wetland habitat patches as ecological components of landscape memory in a highly modified floodplain
TRANSCRIPT

RIVER RESEARCH AND APPLICATIONS
River Res. Applic. 30: 874–886 (2014)
Published online 9 July 2013 in Wiley Online Library(wileyonlinelibrary.com) DOI: 10.1002/rra.2685
WETLAND HABITAT PATCHES AS ECOLOGICAL COMPONENTS OF LANDSCAPEMEMORY IN A HIGHLY MODIFIED FLOODPLAIN
A. ORTMANN-AJKAIa*, D. LÓCZYb, P. GYENIZSEc AND E. PIRKHOFFERd
a Department of Ecology and Hydrobiology, Institute of Environmental Sciences, University of Pécs, Pécs, Hungaryb Department of Environmental Geography and Landscape Conservation, Institute of Environmental Sciences, University of Pécs, Pécs, Hungary
c Department of Cartography and Geoinformatics, Institute of Geography, University of Pécs, Pécs, Hungaryd Department of Climatology and Soil Science, Institute of Environmental Sciences, University of Pécs, Pécs, Hungary
ABSTRACT
Landscape memory and ecological memory are relatively new concepts developed independently in geomorphology and ecology.Eco-geomorphology strives to enrich the concept of landscape memory with that of ecological memory, as ecosystems build inherent partsof landscapes and are controlled by geomorphological factors. This integration is illustrated by a small-scale eco-geomorphological casestudy of links between ecosystems and their hydrogeomorphological template, in the floodplain of a medium size Hungarian River. Despitethat the river is channelized by 1835 and most of the study area is under intensive agricultural cultivation, the seminatural vegetation along thevarious reaches shows significant variations according to geomorphological background variables: detrended correspondence analysis yieldsresults of p= 0.002 significance. Our results prove that geomorphological patterns in floodplains, defined by landscape memory, and therelated vegetation patterns, as manifestations of ecological memory, jointly determine the face of floodplain landscapes. The integration ofecological memory into the definition of landscape memory may help meet the challenges of eco-geomorphology and facilitate a holisticunderstanding of landscapes, and thus be instrumental for reconstructing past environments—both in theory (scientific research) and practice(rehabilitation projects). Copyright © 2013 John Wiley & Sons, Ltd.
key words: ecological memory; landscape memory; eco-geomorphology; floodplain vegetation; geomorphometric indices; DCCA; wetland reconstruction
Received 19 February 2013; Revised 9 May 2013; Accepted 24 May 2013
INTRODUCTION
River floodplains are well-known biodiversity hotspots(Bornette et al., 1998; Ward et al., 2002). High biodiversityis shaped and influenced by geomorphology throughcontrolling ecosystem processes (Renschler et al., 2007).Hydrogeomorphological patches (Thorp et al., 2006) areassociated with catchment and valley geomorphology andflow characteristics, which can be studied and describedwith concepts and techniques of physical geography, forexample catchment properties (Church, 2002; Orr et al.,2008), valley parameters (Lóczy et al., 2011) under naturalconditions and also human activities, for examplechannelization, dams or land use (Apan et al., 2002; Allan,2004; Nakamura et al., 2008). In turn, riverbank andfloodplain geomorphology is also shaped by plants, asdiscussed in details in publications (e.g. Corenblit et al.,2007, 2009; Gurnell et al., 2012).Brierley's (2010) landscape memory concept—‘how
imprints from the past influence the way landscapes look
*Correspondence to: A. Ortmann-Ajkai, Department of Ecology andHydrobiology, Institute of Environmental Sciences, University of Pécs,Pécs, Hungary.E-mail: [email protected]
Copyright © 2013 John Wiley & Sons, Ltd.
and operate’—provides a framework to organize all thesefactors into three thematic and time scale domains:geologic memory, operating over millennia, climatic mem-ory, operating over thousands to hundreds of thousands ofyears and anthropogenic memory, operating over tens orhundreds of years.The floodplains of most large and medium size rivers in
Europe are regulated. Abandoned meanders in the protectedpart of the floodplain, clearly visible on satellite images, aretypical ‘imprints of the past’ (Marston et al., 1995; Apanet al., 2002). Because of their better water supply, they oftenharbour wetland ecosystems, habitats with characteristicaquatic and semiaquatic vegetation—even centuries afterchannelization. These ecosystems are influenced indirectly,through the physical hydrogeomorphic template (landscapememory—Thorp et al., 2006), and at the same time, asecological entities (ecological memory).
The concept of ecological memory
Although ecosystem memory has ever been presentimplicitly in ecology (e.g. for relict species, seed banksand lately in historical ecology), the first exact wordingis by Padisák (1992): the ecological memory of a

WETLAND HABITAT PATCHES AND LANDSCAPE MEMORY 875
phytoplankton community, analogous to the seed bankconcept in botany, is ‘the capacity of past states orexperiences to influence present or future responses ofthe community’.The now widely accepted concept of ecological memory
was formulated a decade ago. According to the firstdefinition (Thompson et al., 2001), ecological memory is‘the result of past environmental conditions and subsequentselection of populations, encoded in the current structure ofbiological communities and reflected in the genetic structureof species. It affects how communities are assembled, and itmay also affect the likelihood that they can be restored’.Peterson's definition (2002) formulates the strong
interdependence between ecological processes and landscapedynamics: ecological memory is ‘the degree to which anecological process is shaped by its past modifications of alandscape’. The importance of vegetation patterns is alsopointed out by him, claiming that ecological memory isencoded in the pattern of vegetation across the landscape.As wetland ecosystems have a history not long enough
to be studied on an evolutionary timescale, Bengtsson's def-inition (Bengtsson et al., 2003) is more operative for us touse. According to this definition, the ecological memory ofan (eco)system is ‘the network of species, their dynamicinteractions between each other and the environment, andthe combination of structures that make reorganization afterdisturbance possible’. It can be divided into internal andexternal components (Schaefer, 2009). Internal or within-patch memory consists of ‘biological legacies’, representedby species presence and their within-patch dynamics: forexample their presence, reproduction and interactions, suchas competition abilities, trophic relations (Bengtsson et al.,2003) and genetic composition (Schaefer, 2009), arediscussed in details in ecological literature. External memoryis defined by between-patch dynamics (Bengtsson et al.,2003), first of all sources and colonization possibilities ofspecies, determined by spatial configuration of the landscape.Although termed as ‘landscape memory’, Gillarova
et al.'s (2008) definition is closer to the ecological memoryconcepts mentioned previously: ‘memory structures repre-sent continuous landscape segments containing a varyingscope of original ecosystems that have been preservedsince historical period of the development of the landscapeby now’.Both common components of the previous definitions are
worth noting: the strong link with landscape ecology andconservational importance. Interdependence oflandscapeand ecological processes is the key idea in Peterson's defini-tion. The link to landscape ecology is also explicit inGillarova's definition, speaking of preserved landscapefragments. Thompson's ‘past environmental conditions’refer to Brierley's landscape history imprints. Bengtsson'sexternal memory, defined by between-patch dynamics
Copyright © 2013 John Wiley & Sons, Ltd.
