vigilance behaviour and alarm calls in a captive group of meerkats, suricata suricatta
TRANSCRIPT

Z. Tierpsychol., 65, 228-240 (1984) @ 1984 Verlag Paul Parey, Berlin und Hamburg ISSN 0044-3573 / Intercode: ZETIAG
Department of Psychology, The Universi ty of Western Ontario, London
Vigilance Behaviour and Alarm Calls in a Captive Group of Meerkats, Suricata suricatta
By GREG MORAN
With 5 figures
Received: April 4 , 1983
Accepted: July 26, 1983
Abstract and Summary
At least one member of captive groups of meerkats (Suricata suricatta) typically is situated in a prominent, elevated location. This study provides an initial description of this pattern of behaviour and evidence to suggest that it serves a vigilance function. While on the ledges, animals behaved in an alert and watchful fashion and emitted two particular classes of vocalization in response to potential dangers to the colony. Other group members responded to these alarm vocalizations with a range of alerting and retreating patterns. The vigilance behaviour system provides a useful model for the investigation of several sociobiological hypotheses and of a sophisticated alarm vocalization network.
Introduction
Altruistic behaviour has become a central topic area in current treatments of ethology and sociobiology (see, for example: ALCOCK 1979; BROWN 1975; DAWKINS 1976, 1979; GOULD 1982; HINDE 1982; WILSON 1975; WITTEN- BERGER 1981). Most definitions of altruistic behaviour have been similar to that given by BROWN (1975) for operational altruism - “giving of aid in the form of arbitrarily defined goods or services to individuals of the same species who are not offspring or direct descendants of the donor and without direct benefits to the donor or its mate” (p. 198). Interest in this behaviour stems from a reevaluation of the relationship between behaviour and inheritance that gave rise to the notion of “kin selection” (HAMILTON 1963; MAYNARD SMITH 1964; WILSON 1975). Essentially, the kin selection hypothesis holds that a gene might be selected for because it predisposes an individual to behave altruistically as long as the beneficiaries are relatives of the donor (see also DAWKINS 1976, 1978, 1979). The investigation of instances where the behav-
US. Copyright Clearance Center Code Statement: 0044-3573/84/6503-0228$02.50/0

Vigilance Behaviour and Alarm Calls in a Captive Group of Meerkats 229
iour of an individual appears to aid another at cost to itself is essential to the elaboration of such theories of gene/kin selection.
Important cases of such behaviour have involved the caretaking of young by individuals other than parents; BROWN (1975) describes several instances in birds; ROWELL (1972) describes “aunts” who assist parents of rhesus young; and ROOD (1974, 1978) has studied helper behaviour in the banded and dwarf mongoose. Another related class of behaviour is alarm calling, where an in- dividual alerts a group but increases its own likelihood of being noticed by the predator (see MAYNARD SMITH 1965). Although some have argued that the pattern need not be altruistic (see for example BROWN 1975; CHARNOV and KREBS 1975), kin selection or gene selection thinking suggests that alarm calling is likely to be a component of an evolutionarily stable strategy (MAYNARD SMITH and PRICE 1973) only among closely related groups of animals. The hypothesis was directly supported by the work of SHERMAN (1977) who stu- died the behaviour of several hundred individually marked Belding’s ground squirrels (Spermophilus beldingi). H e found that warning calls were most fre- quently given by older females who had many relatives in the colony and he calculated that the potential benefits to these relatives outweighed any costs to the calling animal herself. H e also found that the likelihood that a female would give an alarm call was a direct function of the number of relatives in the nearby area. Furthermore, he observed that males moved frequently and, thus, were less likely to be near close relatives at any one time than were the relatively sedentary females. This led to the prediction that males would give fewer warning calls than females because it was less likely that close relatives might benefit. Such was the case. Alarm calling seems a rich area in which to refine some of the more sophisticated hypotheses of current sociobiological thought.
