesante dei contenuti nutritivi e delle caratteristiche organolettiche
dell'alimento.
Si differenzia, inoltre, dalla termizzazione (più spesso usata solo per il latte e suoi derivati)
soprattutto per la durata del trattamento.
Con la pastorizzazione il numero di germi nel latte si riduce di due logaritmi e gli agenti patogeni
trasmissibili attraverso il latte vengono uccisi o inattivati. Tuttavia, numerosi microrganismi
termoresistenti possono sopravvivere alla pastorizzazione (Cantoni C. 2011). Il suo uso è limitato ad
alcuni alimenti in forma liquida (soprattutto latte,vino,birra e succhi di frutta), su cui il processo si
può compiere con particolare efficacia e con limitati effetti avversi.
44

La pastorizzazione del latte viene eseguita soltanto con l'impiego di calore (Jay J. M. 2009).
In anni più recenti a questa metodica si sono affiancati il trattamento HTST (High
Temperature/Short Time) e il trattamento UHT (Ultra High Temperature), per migliorare la
sicurezza dei prodotti alimentari e prolungarne la vita di scaffale. Sono entrambi in grado di
effettuare il risanamento (eliminazione dei batteri potenzialmente patogeni); propriamente, la
pastorizzazione è solo la HTST che utilizza temperature di 70-75°C per 15-20 secondi (Ailas C.
2000); la UHT invece è una sterilizzazione 140-145°C per pochi secondi
(http://www.chimica-online.it/download/calore/conservazione_calore.htm).
Questi processi permettono una più lunga conservazione del prodotto, riuscendo a mantenere quasi
inalterate le caratteristiche organolettiche e nutrizionali.
In Italia, la pastorizzazione del latte fu introdotta con il Regio Decreto 9 maggio 1929 n. 994.
Secondo quanto riportato dalla legge 3 maggio 1989 n. 169, il latte pastorizzato deve possedere le
seguenti caratteristiche:
• essere ottenuto mediante un trattamento che comporti un’elevata temperatura per un breve periodo
di tempo (almeno 71,7°C per 15 secondi o qualsiasi altra combinazione equivalente) o mediante un
trattamento di pastorizzazione che impieghi diverse combinazioni di tempo e temperatura
raggiungendo un effetto equivalente;
• presentare una reazione negativa alla prova della fosfatasi e positiva alla prova della perossidasi.
E' tuttavia autorizzata la fabbricazione di latte pastorizzato che presenti una reazione negativa della
prova di perossidasi a condizione che, sulle confezioni figuri una indicazione del tipo pastorizzato a
temperatura elevata;
• al trattamento termico deve seguire un rapido raffreddamento che porti il latte nel più breve tempo
possibile ad una temperatura non superiore ai 6 °C;
45

• deve avere un contenuto in sieroproteine solubili non denaturate non inferiore all’11% delle
proteine totali, e non inferiore al 14% delle proteine totali nel latte fresco pastorizzato, e non
inferiore al 15,5% delle proteine totali nel latte fresco pastorizzato di alta qualità.
Con il trattamento termico di pastorizzazione abbiamo:
• distruzione dei microrganismi patogeni non sporigeni;
• conservabilità a 4 °C per 6 giorni (fino a 45 giorni per il latte pastorizzato ad alta temperatura);
• denaturazione del 10-25% delle sieroproteine (fino a 50% per il latte ad alta temperatura);
• distruzione fino al 10% della vitamina C.
Viene definito"latte fresco pastorizzato", il latte che perviene crudo allo stabilimento di
confezionamento e che è sottoposto a un solo trattamento termico entro 48 ore dalla mungitura.
Viene definito "latte fresco pastorizzato di alta qualità", il latte fresco pastorizzato ottenuto da latte
crudo, proveniente direttamente dalle stalle, ovvero, da centri di raccolta cooperativi o consortili
avente le caratteristiche igieniche e di composizione stabilite con particolare riferimento al
contenuto di proteine, grasso, di carica batterica totale e di numero di cellule somatiche. Può essere
messo in commercio solo nel tipo intero e con un contenuto in materia grassa non inferiore al 3,5%.
La termizzazione, invece, non è che un semplice riscaldamento del latte a 57-68°C con un tempo
effettivo di riscaldamento di 30 secondi (Cantoni C. 2011), che ha lo scopo di eliminare una certa
percentuale di germi anticaseari, la cui presenza, causerebbe fermentazioni anomale e un
conseguente deprezzamento del prodotto caseario.
Questo trattamento, lascia al tempo stesso quasi inalterata la presenza di batteri utili alla
caseificazione, quali lattobacilli e streptococchi. La combinazione tempo/temperatura fa sì che le
caratteristiche organolettiche e qualitative del latte crudo di partenza, rimangano inalterate.
46

La tecnologia ESL (extended shelf life), permette di avere latte fresco conservabile per 12-21 giorni
e quindi, per un periodo sostanzialmente più lungo, rispetto al latte pastorizzato in modo
tradizionale ( conservabile per 5-6 giorni). Tale periodo di conservabilità, si ottiene grazie a una
serie di misure, finalizzate ad abbassare il contenuto di germi del latte, senza compromettere le
caratteristiche organolettiche.
Tra queste misure figurano:
• una buona qualità del latte crudo;
• un'efficace neutralizzazione della flora microbica con metodi quali; (a) combinazione di
centrifugazione e pastorizzazione; (b) riscaldamento elevato nel metodo diretto a 127°C per
due secondi e raffreddamento rapido a 3°C; oppure (c) microfiltrazione combinata a
pastorizzazione;
• nei metodi di microfiltrazione attualmente in fase di sviluppo, il latte viene scremato nel
separatore. Il latte scremato viene sottoposto a degerminazione mediante ultrafiltrazione
(grandezza dei pori della membrana pari circa a 1 micron), vi si aggiunge quindi la panna e
tutto il prodotto viene infine pastorizzato (72-74 °C per 15-.30 secondi);
• prevenzione di eventuali ricontaminazioni nel processo “downstream” povero di germi o
addirittura asettico (aggiunta del prodotto, serbatoi di stoccaggio e confezionamento);
• temperatura del prodotto in fase di produzione pari a un massimo di 6°C, del prodotto in
commercio pari a massimo di 8°C (Cantoni C. 2011).
47

2.4.3 I patogeni del latte
Poiché il latte è un eccellente fonte di nutrienti, e poiché gli animali possono ospitare microrganismi
responsabili di patologie nell'uomo, non sorprende che il latte crudo possa essere veicolo di alcune
malattie. Alcune sono tipiche patologie animali che possono colpire anche l'uomo (zoonosi),
attraverso il consumo di latte di vacca:
tubercolosi; febbre Q;
brucellosi; listeriosi;
salmonellosi; infezioni stafilococciche/ streptococciche (mastiti);
campilobatteriosi; coliti enteroemorragiche.
I principali patogeni che possono riguardare il latte sono:
Campylobacter. Sono batteri in grado di causare malattie acute e croniche nell'uomo e negli
animali, con sintomatologia tipica rappresentata da diarrea. Le due specie principalmente coinvolte
nei focolai di origine animale sono C. jejuni e C. fetus. Le fonti per l'uomo sono numerose, dal
contatto diretto con animali (sia quelli selvatici che di affezione), al consumo di alimenti (carne,
latte) contaminati, ma anche di acqua contaminata. Il bovino non è né l'ospite né la fonte primaria di
contaminazione, essendo questa rappresentata soprattutto dai volatili e dai suini. Tuttavia, la
presenza di Campylobacter negli allevamenti bovini non è infrequente e quindi, il potenziale di
rischio di contaminazione del latte è presente;
E. coli verocitossici. I ceppi di E.coli verocitossici, di cui E.coli O157:H7 è il principale
rappresentante, sono un gruppo di coliformi che ha causato focolai di malattia con vittime negli
48

USA in seguito a consumo di carne contaminata poco cotta. Sebbene questi ceppi siano stati
identificati in diversi ruminati domestici, il bovino sembra essere il serbatoio principale, mentre non
si conosce il ruolo di altri animali come maiali e animali d'affezione. Questi batteri non causano
forme enteriche nei bovini e per questo motivo gli animali portatori-eliminatori non vengono
facilmente identificati. La carne contaminata in sede di macellazione e poco cotta rappresenta la
fonte primaria d' infezione per l'uomo. Tuttavia, sono stati segnalati anche casi derivanti dal
consumo di latte contaminato;
Listeria monocytogenes. La listeriosi è una malattia che colpisce animali e uomo e può determinare
diverse forme cliniche anche gravi. Vi è una crescente evidenza che nell'uomo la maggior parte
delle forme abbia origine alimentare. Negli animali la fonte principale di contaminazione è
rappresentata da insilati e foraggi contaminati da terra o polvere, ma spesso la sitomatologia clinica
è assente; è infatti possibile isolare il microrganismo dalle feci di diversi animali domestici e
selvatici apparentemente sani;
Yersinia Enterocolitica. Da sottolineare che sono stati segnalati solo alcuni sierotipi (O: 3 e O: 9)
come causa di patologia nell'uomo, patologia caratterizzata soprattutto da forme enteriche. Gli
alimenti contaminati sono considerati il principale veicolo di trasmissione di malattia all'uomo, e tra
questi sono compresi il latte e i prodotti a base di latte;
Mycobacterium avium subsp paratuberculosi (MAP). Questo microrganismo causa la
paratubercolosi nel bovino, ed è stato recentemente oggetto di particolare attenzione, poiché vi sono
crescenti evidenze che possa anche essere implicato nel morbo di Crohn nell'uomo. Da sottolineare
tuttavia, che il ruolo zoonosico di M.avium non è stato ancora dimostrato. La parartubercolosi è
49

presente da tempo negli allevamenti ed il microrganismo può essere eliminato con il latte oltre che
con le feci, anche in assenza di segni clinici. I trattamenti termici ed in particolare la pastorizzazione
non sono in grado di inattivarlo completamente, quindi, vi è la possibilità per l'uomo di assumere
latte contaminato;
Staphylococcus aureus. E' un microrganismo molto diffuso in natura, essendo parte della flora
commensale dell'uomo e degli animali, dove si localizza sulla cute, sulle mucose e nel tratto
intestinale. Nella vacca da latte determina infezioni mammarie persistenti, nell'uomo invece,
provoca intossicazioni, conseguenti all'ingestione delle tossine prodotte dal batterio nell'alimento.
La pastorizzazione è in grado di inattivare la maggior parte dei ceppi ma non la tossina
termostabile. I cibi maggiormente implicati nelle tossinfezioni da tossine stafilococciche sono il
latte e i prodotti a base di latte crudo ( Quaderni di ricerca n 119-2010Lombardia );
Salmonella. La salmonellosi è un’importante malattia negli animali da reddito, per la gravità dei
sintomi e per i risvolti di natura sanitaria conseguenti alla trasmissione all’uomo. Nel bovino la
salmonellosi si manifesta clinicamente come sindrome setticemica, enterica, o abortigena. Possono
essere colpiti animali di tutte le età, anche se la malattia si manifesta più frequentemente nei vitelli.
La diagnosi di laboratorio può essere effettuata mediante isolamento in coltura, PCR, o titolazione
anticorpale. Il rischio principale per l’introduzione di Salmonella in allevamento, è l’ingresso di
soggetti eliminatori, pertanto è opportuno prevedere per gli animali introdotti una quarantena di
almeno 3 settimane, ed un controllo microbiologico sulle feci. Un altro importante fattore di rischio
è la contaminazione ambientale ad opera di animali sinantropi. La misura di prevenzione da
applicare in questo caso è ridurre al minimo la possibilità di contatto fra tutte queste specie e gli
animali allevati, evitando in particolar modo la contaminazione fecale di alimenti e mangiatoie
50

(Barberio A., et al 2009).
2.4.4 Fermentazione prodotti lattiero-caseari e batteri lattici
Il metabolismo microbico è responsabile di molti processi di trasformazione. I prodotti che ne
derivano, possono essere utili o dannosi per l’uomo. Nel primo caso l’uomo ha messo a punto
tecniche per promuovere o guidare questi processi, nel secondo caso l’obiettivo è stato di prevenire
o arrestare le trasformazioni indesiderate. Molti dei cibi che consumiamo sono il risultato di una
trasformazione microbica o possono subirla loro malgrado
(www.mednat.org/alimentazione/fermentazione_cibi_e_bevande.pdf).
La produzione e le caratteristiche, di molti prodotti alimentari, sono dovute alle attività fermentative
dei microrganismi. Formaggi stagionati, sottaceti, e salumi sono tutti alimenti caratterizzati da una
shelf-life più lunga rispetto a quella delle materie prime da cui originano. Oltre ad essere più stabili
dal punto di vista della conservazione, tutti i prodotti fermentati sono caratterizzati da aroma e
sapore propri, che sono il risultato, diretto o indiretto, dell'attività dei microrganismi che attuano la
fermentazione. Il processo fermentativo è in grado di ridurre la tossicità di alcuni alimenti, oppure
può renderne altri tossici.
Dal punto di vista biochimico, la fermentazione è il processo metabolico durante il quale i
carboidrati e i composti ad essi correlati vengono parzialmente ossidati, con conseguente rilascio di
energia in assenza di qualsiasi accettore esterno di elettroni. Sono i composti organici derivanti
direttamente dalla scissione dei carboidrati a fungere da accettori finali degli elettroni, e di
conseguenza, l'ossidazione incompleta dei composti di partenza determina il rilascio solo di una
51

piccola quantità di energia durante tale processo. I prodotti della fermentazione sono rappresentati
da composti organici più ridotti di altri (Jay J. M. 2009).
Il processo di fermentazione è attuato da quelli che collettivamente vengono chiamati “fermenti
lattici”, che intervengono nella trasformazione di materie prime naturali (latte, carni, verdure,
insilati ecc.) in prodotti di notevole valore nutrizionale, quali formaggi o insaccati.
Molti alimenti sono prodotti tutt'oggi sfruttando in maniera non consapevole la crescita di
microrganismi utili naturalmente presenti nelle materie prime o nell'ambiente di produzione. Questi
microrganismi, anche se possono essere inizialmente presenti in basso numero, prendono il
sopravvento sugli agenti di deterioramento e sui patogeni perché, la formulazione dell'alimento o le
condizioni di produzione o, ancora, la stessa crescita dei microrganismi utili, rende l'ambiente
favorevole alla loro crescita a scapito di microrganismi dannosi (Notiziario ERSA 2/2006).
Il gruppo dei batteri lattici comprende microrganismi molto diversi dal punto di vista morfologico
(microrganismi bastoncellari e cocchi) e tassonomico. In qualità di anaerobi obbligati, nel processo
di fermentazione producono acido lattico, che può essere l'unico prodotto di fermentazione
(omofermentazione), oppure, oltre all'acido lattico, si possono formare anche anidride carbonica,
etanolo e/o acido acetico (eterofermentazione). Comunque, il numero di lattobacilli può aumentare
anaerobicamente anche in presenza di ossigeno dell'aria ( anerobi aerotolleranti). I batteri lattici
sono gram positivi, catalasi negativi, prevalentemente immobili, e ad eccezione del genere
Sporalactobacillus, non formano spore. Al gruppo dei batteri lattici attualmente sono ascrivibili
moltissimi generi, tra cui le specie e i ceppi maggiormente utilizzati nelle fermentazioni alimentari
e/o come microrganismi probiotici, appartengono ai generi Lactobacillus, Lactococcus,
Streptococcus, Leuconostoc, Weissella, Oenococcus, Pediococcus, Carnobacterium ed
Enterococcus (www.mednat.org/alimentazione/fermentazione_cibi_e_bevande.pdf).
52

L' habitat naturale dei batteri lattici è costituito dai vegetali intatti e da quelli autodecomposti, come
pure dall'intestino e dalle mucose dell'uomo e dell'animale.
Tali batteri si sono talmente adattati a questi habitat ricchi di sostanze nutritive e di accrescimento,
che non sono più in grado di autosintetizzare alcuni fattori di crescita come, ad esempio, gli
aminoacidi e le vitamine, che trovano disponibili negli alimenti.
Possono crescere in un range di temperatura compreso tra 5°C e 53°C con valori ottimali di
30-40°C , sono acidurici, con un pH ottimale di crescita di 5,5-5,8 potendo crescere anche a pH<5.
In base alla presenza o all’assenza degli enzimi responsabili del tipo di omo o etero-fermentazione
degli zuccheri, rispettivamente gli enzimi fruttosio-1,6-difosfato aldolasi e la fosfochetolasi, le
specie appartenenti al genere Lactobacillus sono divise in 3 gruppi fisiologici:
Gruppo I: Lattobacilli omofermentanti;
Gruppo II: Lattobacilli eterofermentanti facoltativi;
GruppoIII: Lattobacilli eterofermentanti obbligati.
(www.mednat.org/alimentazione/fermentazione_cibi_e_bevande.pdf).
Alcuni microrganismi responsabili degli aromi tipici sono denominati secondari perché la loro
azione si esplica dopo che la fermentazione lattica è avviata. Tra questi ci sono i batteri propionici
ed i micrococchi, ma anche muffe e lieviti. I batteri propionici sono i responsabili della tipica
occhiatura, nonché del gusto del formaggio tipo Emmental. La produzione di gas, di acido acetico e
propionico è la principale azione di questi batteri, ma molto importante è anche la loro capacità di
formare acidi grassi liberi e volatili, che conferiscono la tipica e intensa aroma al prodotto. Possono
inoltre agire sulla caseina destrutturata dai batteri lattici, modificando ulteriormente il substrato.
I micrococchi sono microrganismi capaci di tollerare concentrazioni elevate di cloruro di sodio, da
7,5% a 15%, aerobi stretti o anaerobi facoltativi, poco tolleranti i bassi pH, e hanno un ruolo
importante nella maturazione dei formaggi a maturazione superficiale. Svolgono le seguenti
53