(Bengtsson et al., 2003), first of all sources and colonizationpossibilities of species (species/ecological flows), was deter-mined by the spatial configuration of the landscape, whichbelongs to spatially realistic metapopulation ecology (Hanski,2001), the first among the top 10 research priorities oflandscape ecology (Wu and Hobbs, 2002). Stallins (2006)even considers ecological memory—besides multiple causal-ity, ecosystem engineering and ecological topology—as oneof the four overlapping themes linking geomorphology andecology (Stallins, 2006), offering an operational solution forthe challenge of eco-geomorphology (Thoms and Parsons,2002) and eco-hydromorphology (Vaughan et al., 2009):the synthesis of hydrogeomorphology, ecology and land-scape ecology.The importance for nature conservation is also expressed
by several authors. The guiding principle of Bengtsson'sbaseline article is biodiversity conservation, with specialrespect to human-dominated landscapes, where ecologicalnetworks gain more importance. Ecological memory, as aform of spatial resilience, is considered to be a prerequisiteof ecosystem reorganization (Bengtsson et al., 2003;Gunderson, 2000), reconstruction and rehabilitation.Peterson (2002) states that ecological memory is
encoded in the pattern of vegetation across the landscape.Considering the conservational importance of seminaturalvegetation patches in highly anthropogenic floodplains,and the increasing number of wetland reconstruction pro-cesses, it is also worth considering them in the ecologicalmemory context.Wetland vegetation patches encompass ecological mem-
ory as defined by Bengtsson: refugia for wetland speciestogether with their ecological interaction networks (internalcomponent) and their spatial structure (e.g. alignments asstepping stones), determined by hydrogeomorphic factors,make interpatch species movements possible (external com-ponent). What makes (semi-) natural wetland habitats anespecially promising subject for the linked study of ecolog-ical and landscape memory is that their spatial configurationis strongly determined by geomorphological features, so thegeomorphology–ecology link may be very strong here.Floodplains are usually recent formations, so their history—both before and after regulation—may be well studied.for example from historical maps or remote-sensed images(e.g. Lóczy et al., 2011). This imprint from the past can beregarded as landscape memory (Brierley, 2010)—a uniqueopportunity to link the concepts of landscape memory andecological memory.Our study provides empirical, field-based evidence. Its
objectives are as follows: (1) to characterize geomorphicreaches with vegetation types and identify the geomorpho-logical controls of vegetation types and relate them toreaches; (2) to demonstrate the strength of dependence ofecological memory (encoded in vegetation patches) on
River Res. Applic. 30: 874–886 (2014)
DOI: 10.1002/rra

A. ORTMANN-AJKAI ET AL.876
quantitative hydrogeomorphological variables; (3) to proposean extension of Brierley's ‘landscape memory’ concept toincorporate ‘ecological memory’; and (4) to discuss its impli-cations for nature conservation.To this end, a quantitative description of the patch pattern
of vegetation is presented, followed by a multivariateanalysis of the linkages of these ecological memory-codingobjects with abiotic (hydrogeomorphological) variables onthe example of the floodplain of a medium size river ofSouthwest Hungary, the Kapos, using a detailed geomorpho-logical typology of floodplain reaches (Lóczy et al., 2011)and vegetation types obtained from the Landscape Ecologi-cal Database of Hungarian Habitats (MÉTA) (Molnáret al., 2007).
MATERIAL AND METHODS
Study area
The medium size catchment of the Kapos River covers3295.4 km2 in the Outer Somogy Hills in SouthwestHungary (Lóczy, 2013) (Figure 1). The trunk river is112.7 km long, a fifth-order stream at confluence to the SióCanal. The morphological floodplain has an area of104.2 km2 (3.3% of the catchment area). Its mean discharge(calculated for the period 1995–2005) at a gauge (see onFigure 1) is 6.19m3 s�1, whereas flood discharge is42.4m3 s�1 on the average, and the highest on record is174m3 s�1. The retention of floodwater is restricted to a lessthan 40-m wide active floodplain and groundwater storageon the protected floodplain. As a consequence, floodplaininundation is rather common. The present study concerns
Figure 1. Location of study area (Upper Kapos River) in Southwest Hun
Copyright © 2013 John Wiley & Sons, Ltd.
the uppermost 50-km reach, extending from the spring to theconfluence of the Baranya Canal, near the town Dombóvár.Most important geologic, climatic and anthropogenic
factors, shaping this landscape on different spatial scales,are summarized in Table I.The geological substrate is a Quaternary loess mantle, in
valleys up to 15m, on hills up to 25m in thickness, directlyoverlying Upper Miocene marine and lacustrine clays andsands (Szilárd, 1967; Magyari et al., 2005; Csontos et al.,2005). The Kapos catchment shows remarkable asymmetries:typical slopes on the northern valley side with Pleistoceneterraces are 2.5° to 3.5° and only exceptionally exceed 15°,whereas southern slopes of 15° to 25° also occur.Climate in the Quaternary changed from glacial to warmer
and dry continental, then a more humid submediterranean-type climatic optimum, getting cooler, then dryer up topresent. The predominant climate types today are subatlanticwith a mean annual temperature slightly above 10 °C and anannual precipitation of 680 to 720mm. As valley floors usedto be lined with peat bogs, the predominant soil types arevarieties of Histosols on muck to Fluvisols on alluvial silt.Potential natural vegetation, determined purely by
climatic and geologic factors before human colonization,consisted of zonal mesophilous forests (beech, hornbeamand oak forests) in the hills and azonal riparian forests anda wide array of different wetland habitats, open waters,swamps, mires and sedges, in the floodplains (Borhidiet al., 2012). As in this timescale, climatic and geomorphicconditions may be regarded as stable, boundary conditionsof ecological memory, manifested in recent vegetation pat-terns, are determined predominantly by anthropogenicmemory(land use) summarized in Table Ic.