The current study sets out to describe a related pattern of behaviour which may combine the personal costs of alarm calling and a cost similar to that of caretaking behaviour. Although field studies of the meerkat (Suricata suricatta, Viverridae) are scarce, reports indicate that individuals live in family based groups of up to 25 members. The animals live in burrows; often old termite mounds or tunnels shared with ground squirrels (EWER 1963; LYNCH 1980). The meerkat’s diet is composed largely of insects. The animal’s wide-set frontal eyes are part of a sophisticated visual system that features excellent acuity and depth perception (MORAN et al., in press). In our study of a captive group we have noticed a pattern of behaviour that we tentatively have inter- preted as serving a lookout or vigilance function. Animals sometimes alarm call when displaying this pattern, but the behaviour also removes the in- dividual from normal ongoing activities for significant periods of time; poten- tially a high cost for an animal which obtains most of its food from low- return foraging. This pattern of behaviour could prove particularly fruitful for investigating the implicit trade-off between personal costs and benefits to relatives. The current study provides an initial description of the pattern of behaviour in a captive group of animals and pays special attention to evidence that might support the suggestion that it serves a sentinel role in the group.

230 GREG MORAN
Methods
Animals and Enclosure
Two female and two male meerkats were obtained on extended loan from the Metro- politan Toronto Zoo in the fall of 1978 and were maintained as a single group in the labo- ratory since then. Two animals (Al-JV$; Ma-JVq) were offspring of the other pair (Mi- AD$; Je-AD?) and were approximately two years of age at the outset of the study. The two younger animals are referred to as juveniles rather than adults for convenient reference only. The animals were free to move throughout the enclosure (see Fig. 1). Both rooms were well lit by large windows and the animals were maintained on a natural light cycle. The temperature was maintained at 27-29OC and humidity ranged between 60 and 70%. The rooms included lengths of plastic tubing, a sand-box, a running wheel, a nest-box, and ramps leading to ledges on each window sill. The meerkats were fed a meat and vegetable diet with a vitamidmineral supplement.
Procedures Three series of general observations were carried out in
July/August 1980; July/August 1981 ; and May/June 1982. 20 30-min sessions were performed in each series. Observations were made five times per week except on rare occasions. A single observer made all recordings from the observation area (see Fig. 1 ) using a check sheet. All occurrences of an animal moving up onto or down from one of the three window ledges; the identity of the animal; and, the time of the act were recorded. All four animals
1. General observations.
ROOM A
Fig. 1: Schematic illustration of the enclosure. Ea& ledge could be reached by a ramp from the floor

Vigilance Behaviour and Alarm Calls in a Captive Group of Meerkats 231
were present in 1980. In 1981, Mi-ADJ, Ma-JV?, and AI-JV$ were present at the begin- ning, but Ma-JV? died on the fifth day of the study. Mi-ADd, Je-AD?, and AI-JVJ were present throughout the observations in 1982.
2. Detailed behavioural observations. The actual behaviour of the meerkats when on the ledges was analysed in a series of observations made in 1982. This study was carried out in a room similar to room A in Fig. 1. The room was approximately the same size and had the same contents. Mi-ADJ, Je-AD?, and AI-JVJ were present in the enclosure through- out. Any animal on the ledge was videotaped during 15 30-min sessions. The tapes were later analysed using the following behavioural catalogue:
Alert Stand: Sit: Lazy Sit:
Lying Down: All Fours: Lazy Fours:
Gaze Out: Gaze In: Asleep: Grooming:
Immobile: Walk: Rim:
A. Posture
Body erect; rear legs extended fully; tail used as support. Body erect; hindquarters in contact with the ledge. Slouched, back curled forward, almost doubled over; hindquarters in contact with ground. Chest and ventral area in contact with the ground and supporting the body. Normal quadruped stance. Like Lazy Sit except head up and forelegs are extended, touching the ground but supporting little or no weight. Most of weight supported by hindquarters.
B. Gaze
Head and eyes directed out of the window. Head and eyes directed in to the room. Head drooping, eyes closed. Gaze involved in self-grooming.
C. Locomotion
Not locomoting. Typical unhurried locomotion. Faster locomotion than Walk, usually two forelegs and two rear legs move in synchrony rather than opposition.