funzioni:
• prevengono l'irrancidimento dei grassi decomponendo l'acqua ossigenata prodotta da altri
microrganismi e creando un ambiente riducente per riduzione dei nitrati;
• hanno una maggiore attività lipolitica e proteolitica, rispetto ai fermenti lattici, e sono in
grado di metabolizzare aminoacidi e acidi grassi producendo composti importanti per il
sapore e l'aroma;
• alcuni ceppi sono in grado di produrre sostanze inibitorie per microrganismi patogeni;
• alcune specie sono pigmentate e contribuiscono alla determinazione dell'aspetto della
superficie di formaggi a maturazione superficiale.
Alcuni batteri lattici interessanti per il settore lattiero-caseario, sono pure in grado di produrre delle
sostanze, le batteriocine, attive contro batteri patogeni, come Listeria monocytogenes, che possono
essere veicolati dagli alimenti. Le batteriocine vengono prodotte industrialmente, mediante processi
di fermentazione. Altri ceppi sono produttori di diverse sostanze proteiche, le nisine, con attività
antibatterica; vengono aggiunti, infatti, ai formaggi fusi, come i formaggini, per la loro capacità di
prevenire o bloccare l’attività alterativa di batteri sporigeni eventualmente presenti. Sono tra i pochi
antibiotici che per legge possono essere addizionati agli alimenti
(www.mednat.org/alimentazione/fermentazione_cibi_e_bevande.pdf.).
Tuttavia, affidarsi alla contaminazione naturale delle materie prime con microrganismi utili diventa
sempre più difficile soprattutto nei casi in cui la microflora necessaria può essere assente in latti resi
paucimicrobici o dismicrobici a seguito di pratiche di produzione o dai trattamenti termici. I
microrganismi potrebbero non essere presenti in numero sufficiente da prendere rapidamente il
sopravvento sui microrganismi dannosi, o svilupparsi in ritardo, rallentando il processo di
54

produzione. La comprensione del ruolo dei microrganismi nelle fermentazioni ha portato alla
nascita dell'industria delle “colture starter”. L'uso di starter, specialmente se isolati e selezionati
dalla microflora caratteristica dei formaggi, può rappresentare un notevole interesse per accelerarne
e indirizzarne la maturazione: ciò può addirittura diventare essenziale in condizioni come quelle
odierne. Gli, starter, quindi, danno inizio alla fermentazione e la indirizzano correttamente,
apportando microrganismi utili in quantità sufficienti allo scopo. Con l'aggiunta dell'innesto al latte
in caldaia si apportano acidità, micronutrienti, fattori di crescita, enzimi e soprattutto batteri lattici.
L'innesto può sopperire alla scarsa attività del latte, favorisce l'attività degli enzimi coagulanti,
riduce i tempi di coagulazione, quindi migliora la struttura del coagulo, accelera la sineresi e lo
spurgo dei granuli di cagliata e soprattutto apporta batteri lattici vivi (Notiziario ERSA 2/2006).
2.4.5 Presenza di Yersinia spp nel latte e prodotti a base di latte
Nonostante sia stato provato che la maggior parte dei ceppi di Yersinia enterocolitica isolati nei
prodotti lattiero-caseari non sono patogeni (Walker et al. 1986), laddove si è riscontrata la presenza
di quelli che risultano esserlo, lo rendono comunque un potenziale problema di salute pubblica per
tale settore, poiché responsabile di diversi focolai di malattia per ingestione di prodotti contaminati
(Tacket et al 1984). Inoltre, dal confronto dei risultati raccolti da diversi studi sull'isolamento di
Yersinia enterocolitica nel latte crudo in diversi paesi (Schiemann et al 1978; Hughes 1979;
Rohbach et al 1992), appare evidente una considerevole variabilità nei valori di prevalenza del
microrganismo (Larkin et al 1991).
E' possibile però constatare, da alcuni dati ufficiali risalenti al 2007, un miglioramento della qualità
igienico-sanitaria del latte prodotto nei diversi stati membri dell' Unione Europea; in particolare
nessun campione ha mostrato positività alla contaminazione da Yersinia enterocolitica, rilevato
55

invece nel 1,5% dei campioni dell'anno precedente (EFSA 2009).
L'interesse nel valutare la presenza, e quindi, la frequenza con cui i ceppi patogeni del genere in
questione sono ritrovati nel latte crudo, nasce dal fatto che nell'ultimo decennio, il consumo di
questo prodotto si è diffuso molto nel territorio nazionale. Il costo contenuto, la riduzione
dell'impatto ambientale conseguente alla semplificazione logistica e di imballaggio, il desiderio di
naturalità e il ritorno alle tradizioni, sono alcuni dei fattori che hanno contribuito a consolidare una
realtà produttiva che fino a qualche anno fa era assente nel nostro paese.
Secondo quanto previsto nella Delibera n° 381 del 28 Maggio 2007 della regione Toscana, le
aziende, attualmente abilitate alla vendita diretta di prodotto sono autorizzate dal Servizio Sanitario
Pubblico competente, hanno allevamenti controllati e devono rispettare corrette norme di
autocontrollo con analisi periodiche, per la costante verifica dei requisiti qualitativi e
igienico-sanitari, che per questa tipologia di prodotto sono più restrittivi di quelli previsti per il latte
destinato a essere trattato termicamente.
Sul piano nazionale, i criteri igienici di processo riconducibili alla condizione sanitaria degli animali
e all'igiene delle fasi di mungitura sono stabiliti dall'Intesa Stato-Regioni e province Autonome di
Trento e Bolzano in materia di vendita diretta di latte crudo per l'alimentazione umana” 5/CSR del
25 Gennaio 2007.
In ambito europeo si fa riferimento al REGOLAMENTO CE n° 853/2004, volto ad assicurare un
livello elevato di tutela dei consumatori per quanto attiene la sicurezza dei prodotti volta a garantire
il corretto funzionamento del mercato interno dei prodotti di origine animale, contribuendo al
conseguimento degli obiettivi della politica agricola comune. E' pertanto necessario da una parte
non sottovalutare il potenziale pericolo di contaminazioni microbiche che comporterebbero
conseguenze negative sia per la tutela del consumatore, che per gli allevatori, dall'altra un puntuale
monitoraggio dell'epidemiologia dei patogeni potenzialmente presenti negli allevamenti al fine di
56

mettere in atto tutte le misure precauzionali del caso senza creare allarmismo ingiustificato.
Altra importante considerazione è che non tutti i latti sono uguali, ogni singolo prodotto acquistato
presso i distributori rappresenta e racchiude le caratteristiche del singolo allevamento da cui
proviene e queste, soprattutto laddove le realtà produttive sono maggiormente diversificate,
comportano realmente una pluralità di tipologie di prodotto che difficilmente è percepibile quando
si mescola il latte di più aziende (Lodi et al 2009).
Per ogni unità campionaria sono stati messi in atto metodi standardizzati di isolamento e
d'identificazione dei principali cinque patogeni per il latte crudo:
Stafilococchi coagulasi positivi ( ISO 6888- 1:1999);
Campylobacter spp (UNI EN ISO 10272-1:2006);
Salmonella (ISO 6579 : 2002 /cor 1:2004);
Listeria monocytogenes (UNI EN ISO 11290-1:2005);
E.coli O:157 (ISO 16654 : 2001).
In base alla Ricerca corrente del 2009, già da diversi anni vengono effettuate prove per ricercare:
Carica mesofila totale (NORMA AFNOR 3M 01/1-09/89);
E.coli e coliformi totali (NORMA AOAC 991-14 2002).
A tali analisi ufficiali è stata affiancata, dal 2010, l'analisi quali-quantitativa addizionale per la
ricerca di ceppi patogeni di Yersinia enterocolitica.
Considerando che i punti critici per la contaminazione del latte si concentrano nel momento della
mungitura, si ritiene opportuno e necessario un controllo, da parte dell'allevatore, dell'igiene delle
operazioni di mungitura. Infatti, se la presenza nel latte di batteri responsabili di infezioni
mammarie, quali S. aureus e MAP, è controllabile mediante opportuni piani di controllo, la
contaminazione fecale del latte si controlla solo mediante una buona routine di mungitura.
L'ingestione diretta di latte crudo inquinato da Yersinia può essere causa reale di trasmissione
57

dell'infezione dall'animale all'uomo, poiché il sierotipo O:3 è il responsabile più comune di
gastroenteriti. Le caratteristiche del germe (resistenza e moltiplicazione a basse temperature) fanno
sì, che attraverso le attrezzature meccaniche per la mungitura non sufficientemente pulite, attraverso
le vasche refrigerate di raccolta del latte e soprattutto, a causa di procedimenti di pastorizzazione
imperfetti, la contaminazione di Yersinia enterocolitica rappresenti un reale pericolo per il
consumatore (Aiello P. et al 1984).
2.5 Prodotti a base di carne
L'animale da macello rappresenta, nella catena di produzione degli alimenti, il maggior serbatoio di
microrganismi, soprattutto di specie patogene o patogene facoltative. Queste presenze indesiderate
esercitano un'intensa azione infettante sulle carcasse degli animali macellati, che possono servire
come possibile veicolo del germe e quindi, contaminare i prodotti finiti. Ne deriva un potenziale
rischio sanitario per il personale addetto alla lavorazione, ma soprattutto per il consumatore finale.
Le innovazioni tecnologiche, associate a norme igieniche adeguate, si sono fin ora concentrate
principalmente nei confronti di agenti batterici comunemente conosciuti come: Salmonelle, E.coli,
Stafilococchi, Streptococchi, Pseudomonas, Shighelle, ecc. Recentemente l'attenzione si è
focalizzata su alcuni microrganismi, considerati emergenti, sempre più frequentemente isolati
negli alimenti di origine animale, come Listeria monocytogenes, Campylobacter jejuni, Aeromonas
hydrophyla e Yersinia enterocolitica.
Attualmente grazie ad un sempre maggior utilizzo della catena del freddo nell'industria alimentare,
ed al progresso delle tecniche di isolamento e di identificazione di batteri, supportate da metodi di
biologia molecolare, lo spettro di nocività gastroenterica sembra essersi ampliato. Infatti, l'utilizzo
di tecniche di PCR o d'ibridazione del DNA, ha notevolmente migliorato l'efficienza di isolamento
58

del germe, a partire da campioni di carne suina. Inoltre, se si considera che numerose tossinfezioni
derivano più da errori di manipolazione durante e dopo la sua preparazione, che dall'alimento in sé,
si può comprendere il ruolo di Yersinia enterocolitica come agente a rischio sanitario ( Allegra
A.M., et al 2003).
Le complesse misure di prevenzione attualmente adottate nell’ambito della produzione delle carni,
comprendono strumenti quali la selezione di genotipi resistenti, il bando delle farine animali, la
definizione delle categorie di età dei bovini destinati a macellazione, l’adozione di procedure per
l’asportazione ed eliminazione di materiali che comportano un rischio specifico.
Anche per il controllo dei pericoli microbiologici, la produzione primaria riveste un ruolo di
fondamentale importanza ed è capace di influenzare in maniera determinante le caratteristiche
microbiologiche del prodotto (FSA, 2004). Nel settore delle carni fresche, nelle successive fasi di
macellazione, sezionamento e confezionamento, le procedure e tecnologie adottate hanno la
prevalente finalità di contenere ulteriori contaminazioni e limitare lo sviluppo dei microrganismi.
Alcuni punti di controllo critico nella fase di macellazione, quali ad esempio, eviscerazione e
scuoiamento, hanno notevole importanza nel minimizzare il rischio derivante dalle contaminazioni
e dallo sviluppo di microrganismi patogeni di rilevante impatto epidemiologico (Edward, Fung
2006). Sono limitate o poco applicate le possibilità di impiego di metodi di risanamento (trattamenti
termici o alternativi).
I criteri microbiologici definiti dalla Legislazione Comunitaria attribuiscono notevole importanza ai
microrganismi patogeni veicolati dalle carni. Sono numerosi i criteri di sicurezza e di igiene che
hanno quale matrice alimentare di riferimento le carni. I criteri microbiologici applicabili per la
valutazione delle carcasse degli animali macellati, hanno per finalità la verifica dei risultati ottenuti
da parte dell’OSA con l’attuazione delle procedure igieniche, inclusa la valutazione del trend del
controllo di processo. Soprattutto negli USA sono state sviluppate iniziative e misure anche cogenti,
59

prioritariamente finalizzate al controllo dei patogeni nella macellazione, destinate a rafforzare le
SOPs (Standard Operating Procedures) e rendere proporzionale il monitoraggio microbiologico
rispetto all’efficacia delle misure di controllo igienico. Alcuni dei più importanti agenti batterici
responsabili di malattie ad elevata incidenza trasmesse da alimenti (Campylobacter spp.,
Salmonella spp, E. coli enteropatogeni), sono veicolati attraverso le carni, soprattutto i c.d.
“patogeni enterici” (Huffman, 2002). Questi germi risultano direttamente coinvolti nelle
contaminazioni durante le fasi di macellazione, potendo essere trasferiti dall’intestino alla cute e da
questi, alle carcasse durante le operazioni di eviscerazione e scuoiamento (Nastasijevic et al., 2008;
McEvoy et al., 2000a).
La carne riveste un’importanza fondamentale dal punto di vista nutrizionale e dietetico, ma nello
stesso tempo può costituire un fattore di rischio per la salute pubblica. Le contaminazioni a cui può
andare incontro la carne sono numerose e possono derivare da contaminazioni che si verificano sia
durante la vita dell’animale (contaminazioni endogene), sia durante le varie fasi del processo
produttivo (contaminazione esogene).
Le contaminazioni endogene possono derivare da sostanze somministrate volontariamente
all’animale (farmaci e anabolizzanti) o da sostanze che siano state accidentalmente assunte
dall’animale attraverso alimenti o acqua (pesticidi, metalli pesanti, micotossine, PCB, etc.). Tra le
contaminazioni endogene, un ruolo importante viene assunto dai cosiddetti “pericoli biologici”
responsabili di zoonosi, quali parassiti (Toxoplasma gondii, Trichinella spp, Cysticercus bovis e
Cysticercus cellulosae), batteri (brucelle, micobatteri tubercolari, Burkholderia mallei, Bacillus
antracis etc.) e agenti non convenzionali, come le proteine prioniche delle encefalopatie
spongiformi trasmissibili del bovino e degli ovini (TSE) (Colavita, 2008).
Le contaminazioni endogene delle carni sono essenzialmente riconducibili a batteri responsabili di
tossinfezioni alimentari quali, Salmonella, Campylobacter, Escherichia coli, Yersinia e Aeromonas,
60

Gram-positivi sporigeni quali Clostridium e Bacillus e infine Gram-positivi asporigeni appartenenti
ai generi Listeria e Staphylococcus (Colavita, 2008). Questi microrganismi, si trovano
prevalentemente nel tratto digerente e sulla cute degli animali e possiedono elevata capacità di
sopravvivenza nell’ambiente esterno. I soggetti portatori, che non presentano dunque
sintomatologia clinica, sono i principali “disseminatori” di questi microrganismi nell’ambiente,
ragion per cui alcune fasi della macellazione quali lo scuoiamento, la depilazione, e l’eviscerazione,
rappresentano i momenti in cui possono avvenire le maggiori contaminazioni delle carcasse,
dell’ambiente e delle attrezzature. La qualità igienica della carne dipenderà, dunque, da una parte
dalle contaminazioni derivate dal processo di macellazione e sezionamento, così come dalla
capacità di crescita dei microrganismi durante le fasi di raffreddamento, stoccaggio e distribuzione.
Diversi microrganismi potenzialmente pericolosi possono ritrovarsi nell’intestino, nelle deiezioni e
sulla pelle degli animali sani. Questi microrganismi, quali per esempio, E. coli patogeni,
particolarmente E.coli O157:H7, Salmonella e Campylobacter, sono capaci di provocare delle
infezioni alimentari nell’uomo (Edwards and Fung, 2006).
I microrganismi sono introdotti nella filiera delle carni attraverso gli stessi animali che li veicolano
a livello del tubo digerente e della pelle (Rosset, 1982; Rosset et Liger, 1982; Fournaud, 1978.), che
costituiscono la principale fonte di contaminazione delle carcasse al momento della macellazione
(McEvoy et al., 2000a; Cartier, 1997; Fournaud, 1978).
Gli animali spesso, sono portatori del microrganismo senza presentare sintomi di malattia. Il rilascio
di questi germi nell’ambiente in alcuni casi può essere massivo, costituendo un potenziale enorme
di contaminazione delle superfici corporee di animali a contatto. Ormai da anni l’USDA sostiene
una politica di tolleranza zero nei confronti delle contaminazioni fecali sulle carcasse. Procedure di
sanitizzazione e metodi di intervento vengono utilizzati durante le operazioni di macellazione al
fine di prevenire o eliminare le contaminazioni fecali, le cross-contaminazioni e ridurre la carica
61

batterica (USDA Library, 2002).
Al momento della macellazione la contaminazione delle carcasse è inevitabile, poiché la carne,
inizialmente sterile, entra in contatto con le contaminazioni ambientali (aria, acqua, materiali,
uomo, etc.). Il numero e la specie dei batteri con cui si entra a contatto, in questo stadio sono
determinanti, in quanto, definiscono la qualità igienica dei prodotti che, saranno ottenuti a partire da
queste carni.
Dopo il 1996, la messa in opera dei piani HACCP ha prodotto un’applicazione più regolare delle
misure igieniche di macellazione, ma non può consentire di eliminare completamente il rischio di
contaminazione, in quanto, le superfici preventivamente “sterili” delle carcasse vengono a contatto
con l’ambiente esterno, spesso ricco di possibili fonti di contaminazione.
La superficie della carcassa è essenzialmente sterile (Gill et al., 1976,1978), ma durante il processo
di macellazione le contaminazioni sono inevitabili (Elder et al., 2000; Belk 2001). Durante la
macellazione possono essere trasferiti sulla superficie della carcassa anche 3 log UFC/cm2 (Arthur
et al., 2005). Tali operazioni sono inoltre capaci di trasferire sulla carne eventuali microrganismi
patogeni di cui sono portatori gli animali (McEvoy et al. 2003b).
La scuoiatura della carcassa e la sua eviscerazione sono considerate le fasi che determinano un
elevato rischio di contaminazione. Le contaminazioni possono originare per contatto diretto o
indirettamente da pareti, pavimenti, aria, personale, indumenti, impianti, attrezzature e strumenti
impiegati, quali i coltelli utilizzati per le operazioni di scuoiamento e sezionamento (Fung et al.,
2001). Non sono altresì trascurabili le contaminazioni crociate, che possono verificarsi dal contatto
diretto tra carcasse (Elder et al., 2000). Questo passaggio di microrganismi, tra una carcassa e quelle
contigue, è notevolmente influenzato dalla struttura dell’impianto, dalla velocità della catena di
macellazione e soprattutto dalle attività degli operatori (Belk, 2001).
Il grado di contaminazione della cute degli animali macellati e la localizzazione dello sporco,
62