gary
River Res. Applic. 30: 874–886 (2014)
DOI: 10.1002/rra
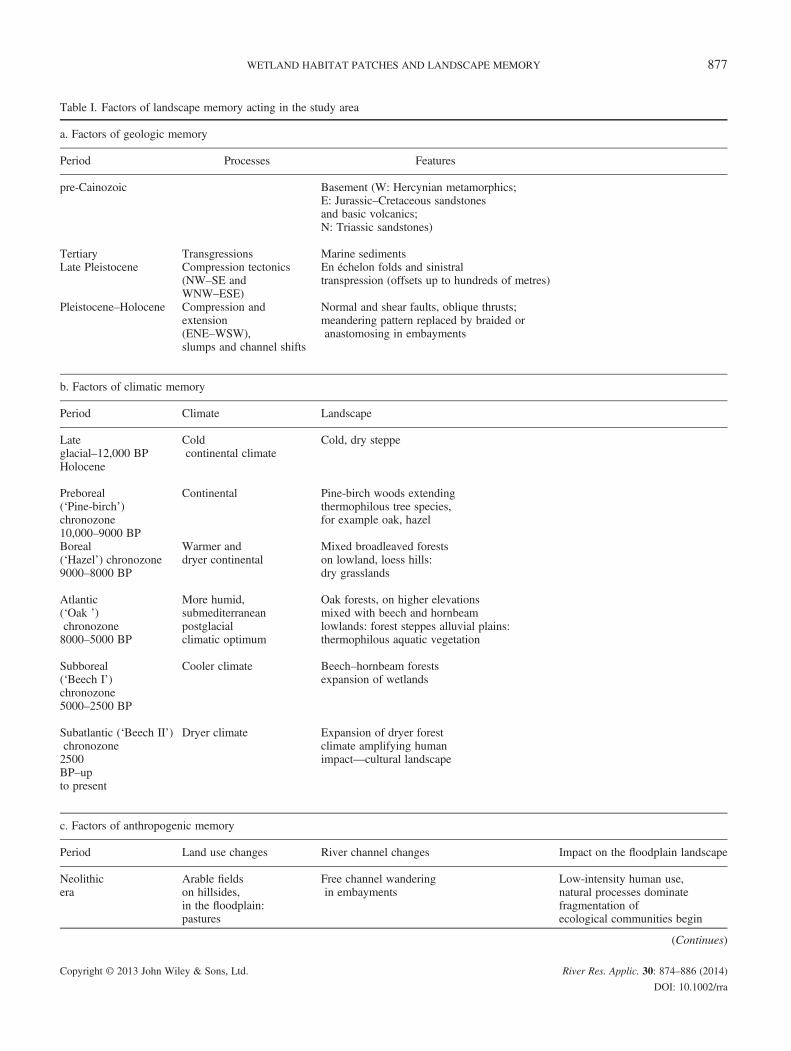
Table I. Factors of landscape memory acting in the study area
a. Factors of geologic memory
Period Processes Features
pre-Cainozoic Basement (W: Hercynian metamorphics;E: Jurassic–Cretaceous sandstonesand basic volcanics;N: Triassic sandstones)
Tertiary Transgressions Marine sedimentsLate Pleistocene Compression tectonics
(NW–SE andWNW–ESE)
En échelon folds and sinistraltranspression (offsets up to hundreds of metres)
Pleistocene–Holocene Compression andextension(ENE–WSW),slumps and channel shifts
Normal and shear faults, oblique thrusts;meandering pattern replaced by braided oranastomosing in embayments
b. Factors of climatic memory
Period Climate Landscape
Lateglacial–12,000 BP
Coldcontinental climate
Cold, dry steppe
Holocene
Preboreal(‘Pine-birch’)chronozone10,000–9000 BP
Continental Pine-birch woods extendingthermophilous tree species,for example oak, hazel
Boreal(‘Hazel’) chronozone9000–8000 BP
Warmer anddryer continental
Mixed broadleaved forestson lowland, loess hills:dry grasslands
Atlantic(‘Oak ’)chronozone8000–5000 BP
More humid,submediterraneanpostglacialclimatic optimum
Oak forests, on higher elevationsmixed with beech and hornbeamlowlands: forest steppes alluvial plains:thermophilous aquatic vegetation
Subboreal(‘Beech I’)chronozone5000–2500 BP
Cooler climate Beech–hornbeam forestsexpansion of wetlands
Subatlantic (‘Beech II’)chronozone2500BP–upto present
Dryer climate Expansion of dryer forestclimate amplifying humanimpact—cultural landscape
c. Factors of anthropogenic memory
Period Land use changes River channel changes Impact on the floodplain landscape
Neolithicera
Arable fieldson hillsides,in the floodplain:pastures
Free channel wanderingin embayments
Low-intensity human use,natural processes dominatefragmentation ofecological communities begin
(Continues)
WETLAND HABITAT PATCHES AND LANDSCAPE MEMORY 877
Copyright © 2013 John Wiley & Sons, Ltd. River Res. Applic. 30: 874–886 (2014)
DOI: 10.1002/rra

Table I. (Continued)
a. Factors of geologic memory
Period Processes Features
Roman era(Province Pannonia)
Intensified deforestation
Mediaeval times Floodplain economy(graze lands,meadows, orchards)established
17–18th century Stockbreedingintensifies, horticulture(fodder andvegetable growing)spreading
Mill canals built parallel with mainchannel; bank reinforcementat settlements
Wetlands maintainedby frequent flooding
19th century Drainage ditches built;drained landconverted tosmall scalearable farming;more intensiveuse of meadows
River channelization, displacement oftributary confluences, land drainagein the floodplain, which isdisconnected from the river channel
Desiccation of peat bogs, fires,alternating with periodsof waterlogging and flooding
1900–1950 Transport lines(roads, new bridgesand railways) built alongfloodplain margin;arable farming intensifying
Raising and reinforcingflood-control dykes,preventing inundations
Increased fragmentation;periods of waterloggingand desiccation alternating
1950–2000 Large-scale arablefarming and poplarplantations introduced
Flood retention reservoirs(used as fish ponds)built along tributary streams;return period of floods: 6–8 years
Human pressure rapidlyintensifying; wetlandsshrinking
2000– Large-scale farmingsurvives, stockbreedingdeclines, lower intensityof pasture andmeadow use
Return period of floods reducingto 3-6 years, uprooted treescause channel deformation(local widening)
Occasional dam breaches,frequent waterloggingprovide sites for the survivalof wetland communities,as representativesof ecological memory;wet meadows degrade
c. Factors of anthropogenic memory
Land use changes River channel changes Impact on the floodplain landscape
A. ORTMANN-AJKAI ET AL.878
Arable fields have been cultivated since the Neolithic era.The regularly flooded floodplains were used as hayfields andpastures. Deforestation intensified in the Roman era, asSouth Transdanubia was part of the Roman Empire (Panno-nia province). For centuries, landscape elements—arablefields, vineyards and forests on the hills; wet meadows,marshes and even fishponds in valley floors—weresustained with proportions changing parallel with long-termpopulation changes (Bándi et al., 1979). As a consequenceof river regulations, completed in the 19th century, thefloodplain desiccated, inundations became less frequentand meadows gained space at the expense of marshes andpresumably turned into dryer degraded ecotypes.
Copyright © 2013 John Wiley & Sons, Ltd.
A detailed geomorphological description of the floodplaincan be found in Lóczy et al. (2011). On the basis ofhydrogeomorphological properties (Table II), four reaches aredistinguished. At the uppermost reach, the Kapos River flowson a narrow floodplain, which becomes partly confined down-stream, and broader floodplain pockets alternate with narrowersections along meandering and locally braided reaches.Meandering-braided reaches accompanied by a floodplain ofrelatively uniform width, divided by a braided section, follow.The uppermost reach is located in a densely forested hilly
area, where original natural vegetation predominates: beech for-ests (Vicio oroboidi –Fagetum) in the hillsides and alder groves(Carici pendulae – Alnetum) along the streams. Further down
River Res. Applic. 30: 874–886 (2014)
DOI: 10.1002/rra
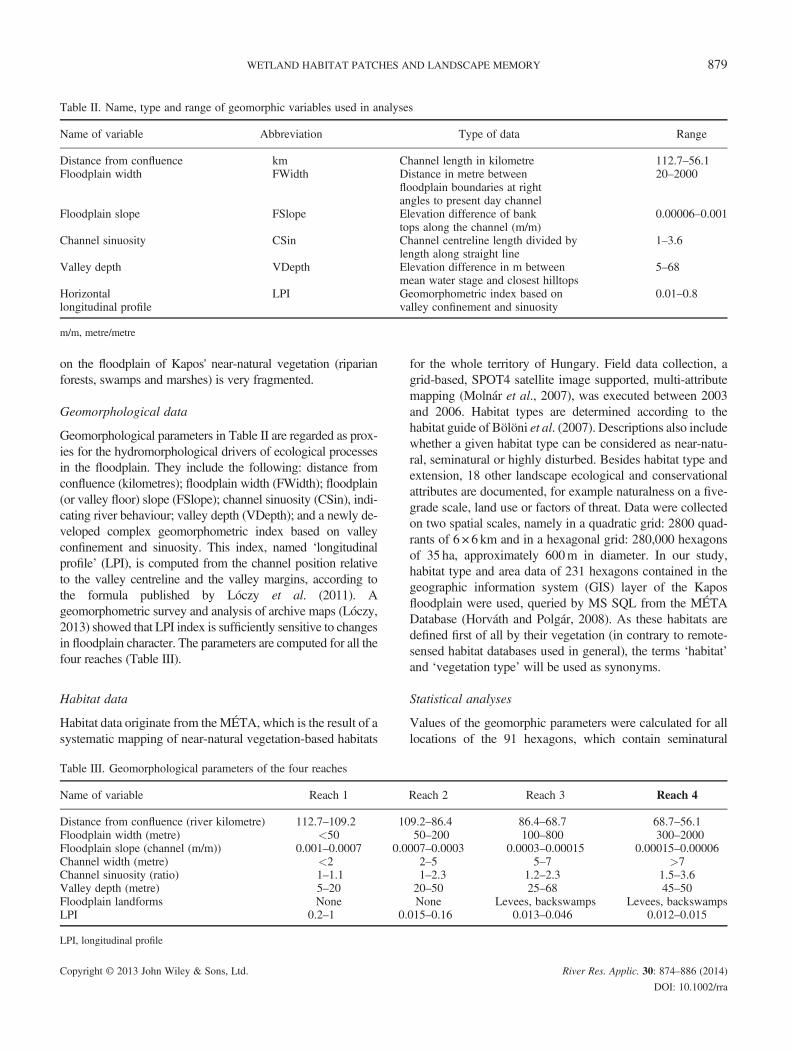
Table II. Name, type and range of geomorphic variables used in analyses
Name of variable Abbreviation Type of data Range
Distance from confluence km Channel length in kilometre 112.7–56.1Floodplain width FWidth Distance in metre between
floodplain boundaries at rightangles to present day channel
20–2000
Floodplain slope FSlope Elevation difference of banktops along the channel (m/m)
0.00006–0.001
Channel sinuosity CSin Channel centreline length divided bylength along straight line
1–3.6
Valley depth VDepth Elevation difference in m betweenmean water stage and closest hilltops
5–68
Horizontallongitudinal profile
LPI Geomorphometric index based onvalley confinement and sinuosity
0.01–0.8
m/m, metre/metre
WETLAND HABITAT PATCHES AND LANDSCAPE MEMORY 879
on the floodplain of Kapos' near-natural vegetation (riparianforests, swamps and marshes) is very fragmented.