The sub-categories within each of the three categories were mutually exclusive and the three categories were scored independently of one another. The behaviour of the animal on the ledge was sampled using an instantaneous sampling technique (SLATER 1978) and 3-s intervals. All observations were pooled irrespective of the identity of the animal on the ledge.
A second set of observations was made of animals which were not on the ledge in order to provide a broad comparison. It was not possible to employ a video record because the constant movement necessary to follow animals would have been disruptive. Therefore, a real-time observational procedure was used. It was necessary to make a few alterations to the catalogue. The gaze categories “Gaze Upward” and “Other Gaze” (all gazes not di- rected upward) replaced “Gaze Out” and “Gaze In”. It was possible to observe all three animals simultaneously and thus a focal animal instantaneous sampling technique was used (ALTMANN 1974). Samples were taken at 5-s intervals. Each session lasted 30min. A single animal was observed for 10min or until the animal moved onto the ledge. The order in which animals were observed was random and each animal was observed for a total of 60 min. A total of 8 30-min sessions were required to achieve this sample.
Samples of vocalizations were recorded in an ad hoc fashion at various times in the colony. Recordings were made using a Sony TC 366-4 tape recorder and a Shure model SM6O microphone which was suspended approximately 1 m above the floor in the centre of room A. The identity of the vocalizing animal and ongoing behaviour were noted in each case. Spectral analyses of the recordings which were relatively free of background noise were made using a Model 7030A Sonograph equipped with a 1 to 16,000
3. Vocalizations.

232 GREG MORAN
Hz Spectrum Analyzer and a Scale Magnifier (Kay Elemetrics Corporation). The clearest sample of a class of vocalization was chosen as a stimulus to be played back to the animals. The vocalization was recorded at 1-s intervals for 5 s. On 8 separate days this stimulus and a white noise of matched intensity and duration were played through a speaker using the same tape recorder. During any given session the white noise and the sample vocalization were eacb presented once and were separated by 2 min. The order of presentation of the two stimuli within a session was determined randomly. The behaviour of the animals prior to, during, and following the stimulus was recorded on videotape.
A t test for correlated observations (ELLINGSTAD and HEIMSTRA 1974) was used in all statistical comparisons. An observation session was treated as the statistical sampling unit.
Results
1. General Observations
All of the animals visited the ledges for substantial periods of time in each year of the study (Fig, 2a). In 1980, the animals did not differ sig- nificantly in the amount of time that they were on the ledge. In the final two years Milf, the oldest male, spent more time on the ledge than did other ani- mals [t(19) = 4.60, p < .001 in 1981 vs. A1-JV$; t(19) = 5.26, p < .001 in 1982 vs. Je-AD?]. Visits to the ledge were of two types: those in which the animal simply went up, investigated briefly, and then came down again; and a second class of visits where the animal remained for a substantial period in a single location and posture. Fig. 2 b illustrates the distribution of these shor- ter and longer visits over the members of the group during the three years of the study. All animals made both types of visits to the ledge. Once again, how- ever, Milf made substantially more of the longer visits during the final two years than did any other animal [t(19) = 3.15, p < .01 in 1981 vs. Al-JV$; t(19) = 3.39, p < .01 in 1982 vs. Je-AD?].
There was at least one meerkat on a ledge during observations for 97% of the time in 1980 and for 95 % of the time in 1982 (Fig. 2a). In 1981 there was no animal on a ledge for 25 % of observation time. In that same year Ma-JV? died after only four sessions. The proportion of time that she spent on the ledge during these four sessions is approximately the same as the per- cent of time that the ledges were empty during this series of observations (Fig. 2 a). Furthermore, this series also was characterized by a very large num- ber of brief visits to the ledge by the older male Mi-ADd (Fig. 2 b). H e did not display such a pattern in the preceding years nor was it characteristic of his behaviour in other studies in our colony.