influenzano in maniera prevedibile la qualità delle contaminazioni, alla fine del processo di
macellazione (Sofos et al., 1999).
2.5.1 Yersinia spp nei prodotti a base di carne
Il mondo animale è un serbatoio importante per ceppi di Yersinia spp. Diversi animali portano
sierotipi unici del batterio, che sono coinvolti nelle infezioni umane. Numerosi studi sono stati
effettuati per isolare ceppi di Yersinia da diversi animali. Tuttavia, la maggior parte dei ceppi isolati
da fonte animale, differiscono sia biochimicamente e sierologicamente da ceppi isolati da individui
umani affetti da yersiniosi. I suini sono stati implicati come un importante serbatoio di sierotipi di
Yersinia, coinvolti nelle infezioni umane. L'incidenza di Yersinia nei maiali varia, oltre che da
paese a paese, anche all'interno della stessa nazione.
I ceppi di Yersinia enterocolitica che appartengono ai sierotipi associati con le malattie umane sono
stati isolati frequentemente da tonsille, lingue e materiale fecale di maiali da macello. Il tasso di
isolamento del microganismo dalle tonsille e lingue dei maiali è generalmente maggiore del tasso
presente nelle feci. In diversi paesi, Yersinia enterocolitica appartenente al biosierotipo 4/O: 3, ha
dimostrato di essere quello predominante in suini asintomatici. L'alta prevalenza dei ceppi O: 3 nei
suini, dimostra che probabilmente esiste un certo livello di specificità per questo ospite animale
(Schaake J., et al 2014). Tale sierotipo è stato isolato esclusivamente anche in diversi paesi europei,
come Danimarca, Belgio, Finlandia, Germania, Svezia e Svizzera. Una prevalenza minore è stata
riscontrata in Italia, in Grecia e in Polonia. Alcuni ricercatori hanno concluso che il ceppo O: 3 è un
normale abitante della cavità orale dei suini ed è coinvolto nelle infezioni umane. Quindi, i risultati
suggeriscono che i suini sono un serbatoio di infezione per il ceppo O: 3 e O: 5.
Inoltre, studi su modelli animali hanno dimostrato che i biosierotipi O: 3 e O: 8, hanno la capacità di
63

colonizzare e persistere per lunghi periodi a livello del tratto intestinale degli animali. Il corso dell'
infezione del biosierotipo O: 3 è asintomatica, anche se l'invasione di sezioni di tessuto intestinale e
diffusione di batteri sono stati osservati durante le prime 3 settimane dopo la somministrazione
orale. Al contrario, solo le tonsille erano efficientemente colonizzate da O: 8 in alcune delle cavie
infettate.
I maiali più giovani contraggono l'infezione dopo le prime 3 settimane in aree contaminate, o
tramite altri animali infettati e rimangono dei portatori faringei asintomatici del batterio per lungo
tempo; un grande numero di batteri viene eliminato nelle successive 2-3 settimane dopo l'infezione
(Schaake J. Et al 2014).
Le carcasse di suini sono spesso tenute in celle frigorifere per 2/4 giorni dopo la macellazione
prima del taglio. I prodotti preconfezionati a base di carne, possono rimanere in celle di
raffreddamento per la vendita al dettaglio per più una settimana, questo dipende dal prodotto, dall'
64
Fig. 6: Fonti di contaminazione di Yersinia enterocolitica per l'uomo (Allegra A.M.et al, 2003).

imballaggio, e dal tasso di turnover dei prodotti. I ceppi patogeni di Yersinia possono propagarsi
considerevolmente nel corso di questo periodo di conservazione, relativamente lungo.
In quanto organismo facoltativo, la crescita è drasticamente influenzata da un'atmosfera gassosa. In
condizioni anaerobiche non è in grado di crescere nel settore delle carni a pH 5.4-5.8, mentre la
crescita avviene a pH 6.0.
La presenza di anidride carbonica inibisce la crescita del microrganismo.
I suini da macello sono il principale serbatoio e la contaminazione delle carcasse avviene durante un
processo di macellazione igienicamente subottimale. Uno studio, ha utilizzato la sierologia per
determinare lo status infettivo di Yersinia nei suini prima della macellazione.
Sulla base dei risultati ottenuti al macello, il 66% dei suini aveva avuto contatto con Yersinia spp
enteropatogena in allevamento e la maggior parte degli allevamenti ospitava Yersinia spp (
Vanantwerpen G., et al 2014).
La prevalenza di ceppi patogeni di Yersinia enterocolitica è determinata con l' utilizzo di tecniche di
biologia molecolare tramite PCR. I prodotti a base di carne suina sono stati ampiamente analizzati a
causa dell'associazione tra Yersinia enterocolitica e i maiali. La fonte primaria di Yersinia patogena
sono i suini da ingrasso. La stretta relazione genetica tra i ceppi isolati nei suini e quelli isolati a
livello umano, è stata ampiamente dimostrata tramite parecchi metodi basati sul DNA.
Le Tonsille di suini infettati dal microrganismo possono facilmente contaminare la carcassa, le
frattaglie e l'ambiente durante i processi di macellazione. Tramite metodi di biologia molecolare, è
stato provato che la carne e frattaglie di suino contaminate da Yersinia sono un importante veicolo
di trasmissione per i ceppi patogeni (Bari Md. L., et al 2011).
In generale, le tonsille sono più produttive per il recupero di ceppi di Yersinia rispetto a lingue e
campioni di tessuto, permettendo tassi di isolamento più elevati rispetto all'esecuzione di tamponi.
La proporzione di ceppi patogeni di Yersinia nelle tonsille rispetto alle feci è di 6:1( Nesbakken et al
65

2003).
Tuttavia, quando il campionamento è effettuato sulla linea di macellazione, esiste la possibilità che
la presenza di Yersinia nelle tonsille sia il risultato della contaminazione dovuta ad un altro animale
durante il trasporto, in stabulazione o anche sulla linea di macellazione (EFSA 2009).
L' infezione è spesso acquisita tramite ingestione di cibo contaminato, in particolare carne di maiale
cruda o poco cotta (EFSA 2009) o tramite il contatto diretto con le carcasse. I suini non manifestano
sintomi clinici, sono dei portatori sani del microrganismo che si trova nella cavità orale, nelle
tonsille e lingua ma anche nei linfonodi, ed eliminano il batterio tramite le feci. Le carcasse possono
essere contaminate con il patogeno nel macello tramite la contaminazione con tonsille o contenuto
intestinale (Fondrevez M. et al 2014).
Questo aumenta la possibilità che Yersinia possa accedere alla catena alimentare, moltiplicandosi
nei prodotti alimentari grazie alla sua capacità di sopravvivere alle basse temperature ( Fredriksoon
Ahomaa et al 2000).
La ragione di questa elevata prevalenza può essere la cross-contaminazione durante i processi di
macellazione. Le tecniche di macellazione e l' igiene di tali processi possono influenzare il tasso di
contaminazione (Andersen 1988).
Inoltre Yersinia enterocolitica è stata isolata anche in campioni di pollame e in campioni di pesce
( Shanmugapriya S., et al 2014).
66

2.6 Scopo della Tesi
Nel presente lavoro di tesi si è voluto esaminare la prevalenza di Yersinia enterocolitica, sia in
alimenti crudi a base di carne, sia pronti al consumo alimentare, sia in campioni di latte crudo,
principalmente di derivazione bovina, che in prodotti lattiero-caseari a base di latte crudo,
provenienti da aziende distribuite sul territorio della Toscana, di competenza dell'Istituto
Zooprofilattico Sperimentale delle regioni Lazio e Toscana sezione di Pisa.
Successivamente, si è analizzato il profilo genetico dei ceppi isolati, sia in fase di sperimentazione,
che precedentemente individuati dal personale del laboratorio alimenti dell'istituto, per valutarne il
grado di patogenicità, analizzando specifici marker quali il gene AIL, YST A e YSTB tramite PCR.
Valutando, inoltre, la loro variabilità genetica, relativamente ai geni YST A, YST B e AIL, che
contraddistingue i diversi ceppi di Yersinia enterocolitica. Alcuni campioni sono stati conservati in
congelatore, prima di procedere all'analisi di laboratorio, altri invece, sono stati analizzati subito,
senza subire processi di congelamento e scongelamento.
L'attenzione verso il genere Yersinia spp nasce dal fatto che negli ultimi decenni, si è registrato un
aumento dei casi di yersiniosi in diversi paesi europei, collocando la malattia al terzo posto tra le
zoonosi maggiormente notificate. La situazione italiana presenta lacune, sia dal punto di vista della
notifica obbligatoria dei casi riscontrati nel paese, sia dal punto di vista della ricerca in laboratorio
del batterio. Infatti, il genere Yersinia non rientra nei criteri microbiologici applicabili ai prodotti
alimentari, in base ai criteri dettati dal Reg. CE N° 2073/2005, e in nessun piano nazionale o
sovranazionale. In letteratura, inoltre, scarseggiano i lavori dove il batterio è stato quantificato nelle
diverse derrate alimentari.
Lo studio è stato fatto anche al fine di fornire un risultato sia dal punto di vista igienico-sanitario,
sia dal punto di vista alimentare per il consumatore. Inoltre, vista le scarse informazioni presenti in
letteratura sulla realtà italiana, ci riproponiamo tramite questo lavoro di tesi, di fare chiarezza su una
67

realtà ancora non del tutto bene compresa sul territorio Toscano, di competenza dell' Istituto
Zooprofilattico Sperimentale delle regioni Lazio e Toscana, sezione di Pisa.
68

3 Materiali e metodi
In questo lavoro di tesi sono stati analizzati 186 campioni, suddivisi in base alla loro tipologia,
come riportato in tabella :
Tutti i campioni (lattiero-caseari) sono a base di latte crudo, non hanno, quindi, subito nessun
processo di termizzazione o pastorizzazione, tecniche che andrebbero a distruggere le cellule
batteriche di Yersinia enterocolitica all'interno dell'alimento, come già illustrato in precedenza. I
campioni di latte crudo bovino/ovino, sono stati prelevati da distributore automatico di latte o da
allevamento, e sono stati conservati a una temperatura di 3°C± 2 in refrigeratore prima di procedere
con le pratiche di analisi. I prodotti a base di carne, insaccati, carne macinata, carne di pollo e
prodotti a base di pesce, sono tutti prodotti a base di carne cruda o di pesce crudo. Dobbiamo
puntualizzare che i prodotti insaccati, a base di carne, in quanto tali, hanno subito un processo di
maturazione e fermentazione da parte della flora lattica in essi contenuta.
69
Tab. 5: Categorie di campioni analizzati.
N° dei campioni Tipologia del campione
73 Carne bovina
37 Carne di suino
5 Carne di pollo
10 Prodotti a base di pesce
21 Prodotti a base di carne
15 Formaggi a latte crudo
24 Latte crudo bovino
1 Latte crudo ovino
Totale 186

I campioni sono tutti a base cruda, in quanto un qualsiasi processo termico, anche solo la semplice
bollitura, andrebbe a distruggere le cellule di Yersinia enterocolitica eventualmente contenute.
I formaggi a base di latte crudo (principalmente ovino) non hanno subito nessun tipo di processo di
termizzazione, ma soltanto processi di fermentazione e maturazione, grazie alla presenza della flora
lattica contenuta nel prodotto stesso. Yersinia enterocolitica viene facilmente distrutta dai normali
metodi di cottura. Alcuni prodotti sono stati analizzati subito, le analisi sono state eseguite sul
prodotto fresco, mentre altri sono stati congelati, a una temperatura di -20 °C, in modo da eseguire
le procedure di analisi in un secondo momento. Il processo di congelamento/scongelamento non
altera le caratteristiche del batterio, che rimane comunque vitale e non altera le caratteristiche
organolettiche dell'alimento senza apparenti segni di deterioramento (Hudson and Mott, 1993).
Per i campioni a base di carne si è proceduto, inizialmente, a una valutazione qualitativa per il
batterio in esame, soltanto in presenza di positività, si è proceduto a una valutazione quantitativa
del microrganismo tramite il metodo MPN. Per i campioni di latte crudo si è proceduto, invece, fin
da subito, ad una valutazione quantitativa tramite diluizioni seriali, fino alla diluizione -2 con
metodo MPN. I diversi campioni analizzati originavano da diverse aree della Toscana, tra cui:
70
Massa( N°campioni 5)
Pisa(N°campioni 5)
Lucca(N° campioni 39)
Livorno(N°campioni 93)
Pistoia(N°campioni 8)
Grosseto(N°campioni 5)
Fig. 7: Provenienza geografica dei campioni analizzati.

3.1 Procedura di analisi qualitativa
L'analisi qualitativa, che è stata applicata, fa riferimento alla metodica standard internazionale EN
ISO 10273: 2003. La procedura consta di diverse fasi.
3.1.1 Arricchimento in terreni liquidi selettivi
25g di campione sono posti all'interno di buste presto chiuso grandi e addizionati con 225 ml di
PSB, in seguito omogeneizzata da stomacher.
Successivamente, 1 ml della sospensione in PSB è stato addizionato a 9 ml di ITC ( Irgasan
Ticarcillin Chlorate broth ). L'arricchimento in brodo ITC è stato proposto per l'isolamento di
Yersinia enterocolitica biosierotipo 4/O: 3, ma non per il biosierotipo 1B/O: 8, 2/O: 9 o 2/O: 5,27.
Il primo terreno liquido viene posto ad incubare a 25°C per 5 giorni senza agitazione, mentre il
secondo è posto a 25°C per 48 ore.
3.1.2 Semina in piastra ed identificazione
Le colture ottenute dal precedente arricchimento, vengono seminate, per mezzo di un'ansa sterile,
sulla superficie dei due seguenti terreni solidi:
CIN (Cefsulodin Irgasan Novobiocin) il quale è preparato addizionando al CIN agar base, il
supplemento selettivo con cefsuldina e novobiocina. E' un terreno altamente selettivo per
l'isolamento di Yersinia. I batteri Gram positivi e parte di quelli Gram negativi, sono inibiti dagli
agenti selettivi presenti nel terreno di base, sodio desossicolato, cristall violetto e Irgasan; gli
antibiotici inibiscono la crescita degli altri enterobatteri. Il mannitolo è presente quale carboidrato
fermentabile: i batteri in grado di fermentarlo inducono un' acidificazione del terreno con
precipitazione del sodio desossicolato e assorbimento del rosso neutro;
SSDC( Salmonella/Shigella agar con Sodium Desoxycholate e Calcium Chloride).
71

Tra la fase di arricchimento e quella di semina in piastra, è previsto un trattamento rapido con KOH
0.5% al fine di eliminare l'eventuale flora “background”, che si è sviluppata durante la fase
precedente.
Con una pipetta sterile da 1ml vengono trasferiti 0.5 ml della coltura ottenuta con PSB in una
provetta contenente 4.5 ml di KOH 0.5% in soluzione salina e subito vortexati. Dopo 20-25 secondi
viene seminata immediatamente, con l'ausilio di un'ansa sterile, la superficie di una piastra CIN agar
così da ottenere colonie ben separate.
La coltura ottenuta in ITC broth, è seminata con ansa sterile sulla superficie di una piastra di
SSDC. Tutte le piastre così seminate, vengono capovolte e poste in termostato a 30°C, segue una
lettura dopo 24 ore e se necessario dopo 48 ore, per verificare la presenza di colonie caratteristiche
di Yersinia enterocolitica.
Sul primo terreno (CIN), le colonie sospette si presentano di piccole dimensioni (1mm o meno), di
colore rosa intenso, lisce, opache, con un centro rosso caratterizzato da bordo traslucido.
Sul secondo terreno, le colonie caratteristiche del microrganismo, si presentano, anche in questo
caso, di piccole dimensioni (1mm o meno) ma di colore grigio con un margine indistinto.
72
Fig. 8: Yersinia enterocolitica su terreno CINagar.

3.1.3 Conferma
Da ciascuna piastra dei due terreni selettivi vengono selezionate cinque colonie caratteristiche e/o
sospette di Yersinia enterocolitica. Tali colonie vengono seminate, tramite l'utilizzo di un'ansa
sterile, su Agar Nutrient (NA) per ottenerne colonie in gran numero e ben isolate. Successivamente,
vengono incubate a 30°C per 24 ore.
Nel caso in cui fossero presenti colonie sospette di Yersinia enterocolitica, si procede con
l'esecuzione delle prove presuntive e test di conferma:
• test presuntivi;
• test biochimici.
La procedura qualitativa ISO 10273: 2003, prevede inoltre, test di biotipizzazione, test per stabilire
criteri di patogenicità e test sierologici, che non sono stati eseguiti durante questo lavoro di tesi.
I test presuntivi previsti sono:
• Ricerca dell'ossidasi:
questa metodica viene eseguita prelevando una porzione di ciascuna colonia caratteristica
selezionata, e usando un'ansa sterile si striscia su una carta da filtro imbevuta con il reagente
ossidasi.
Si considera il test negativo, quando il colore della cartina non vira al malva, violetto o blu intenso
entro 10 secondi.
73

• Ricerca dell'ureasi:
la colonia sospetta viene seminata con un'ansa sterile, appena sotto la superficie del terreno liquido
UIM ( Urea Indole Medium), di colore rosso/arancione. Viene posta in incubazione a 30°C per 24
ore. Lo sviluppo di una colorazione viola/fucsia del terreno indica una reazione ureasi positiva. Se
questo non avviene, la reazione è negativa. Yersinia enterocolitica dà per lo più una reazione
positiva in 1-5minuti. La registrazione del tempo di reazione può avere valore diagnostico.
E' stato anche utilizzato, per il medesimo scopo, un altro terreno liquido, Urea Broth (UB), di colore
giallo chiaro, posto all'interno di piccole provette. Se la colonia inoculata possiede l'ureasi, l'azione
di idrolisi dell'urea presente nel terreno, porta al viraggio del colore da giallo chiaro a fucsia. Al
contrario, se il colore rimane invariato, indica una reazione ureasi negativa.
74
Fig. 9: Oxidase strips.
Fig. 10: UIM negativo/positivo. Fig. 11: UREA BROTH negativo/positivo.