Geomorphological data
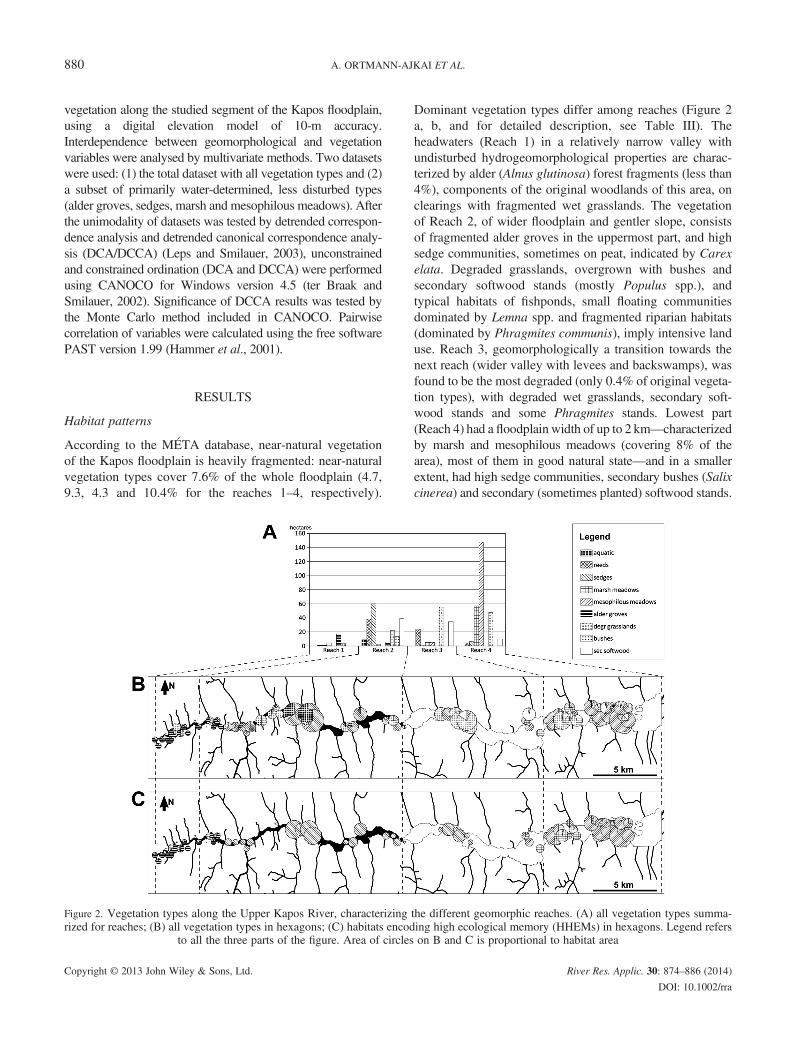
Geomorphological parameters in Table II are regarded as prox-ies for the hydromorphological drivers of ecological processesin the floodplain. They include the following: distance fromconfluence (kilometres); floodplain width (FWidth); floodplain(or valley floor) slope (FSlope); channel sinuosity (CSin), indi-cating river behaviour; valley depth (VDepth); and a newly de-veloped complex geomorphometric index based on valleyconfinement and sinuosity. This index, named ‘longitudinalprofile’ (LPI), is computed from the channel position relativeto the valley centreline and the valley margins, according tothe formula published by Lóczy et al. (2011). Ageomorphometric survey and analysis of archive maps (Lóczy,2013) showed that LPI index is sufficiently sensitive to changesin floodplain character. The parameters are computed for all thefour reaches (Table III).
Habitat data
Habitat data originate from the MÉTA, which is the result of asystematic mapping of near-natural vegetation-based habitats
Table III. Geomorphological parameters of the four reaches
Name of variable Reach 1
Distance from confluence (river kilometre) 112.7–109.2 10Floodplain width (metre) <50Floodplain slope (channel (m/m)) 0.001–0.0007 0.0Channel width (metre) <2Channel sinuosity (ratio) 1–1.1Valley depth (metre) 5–20Floodplain landforms NoneLPI 0.2–1 0.
LPI, longitudinal profile
Copyright © 2013 John Wiley & Sons, Ltd.
for the whole territory of Hungary. Field data collection, agrid-based, SPOT4 satellite image supported, multi-attributemapping (Molnár et al., 2007), was executed between 2003and 2006. Habitat types are determined according to thehabitat guide of Bölöni et al. (2007). Descriptions also includewhether a given habitat type can be considered as near-natu-ral, seminatural or highly disturbed. Besides habitat type andextension, 18 other landscape ecological and conservationalattributes are documented, for example naturalness on a five-grade scale, land use or factors of threat. Data were collectedon two spatial scales, namely in a quadratic grid: 2800 quad-rants of 6 × 6 km and in a hexagonal grid: 280,000 hexagonsof 35 ha, approximately 600m in diameter. In our study,habitat type and area data of 231 hexagons contained in thegeographic information system (GIS) layer of the Kaposfloodplain were used, queried by MS SQL from the MÉTADatabase (Horváth and Polgár, 2008). As these habitats aredefined first of all by their vegetation (in contrary to remote-sensed habitat databases used in general), the terms ‘habitat’and ‘vegetation type’ will be used as synonyms.
Statistical analyses
Values of the geomorphic parameters were calculated for alllocations of the 91 hexagons, which contain seminatural
Reach 2 Reach 3 Reach 4
9.2–86.4 86.4–68.7 68.7–56.150–200 100–800 300–2000
007–0.0003 0.0003–0.00015 0.00015–0.000062–5 5–7 >71–2.3 1.2–2.3 1.5–3.620–50 25–68 45–50None Levees, backswamps Levees, backswamps
015–0.16 0.013–0.046 0.012–0.015
River Res. Applic. 30: 874–886 (2014)
DOI: 10.1002/rra
A. ORTMANN-AJKAI ET AL.880
vegetation along the studied segment of the Kapos floodplain,using a digital elevation model of 10-m accuracy.Interdependence between geomorphological and vegetationvariables were analysed by multivariate methods. Two datasetswere used: (1) the total dataset with all vegetation types and (2)a subset of primarily water-determined, less disturbed types(alder groves, sedges, marsh and mesophilous meadows). Afterthe unimodality of datasets was tested by detrended correspon-dence analysis and detrended canonical correspondence analy-sis (DCA/DCCA) (Leps and Smilauer, 2003), unconstrainedand constrained ordination (DCA and DCCA) were performedusing CANOCO for Windows version 4.5 (ter Braak andSmilauer, 2002). Significance of DCCA results was tested bythe Monte Carlo method included in CANOCO. Pairwisecorrelation of variables were calculated using the free softwarePAST version 1.99 (Hammer et al., 2001).