2. Detailed Behavioural Observations
Animals on the ledge behaved quite differently from those in other loca- tions in the enclosure (Fig. 3). In the first place, although animals were fre- quently immobile when both on and off the ledge, meerkats on the ledge spent over 70% of their time in an alert stand or a sitting posture, whereas these postures were seen less than 2 % of the time when animals were not on the ledge. O n the other hand, the four most relaxed postures were shown a total

Vigilance Behaviour and Alarm Calls in a Captive Group of Meerkats 233
.b' 1980
l l i ) 112 1 1981
I
I ~
H
il I
1982
A H i I N T I
I
1
i
__ M O
'"Y
Fig. 2: (a) The proportion of total time during which each animal was on a ledge for each period of observation. The proportion of time during which no animal was on the ledge also is indicated. Data represents 20 30-min sessions in each year. (b) Number of visits by each animal to a ledge during each of the three series of observations. Visits are divided into those of less than 1 min duration and those longer than 1 min. (':.) Data shown for Ma-JVQ
in 1981 represents only the first four sessions prior to her death
of only 1 8 % of the time by animals on the ledge as compared to 87 % by animals in other locations. The meerkats were observed sleeping or grooming much less often on the ledge (total 8 %) than on the ground (32 %). Animals on the ledge directed their gaze out of the window 5 3 % of the time. These results indicate that the animals behaved very distinctively while on the ledge despite the fact that the different sampling procedures make a formal statistical analysis inappropriate.

234 GREG MORAN
'('('[ ON LEDGE
' " (J [ NOT ON 1 F DGE
Fig. 3: Upper panel illustrates the proportion of time that an animal on a ledge was engaged in various activities (see text for an explanation of categories). Per- centages are based on 450min of videotaped record during which the ledge was empty for a total of 8.5 min. Lower panel illustrates the proportion of time that ani- mals were engaged in a variety of activities when not on the ledge (see text for procedural details). Time spent in nest boxes and the running wheel is not represented
3. Vocalizations
Two types of vocalization were emitted disproportionately often by meer- kats on ledges and elicited reliable reactions. Unfortunately, the vocalizations occurred relatively infrequently and were elicited by stimuli beyond the con- trol of observers. Therefore the gathering of systematic quantitative observa- tions was difficult.
A set of sonograms of examples of the hoot vocalization are shown in the left hand panel of Fig. 4. The hoot is one member of a class of vocalizations whose members share a similar acoustic structure but sound quite distinctive to the ear and occur in dissimilar circumstances (see MORAN 1980). The upper- most panel reveals that the hoot does not contain acoustic energy of more than approximately 3 kHz. The vocalization is characterized by a fundamental fre- quency in the range of 600-900 H z with formant frequencies ranging as high as approximately 2500 Hz. Hoots show little modulation. Durations vary across examples of hoots. For example, the individual hoots in the bottom panel of Fig. 4 are approximately .2 s in duration whereas the hoots in the second panel last approximately .75 s. Some of this variability may be attri- butable to individual differences; the series of hoots in the bottom panel of Fig. 4 are alternating vocalizations made by two animals. The frequency of hoots and the degree of modulation for each animal is quite distinct. Hoots

Vigilance Behaviour and Alarm Calls in a Captive Group of Meerkats 235
Fig. 4: Sonographic representation of hoot and bark vocalizations. The uppermost hoot was analysed using a narrow-band (50 kHz) and a 16 kHz range. The remainder of the hoots were analysed using a wide band (300 kHz) filter and a 4 kHz range. All examples
of the bark were analysed using the wide-band filter and the 16 kHz range
were associated with the presence of a hawk circling in the area or the fly-past of one of many high-flying aircraft on a nearby international airroute. Observers reliably agreed that these stimuli elicited the hoots. The reaction of colony members to hoots almost always involved a cessation of activity and some degree of alerting or taking to cover. There appeared to be a relationship between the intensity of the hoot and the group’s reaction. A loud hoot elicited immediate flight to nest boxes and tubes; a less intense hoot was followed by a cessation of activity and a brief scanning of the environment from an alert stand posture, and perhaps a visit to the ledge and a scan of the surrounding area.