• Ricerca della triptofano deaminasi:
alla coltura inoculata nella provetta con terreno UIM, dopo incubazione, viene aggiunta una
quantità di 0.1- 0.2 ml di Reattivo di Kovac per il rilevamento dell'indolo. Se il ceppo possiede
l'enzima triptofano-deaminasi, questo scinde il triptofano contenuto nel terreno liberando indolo,
che essendo più volatile affiora in superficie. La formazione di quest'ultimo, è valutata visivamente
attraverso lo sviluppo di un anello rosso, sulla superficie della coltura, che avviene circa in 15
minuti, spesso anche in tempi più brevi.
• Semina in Triple Sugar Iron Agar (TSI Agar):
le colonie sospette vengono seminate sul TSI agar per strisciamento sulla sua superficie inclinata
(becco di clarino) e per infissione, con un' ansa sterile, il fondo dell'agar. Le provette di terreno
seminate vengono poste in termostato a 30°C per 24 ore.
Il profilo tipico dell'attività dei ceppi di Yersinia enterocolitica è il fondo e il becco giallo senza
bolle o spaccature del terreno, questo esprime la loro capacità di fermentare alcuni zuccheri
contenuti nel terreno senza la produzione di gas.
75
Fondo giallo e becco rosso> glucosio +Fondo e becco rosso> gluc./lattosio e saccarosio-Fondo e becco giallo> lattosio e/o saccarosio +Fondo nero> produzione di H2SBolle o spaccature del terreno> formazione di gas
Fig. 12: Agar positivo.

A questo punto, si procede con l'identificazione biochimica delle colonie che hanno dimostrato di
possedere le caratteristiche presuntive ricercate.
• Test Biochimici:
Su ogni ceppo fin qui caratterizzato, è stato eseguito il sistema miniaturizzato API20E
(Biomérieux®), al fine di ottenere una conferma di tipo biochimico a livello di specie.
Nel momento in cui si ottiene un buon grado di identificazione (ID%) del ceppo alla specie di
appartenenza Yersinia enterocolitica, si assegna un numero di identificazione e si inserisce in
cryobank a -80°C.
76
Fig. 13: Profilo biochimico di Yersinia enterocolitica su API 20E (Biomérieux®).

77
Fig. 14: Diagramma di flusso: Procedura qualitativa.

3.2 Procedura analisi quantitativa: il metodo MPN
Oltre alla valutazione qualitativa, si procede a una valutazione quantitativa, attraverso un'analisi di
tipo statistico, tramite il metodo MPN ( Most Probable Number): il metodo permette di stimare la
densità della popolazione microbica ottenuta, dopo aver preparato la soluzione madre e le diluizioni
seriali decimali del campione originario, all'interno di buste presto - chiuso e provette.
La tecnica di conteggio MPN o tecnica di conteggio con tubi multipli, è una procedura di
numerazione utilizzata per stimare la densità di una popolazione di microrganismi vitali, in un dato
campione. Si tratta di un metodo statistico, basato sulla probabilità di rilevare sviluppo microbico,
dopo coltura in tubi multipli, di substrato liquido, di diluizioni seriali del campione. Lo sviluppo
microbico, dopo incubazione dei brodi inoculati, può essere valutato osservando l' eventuale
intorbidamento della coltura, oppure, valutando particolari attività metaboliche del microrganismo o
della popolazione microbica da numerare (produzione di gas, di acidi ecc.). La tecnica è utilizzata in
associazione con una tavola statistica, che fornisce il valore del numero più probabile di
microrganismi per varie combinazioni di tubi positivi.
Il campione, dovrebbe essere diluito in maniera tale che le diluizioni più spinte non presentino
microrganismi (“diluizione all’estinzione”). I migliori risultati si hanno quando, tutti i tubi inoculati
con le diluizioni più basse risultano positivi e tutti i tubi inoculati con le diluizioni più alte risultano
negativi.
Per aumentare l’accuratezza statistica dell’MPN è necessario inoculare con ciascuna diluizione del
campione più tubi di brodo nutritivo.
La variabilità statistica della distribuzione batterica è meglio stimata al crescere dei tubi inoculati o
che sono praticamente inoculabili con ogni diluizione.
78

Come già accennato, la stima del MPN/ml o g di campione, è letta su apposite tavole MPN, in base
al numero di tubi positivi per ciascuna serie inoculata con una data diluizione.
Esistono tavole MPN per inoculi di 3, 5 e 10 tubi.
Una prima tavola statistica per la determinazione del MPN fu proposta da Mc Crady nel 1915. Una
versione più recente delle tabelle MPN è stata proposta da de Man nel 1983. In queste tavole, oltre
al numero più probabile di microrganismi, è riportato l’intervallo, con il valore massimo e minimo,
entro il quale cade il 95% degli altri valori possibili per una specifica combinazione di risultati
(http://wpage.unina.it/villani/eLAB6.html).
Oppure, si può ricorrere al calcolo del valore MPN, tramite la semplice formula qui di seguito, e poi
si procede al confronto con la tabella di rifermento:
MPN=N⋅F⋅Vs
V
N: valore letto sulla tabella di riferimento
F: fattore di diluizione della serie
V: fattore di diluizione base della tabella (1-0,1-0,5)
Vs: quantità di riferimento (es.1 ml, 25 g)
Questo metodo di analisi, offre diversi vantaggi:
• è relativamente semplice;
• vi è maggiore probabilità, rispetto al metodo di conta in piastra convenzionale, che i risultati
ottenuti da laboratori diversi siano concordanti;
• utilizzando appropriati terreni selettivi e differenziali, il metodo consente la determinazione
di specifici gruppi di microrganismi (Jay J.M. 2005).
79

Ciascuna busta e ogni provetta, rappresenta una singola prova qualitativa sviluppata singolarmente
fino alla lettura finale.
Le tre serie di provette permettono di alzare la sensibilità del metodo fino a 0,03 MPN/ml.
La procedura consiste nelle seguenti fasi:
1. si dispensano 10 ml di campione in ogni busta presto- chiuso piccola (3 in totale) da
stomacher, a cui vengono aggiunti 90 ml di PSB. In questo modo si ottiene la diluizione “
00” ;
2. viene preparata allo stesso modo un quarta busta, usata come soluzione madre del campione
(diluizione 1:10), che verrà distribuita in tre serie di provette;
3. la prima serie di provette, si ottiene distribuendo 10 ml di soluzione madre in tre provette
vuote, le quali rappresentano la diluizione “0” del campione originale;
4. la seconda serie di provette, si ottiene ripartendo 1 ml della soluzione madre in ciascuna
delle tre provette con 9ml di PSB, otteniamo la diluizione “ -1”;
5. la terza serie di provette, si ottiene dispensando 0,1 ml di soluzione madre, in ciascuna delle
tre provette contenenti 9,9 ml di PSB, in modo da ottenere la diluizione “ -2”.
80
Diluizione “00”
Diluizione “00”
Diluizione “00”

Le buste presto-chiuso e le provette vengono messe ad incubare a 25°C per 5 giorni non in
agitazione. Successivamente, si procede alla semina in piastre CIN con pre-trattamento in KOH al
0,5% ( semina in piastra entro 20 secondi ). Dalla combinazione dei risultati di buste e provette,
attraverso la consultazione della tabella riportata nel metodo USDA 2008, si ottiene il valore
ricercato espresso come numero più probabile.
81
Diluizione “0” Diluizione “-2”Diluizione “-1”
Soluzione madre per diluizioni scalari

Tabella MPN (3 tubi per diluizione) Diluizione 10 -1/0 – 0,lg:
82
Tab. 6: Schema di lettura metodo MPN.
Limiti di confidenzaCombinazioni MPN/g Al 95%positive Inferiore superiore
0-0-0 <0,03 <0,005 0,090-0-1 0,03 <0,005 0,090-1-0 0,03 <0,005 0,130-2-0 _ _ _1-0-0 0,04 <0,005 0,201-0-1 0,07 0,01 0,211-1-0 0,07 0,01 0,231-1-1 0,11 0,03 0,361-2-0 0,11 0,03 0,362-0-0 0,09 0,01 0,362-0-1 0,14 0,03 0,372-1-0 0,15 0,03 0,442-1-1 0,2 0,07 0,892-2-0 0,21 0,04 0,472-2-1 0,28 0,10 1,502-3-0 _ _ _3-0-0 0,23 0,04 1,203-0-1 0,39 0,07 1,303-0-2 0,64 0,15 3,803-1-0 0,43 0,07 2,103-1-1 0,75 0,14 2,303-1-2 1,2 0,30 3,803-2-0 0,93 0,15 3,803-2-1 1,5 0,30 4,403-2-2 2,1 0,35 4,703-3-0 2,4 0,36 13,03-3-1 4,6 0,71 24,03-3-2 11 1,50 48,03-3-3 >11,0 >1.50 >48,0

3.3 Protocollo operativo per la ricerca dei geni AIL, YST A e YST B nei ceppi di campo di Yersinia enterocolitica (Thoener P., et al, 2002- ISS rapporti ISTISAN 97/31)
Protocollo utilizzato per descrivere le modalità operative per l'estrazione del DNA, da ceppi
batterici di Yersinia enterocolitica, isolati dal laboratorio alimenti dell'Istituto Zooprofilattico e la
successiva ricerca mediante PCR dei geni AIL, YST A e B.
Sono stati sottoposti ad analisi di biologia molecolare, per la ricerca dei geni di patogenicità AIL e
YST A, mentre per i ceppi non patogeni YST B , tutti i ceppi di Yersinia enterocolitica isolati dal
laboratorio alimenti dell' Ist. Zooprofilattico Sperimentale delle Regioni del Lazio e della Toscana,
sezione di Pisa, dall'anno 2010 ad Ottobre 2015, per un totale di 59 ceppi batterici analizzati.
83
Tab. 7: Matrici alimentari fonti di isolamento di Yersinia enterocolitica conservati in microbank a -80°C.
Tipologia di alimento Ceppi isolati N°
Mozzarella 4
Intestino di cane 1
Latte bovino crudo 49
Feto ovino 1
Fusi di pollo 3

I ceppi batterici isolati tramite terreni selettivi (CIN) e identificati mediante test biochimici
miniaturizzati (API 20E Biomérieux®), vengono sottoposti alla procedura, la quale è formata da
diversi step:
• i ceppi individuati, o isolati precedentemente dal laboratorio, e conservati in criobank,
vengono nuovamente messi in coltura su piastre di terreno CIN e vengono posti ad incubare
per 24 ore a 25°C;
• successivamente, una sola colonia batterica viene seminata in brodo nutritivo TSYEB e
messa a incubare per 24 ore a 25°C.
La coltura batterica ottenuta, necessaria per l'estrazione del DNA, ha una concentrazione di ~ 107
-108 UFC/ml.
• Estrazione del DNA mediante bollituta ( METODO ISS)
Il DNA è stato estratto secondo il metodo descritto da ISS Rapporti ISTISAN 97/31, apportando
alcune modifiche.
Si trasferisce 1 ml di brodo di coltura in una eppendorf da 1,5 ml contrassegnata con nome/numero
del ceppo batterico sottoposto ad identificazione. Successivamente, si centrifuga la coltura a 1700 g
(RCF) per 10 minuti.
Il sovranatante viene eliminato e il DNA batterico viene risospeso con 1 ml di acqua distillata
sterile; le fasi appena descritte vengono eseguite in condizioni di sterilità.
Si accende il termoblocco, e le eppendorf contenenti il DNA batterico, vengono poste ad incubare
a 100 °C per 10 minuti. Il lisato cellulare, così ottenuto, viene conservato a -20°C fino
all'esecuzione della PCR.
84

• Preparazione della soluzione mix
Si procede, dunque, alla preparazione della mix, che conterrà tutti reagenti necessari
all'amplificazione del DNA estratto.
Durante la preparazione della soluzione tutti i reagenti necessari devono essere posti a contatto con
ghiaccio, in modo che la Taq polimerasi, particolarmente sensibile ai ripetuti congelamenti e
scongelamenti, non si deteriori. In questo studio è stata utilizzata una Taq polimerasi, Go Taq®
Flexi DNA polymerase Promega e il buffer, Green Go Taq Flexi® Buffer Promega, seguendo le
indicazioni del produttore per la preparazione della soluzione mix.
La preparazione della soluzione deve essere eseguita con l'ausilio di una cappa adibita solo alla
composizione della mix, così come l'aggiunta del DNA estratto, che avverrà sotto la sua apposita
cappa.
• Calcolo del volume della soluzione mix
Si considera il numero dei campioni di DNA da amplificare, un controllo negativo di
amplificazione, un controllo positivo e viene aggiunto un campione in eccesso. Il volume finale
necessario per l'amplificazione è di 50 µl, di cui 5 µl sono rappresentati dal DNA campione.
Per la preparazione della mix vengono utilizzati reagenti della ditta Promega:
5X Green Go Taq Flexi® Buffer 10 μl per campione (contenente il colorante SYBR Green);
Mgcl2 (25mM ) 2 μl per campione;
dNTp (10mM ) 1 μl per campione;
primer forward (20mM) 1 μl per campione ;
primer reverse (20mM) 1μl per campione;
Taq Go Flexi®polimerase (5u/ml) 0, 25 μl per campione;
85

DNA estratto 5 μl.
Il volume dell'acqua bidistillata Rnaasi/Dnasi free, si calcola sottraendo il volume totale da
amplificare (Vtot = 45μl x n° di campioni) alla somma dei volumi dei vari reagenti per la mix.
I primer utilizzati in questo studio sono:
Sequence (5'-3') Amplicon length (bp)
AIL
forward TAATGTGTACGCTGCGAG
reverse GACGTCTTACTTGCACTG
YST A
forward ATCGACACCAATAACCGCTGAG
reverse CCAATCACTACTGACTTCGGCT
YST B
forward GTACATTAGGCCAAGAGACG
reverse GCAACATACCTCACAACACC
86
351
79
146

• Amplificazione del DNA
Preparata la soluzione mix, vengono aggiunti 5μl di DNA estratto da amplificare sotto cappa adibita
a questa specifica fase.
Il termociclizzatore viene avviato e vengono impostate le condizioni per la PCR.
Condizioni di amplificazione:
L'amplificato, così ottenuto, viene conservato a -20°C.
L' analisi procede tramite elettroforesi su gel di agarosio.
87
-1ciclo di denaturazione a 95°C per 10 minuti- 25 cicli melting a 95°C per 15 secondi- Fase di annealing: 52°C per 30 secondi (per il gene AIL) 55°C per 30 secondi(per il gene YST A-B)
- Fase di elongation a 72C° per 30 secondi- Final extension a 72°C per 10 minuti
Fig. 15: Termociclizzatore per PCR.

• Elettroforesi su gel di agarosio al 2%
Si procede alla preparazione del gel di agarosio al 2%, per l' osservazione degli amplificati di YST
A, YST B e il gene AIL.
Si pesano i grammi di agarosio necessari, i quali vengono sciolti in un adeguato volume di
BUFFER TAE all'1%, mediante l'ausilio di una piastra riscaldante, in base alla grandezza del gel da
preparare.
Successivamente, si aggiungono adeguati microlitri di GEL REDTM(Biotium), il quale è una
molecola intercalante gli acidi nucleici, utilizzato in sostituzione del bromuro di etidio (sostanza
mutagena) utilizzato in passato, ed omogeneizziamo la soluzione.
Si distribuisce il gel ancora allo stato liquido, nell'apposita vaschetta, dopo aver posizionato il
pettine, cercando di non formare bolle durante il processo di solidificazione.
Il gel solidificato viene posizionato nella camera elettroforetica contenente BUFFER TAE all'1%. Il
pettine viene estratto e vengono caricati i pozzetti del gel con i campioni di DNA amplificato: 10 μl
di campione.
Per finire, viene posizionato il Marker da 100 bp (Gene RulerTM DNA Ladder. Thermofisher
ScientificTM ). La camera elettroforetica viene chiusa con l'apposito coperchio, si collegano gli
elettrodi, positivo e negativo, in base ai colori, e il voltaggio viene impostato a 100 mv per circa 40
minuti di tempo.
88
Fig. 16: Gel di agarosio.

• Osservazione del gel tramite transilluminatore
Si procede con l'accensione del transilluminatore e il gel viene posizionato al centro del suo
carrello scorrevole.
Si osserva l'immagine ottenuta tramite raggi UV, comparando le bande ottenute dagli amplificati
con quelle del marker, controllando che il controllo negativo non presenti bande.
89
Fig. 17: Transilluminatore chiuso / apertura del carrello interno.