RESULTS
Habitat patterns
According to the MÉTA database, near-natural vegetationof the Kapos floodplain is heavily fragmented: near-naturalvegetation types cover 7.6% of the whole floodplain (4.7,9.3, 4.3 and 10.4% for the reaches 1–4, respectively).
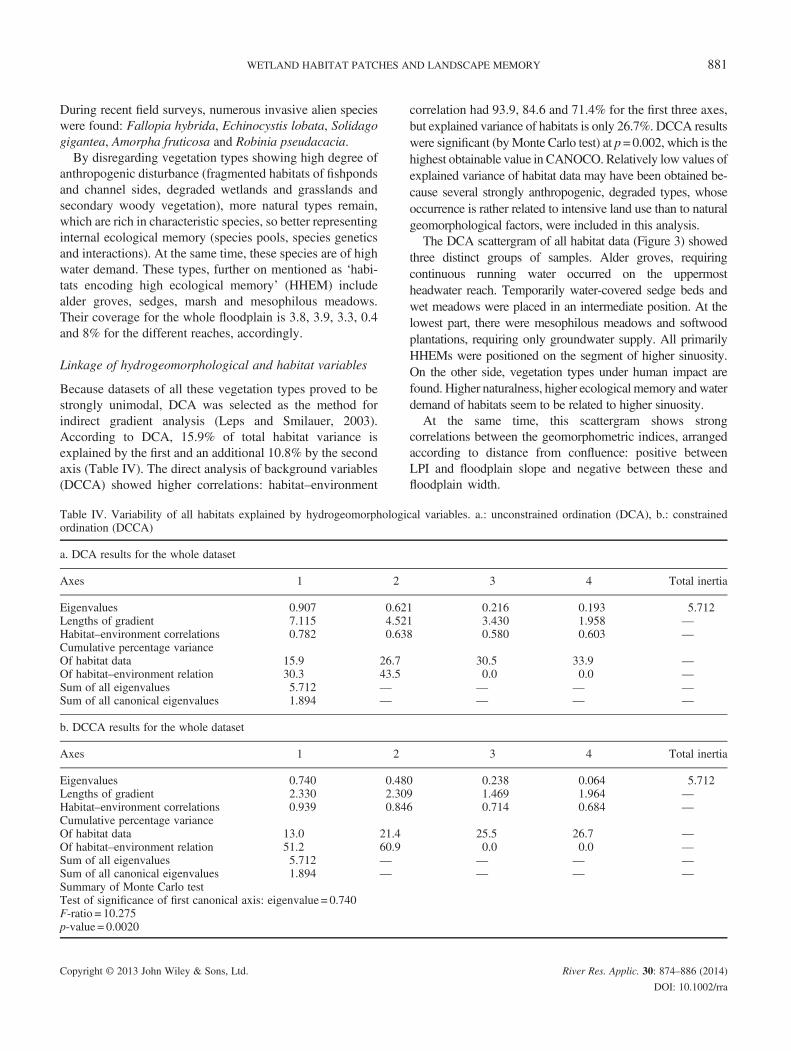
Figure 2. Vegetation types along the Upper Kapos River, characterizingrized for reaches; (B) all vegetation types in hexagons; (C) habitats encod
to all the three parts of the figure. Area of circles
Copyright © 2013 John Wiley & Sons, Ltd.
Dominant vegetation types differ among reaches (Figure 2a, b, and for detailed description, see Table III). Theheadwaters (Reach 1) in a relatively narrow valley withundisturbed hydrogeomorphological properties are charac-terized by alder (Alnus glutinosa) forest fragments (less than4%), components of the original woodlands of this area, onclearings with fragmented wet grasslands. The vegetationof Reach 2, of wider floodplain and gentler slope, consistsof fragmented alder groves in the uppermost part, and highsedge communities, sometimes on peat, indicated by Carexelata. Degraded grasslands, overgrown with bushes andsecondary softwood stands (mostly Populus spp.), andtypical habitats of fishponds, small floating communitiesdominated by Lemna spp. and fragmented riparian habitats(dominated by Phragmites communis), imply intensive landuse. Reach 3, geomorphologically a transition towards thenext reach (wider valley with levees and backswamps), wasfound to be the most degraded (only 0.4% of original vegeta-tion types), with degraded wet grasslands, secondary soft-wood stands and some Phragmites stands. Lowest part(Reach 4) had a floodplain width of up to 2 km—characterizedby marsh and mesophilous meadows (covering 8% of thearea), most of them in good natural state—and in a smallerextent, had high sedge communities, secondary bushes (Salixcinerea) and secondary (sometimes planted) softwood stands.
the different geomorphic reaches. (A) all vegetation types summa-ing high ecological memory (HHEMs) in hexagons. Legend referson B and C is proportional to habitat area
River Res. Applic. 30: 874–886 (2014)
DOI: 10.1002/rra
WETLAND HABITAT PATCHES AND LANDSCAPE MEMORY 881
During recent field surveys, numerous invasive alien specieswere found: Fallopia hybrida, Echinocystis lobata, Solidagogigantea, Amorpha fruticosa and Robinia pseudacacia.By disregarding vegetation types showing high degree of
anthropogenic disturbance (fragmented habitats of fishpondsand channel sides, degraded wetlands and grasslands andsecondary woody vegetation), more natural types remain,which are rich in characteristic species, so better representinginternal ecological memory (species pools, species geneticsand interactions). At the same time, these species are of highwater demand. These types, further on mentioned as ‘habi-tats encoding high ecological memory’ (HHEM) includealder groves, sedges, marsh and mesophilous meadows.Their coverage for the whole floodplain is 3.8, 3.9, 3.3, 0.4and 8% for the different reaches, accordingly.
Linkage of hydrogeomorphological and habitat variables
Because datasets of all these vegetation types proved to bestrongly unimodal, DCA was selected as the method forindirect gradient analysis (Leps and Smilauer, 2003).According to DCA, 15.9% of total habitat variance isexplained by the first and an additional 10.8% by the secondaxis (Table IV). The direct analysis of background variables(DCCA) showed higher correlations: habitat–environment
Table IV. Variability of all habitats explained by hydrogeomorphologiordination (DCCA)
a. DCA results for the whole dataset
Axes 1 2
Eigenvalues 0.907 0.62Lengths of gradient 7.115 4.52Habitat–environment correlations 0.782 0.63Cumulative percentage varianceOf habitat data 15.9 26.7Of habitat–environment relation 30.3 43.5Sum of all eigenvalues 5.712 —Sum of all canonical eigenvalues 1.894 —
b. DCCA results for the whole dataset
Axes 1 2
Eigenvalues 0.740 0.48Lengths of gradient 2.330 2.30Habitat–environment correlations 0.939 0.84Cumulative percentage varianceOf habitat data 13.0 21.4Of habitat–environment relation 51.2 60.9Sum of all eigenvalues 5.712 —Sum of all canonical eigenvalues 1.894 —Summary of Monte Carlo testTest of significance of first canonical axis: eigenvalue = 0.740F-ratio = 10.275p-value = 0.0020
Copyright © 2013 John Wiley & Sons, Ltd.
correlation had 93.9, 84.6 and 71.4% for the first three axes,but explained variance of habitats is only 26.7%. DCCA resultswere significant (byMonte Carlo test) at p=0.002, which is thehighest obtainable value in CANOCO. Relatively low values ofexplained variance of habitat data may have been obtained be-cause several strongly anthropogenic, degraded types, whoseoccurrence is rather related to intensive land use than to naturalgeomorphological factors, were included in this analysis.The DCA scattergram of all habitat data (Figure 3) showed
three distinct groups of samples. Alder groves, requiringcontinuous running water occurred on the uppermostheadwater reach. Temporarily water-covered sedge beds andwet meadows were placed in an intermediate position. At thelowest part, there were mesophilous meadows and softwoodplantations, requiring only groundwater supply. All primarilyHHEMs were positioned on the segment of higher sinuosity.On the other side, vegetation types under human impact arefound. Higher naturalness, higher ecological memory andwaterdemand of habitats seem to be related to higher sinuosity.At the same time, this scattergram shows strong
correlations between the geomorphometric indices, arrangedaccording to distance from confluence: positive betweenLPI and floodplain slope and negative between these andfloodplain width.