Sonagrams of examples of the bark vocalization are presented in the right hand panel of Fig. 4. The call features a packet of intense energy in the 0- 3 kHz range and a second series of frequencies in the 7-11 kHz range. Less
236 GREG MORAN
intense acoustic energy is distributed between these ranges giving the bark a far noisier character than the relatively pure sound of the hoot. The overall pattern of individual barks is relatively stereotyped. Barks occur in bursts with 0.5-1.0 s intervals between each call. The animals barked most com- monly when they saw dogs in a nearby roof kennel or were confronted by sudden unexpected events. These included a loud noise in the enclosure; attempts to catch an animal for medical purposes; and, the sudden appearance of a window cleaner outside their sixth floor ledge. Barks tended to elicit immediate retreat from the other members of the colony. As with the hoot, however, there seemed to be a direct relationship between the intensity of the hoot and the likelihood and speed of retreat to cover by the members of the group. Other factors also appeared important. For example, low intensity hoots made by Al- JV 8 when another animal scratched a tube seldom drew a response.
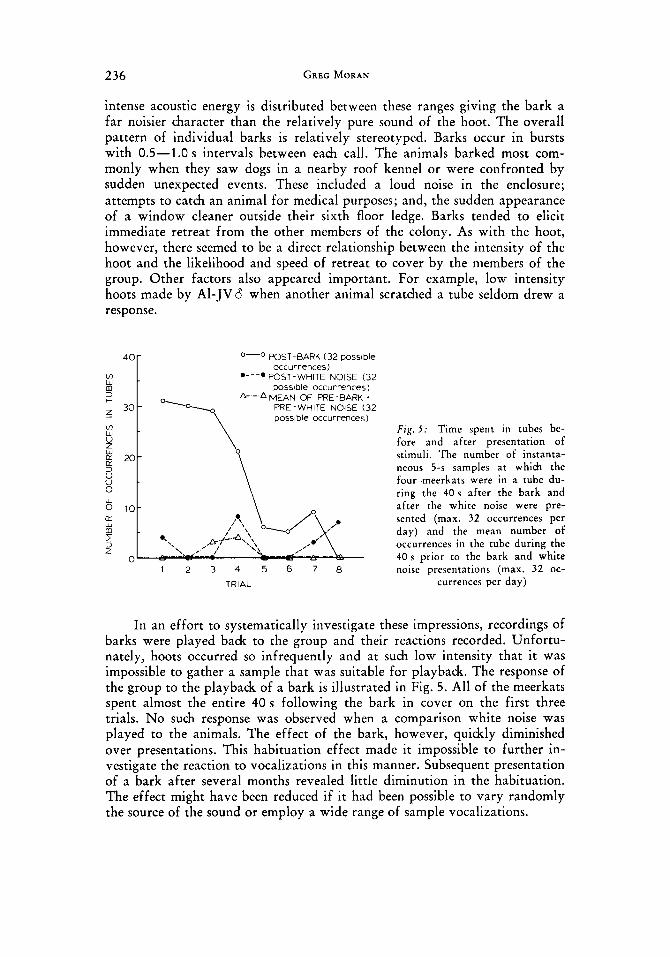
O-O POST-BARK (32 possible occurrences)
*---* POST-WHITE NOISE (32 possible occurrences)
& - MEAN OF PRE-BARK * PRE-WHITE NOISE (32 possible occurrences)
F t g . 5 : Time spent in tubes be- fore and af ter presentation of stimuli. The number of instanta- neous 5-s samples a t which the four meerkats were in a tube du- ring the 4 0 s after the bark and af ter the white noise were pre- sented (max. 32 occurrences per day) and the mean number of occurrences in the tube during the 4 0 s prior to the bark and white noise presentations (max. 32 oc-
currences per day) 1 2 3 4 5 6 7 8
TRIAL
In an effort to systematically investigate these impressions, recordings of barks were played back to the group and their reactions recorded. Unfortu- nately, hoots occurred so infrequently and at such low intensity that it was impossible to gather a sample that was suitable for playback. The response of the group to the playback of a bark is illustrated in Fig. 5. All of the meerkats spent almost the entire 40 s following the bark in cover on the first three trials. No such response was observed when a comparison white noise was played to the animals. The effect of the bark, however, quickly diminished over presentations. This habituation effect made it impossible to further in- vestigate the reaction to vocalizations in this manner. Subsequent presentation of a bark after several months revealed little diminution in the habituation. The effect might have been reduced if it had been possible to vary randomly the source of the sound or employ a wide range of sample vocalizations.