4 Risultati
In questo lavoro di tesi sono stati analizzati 186 campioni, di matrici alimentari differenti,
pervenuti all' Istituto Zooprofilattico Sperimentale delle regioni Lazio e Toscana, dal Marzo 2015
ad Ottobre 2015, così suddivisi:
Il campionamento è stato eseguito sull'alimento fresco, quando possibile, oppure, in seguito a
congelamento/scongelamento ad una temperatura di -20°C, su prodotti crudi o pronti al consumo
umano. I campioni di latte crudo, prevalentemente di origine bovina, sono stati prelevati da
distributori sparsi sul territorio toscano e conservati in refrigeratore a una temperatura di 3°C±2,
prima di dare inizio alle analisi. Yersinia enterocolitica non subisce danni durante il processo di
congelamento, in quanto, essendo un organismo psicrofilo, è in grado di crescere anche a una
temperatura di 0°C / 4°C. Inoltre, la sua crescita non comporta alterazioni organolettiche
dell'alimento o apparenti segni di deterioramento (Hudson and Mott, 1993). Per quanto riguarda i
90
Tab. 8: Campioni analizzati dal Marzo 2015 ad Ottobre 2015.
N° dei campioni Tipologia del campione
73 Carne bovina
37 Carne di suino
5 Carne di pollo
10 Prodotti a base di pesce
21 Prodotti a base di carne
15 Formaggi a latte crudo
24 Latte crudo bovino
1 Latte crudo ovino
Totale 186

prodotti a base di latte, si tratta sempre di prodotti a base di latte crudo, che non hanno subito
processi di pastorizzazione o termizzazione. I formaggi analizzati sono per la maggior parte di
origine ovina.
L' analisi qualitativa eseguita sui prodotti a base di carne, di pesce e sui formaggi a latte crudo
ovino, hanno dato esito negativo. Si è riscontrata la presenza di Yersinia enterocolitica soltanto in 5
campioni di latte crudo bovino su un totale di 24 campioni, analizzati tramite analisi quantitativa
( MPN), con una percentuale del 2,7% sul totale dei campioni analizzati, e con una prevalenza del
20,8 % nei campioni di latte crudo bovino. I campioni di latte risultati positivi alle analisi sono stati
prelevati da bovine allevate nei territori delle province di Massa e Lucca. Successivamente, i ceppi
isolati sono stati identificati con un numero di serie e conservati in microbank a -80°C, in attesa di
ulteriori analisi.
91
Tab. 9: Prevalenza di Yersinia enterocolitica nei campioni analizzati.
Fig. 18: Prevalenza di Yersinia enterocolitica nei campioni dilatte crudo bovino analizzati.
Campioni Campioni CampioniTotali Latte crudo bovino Positivi enterocolitica
186 24 5 20,8%
Prevalenza Yersinia
20,8% Prevalenza Yersiniaenterocolitica
nel latte crudo bovino
79,2% campioni negatividi latte crudo bovino

4.1 Risultati dell'analisi molecolare
I 5 ceppi di Yersinia enterocolitica che sono stati isolati in questo lavoro di tesi, sono risultati
apatogeni, tramite le analisi di biologia molecolare eseguite successivamente. Non presentano,
quindi, i seguenti marker molecolari di patogenicità, gene AIL e YST A. Come già detto in
precedenza, in questo lavoro di tesi (Cap. 1 Introduzione), la presenza dei geni AIL e YST A è
fondamentale per il processo di patogenesi del batterio. Infatti, l'assenza di tali geni fa sì che il
microrganismo, non sia in grado di prendere contatto con le cellule epiteliali del tratto
gastro-intestinale, venendo facilmente eliminato dall'organismo ospite, senza instaurare l'infezione.
Lo scopo principale delle analisi di biologia molecolare, eseguite in seguito al protocollo
qualitativo/quantitativo, tramite PCR, è quello di andare a valutare la patogenicità, tramite la ricerca
dei geni AIL e YST A, che contraddistingue la specie batterica, e la diffusione del gene YST B nei
92
Fig. 19: Percentuale di campioni risultati positivi a Yersinia enterocolitica sul totale dei campioni analizzati.
Campioni positivi
Campioni negativi
Campioni positivi a YersiniaEnterocolitica 2,7%
Campioni negativi a Yersinia enterocolitica 97,3%

ceppi non patogeni.
Per questo, sono stati analizzati tutti i ceppi di Yersinia enterocolitica, conservati in criobank a –
80°C, isolati dall' Istituto Zooprofilattico dall'anno 2010 fino ad Ottobre 2015.
In questo lavoro è stata valutata la presenza di differenti geni di patogenicità a livello
cromosomico, che risultano essere maggiormente conservati nei diversi ceppi di Yersinia
enterocolitica. I marker di patogenicità ricercati sono tre: il gene AIL, il gene YST A e il gene YST
B.
La presenza di tali geni è stata verificata tramite PCR e successiva elettroforesi su gel di agarosio.
Dai dati ottenuti dal nostro lavoro, è possibile evidenziare la presenza di soltanto due ceppi
patogeni su un totale di 59 ceppi isolati (prevalenza del 3,4%), essi presentano sia il gene AIL, che
il gene YST A.
Tramite la Fig.20 raffigurante il gel, possiamo notare le bande di DNA che indicano la presenza sia
del gene AIL, sia del gene YST A, nello stesso ceppo batterico patogeno in esame. L'amplificato del
gene AIL ha una lunghezza di 351 bp e si posiziona più in alto rispetto all'amplificato del gene YST
93
Fig. 20: Elettroforesi su gel di agarosio. Le bande evidenziano la presenza del Gene AIL e del gene YST A in uno stesso ceppo in esame.
Amplificato AIL351 bp
Amplificato YST A79 bp

A, 79 bp, che invece si posiziona più in basso nel gel. E' nota in letteratura, la stretta correlazione
dei due geni, la presenza del gene AIL preannuncia la presenza del gene YST A e viceversa
(Platt-Samoraj A. et al, 2006).
94
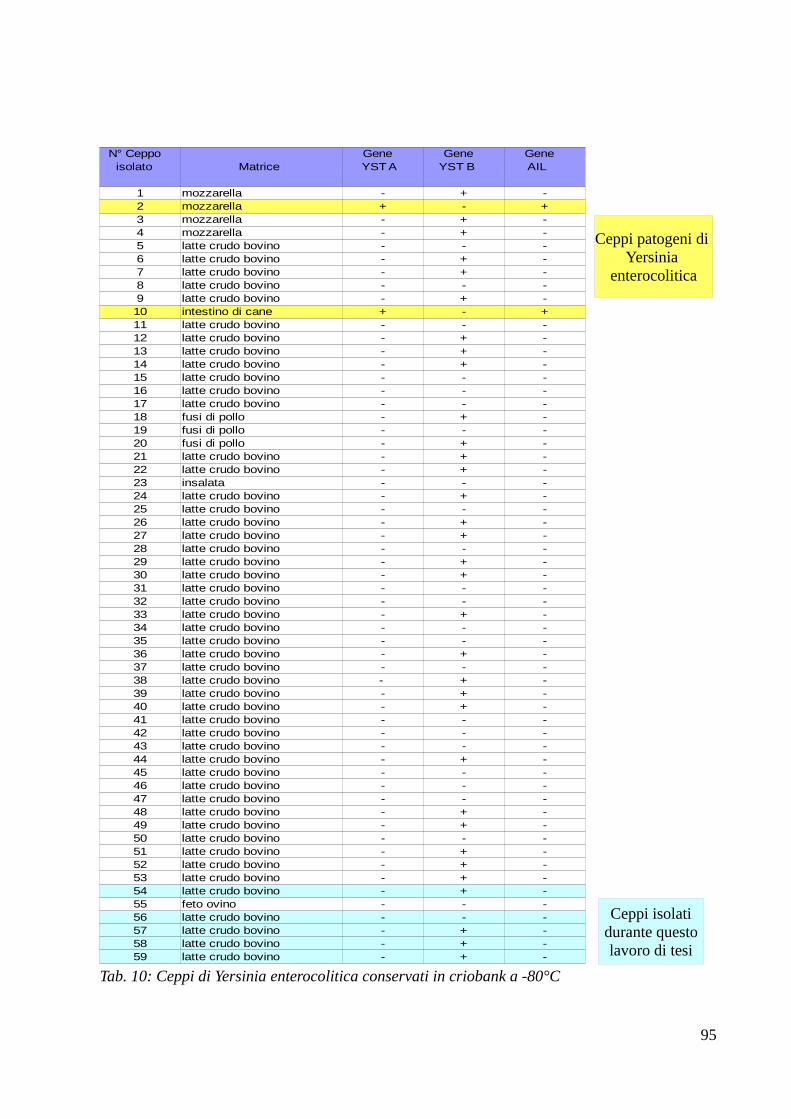
95
Ceppi patogeni di Yersinia
enterocolitica
Ceppi isolatidurante questolavoro di tesi
Tab. 10: Ceppi di Yersinia enterocolitica conservati in criobank a -80°C
N° Ceppo Gene Gene Gene isolato Matrice YST A YST B AIL
1 mozzarella - + -2 mozzarella + - +3 mozzarella - + -4 mozzarella - + -5 latte crudo bovino - - -6 latte crudo bovino - + -7 latte crudo bovino - + -8 latte crudo bovino - - -9 latte crudo bovino - + -10 intestino di cane + - +11 latte crudo bovino - - -12 latte crudo bovino - + -13 latte crudo bovino - + -14 latte crudo bovino - + -15 latte crudo bovino - - -16 latte crudo bovino - - -17 latte crudo bovino - - -18 fusi di pollo - + -19 fusi di pollo - - -20 fusi di pollo - + -21 latte crudo bovino - + -22 latte crudo bovino - + -23 insalata - - -24 latte crudo bovino - + -25 latte crudo bovino - - -26 latte crudo bovino - + -27 latte crudo bovino - + -28 latte crudo bovino - - -29 latte crudo bovino - + -30 latte crudo bovino - + -31 latte crudo bovino - - -32 latte crudo bovino - - -33 latte crudo bovino - + -34 latte crudo bovino - - -35 latte crudo bovino - - -36 latte crudo bovino - + -37 latte crudo bovino - - -38 latte crudo bovino - + -39 latte crudo bovino - + -40 latte crudo bovino - + -41 latte crudo bovino - - -42 latte crudo bovino - - -43 latte crudo bovino - - -44 latte crudo bovino - + -45 latte crudo bovino - - -46 latte crudo bovino - - -47 latte crudo bovino - - -48 latte crudo bovino - + -49 latte crudo bovino - + -50 latte crudo bovino - - -51 latte crudo bovino - + -52 latte crudo bovino - + -53 latte crudo bovino - + -54 latte crudo bovino - + -55 feto ovino - - -56 latte crudo bovino - - -57 latte crudo bovino - + -58 latte crudo bovino - + -59 latte crudo bovino - + -

Il maggior numero di ceppi, 49, sono stati isolati dal latte crudo bovino. Nessuno dei ceppi isolati
risulta essere patogeno, non presentano né il gene AIL, né il gene YST A. Si nota, che 27 ceppi
isolati dal latte crudo bovino presentano il gene YST B, mentre gli altri 22 ceppi non presentano
nessuno dei marker molecolari ricercati.
96
Tab. 11: Prevalenza del gene YST B in ceppi di Yersinia enterocolitica isolati da latte crudo bovino.
Ceppi isolati dal latte YST B NO MARKER
49 27 (55,1%) 22 (44,9%)
Fig. 21: Rappresentazione grafica dei ceppi di Yersinia enterocolitica isolati da campioni di latte crudo bovino dall'anno 2010 al mese di Ottobre 2015, in base alla presenza di YST B
YST B
NO MARKER
Ceppi isolati da lattecrudo bovino senza
marker ricercatiPrevalenza del 44,9%
Ceppi isolati da latte crudo bovino con YST B
Prevalenza del 55,1%

Considerando il totale dei ceppi isolati dall'anno 2010 ad Ottobre 2015, pari a 59 ceppi batterici,
emerge che 32 di questi, presentano il gene YST B con una prevalenza del 54,2%. Mentre, i restanti
25 ceppi, con una prevalenza del 42,4%, non mostrano nessuno dei marker genetici ricercati in
questo studio. Inoltre, soltanto due dei 59 ceppi risultano essere patogeni, come già detto in
precedenza, presentano, sia il gene AIL, sia il gene YST A, con una prevalenza del 3,4%, così come
mostra la tabella seguente:
97
Tab. 12: Prevalenze dei geni AIL, YST A e YST B nei ceppi isolati totali dal 2010 ad Ottobre 2015.
Gene Prevalenza del gene
AIL 3,4%
YST A 3,4%
YST B 54,2%
NO MARKER 42,4%

98
Markerda 100bp
Fig. 23: Elettroforesi su gel di agarosio per la ricerca del gene YST B.
AIL 3,4%YST A 3,4%
Marker 100 bp
YST B Controllopositivo
YST B Ceppo risultatopositivo
YST B lunghezza dell'amplificato
146 bp
K- 1 2 3 4 5
Fig. 22: Prevalenze dei geni AIL, YST a e YST B nei ceppi totali isolati dall' anno 2010 ad Ottobre 2015.
AIL
YST A
YST B
NO MARKER
YST B54,2%
NO MARKER42,4%

Le Figure 23 e 24, mostrano due corse elettroforetiche su gel di agarosio al 2%, eseguite sui
medesimi ceppi isolati. Da notare, come la presenza di YST A non è accompagnata dalla presenza
del gene YST B e viceversa. La banda corrispondente al gene YST A è presente soltanto nel ceppo
di riferimento ( controllo positivo ), il quale non mostra la banda corrispondente al gene YST B.
Nel nostro caso, le analisi hanno rivelato che la presenza del gene YST A non è correlata alla
presenza del gene YST B e viceversa. Questo risultato è in accordo con i dati presenti in letteratura,
come nel caso dello studio eseguito da Peruzy M.F., nel 2014, in cui i ceppi analizzati, appartenenti
al biotipo 1A, presentavano il gene YST B nel 100% dei casi, ma non il gene YST A (Peruzy M.F.,
99
YST BControllopositivo
YST BCeppo risultatopositivo
K- 1 2 3 4 5
Marker
K- 2 3 4 5 6 7 8 9 10
MarkerAssenza della banda di YST B
Fig. 24: Elettroforesi su gel di agarosio per la ricerca del gene YST A e del gene YST B nei medesimi ceppi.

2014)
4.2 Discussione dei risultati
L'attenzione verso il genere Yersinia spp nasce dal fatto che negli ultimi decenni, si è registrato un
aumento dei casi di yersiniosi in diversi paesi europei, collocando la malattia al terzo posto tra le
zoonosi maggiormente notificate. La situazione italiana presenta lacune, sia dal punto di vista della
notifica obbligatoria dei casi riscontrati nel paese, sia dal punto di vista della ricerca in laboratorio
del batterio. Infatti, il genere Yersinia non rientra nei criteri microbiologici applicabili ai prodotti
alimentari, in base ai criteri dettati dal Reg. CE N° 2073/2005, e in nessun piano nazionale o
sovranazionale. In letteratura inoltre, scarseggiano i lavori che permettono una quantificazione del
batterio nelle diverse derrate alimentari. Non sono, quindi, disponibili dei reali tassi di incidenza nel
nostro paese; e poiché la maggior parte di casi di yersiniosi si verifica sporadicamente senza
un'origine apparente, l'epidemiologia dell'infezione da Yersinia enterocolitica è complessa e non del
tutto compresa (Fredriksson-Ahomaa et. al 2003).
Il mondo animale è un serbatoio importante per ceppi di Yersinia spp. Diversi animali portano
sierotipi unici del batterio che sono coinvolti nelle infezioni umane. Numerosi studi sono stati
effettuati per isolare ceppi di Yersinia da diversi animali. Tuttavia, la maggior parte dei ceppi isolati
da fonte animale, differisce sia biochimicamente, sia sierologicamente da ceppi isolati da individui
umani affetti da yersiniosi. I suini sono stati implicati come un importante serbatoio di sierotipi di
Yersinia coinvolti nelle infezioni umane. L'incidenza di Yersinia nei maiali varia, oltre che da paese
a paese, anche all'interno della stessa nazione. I ceppi di Yersinia enterocolitica che appartengono
ai sierotipi associati con le malattie umane, sono stati isolati frequentemente da tonsille, lingue e
materiale fecale di maiali da macello. Il tasso di isolamento del microrganismo dalle tonsille e
100

lingue dei maiali è generalmente maggiore del tasso presente nelle feci. In diversi paesi, Yersinia
enterocolitica appartenente al biosierotipo 4/O: 3, ha dimostrato di essere quello predominante in
suini asintomatici. L'alta prevalenza dei ceppi O: 3 nei suini dimostra, che probabilmente, esiste un
certo livello di specificità per questo ospite animale (Schaake J., et al 2014).
La stretta relazione genetica tra i ceppi isolati nei suini e quelli isolati a livello umano, è stata
ampiamente dimostrata tramite parecchi metodi basati sul DNA.
Le tonsille di suini infettati dal microrganismo possono facilmente contaminare la carcassa, le
frattaglie e l'ambiente durante i processi di macellazione. Tramite metodi di biologia molecolare è
stato provato che la carne e le frattaglie di suino, contaminate da Yersinia, sono un importante
veicolo di trasmissione per i ceppi patogeni ( Bari Md. L., et al 2011).
Pur essendo la carne suina cruda una delle principali fonti di infezione per l'uomo di Yersinia
enterocolitica, nel nostro caso, le analisi sui campioni di carne di maiale hanno dato esito negativo.
L'animale da macello rappresenta, nella catena di produzione degli alimenti, il maggior serbatoio di
microrganismi, soprattutto di specie patogene o patogene facoltative. Questi esercitano un'intensa
azione infettante sulle carcasse degli animali macellati, che possono servire come possibile veicolo
del germe, e quindi, contaminare i prodotti finiti. Inoltre, se si considera che numerose tossinfezioni
derivano più da errori di manipolazione durante e dopo la preparazione, che dall'alimento stesso, si
può comprendere il ruolo di Yersinia enterocolitica come agente a rischio sanitario (Allegra A.M.,
et al 2003).
I ceppi di Yersinia enterocolitica associati con le malattie umane sono stati isolati frequentemente
da tonsille, lingue e materiale fecale di maiali da macello. I suini sono il principale serbatoio del
sierotipo 4/O:3 considerato a bassa patogenicità, anche se l'animale non sviluppa sintomatologia. La
contaminazione delle carcasse avviene tramite il contatto con tonsille o contenuto intestinale
101

dell'animale contaminato dal batterio, oppure, durante il processo di macellazione, in condizioni di
scarsa igiene o tramite procedure non ottimali. Quindi, le tecniche di macellazione e l'igiene di tali
processi possono influenzare il tasso di contaminazione delle carni (Andersen 1988).
E' da evidenziare l'elevato rischio di contaminazione dei prodotti durante l'asportazione della lingua,
della faringe e nell'esecuzione della visita ispettiva post-mortem. Il poco frequente isolamento di
questo microganismo in prodotti a base di carne suina, ad eccezione di quelli contenenti la lingua,
può essere dovuto a metodologie di ricerca poco sensibili; studi recenti, indicano che l'utilizzo di
terreni più selettivi o di tecnologie più sofisticate, aumenti la percentuale di isolamento di ceppi
patogeni di Yersinia enterocolitica.
Da quanto sopra esposto, la prevenzione della contaminazione delle carcasse avviene ponendo
attenzione alle seguenti fasi di macellazione:
• rimozione dell'intestino;
• escissione della lingua e delle amigdale;
• incisione dei linfonodi mandibolari o disossamento della testa.
Alcuni autori hanno appurato che l'utilizzo di bung cutter, associato alla chiusura del retto,
determini una notevole diminuzione della contaminazione della carcassa da parte di Yersinia
enterocolitica, rispetto all' eviscerazione manuale (Allegra A.M. et al, 2003).
Contrariamente ai risultati da noi ottenuti, molti studi dimostrano come la carne di suino sia una
fonte di contaminazione di ceppi di Yersinia enterocolitica, patogeni per l'uomo. Infatti, nello studio
eseguito da Roccato A. nel 2015, su 158 campioni di carne suina, si è evidenziata una positività per
Yersinia enterocolitica in 69 campioni. Mentre, Fondrevez in Francia, nel suo lavoro del 2014, dopo
aver analizzato 3120 campioni di carne di maiale, rilevò una prevalenza di 13,7% (414 campioni
positivi) di ceppi patogeni appartenenti a Yersinia enterocolitica. In questo studio, il target di
campionamento furono le tonsille, in quanto maggiormente rappresentative della contaminazione da
102

parte del batterio dell'animale suino, rispetto al contenuto intestinale o rispetto alle feci. La
prevalenza del 13,7% di Yersinia enterocolitica patogena trovata da Fondrevez, risultò essere bassa
rispetto alle altre prevalenze di altri stati europei. Infatti, in Germania si aveva una prevalenza del
22%, 34% in Svizzera, 44% nel Regno unito tra il 2003 e il 2005, del 37,4% nel 2008 in Belgio, del
93% in Spagna, e del 44 % in Belgio, del 32% in Italia tra il 2005 e l'anno 2007. (Fondrevez M., et
al, 2014).
Anche Paixao R. nella sua ricerca del 2012, su 720 campioni tra tamponi di lingua e tonsille di
maiale, tamponi ambientali e campioni di frammenti di maiale, individuò 442 campioni positivi
alla presenza di Yersinia enterocolitica. Circa il 72,40% dei ceppi isolati appartenevano al biotipo
4/O: 3 (Paixao R. et al, 2012).
Nel nostro caso l'assenza di esito positivo delle analisi, ci fa presupporre che i processi di
macellazione degli animali avvengano in maniera ottimale, che non si verifichi in alcun modo
103
Fig. 25: Prevalenza di Yersinia enterocolitica nei Paesi europei tra il 2003 e il 2014 in campioni dicarne di suino (Fondrevez M., et al, 2014).