cal variables. a.: unconstrained ordination (DCA), b.: constrained
3 4 Total inertia
1 0.216 0.193 5.7121 3.430 1.958 —8 0.580 0.603 —
30.5 33.9 —0.0 0.0 —— — —— — —
3 4 Total inertia
0 0.238 0.064 5.7129 1.469 1.964 —6 0.714 0.684 —
25.5 26.7 —0.0 0.0 —— — —— — —
River Res. Applic. 30: 874–886 (2014)
DOI: 10.1002/rra

Figure 3. Scattergram of detrended correspondence analysis ofgeomorphic indices and all vegetation types
A. ORTMANN-AJKAI ET AL.882
Including only the primarily water-determined habitattypes, HHEMs in the analysis and DCA correlation resultswere also high: between habitat and geomorphometricvariables 95.4% for the first, 73% for the second and thirdaxes, cumulative explained variance above 50% (Table V).Direct analysis of background variables by DCCA again
showed strong habitat–environment correlation: 96.5% forthe first and 86% for the second and third axes andexplained cumulative variance of habitats was 74.5%.The DCCA scattergram (Figure 4) clearly displayed the
distinct longitudinal pattern and recognizable qualitatively
Table V. Variability of habitats encoding high ecological memoryunconstrained ordination (DCA), b.: constrained ordination (DCCA)
a. DCA results for primarily water-determined vegetation types
Axes 1 2
Eigenvalues 0.867 0.1Lengths of gradient 2.623 1.9Habitat–environment correlations 0.954 0.7Cumulative percentage varianceof habitat data 42.0 48.8of habitat–environment relation 49.8 64.4
b. DCCA results for primarily water-determined vegetation types
Axes 1 2
Eigenvalues 0.800 0.4Lengths of gradient 2.066 2.2Habitat–environment correlations 0.965 0.8Cumulative percentage varianceof habitat data 38.8 59.2of habitat–environment relation 52.8 86.1
Copyright © 2013 John Wiley & Sons, Ltd.
on themaps: headwater-to-lower reach sequence of vegetationtypes of descending water demand, the separation of the moredisturbance-tolerant sedge type and the strong correlationbetween geomorphometric variables.The pairwise correlations of geomorphometric and
habitat variables (Table VI) support the patterns visibleon scattergrams. Habitat variables were negatively corre-lated in most cases. Although these results are not signif-icant, they suggest that these types occupy differentniches. Positive significant correlation was found only be-tween marsh and mesophilous meadows. These types, oc-curring together on Reach 4, are often difficult todistinguish, as they merge into each other gradually viaa broad ecotone. Habitat variables showed strong pairwisecorrelations with geomorphometric variables. Alder grovescorrelated significantly (p< 0.05) with all geomorphometricvariables, presenting high positive correlation values withLPI and slope (confined, relatively steep headwater reach,clear running water) and smaller negative correlation valueswith other variables. Mesophilous meadows had four,whereas marsh meadows had three significant correlationsof five cases (p< 0.05). Mesophilous meadows showed highpositive correlation with floodplain width: they occur in thelowermost surfaces, with no direct water cover but havingsubstantial subsurface water supply. Marsh meadows stoodout with high positive correlation with sinuosity, supportingthe empirical fact that they often occur in abandoned, butseasonally and partially flooded meanders. Sedge beds areexceptional: because they occur along all reaches, needingonly small temporarily flooded growing sites, they showedno significant correlation with any geomorphometric
(HHEMs) explained by hydrogeomorphological variables. a.:
3 4 Total inertia
40 0.047 0.000 2.06405 1.899 0.000 —31 0.734 0.000 —
51.1 0.0 —0.0 0.0 —
3 4 Total inertia
22 0.112 0.204 2.06484 2.277 2.911 —57 0.859 0.000 —
64.6 74.5 —0.0 0.0 —
River Res. Applic. 30: 874–886 (2014)
DOI: 10.1002/rra

Table VI. Pairwise correlations between geomorphic and vegecoefficients). Significant p values (p< 0.05) and high correlation
puncorr/ correlation Mesophilous meadows Sedges Ma
Mesophilous meadows — 0.52Sedges �0.07 —Marsh meadows 0.39 �0.1Alder �0.13 �0.11FWidth 0.55 �0.16FSlope �0.33 0.00LPI �0.25 �0.09CSin 0.21 �0.09VDepth 0.11 �0.02
FWidth, floodplain width; FSlope, floodplain slope; LPI, longitudinal profi
Figure 4. Scattergram of detrended canonical correspondence analysis of geomorphic indices and habitats encoding high ecologica
memory
WETLAND HABITAT PATCHES AND LANDSCAPE MEMORY 883
Copyright © 2013 John Wiley & Sons, Ltd.
-l
variables or reaches (despite that they were the dominantvegetation type of Reach 2).
DISCUSSION
Processes and patterns
Brierley (2010) distinguishes geologic, climatic andanthropogenic memory, with timescales of millennia,thousands from hundred thousands and ten-to-thousandsyears, respectively.The recent patterns of vegetation patches—representing
ecological memory—are remnants of pre-Neolithic naturalvegetation. It was determined dominantly by large-scaleQuaternary geologic and climatic factors, reflected in largerpatches of natural types.
tationvalues
rsh me
0.00.3—�0.00.3
�0.2�0.10.50.1
le; CS
Present habitat structure is fragmented, consisting ofpatches of natural and seminatural types of some hectares,mainly controlled by human factors operating on the samespatial scale (primarily agriculture because the Neolithicand river regulations intensified in the 19th century).The high vegetation–geomorphology correlation values,
and the fact that in the case of HHEMs DCA and DCCAresults were very similar (Table IV), imply that the recentspatial distribution of habitats is determined in a high degreeby geomorphological factors, that is elements of geologicand climatic memory, despite the fact that they occur in aregulated floodplain, dominated by intensive land use.Primarily water-determined HHEMS and other types were
arranged on the two sides of the DCA scattergram (Figure 3).All HHEMs are found in the ‘sinuous’ part of the diagram;on the other side, more anthropogenic types prevail. Thereseems to be a positive link between sinuosity—highest inReaches 2 and 4 (Table III)—naturalness, water demandand ecological memory of habitats, suggesting the existenceof one or more explaining factor(s), which seem(s) to relateto sinuosity. Lateral channel shift widens the floodplain anddecreases valley slope, creating embayments. In the embay-ments, meanders have more space to develop together withfloodplain forms attached to them, for example point bars,natural levees and backswamps. The richness of geomor-phological features creates a wide array of abiotic habitattypes with different inundation regimes, groundwater levelsand sediment grain size distributions from medium-grainedsand to silt. At the same time, the lower levels of em-bayments with higher flood risk are less preferred by inten-sive agriculture, so near-natural vegetation types (includingwet and mesophilous meadows, created by traditionalagriculture) have more chance to be preserved.As river regulation has significantly altered land use, we
may assume that differences between the reaches have beenreduced. The river reaches still show significant variations inhabitat structure and composition even after regulation.More natural, primarily water-determined vegetation types,
variables (upper triangle: p values; lower triangle: correlation(>0.5 or<�0.5) are in bold
adows Alder FWidth FSlope LPI CSin VDepth
0 0.21 0.00 0.00 0.02 0.04 0.314 0.30 0.12 1.00 0.42 0.41 0.82
0.73 0.00 0.01 0.09 0.00 0.084 — 0.00 0.00 0.00 0.00 0.004 �0.39 — 0.00 0.00 0.00 0.009 0.62 �0.79 — 0.00 0.00 0.008 0.70 �0.67 0.88 — 0.00 0.008 �0.30 0.49 �0.51 �0.45 — 0.008 �0.52 0.57 �0.84 �0.79 0.45 —
in, channel sinuosity; VDepth, Valley depth
River Res. Applic. 30: 874–886 (2014)
DOI: 10.1002/rra

A. ORTMANN-AJKAI ET AL.884
serving as HHEMs (alder groves, sedge beds and marsh andmesophilous meadows), unambiguously characterizeindividual reaches, despite their fragmentary occurrence(4%). Even anthropogenic vegetation types (e.g. degradedgrasslands) are bound to geomorphological variables andto reaches determined by them. Land use effects are moreexplicit in the occurrence of more degraded vegetation types,reflected by the lower explained cumulative percentagevariance values and greater difference between DCA andDCCA results.