Vigilance Behaviour and Alarm Calls in a Captive Group of Meerkats 237
Discussion
Members of a captive colony of meerkats devote substantial amounts of time to an activity which appears to serve a vigilance function. All group members spent time on ledges that provided a panoramic view of the sur- rounds. At least one meerkat was on a ledge at almost all times except for a period after the death of a group member. The death of this animal also disrupted other normal behaviour patterns of the group. In fact, the propor- tion of time during which the ledge was empty corresponded almost exactly to the proportion of time that the female had spent on the ledge prior to her death. All of this evidence suggests that the pattern is an integrated feature of the animals’ behaviour. Analysis of the behaviour of animals while on the ledges supported the notion that while there, they are attentive to potential dangers to the colony and spend relatively little time engaged in activities incompatible with vigilance. Finally, meerkats on ledges frequently emitted hoots and barks at the appearance of potential dangers outside the colony and response to these vocalizations suggests that they carry a warning message.
Alarm calling has been studied in several species of ground squirrels (e.g. DUNFORD 1977; LEGER and OWINGS 1978; SHERMAN 1977) and prairie dogs (e.g. SLOBODCHIKOFF and COAST 1980; SMITH et al. 1977), but the pattern of vigilance described in this study potentially carries the additional personal cost of preventing the individual from foraging for an extended period of time. The functional explanation of the selection of such a pattern of behav- iour that apparently takes a high toll of personal fitness is open to some speculation. The pattern cannot simply be attributed to kin-selection without further investigation. Several not mutually exclusive hypotheses are open to study in the field. Firstly, the alarm calls actually might reduce the likelihood of the caller being preyed upon (see arguments by CHARNOV and KREBS 1975), although this account might be difficult to apply to extended vigilance. Secondly, kin-selection might be a contributing factor if the species’ natural predators or competing conspecific groups demand unusual watchfulness. Conditions would then be optimal for vigilance to benefit direct offspring or more distant rela- tives in a closely related group. Finally, vigilance and alarm calling might be examples of reciprocal altruism (TRIVERS 1971) because the role of benevolent sentinel is taken by all group members.
The alarm vocalizations observed in the study are of interest in their own right. MULLIGAN and NELLIS (1975) describe an alarm bark for the distantly related mongoose, Herpestes auropunctatus, that is similar to that described here. They did not observe an alarm call analogous to the hoot, but described a call with similar harmonic structure seen in other contexts. We also have observed such calls in our colony. These calls, however, sound very different and occur in contexts that do not suggest imminent danger (MORAN 1980).
The variation in alarm calls may provide a model for investigating what BEER (1975) calls syntactic and contextual coding of meaning (see also SMITH 1977). Coding by form or syntax is suggested by the observation that the intensity of both hoots and barks appears to determine the nature of other

238 GREG MORAN
animals’ response. MARLER (1975) has proposed that such a graded system carries more sophisticated information than an all-or-none signal system. SORENSEN (1981) has made the related observation in our colony that the response to a hoot depends on the identity of the vocalizing animal. In this case, contextual coding of meaning could be provided by the call’s association with a particular individual or behaviour. Thus, the meerkat may differentially encode particular characteristics of a predator or related danger in the hoot and the bark and their context. Evidence of such complex coding has been uncovered by several researchers (e.g. BEER 1976, 1979; CHENEY and SEYFARTH 1981; OWINGS and LEGER 1980; STRUHSAKER 1967) and may prove more pre- valent than originally expected.
This series of observations suggest complexities of social behaviour and communication that have seldom been observed in a non-primate species. Further captive study and long-overdue field studies of the meerkat promise to make important contributions to our understanding of the process of natural selection and vocal communication networks.
Zusammenfassung
In jeder gefangen gehaltenen Gruppe von Erdmannchen (Suricata suri- catta) gab es mindestens ein Tier, das sich gewohnlich auf eine erhohte Stelle mit guter Aussicht setzte.