contaminazione delle carni tramite tonsille e faringe o di materiale fecale, durante il processo di
macellazione ed eviscerazione, e che non ci sia una cross-contaminazione tra gli animali durante il
trasporto o durante la fase di stabulazione in allevamento.
Le analisi sui campioni di carne di pollo hanno dato anch' esse esito negativo. Si è voluto procedere
con l'analisi dei campioni di questa matrice alimentare, in quanto in letteratura sono presenti diversi
lavori sulla prevalenza di Yersinia enterocolitica in questa tipologia di carne. Il pollame comprende
una porzione sostanziale della dieta umana. Infatti, il consumo di carne di pollo è notevolmente
aumentato. Capita R. nel suo studio svolto nel 2002 in Spagna, tramite l'analisi di 40 carcasse di
pollame refrigerate, osservò che 26 campioni erano contaminati da batteri appartenenti a Yersinia
spp. (prevalenza del 65%) e 20 di esse erano contaminate da Yersinia enterocolitica, con una
prevalenza del 50% (Capita R., et al 2002).
Shanmugapriya nel 2014 in India, attraverso l'analisi di 44 campioni alimentari, suddivisi in 20
campioni di pollo e 24 campioni di pesce, individuò una prevalenza di Yersinia enterocolitica del
25% su 6 campioni di carne di pollo risultati positivi alle analisi. La prevalenza risultava essere
molto elevata rispetto ai report precedenti ( Shanmugapriya S., et al, 2014).
Il tasso di incidenza di Yersinia enterocolitica nella carne di pollo rilevata in letteratura è molto
variabile. Alcuni autori giustificano la bassa incidenza con la scarsa capacità competitiva del
batterio nei campioni di cibo. In accordo con quanto detto sopra, la presenza del batterio nelle
carcasse di pollo è attribuita a una contaminazione post-cottura e a una moltiplicazione del
batterio durante la fase di refrigerazione. Per questa ragione, l'igiene delle fasi di processazione
della carne di pollo è un fattore molto importante per lo sviluppo e la proliferazione di Yersinia
enterocolitica (Capita R., et al, 2002).
In passato, infatti, sono stati isolati ceppi di Yersinia enterocolitica anche da campioni di fusi di
104

pollo dal laboratorio alimenti dell' istituto IZSLT sezione di Pisa. I 3 ceppi isolati sono risultati
essere non patogeni, tramite l'esecuzione delle analisi di biologia molecolare eseguite nel corso di
questo lavoro di tesi, con una prevalenza del 5,08% sul totale dei ceppi conservati in microbank.
Non presentano quindi, i marker di patogenicità da noi ricercati, gene AIL e YST A. Tuttavia, due di
questi ceppi isolati presentano il gene YST B, possiamo quindi ipotizzare la loro appartenenza al
biotipo 1A.
Il basso numero dei campioni di carne di pollo analizzati tra Marzo e Ottobre 2015, non ci ha
permesso di fare valutazioni più puntuali sulla prevalenza del microrganismo in questa matrice
alimentare.
In questo lavoro di tesi sono stati analizzati anche 73 campioni di carne cruda bovina, sia di origine
toscana, ma anche carne di importazione, infatti, 67 campioni sono di derivazione brasiliana.
L'analisi di questa matrice alimentare ha dato, anche in questo caso, esito negativo. I risultati
ottenuti sono in forte contrasto con i dati presenti in letteratura.
Infatti, nello studio condotto da Logue C.M. nel 1996 in Irlanda, tramite l'analisi di 340 campioni,
si individuò una prevalenza molto alta di Yersinia spp, del 89% in campioni di carne cruda, del
49% in campioni di carne cotta e una prevalenza più bassa, del 18,8%, in prodotti di carne
confezionati. Nel 9% dei campioni di carne cruda furono isolati ceppi patogeni di Yersinia
enterocolitica, questo si verificò anche nel 2% dei campioni di carne cotta. I sierotipi patogeni
maggiormente isolati furono O: 3 e O: 5,27 nella carne di manzo irlandese.
Il numero iniziale di Yersinia era generalmente < 2,7 log10cfu/g nella carne cruda, e <0,7
log10cfu/g nella carne cotta. L'aumento del numero delle cellule batteriche patogene durante il
periodo di refrigerazione della carne, poteva indurre l'insorgenza di sintomatologia clinica soltanto
in casi di ingestione di carne contaminata cruda o poco cotta. Esisteva un pericolo simile anche nel
105

caso in cui le carni cotte risultassero contaminate dopo i processi di trasformazione.
La dose infettiva minima per l'uomo non è stata accuratamente determinata, ma l'ingestione di
3,5x109 di cellule di Yersinia enterocolitica poteva indurre sindrome diarroica il giorno stesso
dell'avvenuta ingestione. Studi in vivo sui topi, dimostrarono che un livello di cellule di Yersinia
enterocolitica pari a 2x109 , causava infezione a livello del fegato, della milza, causando infine la
morte. Questo suggeriva come la dose infettante del microrganismo per l'uomo potesse essere molto
elevata (Logue C.M. et al, 1996).
Un altro studio, condotto in Giappone nel 1997, metteva in evidenza l'isolamento di ceppi di
Yersinia spp sia in campioni di carne di manzo, che di maiale e di pollo; 38 campioni di suino, su
un totale di 1278 campioni, risultarono contaminati da Yersinia enterocolitica, con una prevalenza
del 3,0%, mentre nella carne di manzo e pollo la prevalenza risultava minore, pari a 0,3%
(Fukushima H. et al, 1997).
In uno studio eseguito nella città di San Paolo, in Brasile, da Tassinari A.R. nel 1994, si
esaminarono campioni di carne e campioni a base di carne, rilevando una prevalenza del 40%,
confermando il ruolo di fonte di contaminazione dei prodotti a base di carne per l'uomo (Tassinari
A.R. et al, 1994).
Lo studio precedentemente citato, è in completo contrasto con i risultati negativi da noi ottenuti
sui campioni di carne di manzo, provenienti dal Brasile. Il mancato isolamento di ceppi di Yersinia
enterocolitica ci fa ben sperare sul miglioramento delle condizioni igienico sanitarie in questa area
geografica, sia per quanto riguarda i processi di macellazione attuati, sia per le norme
igienico-sanitarie di mantenimento degli animali durante la loro vita in allevamento.
Per le matrici alimentari di cui disponiamo di un numero consistente di campioni e analisi, si è
tentato di prevedere un'attesa di ritrovamento di Yersinia enterocolitica nel nostro territorio di
106

competenza. Essendo i nostri positivi pari a zero, l'elemento di valutazione più affidabile è
l'intervallo di confidenza. Inserendo i nostri dati in un calcolatore on-line ( utilizzando un sistema di
riferimento europeo: EURL Lm TECHNICAL GUIDANCE DOCUMENT for conducting shelf-life
studies on Listeria monocytogenes in ready-to-eat foods Version 3 – 6 June 2014 ) otteniamo i limiti
inferiore e superiore; per la carne di manzo, di cui disponiamo di 73 campioni, avremo un limite
inferiore di 0,03% (3 campioni positivi ogni 10000 analizzati), ed un limite superiore del 5% (5
campioni positivi per ogni 100 analizzati), con una confidenza del 95%. Mentre, per la carne di
suino, di cui disponiamo di 37 campioni, avremo un limite inferiore dello 0,07% (pari a 7 campioni
positivi ogni 10000 analizzati), e un limite superiore del 9% (pari a 9 campioni su 100 analizzati),
con un intervallo di confidenza del 95%.
La presenza di Yersinia enterocolitica nel latte crudo è da tempo conosciuta, il latte quindi, è uno
degli alimenti di contaminazione umana più frequente. Dalle nostre analisi, su 24 campioni di latte
crudo bovino, è emersa una prevalenza del 20,8%, questo è in accordo con altri studi precedenti
presenti in letteratura.
Nihal Yucel nel 2005 in Turchia, similmente al nostro studio, ha evidenziato come il latte crudo o
latte inadeguatamente pastorizzato possa essere coinvolto nella trasmissione di yersiniosi tra
animale e uomo. L'analisi di 100 campioni di latte evidenziò la presenza di Yersinia spp in 55
campioni, un' incidenza del 55%. Di questi campioni, 25 risultarono contaminati da Yersinia
enterocolitica (45,3% di incidenza). Mentre i campioni di formaggio, risultati positivi furono
14/100 con un'incidenza del 14%, soltanto 5 però risultarono positivi per Yersinia enterocolitica,
con un'incidenza del 35,7% (Yucel N., Ulusoy H. 2006). Anche in altre parti del mondo, la
prevalenza di Yersinia enterocolitica è alta: 10% in Danimarca (Christensen 1982), 11% in Australia
( Ibrahim, Mac Rae 1991), 36% in Marocco (Hamama et al 1992), 37 % in Italia ( Franzin et al
1984).
107

Aiello nel 1984, nei suoi studi su 316 campioni di latte crudo, provenienti da aziende presenti in
Sicilia, ottenne una positività al batterio in 8 campioni (prevalenza del 2,5%), derivanti sia da
animali di importazione che indigeni (Aiello et al 1984).
Il latte crudo e prodotti lattiero-caseari, permettono la trasmissione di yersiniosi umana. Esiste la
possibilità che il latte risulti contaminato, in quanto, sottoposto a tecniche di pastorizzazione
inadeguate che impediscono la completa eliminazione dei batteri patogeni, oppure, può essere
108
Dato risalente al1984 nel latte crudo bovino
Prevalenza ottenuta nel lattecrudo bovino
Tramite questo lavoro di tesi
Fig. 26: Prevalenza di Yersinia enterocolitica ottenute dal latte crudo bovino in questo lavoro di tesi confrontata con le prevalenze nel mondo dal 1982 all'anno 1992. (Yucel 2005)
Fig. 27: Prevalenza di Yersinia enterocolitica ottenute dal latte crudo bovino in questo lavoro di tesi confrontata con le prevalenze nel mondo.
Nazioneenterocolitica
Turchia 45,3%
Danimarca 10%
Australia 11%
Marocco 36%
Italia 37%
Dati da questo 20,8%lavoro di tesi
Prevalenza di Yersinia

ricontaminato successivamente, tramite manipolazione da operatore, oppure, tramite l'ambiente o
tramite i contenitori in cui viene conservato prima della messa in commercio (Schiemann D.A.,
Toma S. 1978).
In questo lavoro di tesi, le analisi sui prodotti lattiero-caseari a base di latte crudo ovino, non hanno
rilevato la presenza di Yersinia enterocolitica. La completa assenza del batterio nei prodotti
fermentati, formaggi a latte crudo, ma anche prodotti a base di carne e insaccati, è dovuta
probabilmente al processo di stagionatura e fermentazione, da parte dei batteri lattici, i quali sono
presenti o vengono aggiunti a questi alimenti durante la fase di produzione, tramite l'utilizzo di
“colture starter”. Esse danno inizio e indirizzano correttamente il processo di fermentazione,
permettendo anche un aumento del numero iniziale dei batteri lattici presenti nel prodotto, i quali se
presenti in numero esiguo, non sono in grado di dare inizio al processo fermentativo
spontaneamente, e non riescono a contrastare l' eventuale presenza di batteri patogeni
contaminanti l'alimento. Infatti, molti dei batteri lattici, utilizzati nei processi di produzione
alimentare, sono in grado di produrre sostanze con azione inibitoria (come le batteriocine) nei
confronti di batteri patogeni, inoltre, creano ambienti ostili, per esempio modificando il pH
dell'alimento, per quei microrganismi dannosi per la salute umana
(www.mednat.org/alimentazione/fermentazione_cibi_e_bevande.pdf.).
Infatti, Schiemann nel 1978, eseguì una serie di analisi su campioni di latte crudo e prodotti lattiero-
caseari, come formaggi, rilevando un' incidenza di Yersinia enterocolitica nei campioni di latte
crudo del 18,2%, rispetto all'incidenza nei campioni di formaggio che risultò essere circa la metà
9,2%. Inoltre, un campione di cagliata risultato positivo per Yersinia enterocolitica, conservato a
4°C, risultò poi essere negativo al batterio dopo 8 settimane. Quindi, i processi di manifattura del
formaggio apparirono deleteri per la crescita di Yersinia enterocolitica, essendo il tasso di
isolamento nei formaggi fermentati la metà del tasso individuato nei campioni di latte crudo. La
109

distruzione di Yersinia enterocolitica, sia durante la fabbricazione, che nella stagionatura del
formaggio, non è inaspettato, in quanto, le modifiche effettuate dai batteri lattici utilizzati nel
processo di produzione, sono dannose per molti batteri. Questo antagonismo risulta, non solo dall'
abbassamento del pH dalla produzione di acido lattico, ma anche da sostanze antibiotiche prodotte
da alcuni batteri lattici. Il prodotto finale fermentato non è favorevole a una moltiplicazione o alla
sopravvivenza di batteri gram negativi come Yersinia enterocolitica. E' improbabile che Yersinia
enterocolitica sopravviva al processo di pastorizzazione, è molto più probabile che il batterio sia un
contaminante post-pastorizzazione che si verifica con bassa frequenza (Schiemann D.A. 1978).
L'esito negativo delle nostre analisi eseguite su formaggi a latte crudo ovino, ci fa ipotizzare che il
processo di stagionatura e di fermentazione possono aver in qualche modo influito sull'eventuale
sviluppo di batteri patogeni. Quindi, anche nel caso in cui Yersinia enterocolitica fosse presente
nelle prime fase di maturazione del prodotto caseario, l'azione della flora lattica e le sue
modificazioni apportate al prodotto, possono aver inibito lo sviluppo ulteriore del batterio, in questo
modo, i protocolli di analisi non ne hanno evidenziato la presenza. Questo si può imputare, da una
parte all' azione antagonista del processo fermentativo lattico, dall'altra, all'eventuale bassa
discriminazione dei procedimenti di analisi, ma anche a una situazione igienico sanitaria ottimale
durante i processi di mungitura degli animali e standard di produzione elevati.
Dai dati relativi al conteggio di Yersinia enterocolitica nei campioni di latte crudo bovino risultati
positivi, si può considerare che quando è presente, lo è a concentrazioni relativamente basse, infatti,
abbiamo riscontrato valori che variano da 0,036 a 2,1 MPN /ml, mentre in letteratura si riporta una
dose infettante per l'uomo molto più alta, di almeno 108 batteri/ml per un individuo
immunocompetente (Fleming e Hunt, 2006). Generalmente, l'instaurarsi della malattia, yersiniosi,
non è determinata tanto dalla presenza del batterio nell'alimento, non è una relazione causa-effetto,
110

l'insorgenza dello stato clinico dipende generalmente, sia dalla carica infettante del batterio
presente nell'alimento, ma anche dalle condizioni di salute in cui si trova il soggetto. Infatti, un
individuo immunodepresso sarà sicuramente più suscettibile all'insorgenza di yersiniosi, rispetto ad
un soggetto immunocompetente.
L'età dell'ospite e il suo stato di salute e il sierotipo del batterio coinvolto nell'infezione, sono
importanti nel definire l'evoluzione della malattia (Cap.1 Introduzione. Geni di patogenicità e
yersiniosi). Inoltre, dai dati ottenuti dal Centro di Riferimento Regionale delle Tossinfezioni
Alimentari (CeRRTA), raramente il focolaio d'infezione, seppur sporadico, è in stretta correlazione
con la presenza di Yersinia enterocolitica, negli eventuali alimenti incriminati, quando è possibile la
loro identificazione.
Quindi, considerata la ristrettezza dei criteri microbiologici del latte crudo bovino, sia in
considerazione dei risultati da noi ottenuti, sia per il fatto che il latte crudo va consumato previa
bollitura (come previsto dall'o.m. 10 Dicembre 2008 e successive modifiche), è possibile tracciare
un quadro generale rassicurante sulla qualità igienico-sanitaria del latte crudo bovino prodotto e
distribuito nelle province di Lucca, Massa, Livorno e Pisa di competenza dell' Istituto
Zooprofilattico Sperimentale del Lazio e Toscana.
In questo lavoro è stata valutata anche la presenza di differenti geni di patogenicità a livello
cromosomico, che risultano essere maggiormente conservati nei diversi ceppi di Yersinia
enterocolitica.
I marker di patogenicità ricercati sono tre: il gene AIL, il gene YST A e il gene YST B. Alcuni
ceppi patogeni di Yersinia enterocolitica sono dotati di un plasmide pYV, che contiene una serie di
geni, che codificano per l'espressione di proteine (da 16 a 20), localizzate sulla membrana esterna
111