Components of ecological memory
This spatial pattern of habitat patches of floodplains, ascolonization routes—ecological or green corridors(Ortmann-Ajkai and Horváth, 2010)—for species, definethe external component of the ecological memory and alsoprovide refugia for species and their interaction networks(internal components of ecological memory (Bengtssonet al., 2003)).In our case study, this is well represented by mesophilous
forest species, which—as internal components of ecologicalmemory—are common in the relatively natural forests of thehills, but relicts of former floodplain forests in the lower partof the floodplain. Although species presence has not beenexplicitly studied here, because we have concentrated onhabitat–geomorphology interactions, we have some formerdata from local floristic studies. Király (1998, 2006) havefound such species, for example Equisetum telmateia,Dryopteris carthusiana and Polygonatum latifolium, thatform secondary Alnus stands of Reach 4. These relictpopulations may be regarded as external components ofecological memory, perhaps as ‘sinks’ (Pulliam, 1988)being maintained by ‘source’ population above via thefloodplain green corridor. The same have been observed ina neighbouring, similar landscape, where in Reach 1position marginal populations of some mesophilous forestspecies (Asarum europaeum, E. telmateia, Festuca drymeia,Knautia drymeia, P. multiflorum and Staphylea pinnata); inlower parts of this floodplain, C. pendula, Galeobdolonluteum and Aegopodium podagraria were found(Ortmann-Ajkai, unpublished data). These findings alsopoint at the fact that secondary tree stands may serve asrefugia for forest species.
CONCLUSIONS
Vegetation is an intrinsic part of all (except entirely man-made) landscapes. Vegetation patches are indirectlyinfluenced by the physical (hydrogeomorphic) pattern,which in turn reflects landscape memory; at the same time,as ecological entities, also affected by their own ecologicalmemory—according to the principles of eco-geomorphology.
Copyright © 2013 John Wiley & Sons, Ltd.
Complex landscape studies—including landscape memorystudies—should also include ecological factors (reflectingecological memory).We propose that in addition to the classic elements of
landscape memory—geologic, climatic and anthropogenicmemory (Brierley, 2010)— and also ecological memorycomponents, species composition and in-patch andbetween-patch interactions should be considered. The factthat interactions between patches are controlled by thespatial structure of memory patches, which in turn arestrongly influenced by hydrogeomorphic factors, under-lines that floodplain wetlands—even fragmented ones instrongly regulated floodplains are first-class objects forstudies on ecological and landscape memory. HHEMspreserve both internal and external components ofecological memory: floodplain-specific species pools andspecies interactions and a spatial ecological network ofgreen islands or stepping stones, able to survive for along time, if only they are well supplied with water.Their species—if the environmental conditions arefavourable or made favourable, for example by revitaliza-tion projects—have the ability for recolonizing thesurroundings, therefore HHEMs are the starting pointsfor restoration.Their conservation role is even more important in the
present period of accelerated climate change, when agricul-tural activity shifts from the hills towards the floodplains,rather taking the flood risk there than drought risk on higherlying areas. Hydrogeomorphological patterns in floodplains—defined by landscape memory: geologic, climatic andanthropogenic memory and wetland ecosystem patchesdefined and encoded by ecological memory—jointly buildthe present day floodplain landscapes. The integration ofecological memory into traditional landscape memorypromotes the holistic understanding of landscapes,necessary for the reconstruction of past natural environ-ments—both in theory (scientific research) and practice(rehabilitation programmes).
ACKNOWLEDGEMENTS
Authors are grateful to the Institute of Ecology and Botany ofthe Hungarian Academy of Sciences for the data from theMÉTA Database. Authors are grateful for the financialsupport from the Hungarian Science Foundation (OTKA,contract number: K 68903). The funding within theframework of the Baross Gábor Grant (PTE TM09) is alsoacknowledged. We are also grateful to more anonymousreviewers and to Christian Wolter for their suggestions,especially for the recommendation to elaborate the ecologicalmemory concept. Thanks go to RichardWilliamMcIntosh forchecking English grammar and style.
River Res. Applic. 30: 874–886 (2014)
DOI: 10.1002/rra

WETLAND HABITAT PATCHES AND LANDSCAPE MEMORY 885
REFERENCES
Allan JD. 2004. Landscapes and riverscapes: the influence of land use onstream ecosystems.. Annual Review of Ecology, Evolution, andSystematics 35: 257–284. DOI: 10.1146/annurev.ecolsys.35.120202.110122
Apan AA, Raine SR, Paterson MS. 2002. Mapping and analysis of changein the riparian landscape structure of the Lockyer Valley Catchment,Queensland, Australia. Landscape and Urban Planning 59: 43–47.DOI:10.1016/S0169-20460100246-8 DOI:10.1016/S0169-2046(01)00246-8
Bándi G, Petres F, Maráz B. 1979. Baranya megye az őskorban. (Baranyacounty in the prehistoric). In Baranya Megye Története az Őskortól aHonfoglalásig, Bándi G (ed.). Baranya Megyei Levéltár: Pécs (inHungarian).
Bengtsson J, Angelstam P, Elmquist T, Emanuelsson U, Folke C, Ihse M,Moberg F, Nyström M. 2003. Reserves, resilience and dynamiclandscapes. Ambio 32: 389–396. DOI: 10.1579/0044-7447-32.6.389
Bölöni J, Molnár Zs, Illyés E, Kun A. 2007. A new habitat classificationand manual for standardized habitat mapping. Annali di Botanica(nuova serie) 7: 55–76.
Borhidi A, Kevey B, Lendvai G. 2012. Plant Communities of Hungary.Akadémiai: Budapest.
Bornette G, Amoros C, Piegay H, Tachet J, Hein T. 1998. Ecological com-plexity of wetlands within a river landscape. Biological Conservation 85:35–45. DOI:10.1016/S0006-3207 (97)00166-3 DOI:10.1016/S0006-3207(97)00166-3
ter Braak CJF, Smilauer P. 2002. CANOCO 4.5. CANOCO ReferenceManual and CanoDraw for Windows User's Guide. Software forCanonical Community Ordination Version 4.5. Wageningen and CeskeBudejovice: Biometris.
Brierley GJ. 2010. Landscape memory: the imprint of the past on contem-porary landscape forms and processes. Area 42: 76–85. DOI: 10.1111/j.1475-4762.2009.00900.x
Church M. 2002. Geomorphological thresholds in riverine landscapes.Freshwater Biology 47: 541–557.