In jeweils halbstundigen Beobachtungen, die in drei Serien von je 2Omal uber 3 Jahre verteilt waren, sai3 im Mittel in uber 95 % der Zeit mindestens ein Tier auf solchem Posten. Ausnahmen betrafen Zeiten nach dem Tod eines Koloniemitglieds.
Alle Tiere sai3en haufig auf solchen Posten, aber in den letzten beiden Beobachtungsperioden vor allem das erwachsene Mannchen. Es verharrte in den beiden letzten Jahren auch jeweils fur langere Zeit auf diesem Posten als die anderen Tiere.
Die auf dem Aussichtsposten befindlichen Tiere verbrachten mehr Zeit aufgerichtet und die Umgebung musternd als die Tiere, die sich anderswo auf- hielten.
Die meisten Alarmrufe kamen von Tieren auf dem Aussichtsposten. Sowohl die Alarmrufe der Individuen wie die Reaktionen der anderen darauf sind verschieden. Auf Tonbandrufe reagieren die Tiere zunachst mit Deckung- suche, doch verliert sich die Reaktion mit haufigerer Wiederholung.
Das Sitzen an erhohter Stelle scheint ein wichtiger Bestandteil der Sozial- struktur zu sein und als Wachter-Amt zu dienen. Wachesitzen ist unvertraglih mit Fressen und anderem Selbsterhaltungs-Verhalten. Das Wachter-Amt der Surikaten weist auf einen hohen Verwandtschaftsgrad innerhalb der Gruppen hin.

Vigilance Behaviour and Alarm Calls in a Captive Group of Meerkats 239
Acknowledgements
This research was supported by grants to the author from the Natural Sciences and Engineering Research Council of Canada. I would like to thank the following for their' help in data collection at various times during the study: Linda SORENSEN, Mary GILBERT, Phil KRAEMER, Barb DESROCHERS, Kathy PETRON, and Vaughn SUNDAY. Thanks to K. P. OSSENKOPP for translating the Germany summary and to Wanda A. MARKS for her patience through many revisions. The animals observed in this study were provided on extended loan to the author by the Metropolitan Toronto ZOO.
Literature Cited
ALCOCK, J. (1979): Animal Behavior, 2nd ed. Sinauer Assoc., Sunderland, Mass. ALTMANN, J. (1974) : Observational study of behaviour: sampling methods. Behaviour 49,
BEER, C. (1975): Multiple functions and gull display. In: Function and Evolution in Behaviour. (BAERENDS, G., C. BEER and A. MANNING, eds.) Oxford Univ. Press, Oxford, pp. 17-54 BEER, C. (1976): Some complexities in the communication behavior of gulls. Ann. N.Y. Acad. Sci. 280, 413-432 BEER, C. (1979): Vocal communication between laughing gull parents and chicks. Behaviour 70, 119-146 BROWN, J. L. (1975): The Evolu- tion of Behavior. W. W. Norton, New York.
CHARNOV, E. L., and J. R. KREBS (1975): The evolution of alarm calls: altruism or manipulation? Am. Nat. 109, 107-112 CHENEY, D. L., and R. M. SEYFARTH (1981): Selective forces affecting the predator alarm calls of vervet monkeys. Behaviour 76, 25-61.
DAWKINS, R. (1976): The Selfish Gene. Oxford Univ. Press, Oxford, New York D~WKINS, R. (1978): Replicator selection and extended phenotype. Z. Tierpsychol. 47, 61-76 DAWKINS, R. (1979): Twelve misunderstandings of kin selection. 2. Tierpsychol. 51, 184-200 DUNFORD, C. (1977): Kin selection for ground squirrel alarm calls. Am. Nat.
ELLINGSTAD, V., and N. W. HEIMSTRA (1974): Methods in the Study of Behaviour. Brooks/Cole, Monterey, Calif. EWER, R. F. (1963): The behaviour of the meerkat, Suricata suricatta (Schreber). 2. Tierpsychol. 20, 570-607.
227-267.