della parete. Il plasmide è presente soltanto nei ceppi virulenti, ma viene facilmente perso durante le
manipolazioni in laboratorio e durante le fasi di coltura. Per questo abbiamo focalizzato la nostra
attenzione su geni di natura cromosomica maggiormente conservati.
Il gene AIL codifica per una proteina di adesione che è coinvolta nel processo di invasione del
microrganismo. La presenza del gene AIL è strettamente correlata alla virulenza del batterio, si
ritrova solo ed esclusivamente nei ceppi patogeni di Yersinia enterocolitica (Fabrega A., Vila J.
2012). Inoltre, è nota la stretta correlazione tra la presenza del gene AIL e la presenza del gene YST
A.
I geni YST sono deputati all' elaborazione della enterotossina termostabile. I biotipi patogeni
primari, hanno la capacità di produrre enterotossina termostabile, grazie alla presenza del gene YST
A. La proteina yst è presente in diverse forme, codificata da forme differenti del gene YST, forma
A, B e C, di cui ancora oggi non se ne conosce pienamente il ruolo. I ceppi di Yersinia
enterocolitica appartenenti al biotipo 1A, ritenuti in passato non patogeni, riescono a produrre una
tossina termostabile a una temperatura di 28°C, grazie alla presenza del gene cromosomico YST B.
La produzione della tossina B, può essere indotta dal pH presente nell'ileo e questo suggerisce come
YST B, sia un importante fattore di virulenza nei batteri facenti parte del biotipo 1A (Singh I., Virdi
J.S., 2004).
La presenza di tali geni, è stata verificata tramite PCR e successiva elettroforesi su gel di agarosio.
I ceppi che presentano il gene YST B appartengono sicuramente al biotipo 1A. In passato i ceppi
appartenenti a questo biotipo, erano ritenuti ceppi apatogeni, ma recentemente, studi hanno
dimostrato, che vista la loro capacità di creare enterotossina termostabile, possono essere coinvolti
nell'insorgenza di malattie gastro-intestinali nell'uomo. Inoltre, il decorso della malattia è identico a
quella causata dai biotipi virulenti primari ( Platt-Samoraj A. et al, 2006). Peraltro, è stata
evidenziata la presenza di ceppi appartenenti al biotipo 1A in pazienti con manifestazioni cliniche
112
Marker YST B YST BMarker YST B YST B
Marker
K- 1 3 5 9
K- 1 2 3 4 5 6 7 8 9 10

simili a quelle provocate da bio-sierotipi patogeni (Thoener P. et al 2003).
Di tutti i sei biotipi di Yersinia enterocolitica, il biotipo 1A è quello più eterogeneo. I batteri
appartenenti a questo biotipo sono ubiquitari, e vengono isolati da diversi tipi di ambienti, come il
suolo, l' acqua e diverse tipologie di cibo, incluso vegetali e prodotti animali e da differenti specie
animali (Paixao R. et al, 2012).
In accordo con i nostri risultati, Roccato A. nel suo lavoro del 2015, ha individuato il gene YST B
in 47 campioni su un totale di 383, e soltanto in 9 casi ha individuato la presenza del gene AIL
(Roccato A. et al, 2015).
Inoltre, Paixao nel 2012, attraverso lo studio di 792 campioni, isolò 122 ceppi appartenenti al
biotipo 1A, tramite l'analisi di campioni di maiale e di tamponi ambientali provenienti dalle
superfici dei macelli. Nessun ceppo appartenente al biotipo 1A venne però isolato dalle tonsille e
dalle lingue dei maiali, in questi campioni furono individuati soltanto ceppi del biotipo 4/O: 3
patogeno per l'uomo (Paixao R. et al, 2012).
Thoerner nel 2003, individuò che il genotipo predominante nell'80% dei ceppi isolati di Yersinia
enterocolitica, appartenenti al biotipo 1A, presentavano il gene YST B, dimostrando che questi
ceppi erano privi del plasmide di virulenza, ma, per esempio, il gene INV risultava presente ma non
funzionale in molti dei ceppi (Peruzy M.F. 2014). L'assenza di un marcatore per la virulenza in
ceppi del biotipo 1A isolati da casi clinici umani, può essere illustrativa della patogenicità
intrinseca debole di questo biotipo. Il risultato clinico è probabilmente determinato più da fattori
dell'ospite che da fattori di virulenza batterica (Thoener P.,et al 2003).
Delibato E. nel suo studio del 2015, similmente ai dati ottenuti dalle nostre analisi, ottenne risultati
positivi per il gene YST B in ceppi appartenenti al biotipo 1A, e risultati negativi per
l'individuazione dei geni AIL e YST A. Lo studio evidenziò anche la necessità di caratterizzare a
livello genotipico i ceppi di Yersinia enterocolitica, allo scopo di giungere in tempi rapidi alla
113

diagnosi molecolare di tale patogeno (Delibato E. et al, 2015).
Dalle nostre analisi, abbiamo potuto differenziare i ceppi non patogeni, evidenziando la presenza
nel loro genoma del gene YST B, da quelli patogeni primari che presentano invece, sia il gene AIL,
sia il gene YST A (prevalenza 3,4%). La classificazione dei ceppi non patogeni (96,6%) al biotipo
1A, è fatta in base alla presenza nel loro genoma del gene YST B, gene che invece è completamente
assente nei ceppi patogeni. Inoltre, la percentuale dei ceppi non patogeni, 96,6%, si suddivide, a sua
volta, in un 54,2% di prevalenza di ceppi dotati del gene YST B, e un altro 42,4% di ceppi batterici
che non presentano nessuno dei marker ricercati in questo lavoro. Osserviamo che il genotipo
predominante nei ceppi analizzati è YST B+, caratteristico dei ceppi appartenenti al biotipo 1A.
I dati raccolti, suggeriscono la necessità di identificare target genici altamente conservati da
utilizzare per sviluppare un saggio molecolare, da impiegare come screening, per la presenza di
Yersinia enterocolitica negli alimenti. Tale approccio, risulterebbe particolarmente utile, alla luce
anche della complessità del metodo ISO, per l'isolamento del microrganismo. Inoltre, i dati ottenuti
suggeriscono che al fine della caratterizzazione degli isolati di Yersinia enterocolitica, è necessario
ampliare il pannello di geni di virulenza da indagare, in modo da aumentare il grado di
caratterizzazione genica possibile da effettuare sui ceppi batterici isolati.
Gli attuali metodi di isolamento di Yersinia enterocolitica, richiedono lunghi periodi d'incubazione,
di conseguenza, è sempre più sentito il bisogno di avere a disposizione nuovi metodi, che siano più
rapidi e possibilmente più sensibili. Da qui, la necessità dello sviluppo di un metodo molecolare, in
grado di determinare in maniera rapida, la presenza del batterio da campioni alimentari.
114

5 Conclusioni
I dati ottenuti in questo lavoro di tesi, tramite l'analisi qualitativa e quantitativa MPN, hanno
mostrato che in 186 campioni di matrici alimentari analizzati, soltanto 5 sono risultati positivi alla
presenza di Yersinia enterocolitica, con una percentuale del 2,7% rispetto a tutti i campioni
analizzati. I ceppi isolati sono stati ritrovati solo in campioni di latte crudo bovino con una
prevalenza del 20,8%.
Tutti i ceppi isolati sono risultati non patogeni grazie alle analisi di biologia molecolare eseguite.
Non presentano i geni marker di patogenicità ricercati in questo lavoro di tesi, gene AIL e gene
YST A.
L'interesse nel quantificare il microrganismo presente nelle matrici alimentari, nasce dal fatto che in
letteratura, sono scarsi i dati riguardanti tale aspetto, inoltre, sono carenti gli studi di quantificazione
del batterio in campioni alimentari. Ciò potrebbe essere utile al fine di valutare il comportamento
del microrganismo riguardo ai diversi fattori che possono influenzare sviluppo e crescita nel
prodotto.
I risultati del conteggio denotano, che nei campioni in cui sono stati isolati ceppi di Yersinia
enterocolitica, le cellule batteriche sono comunque presenti in piccole concentrazioni. La
contaminazione è prevalentemente nelle materie prime, come si evidenzia dai nostri dati e da quelli
presenti in bibliografia.
Dalle analisi di biologia molecolare, tramite PCR, possiamo confermare la variabilità per i geni
ricercati, AIL, YST A e YSTB, che caratterizza i batteri appartenenti a Yersinia enterocolitica. Pur
essendo AIL e YST A, geni cromosomici altamente conservati e fortemente coinvolti nel processo
patogenetico del batterio, i dati ottenuti suggeriscono la necessità di ampliare lo spettro di geni di
patogenicità altamente conservati da saggiare e ricercare, in modo da avere una maggiore
115

caratterizzazione dei ceppi isolati.
Infine, possiamo tracciare un quadro generale rassicurante sulla qualità e l'aspetto igienico-sanitario
delle matrici alimentari analizzate, relativamente alla presenza di Yersinia enterocolitica, prelevate
nelle aree geografiche della Toscana di competenza dell'Istituto Zooprofilattico Sperimentale delle
regioni Lazio e Toscana sezione di Pisa.
116

Bibliografia
• Ahvonen P., Thal E., Vasenius H. Occurrence of Yersinia enterocolitica in animal in Finland
and Sweden. Contibutions to Microbiology and Immunology 2: 135-136 (1973).
• Aiello P., Di Noto A. M., Guercio V. Isolamento di Yersinia enterocolitica in latte bovino
crudo. Istituto Zooprofilattico Sperimentale della Sicilia (1984).
• Alais C. Scienza del latte, 3a ed., Tecniche Nuove. pp: 471 (2000).
• Allegra A.M., Fezia G., Fontana E., Roceri M., Sommariva M., Tinelli F. Indagine sulla
presenza di Yersinia enterocolitica in suini macellati. Il Progresso Veterinario n°3 (2003).
• Andersen J.K. Contamination of freshly slaughtered pig carcasses with human pathogenic
Yersinia enterocolitica. Int. J.Food Microbiol. Vol 7 (3): 193-202 (1988).
• Andreis, Ottaviani. Manuale della sicurezza microbiologica degli alimenti e delle acque
(2002).
• Arthur T.M., Wheeler T.L. Shackeloford S.D., Bosilevac J.M., Nou X. and Koohmaraie M.
Effects of low-dose, low-penetration electron beam irradiation of chilled beef carcass
surface cuts on Escherichia coli O157:H7 and meat quality. J. Food Prot. Apr; 68(4):
666-672 (2005).
• Barberio A., Badan M., Vincenzoni G. Diagnosi e controllo della Salmonellosi nelle bovine
da latte. Large Animal Review 15: 147-152 (2009).
• Bari Md. L., Hossain Anwar M., Kenji Isshiki, Dike Ukuku. Behavior of Yersinia
enterocolitica in Foods. Journal of Pathogens (2011).
• Belk K.E. Beef decontamination technologies. National Cattlemen’s Beef Association.
http://www.beef.org/documents/ACFFC.pdf (2001).
117

• Bhaduri S. Effect of salt and acidic pH on the stability of virulence plasmid (pYV) in
Yersinia enterocolitica and expression of virulence-associated characteristics. Food
Microbiology. Vol. 28, n 1: 171-173 (2011).
• Bliska J.B., Falkow S. Bacterial resistance to complement killing mediated by ail protein of
Yersinia enterocolitica. Proc. Natl.Acad.Sci.USA. 89: 3561-3565 (1992).
• Bottone E.J. Yersinia enterocolitica: the charisma continues. Clin.Microbiol. Rev. 10:
257-276 (1997).
• Bottone E.J. Yersinia enterocolitca: overview and epidemiologic correlates. Microbes and
infections 1: 323-333 (1999).
• Bramley A.J., Mc Kinnon C.H. The microbiology of raw milk. In: Dairy Microbiology,
vol.1 Ed.: Robinson R.K., Elsierver , London, England, Pagg. 163-208 (1990).
• Cantoni C., Kramer J. Alimenti, Microbiologia e Igiene. Ed. Tecniche nuove (2011).
• Capita R., Alonso-Calleja C., Prieto M., del Camino Garcia-Fernandez M., Moreno B.
Incidence and pathgogenicity of Yersinia spp. Isolates from poultry in Spain. Food
Microbiology 19: 295-301 (2002).
• Cartier P. Le point sur de la qualité microbiologique de la viande bovine. Collection
Interbev « le point sur » (1997).
• Cattabiani F. Alcuni aspetti della patogenicità di Yersinia enterocolitica. Ann.Fac.Medic.
Vet. Di Parma Vol.XXII: 313- 321 (2002).
• Christensen S.G. The prevalence of Yersinia enterocolitica in slaughter anilas water and raw
milk in Denmark. In T.A. Roberts B, Hobbs J.H. B. Christian, e N. Skovgaard (Eds),
Psychrotrophic microorganisms in spoilage and pathogenicity. London: Academic Press.
(1982).
• Colavita Giampaolo. Igiene e Tecnologie degli Alimenti di Origine Animale. Le point
118

Vétérinaire Italie (2008).
• Del Bono, Renon Igiene ed educazione alimentare Agriteam (2001).
• Delibato E., Pucci E., Morleo A., Capuano F., proroga Y., Bilei S., Slawomir O., Luzzi I., De
Medici D. Yersinia enterocolitica patogena: sviluppo di metodologie molecolari per la
caratterizzazione dei geni di virulenza. Sidilv (2015).
• Edwards J.R., Fung D.Y.C. Prevention and decontamination of Escherichia coli O157:H7 on
raw beef carcasses in commercial beef abattoirs. Journal of Rapid Methods and Automation
in Microbiology. 14: 1-95 (2006).
• EFSA journal 7(11):1374 (2009).
• EFSA. Monitoring and identification of human enteropathogenic Yersinia spp. The EFSA
Journal 595: 1-30 (2007).
• Elder R.O., Keen J.E., Siragusa, G.R., Barkocy-Gallagher G.A., Koohmaraie M. and
Laegreid W.W. Correlation of enterohemorrhagic Escherichia coli O157:H7 prevalence in
feces, hides, and carcasses of beef cattle during processing. Proc. Natl Acad. Sci. USA 97,
2999-3003 (2000).
• Elmoslemany A.M., Keefe G.P., Dohoo I.R., Jayarao B.M. Risk factors for bacteriological
quality of bulk tank milk in prince Edward island dairy herds. Part.1 : Overall risk factors.
Journal of Dairy Science, 92: 2634-2643 (2009).
• Estrazione del DNA secondo la tecnica della bollitura. ISS rapporti ISTISAN 97/31 pag.
43-44.
• EURL Lm TECHNICAL GUIDANCE DOCUMENT for conducting shelf-life studies on
Listeria monocytogenes in ready-to-eat foods Version 3 – 6 June (2014 ).
• European Food Safety Authority. Community summary report on trends and source of
zoonoses and zoonotic agents in the european union in 2007. EFSA Journal 223: 5 -312
119

(2009).
• Fenwick S.G., Madie P., Wilks C.R. Duratione of carriage and trasmission of Yersinia
enterocolitica biotype 4, serotype O:3 in dogs. Epidemiology and infections 113: 471-477
(1994).
• Fleming D., Hunt D, Biological safety principles and practices. ASM press, Washington,
DC, USA, (2006).
• Fondrevez M., Minvielle B., Labbè A., Houdayer C., Rose N., Esnault E., Denis M.
Prevalence of pathogenic Yersinia enterocolitica in slaughter-aged pigs durng a one-year
survey, 2010-2011, France. International Journal of Food Microbiology 174: 56-62 (2014).
• Food Standard Agency (FSA). Clean Beef Cattle for slaughter. A guide for producers.
Published by the Food Standard Agency November 2004, Crown Copyright 2004, Printed in
England 5k FSA/0951/1104 (2004).
• Fournaud J. Filière viande, 3: 15 – 20. (1978).
• Frabrega A., Vila J. Yersinia enterocolitica: pathogenesis, virulence and antimicrobial
resistance. Enferm. Infecc. Microbiol.Clin. 30(1):24-32 (2012).
• Francis D.W., Spaulding P.L., Lovet J. Enterotoxin production and thermal resistance of
Yersinia enterocolitica in milk. Appl. Envir.Microbiol, July, p.174-176 (1980).
• Franzin L., Fantino P., Vigotto V. Isolation of Yersinia enterocolitica and Yersinia
enterocolitica-like organisms from raw milk in Italy. Curr. Micriobol., 10: 357-362 (1984).
• Frederiksen, W. A study of some Yersinia pseudotuberculosis-like bacteria (“Bacterium
enterocoliticum” and Pasteurella X), p. 103–104. In Proceedings of the 14th Scandinavian
Congress on Pathology and Microbiology, Oslo, Norway (1964).
• Fredriksson- Ahoma M., Hielm S., Korkeala H. Hig prevalence of yadA-positive Yersinia
enterocolitica in pigs tounge s and miced meat at retail level in Finland. Journal of food
120

Protection 62: 123-127 (2000).
• Fredriksson-Ahomaa M., Korte T. , Korkeala H. Trasmission of yersinia enterocolitica 4/O:3
to pets via contamineted pork. Letters in Applied Microbiology 32: 375-378 (2001).
• Fredriksson-Ahomaa M., Meyer C., Bonke R., Stüber E., Wacheck S. Characterization
of Yersinia enterocolitica 4/O: 3 isolates from tonsils of Bavarian slaughter pigs. Letters in
Applied Microbiology Vol. 50: 412-418 (2010).
• Fredriksson-Ahomaa M., Niskanen T., Bucher M., Korte T., Stolle A., Korkeala H. Different
Yersinia enterocolitica 4/O: 3 genotypes found in pig tonsils in Southern Germany and
Finland. Syst.Appl.Microbiol. 26: 132-137 (2003).
• Fredriksson-Ahomaa M., Stolle A., Siitonen A., Korkeala H. Sporadic human Yersinia
enterocolitica infections caused by bioserotype 4/O:3 originate mainly from pigs. Journal of
Medical Microbiology. 55:747-749 (2006).
• Fukushima H., Hoshina K., Itogawa H., Gomyoda M. Introduction into Japan of pathogenic
Yersinia Through imported pork, beef and fowl. Int. Journal of Food Microbiology 35:
205-212 (1997).
• Galindo C.L, Rosenzweig J.A., Kirtley M.L., Chopra A.K. Pathogenesis of Y.enterocolitica
and Y. pseudotuberculosis in human yersiniosis. Journal of Pathogens (2011).
• Gehringer G. Multiplication of bacteria during farm storage. In. Factors inflencing the
bacteriological quality of raw milk. International Dairy Federation Bulletin, Document 120
(1980).
• Gemski P., Lazere J.R., Casey T. Plasmid associated with pathogenicity and calcium
dependency of Yersinia enterocolitica. Infect. Immun., 17: 682- 685 (1980).
• Gill C.O., Penney N. and Nottingham P.M. Effect of delayed evisceration on the microbical
quality of meat. Applied and Environmental Microbiology. 31: 465 – 468 (1976).
121