Corenblit D, Tabacchi E, Steiger J, Gurnell AM. 2007. Reciprocalinteractions and adjustments between fluvial landforms and vegetationdynamics in river corridors: a review of complementary approaches.Earth-Science Reviews 84: 56–86. DOI:10.1016/j.earscirev.2007.05.004
Corenblit D, Steiger J, Gurnell AM, Naiman RJ 2009. Plants intertwine flu-vial landform dynamics with ecological succession and natural selection:a niche construction perspective for riparian systems. Global Ecologyand Biogeography 18: 507–520. DOI: 10.1111/j.1466-8238.2009.00461
Csontos L, Magyari Á, Van Vliet-Lanoë B, Musitz B. 2005. Neotectonicsof the Somogy Hills (part II): evidence from seismic sections.Tectonophysics 410(1–4): 63–80. DOI:10.1016/j.tecto.2005.05.049
Gillarova H, Trpák P, Trpáková I, Sykorová Z, Pecharová E. 2008.Landscape memory as a solution of the ecological stability ofthe territory after mining. Gospodarska surowcami mineralnymi24: 289–298. downloaded from: http://meeri.eu/Wydawnictwa/GSM2431/10-gillarova-i-inni.pdf, accessed 30.01.2013.
Gunderson LH. 2000. Ecological resilience – in theory and application.Annual Review of Ecology and Systematics 31: 425–39
Gurnell AM, Bertoldi W, Corenblit D. 2012. Changing river channels: theroles of hydrological processes, plants and pioneer fluvial landforms inhumid temperate, mixed load, gravel bed rivers. Earth-Science Reviews111: 129–141. DOI: 10.1016/j.earscirev.2011.11.005
Hammer Ø, Harper DAT, Ryan PDT. 2001. PAST: paleontological statis-tics software package for education and data analysis. PaleontologiaElectronica 4: 1–9. http://palaeo-electronica.org/2001_1/past/issue1_01.htm, accessed on 30.02.2013.
Hanski I. 2001. Spatially realistic theory of metapopulation ecology.Naturwissenschaften 88: 372–381. DOI: 10.1007/s001140100246
Copyright © 2013 John Wiley & Sons, Ltd.
Horváth F, Polgár L. 2008. MÉTA SQL expert interface and access service.Acta Botanica Hungarica 50(Suppl): 35–45. DOI: 10.1556/ABot.50.2008.Suppl.3
Király G. 1998. Adatok a Délkelet-Dunántúl flórájához. (Floristical dataconcerning South-East Transdanubia, Hungary). Somogyi MúzeumokKōzleményei 13: 211–215.
Király G. 2006. Kiegészítések Külso-Somogy edényes flórájánakismeretéhez. (Additions to the vascular flora of Külsõ-Somogy region,Hungary). Somogyi Múzeumok Közleményei 17: 31–40.
Leps J, Smilauer P. 2003. Multivariate Analysis of Ecological Data UsingCANOCO. Cambridge University Press: Cambridge
Lóczy D. 2013. Hydromorphological-Geoecological Foundations of Flood-plain Management: Case Study from Hungary. Lambert AcademicPublishing: Saarbrücken
Lóczy D, Pirkhoffer E, Gyenizse P. 2011. Geomorphometric floodplainclassification in a hill region of Hungary. Geomorphology 147-48: 61–72.DOI:10.1016/j.geomorph.2011.06.040
Magyari Á, Musitz B, Csontos L, van Vliet-Lanoë B. 2005. Quaternaryneotectonics of the SomogyHills, Hungary (part I): evidence from field obser-vations. Tectonophysics 410: 43–62. DOI:10.1016/j.tecto.2005.05.044
Marston RA, Girel J, Pautou G, Piegay H, Bravard JP, Arneson, C. 1995.Channel metamorphosis, floodplain disturbance, and vegetation develop-ment: Ain River, France. Geomorphology 13: 121–131. DOI:10.1016/0169-555X(95)00066-E
Molnár Zs, Bartha S, Seregélyes T, Illyés E, Botta-Dukát Z, Tímár G, HorváthF, Révész A, Kun A, Bölöni J, Biró M, Bodonczi L, Deák ÁJ, Fogarasi P,Horváth A, Isépy I, Karas L, Kecskés F, Molnár Cs, Ortmann-né Ajkai A,Rév Sz. 2007. A grid-based, satellite-image supported, multi-attributedvegetation mapping method (MÉTA). Folia Geobotanica 42: 225–247.DOI: 10.1007/BF02806465
Nakamura F, Kawaguchi Y, Nakano D, Yamada H. 2008. Ecologicalresponses to anthropogenic alterations of gravel-bed rivers in Japan, fromfloodplain river segments to microhabitat scale: a review. In Gravel-bedRivers VI: From Process Understanding to River Restoration, HabersackH, Piegay H, Rinaldi M (eds). Elsevier: Amsterdam
Orr HG, Large ARG, Newson MD, Walsh CL. 2008. A predictive typologycharacterizing hydromorphologyGeomorphology 100: 32–40. DOI:10.1016/j.geomorph.2007.10.022
Ortmann-Ajkai A, Horváth F. 2010. Green corridors or degradation routes?Effects of waterside habitats on landscape health. In Botta-Dukát Z,Salamon-Albert É (eds): 19th International Workshop of EuropeanVegetation Survey, Book of Abstracts, p.111. University of Pécs: Pécs
Padisák J. 1992. Seasonal succession of phytoplankton in a large shallowlake (Balaton, Hungary) - a dynamic approach to ecological memory,its possible role and mechanisms. Journal of Ecology 80: 217–230.
Peterson GD. 2002. Contagious disturbance, ecological memory, and theemergence of landscape pattern. Ecosystems 5: 329–338.
Pulliam HR. 1988. Sources, sinks, and population regulation. AmericanNaturalist 132: 652–661.
Renschler CS, Doyle MW, Thoms MC. 2007. Geomorphology and ecosys-tems: challenges and keys for success in bridging disciplines. Geomor-phology 89: 1–8. DOI: 10.1016/j.geomorph.2006.07.011
Schaefer V. 2009. Alien invasions, ecological restoration in cities and theloss of ecological memory. Restoration Ecology 17: 171–176. DOI:10.1111/j.1526-100X.2008.00513.x
Stallins JA. 2006. Geomorphology and ecology: unifying themes for com-plex systems in biogeomorphology. Geomorphology 77: 207–216. DOI:10.1016/j.geomorph.2006.01.005.
Szilárd J. 1967. A Külső-Somogy Kialakulása és Felszínalaktana (Develop-ment and Geomorphology of the Outer Somogy Hills), FöldrajziTanulmányok/Geographical Studies 7. Akadémiai: Budapest.
Thompson JN, Reichman OJ, Morin PJ, Polis GA, Power ME, Sterner EW,Couch CA, Gough L, Holt R, Hooper DU, Keesing F, Lovell CR, Milne
River Res. Applic. 30: 874–886 (2014)
DOI: 10.1002/rra

A. ORTMANN-AJKAI ET AL.886
T, Molles MC, Roberts DW, Strauss SY. 2001. Frontiers of ecology.BioScience 51: 15–24.
Thoms MC, Parsons M. 2002. Eco-geomorphology: an interdisciplinaryapproach to river science. The structure, function and managementimplications of fluvial sedimentary systems. Publications of theInternational Association of Hydrological Science 276: 113–120
Thorp JH, ThomsMC, DelongMD. 2006. The riverine ecosystem synthesis:biocomplexity in river networks across space and time. River Researchand Applications 22: 123–147. DOI: 10.1002/rra.901
Copyright © 2013 John Wiley & Sons, Ltd.
Vaughan IP, Diamond M, Gurnell AM, Hall KA, Jenkins A, Milner NJ,Naylor, JA, Sear DA, Woodward G, Ormerod SJ. 2009. Integratingecology with hydromorphology: a priority for river science andmanagement. Aquatic Conservation: Marine and Freshwater Ecosystems19: 113–25. DOI: 10.1002/aqc.895
Ward JV, Tockner K, Arscott DB, Claret C. 2002. Riverine landscape diversity.Freshwater Biology 47: 517–539. DOI: 10.1046/j.1365-2427.2002.00893.x
Wu J, Hobbs R. 2002. Key issues and research priorities in landscapeecology: an idiosyncratic synthesis. Landscape Ecology 17: 355–365
River Res. Applic. 30: 874–886 (2014)
DOI: 10.1002/rra