111, 782-785.
GOULD, J. L. (1982): Ethology. Norton, New York. HAMILTON, W. D. (1963): The evolution of altruistic behavior. Am. Nat. 97, 354-356
HINDE, R. A. (1982): Ethology. Oxford Univ. Press, Oxford. LEGER, D. W., and D. H. OWINGS (1978): Responses to alarm calls of California
ground squirrels: effects of call structure and maternal status. Behav. Ecol. Sociobiol. 3, 177-186 LYNCH, C. D. (1980): Ecology of the suricate, Suricata suricatta and the yellow mongoose, Cylictis penicillata with special reference to their reproduction. Memoirs Nasio- nale Mus. 14, 1-145.
~ ~ R L E R , P. (1975): Social organization, communication and graded signals: the chim- panzee and the gorilla. In: Growing Points in Ethology. (BATESON, P. P. G., and R. A. HINDE, eds.) Cambridge Univ. Press, Cambridge, pp. 239-280 MAYNARD SMITH, J. (1964): Kin selection and group selection. Nature 201, 1145-1147 MAYNARD SMITH, J. (1965): The evolution of alarm calls. Am. Nat. 99, 59-63 MAYNARD SMITH, J., and G. R. PRICE (1973): The logic of animal conflict. Nature 246, 15-18 MORAN, G. (1980): Behavioral studies of the meerkat. Ann. Proc. Am. Assoc. Zoo Veterinarians MORAN, G., B. TIMNEY, L. SORENSEN and B. DESROCHERS (1983): Binocular depth perception in the meerkat (Suri- cata suricatta). Vision Res. 23, 965-969 MULLIGAN, B. E., and D. W. NELLIIS (1975): Vocal repertoire of the mongoose, Herpestes auropunctatus. Behaviour 55, 247-267.
OWINGS, D. H., and D. W. LEGER (1980): Chatter vocalizations of California ground squirrels: predator- and social-role specificity. 2. Tierpsychol. 54, 163-184.

240 MORAN, Vigilance Behaviour and Alarm Calls in a Captive Group of Meerkats
ROOD, J. P. (1974): Banded mongoose males guard young. Nature 248, 176-177 ROOD, J. P. (1978): Dwarf mongoose helpers at the den. Z. Tierpsychol. 48, 277-287 ROWELL, F. T. (1972): Social Behaviour of Monkeys. Penguin, Harmondsworth, Middlesex.
SHERMAN, P. W. (1977): Nepotism and the evolution of alarm calls. Science 197, 1246-1253 0 SLATER, P. J. B. (1978): Data collection. In: Quantitative Ethology. (COLGAN, P. W,. ed.) John Wiley, New York, pp. 7-24 SLOBODCHIKOFF, C. N., and R. COAST (1980): Dialects in the alarm calls of prairie dogs. Behav. Ecol. Sociobiol. 7, 49-53 SMITH, W. J. (1977): The Behavior of Communicating. Harvard Univ. Press, Cambridge, Mass. SMITH, W. J., S. L. SMITH, E. C. OPPENHEIMER and J. G . DEVILLA (1977): Vocaliza- tions of the black-tailed prairie dog, Cynomys ludovicianus. Anim. Behav. 25, 152-164 . SORENSEN, L. (1981) : The behavioural repertoire of the meerkat, Suricata suricatta. Unpubl. M. A. Thesis, Univ. of Western Ontario, London STRUHSAKER, T. T. (1967): Auditory communication among vervet monkeys (Cercopithecus aethiops). In: Social Communication Among Primates. (ALTMANN, S. A., ed.) Univ. of Chicago Press, Chicago, pp. 281-324.
TRIVERS, R. L. (1971): The evolution of reciprocal altruism. Qu. Rev. Biol. 46, 35-57. WILSON, E. 0. (1975): Sociobiology. Belknap Press, Cambridge, Mass. WITTEN-
BERGER, J. F. (1981): Animal Social Behavior. Wadsworth, Belmont, Calif.
Author’s address: G . MORAN, Department of Psychology, University of Western Ontario, London, Ontario N6A 5C2, Canada.