• Gill C.O., Penney N. and Nottingham P.M. Tissue sterility in uneviscerated carcasses.
Applied and Environmental Microbiology. 36: 356 – 359 (1978).
• Grahek-Ogden D., Schimmer B., Cudjoe K.S., Nygard K., Kapperud G., Outbreak of
Yersinia enterocolitica serogroup O:9 infection and processed pork, Norway. Emerg. Infect.
Dis. 13: 754-756 (2007).
• Grant T., Bennet-Wood V., Robins-Browne R.M. Characterization of the interaction between
Yersinia enterocolitica biotype 1A and phagocytes and epithelial cells in vitro. Infect.
Immun. 67: 4367-4375 (1999).
• Grant T., Bennett-Wood V., Robins-Browne R.M. Identification of virulence- associated
characteristics in clinical isolated of Yersinia enterocolitica lacking classical virulence
markers. Infect. Immun. 66: 1113-1120 (1998).
• Gyles L.C., Prescott J.F., Songer G.J., Theen C.O. Pathogenesis of bacterial infections in
animals. Blackwell Publishig (2004).
• Hamama A., Marrakchi A., Othamani F. Occurrence of Yersinia enterocolitica in milk anch
dairy product in Morocco. Int. J. food Microbiol., 16 (1): 69-77 (1992).
• Haoxuan Z., Yong S., Zhengguo M., Bo J. Investigation of virulence genes in clinical
isolates of Yersinia enterocolitica. FEMS Immunol. Med. Microbiol. 53: 368-374 (2008).
• Holm C., Jepsen L., Larsen M., Jespersen L. Predominant microflora of downgraded
Danish bulk tank milk. Journal of Dairy Science, 87: 1151-1157 (2004).
• http://wpage.unina.it/villani/eLAB6.html.
• http://www.antropozoonosi.it/Malattie/Yersiniosi/Yersiniosi.php
• http://www.chimica-online.it/download/calore/conservazione_calore.htm.
• Hudson J.A., Mott S.J. Growth of Listeria monocytogenes, Aeromonas hydrophila and
122

Yersinia enterocolitica on cooked beef under refrigeration and mild temperature abuse.
Food Microbiol. 10: 429-437 (1993).
• Huffman R.D. Current and future technologies for the decontamination of carcasses and
fresh meat. Meat Science. 62: 285-294 (2002).
• Hughes D. Isolation of Yersinia entercolitica from milk and dairy farm in Australia. J. Appl.
Bacterial 46:125-130 (1979).
• Ibrahim A., Rae I.C. Mac isolation of Yersinia enterocolitica and related species from red
meat and milk. J. Fodd Sci., 56 (6): 1524-1525 ( 1991).
• Igiene e sicurezza del latte bovino alla stalla: sistema integrato di diagnosi. Quaderni di
Ricerca n°: 119-Ottobre. Lombardia (2010).
• ISO 10273: 2003. Microbiology of Food and Animal feeding stuffs- Horizontal Method for
the detection of presumptive pathogenic Yersinia enterocolitica.
• ISO 16654: 2001. Microbiology of food and animal feeding stuffs-horizontal method for
detection of E.coli O:157.
• ISO 6579: 2002 /cor 1 1: 2004. Microbiology of food and animale feeding stuffs-horizontal
method for the detection of Salmonella spp.
• ISO 6888-1:1999. Microbiology of food and animal feeding stuffs-horizontal method for the
enumeration of coagulase-positive staphylococci (Staphylococcu aureus and other species).
• IZSLER – Istituto Zooprofilattico Sperimentale della Lombardia e dell' Emilia Romagna.
Dati analitici qualità del latte. Programma di pagamento del latte in base alla qualità (2010).
• Jay J.J., Loessner M.J., Golden A.D. Microbiologia degli alimenti (2009).
• Kaminska S., Sadkowska-todys M. Yersiniosi in Poland in 2012. PRZEGL epidemiol. 68:
235 – 238 (2014).
123

• Kapperud G., Namork E., Skurnuk M., Nesbakken T. Plasmid-mediated surface fibrillae of
Yersinia pseudotubercolosis and yersinia enterocolitica: relationship to outer membrane
protein YOP1 and possible importance for pathogenesis. Infect. Immun., 55: 2247- 2254
(1987).
• Kelly P.T., O'Sullivan K.,Berry D.P., More S.J., Meany W.J., O'Callaghan E.J., O'Brien B.,
Farma management factors associated with bulk tank total bacterial coun t in Irish dairy
herds during 2006/2007. Irish Veterinary Journal, 62:36-42 (2009).
• Kontiainen S., Sivonen A., Renkonen O.V. Increased yields of pathogenic Yersinia
enterocolitica strains by cold enrichment. Scandinavia Journal of infections disease 26:
685- 691 (1994).
• Kurweii R., Busse M. Total count and microflora of freshly drawn milk. Milchwisenschaft
28: 427 (1973).
• Lahuerta A., Westrell T., Takkien J., Boelaert F., Rizzi V., Helwigh B. et al. Zoonoses in the
European Union: Origin, distribution and dynamics. The EFSA-ECDC summary report
2009. Euro Surveill 16: 1-4 (2011).
• Larkin L.L, et al. Incidence of Yersinia enterocolitica in raw milk as related its quality.
Milchwissenschaft. 46: 500-502 (1991).
• Lian C.J, Hwang W.S., Kelly J.K., Pai C.H. Invasiviness of Yersinia enterocolitica lacking
the virulence plasmid: an in vivo-study. J.Med.Microbiol. 24: 219-226 (1987).
• Lodi R., Brasca M. latte crudo: pro e contro. Un contributo al dibattito. Sci.
dell'alimentazione rev. n°3 anno 38 (2009).
• Logue C.M., Sheridan J.J., Wauters G., Mc Dowell D.A., Blair I.S. Yersinia spp. And
numbers, with particular reference to Yersinia enterocolitica bio/serotypes, occurring on
Irish meat and meat products, and the influence of alkali treatment on their isolation.
124

International Journal of Food Microbiology 33: 257-274 (1996).
• Martin L., Cabanel N., Lesoille C., Mènard T., investigation of an unusual increase in
human yersiniosis in Creuse, France. International juornal of Infectious Diseases. 34: 76-78
(2015).
• McEvoy J.M., Doherty A.M., Sheridan J.J., Thomson-Cartier F.M., Garvey P., McGuire L.,
Blair I.S. and McDowell D.A. The prevalence and spread of Escherichia coli O157:H7 at a
commercial beef abattoir. Journal of Applied Microbiology. 95: 256-266 (2003b).
• McEvoy, J. M., Doherty A. M., Finnerty M., Sheridan J. J., McGuire L., Blair I. S.,
MacDowell D. A., and Harrington D. The relationship between hide cleanliness and
bacterial numbers on beef carcasses at a commercial abattoir. Letters in Applied
Microbiology. 30: 390-395 (2000a.).
• McIver, M. A., and R. M. Pike. Chronic glanders-like infection of face caused by an
organism resembling Flavobacterium pseudomallei Whitmore. Clinical miscellany, vol. 1.
Mary Imogene Basset Hospital, Cooperstown, N.Y. p. 16–21 (1934).
• Miller V.L., Falkow S. Evidence for two genetic loci in Yersinia enterocolitica that can
promote invasion of epithelial cells. Immun., 56: 1242- 1248 (1988).
• Mucchetti G., Nevaini E. Microbiologia lattiero-casearia. Ed. Tecniche Nuove, Milano
(2006).
• Murphy S.C., Boor K.J. Sources and causes of high bacteria counts in raw milk: an
abbreviated review (2000).
• Nastaijevic I., Mitrovic R., Buncic S. Occurrence of Escherichia coli O157 on hides of
slaughtered cattle. Letters in Applied Microbiology. 46: 126-131 (2008).
• Nesbakken T., Eckner K., Hoidal H.K., Rotterud O.J. Occurence of Yersinia enterocolitica
and campylobacter spp. In slaughter pigs and consequences for meat inspection, slaugheting
125

and dressing procedures. International Journal of Food Microbiology, 80: 231-240 (2003).
• NORMA AFNOR 3M 01/1- 09/89- 3M (TM) PETRIFILM (TM) AEROBIC COUNT
PLATE.
• NORMA AOAC 991.14 2002. Official method coliform and E.coli counts in foods.
• Notiziario ERSA 2/2006. Zanchetta S. Qualità microbiologica del latte e difetti del
formaggio.
• Paerregaard A., espersen F., jensen O.M., Silurnik M. Interctions between Yersinia
enterocolitica and rabbit ileal mucus: growth, adhesion, penetration and susequent changes
in surface hydrophobicity and ability to adhere to ileal brush border membrane vesicles.
Infect.Immun., 59: 253- 260 (1991).
• Pai C.H., Mors V. Production of enterotoxin by Yersinia enterocolitica. Infect. Immun., 19:
908- 911 (1978).
• Paixao R., et al. Characterization of Yersinia enterocolitca Biotype 1A Strains isolated from
Swine Slaughterhouse and Markets. The Scientific World Journal Volume 2013 (2012).
• Peruzy M.F. Moleculare characterization of different virulence genes of Yersinia
enterocolitica by using SYBR Green real-time PCR. Società italiana di Diagnostica di
Laboratorio Veterinaria (2014).
• Platt-Samoraj A., Ugorski M., Szcreba A., Wojciech K., Proca jlo Z. Analysis of the
presence of ail, yst A and yst B genes in Yersinia enterocolitica strains isolated from aborting
sows and aborted fetuses. J.Vet: Med B53: 341-346 (2006).
• Portnoy D.A., Falkow S. Virulence- associated plasmid from Yersinia enterocolitica and
Yersinia pestis. J. Bacteriol., 148: 877 – 883 (1981).
• Portnoy D.A., Martinez R.J. Role of plasmids in the pathogenicity of Yersina species.
Curr.Topics Microbiol. Immunol. 118: 29-51 (1985).
126

• Portnoy D.A., Wolf-watz H., Bolin I., Beeder A.B., Falkow S. Characterization of common
virulenc e plasmids in yersinia species and their role in the expression of outer membrane
proteins. Infect. Immun., 43: 108- 114 (1984).
• Quaderni di zooprofilassi- entrobatteri patogeni: rapporto regionale della sorveglianza di
laboratorio (2012) N° 12 Agosto (2014).
• Rahuma N., Ghenghesh K.S., Aissa R.B., Elamaari A. Carriage by the housefly (Musca
Domestica) of multiple antibiotic-resistant bacteria that are potentially pathogenic to humans
, in hospital and other urban environments in Misurata, Libya. Annual of Tropical Medicine
and Parasitology vol: 99, n 8: 795-802 ( 2005).
• Regolamento (CE) N. 853 /2004 Del parlamento Europeo e del Cosiglio del 29 Aprile 2004.
• Reinemann D.J., Mein G.A., Bray D.R., Reid D., Britt J.s. Troubleshooting high bacteria
counts in farm milk. http:/www.uwex.edu/MilkQuality/PDF/A3705.pdf (1997).
• Reinemann D.J., Ruegg P. An investigation of ATP bioluminescence and quantitaive
bulktank cultures to assess cleanliness of milking machines. ASAE annual International
Meeting Milwaukee, Wisconsin, USA July 10-13 (2000).
• Robin-Browne R.M. Yersinia enterocolitica. In food Microbiology, Fundamentals and
Frontiers ed.Doyle M.P., Beuchat I.R. and Montville T.J. pp 192-215. Washington DC: ASM
Press. (1997).
• Robins-Browne R.M., Miliotis M.D., Cianciosi S., Miller V.L., Falkow S., Morris Jr. J.G.
Evaluation of DNA clony hybridization and other techniques for detection of virulence in
Yersinia species. Journal of Clinical Microbiology, vol. 27, n 4: 644-650 (1989).
• Roccato A., Barco L., Lattini A.A., De Pasquale F., Ramon E., Dalla Pozza M.C., Sillo S.,
Ricci A. Isolamento e caratterizzazione di Yersinia enterocolitica da vegetali e alimenti di
origine suina. Sidilv (2015).
127

• Rochbach B.W., Draughon F.A., Davison P.M., Oliver S.P. Prevalence of Listeria
monocytogenes, Campylobacter jejuni, Yersinia enterocolitica and Salmonella in bulk tank
milk: risk factors and risk for human exposure. J.Food.Prot. 55: 93-97 (1992).
• Rosset R. Etat des animaux avant abattage. Hygiène et technologie de la viande fraîche.
Edition du CNRS. (1982)
• Rosset R. et Liger R. Nature des porteurs de germes. Hygiène et technologie de la viande
fraîche. Edition du CNRS. (1982).
• Schaake J., Drees A., Gruning P., et al. Essential role of invasin for colonization and
persistance of Yersinia enterocolitica in its natural reservoir host, the pig. Infection and
Immunity Vol. 82: 960-969 (2014).
• Schiemann D.A. Association of Yersinia enterocolitica with the manufacture of cheese and
occurrence in pasteurizzed milk. Appll. and Enviromental Microbiology Vol 36, N°2:
274-277 (1978).
• Schiemann D.A., Toma S. Isolation of Yersinia enterocolitca from raw milk. Appl.Environ.
Microbiol 35:54-58 (1978).
• Schleifstein, J., and M. B. Coleman. An unidentified microorganism resembling B. lignieri
and Past. pseudotuberculosis and pathogenic for man. N. Y. State J. Med. 39:1749–1753
(1939).
• Shanmugapriya S., Senthilmurugan T., Thayumanavan. Genetic diversity among Yersinia
enterocolitica isolated from chicken and fish in and around Coimbatore City, India. Iranian
J.Pub Health. 43: 835-844 (2014).
• Singh I., Virdi J.S. Production of Yersinia stable toxin (YST) and distribuition of yst genes
in biotype 1A strains of Yersinia enterocolitica. Journal of Medical Microbiology, 53:
128

1065-1068 (2004).
• Skurnik M. Expression of antigens encoded by virulence plasmid of Yersinia enterocolitica
under different growth conditions. Infect. Immun., 47: 183 – 190 (1985).
• Sofos J.N., Belk K.E., and Smith G.C. Processed to reduce contamination with pathogenic
microorganisms in meat. Con. Meat. Sci. Tech. 45: 596-605 (1999).
• StefanelliS., Ceccarelli M., Paladini A. Isolamento di Yersinia enterocolitica in pazienti con
enterite acuta a Pisa. Ann. Ist. Super. Sanità Vol.22, n° 3: 997-1000 (1986).
• Stern N. J., Pierson M. D., Katula A.W. Growth and competitive nature of Yersinia
enterocolitica in whole milk. J. Food. Sci. Vol. 45: 972-974 (1980).
• Swaminathan B., Harmon M.C., Mehlman I.J. A review: Yersinia enterocolitica. J. App.
Bacter. Vol. 52: 151-183 (1982).
• Tacket C.O., Narain J.P., Sattin R., et al . A multistate outbreak of infections caused by
Yersinia enterocolitica trasmitted by psteurized milk. J. Americ. Med. Assoc. 251: 483-486
(1984).
• Tassinari A.R, Franco B.D., Landgraf M. Incidence of Yersinia spp. In food in Sao Paulo,
Brazil. Int.J. Food Microbiology, 21(3): 263-270 (1994).
• Thibodeau V., Frost E. H., Chénier S., Quessy S. Presence of Yersinia enterocolitica in
tissue of orally inoculed pigs and the tonsils and feces of pigs at slaughter. Can. J. Vet. Res.
63: 96-100 (1999).
• Thoener P., Bin Kingombe C.I., Bogli-Stuber K., et all.PCR detection of virulence genes in
Yersinia enterocolitica and Yersinia pseudotubercolosis and investigation of virulence gene
distribution. Applied and Enviromental Microbiology Vol. 69 n°3: 1810-1816 ( 2003).
• Toora S., Budu-Amoako E., Ablett R.F., Smith J. Effect of high-temperature short-time
pasteurization, freezing and thawing and costant freezing, on the survival of Yersinia
129

enterocolitica in milk. J. Food Protect. 55: 803-805 (1992).
• Tortora G.L., Funke B.R., Case C.L. Elementi di microbiologia, pp 179 Pearson Education
Italia (2008).
• UNI EN ISO 10272- 1: 2006. Microbiology of food and animal feeding stuffs-horizontal
method for detection and enumeration of Campylobacter spp.
• UNI EN ISO 11290- 1: 2005. Metodo orizzontale per la ricerca e la conta di Listeria
monocytogenes.
• United State Department of Agriculture (USDA). Guidance for Minimizing the Risk of
Escherichia coli O157:H7 and Salmonella in Beef Slaughter Operations. National
Agricoltural Library/USDA. September (2002).
• USDA. USDA/FSIS-MLG Appendix 2.03 Laboratory Guidebook. Most Probable Number
and Tables, 01/28/08. QD-F-Micro- 0004.03. Disponibile al sito: www.
fsis.usda.gov/wps/wcm/connect/8872ec11- d 6 a 3 - 4 f c f - 8 6 d f - 4 d 8 7 e 5 7 7 8 0 f 5 /
MLG_Appendix_2_03.pdf?MOD=AJPERES (2008).
• Vanantwerpen G, Van Damme I, De Zutter L, Houf K.Seroprevalence of enteropathogenic
Yersinia spp. in pig batches at slaughter. Prev Vet Med. (2014).
• Walker S.J., Gilmour A. The incidence of Yersinia enterocolitica and Yersinia entercolitica
-like organisms in raw and pasteurized milk in Northern Ireland. J.Appl. Bacterial 61:
133-138 (1986).
• World Health Organization (WHO). Risk profile for enterohemorrhagic E.coli including the
identification of the commodities of concern, including sprouts, ground beef, and pork.
Codex Alimentarius. ftp://ftp.fao.org/codex/ccfh35/fh0305de.pdf (Ultimo accesso novembre
2008).
• www.mednat.org/alimentazione/fermentazione_cibi_e_bevande.pdf. Microbiologia degli
130

alimenti
• Yanagawa Y., Maruyama T., Sakai S., Isolation of Yersinia enterocolitica and Yersinia
pseudotubercolosis from apparently healthy dogs and cats. Microbiology and immunology
22: 643-646 (1978).
• Yucel N., Ulusoy. H. A Turkey survey of hygiene indicator bacteria and Yersinia
enterocolitica in raw milk and cheese samples. Food control 17: 383-388 (2006).
131