vertical distribution of aurelia labiata (scyphozoa) jellyfish in roscoe bay is similar during flood...
TRANSCRIPT

Journal of Sea Research 64 (2010) 422–425
Contents lists available at ScienceDirect
Journal of Sea Research
j ourna l homepage: www.e lsev ie r.com/ locate /seares
Vertical distribution of Aurelia labiata (Scyphozoa) jellyfish in Roscoe Bay is similarduring flood and ebb tides
David J. AlbertRoscoe Bay Marine Laboratory, 4534 W 3rd Avenue, Vancouver, Canada V6R 1N2
E-mail address: [email protected].
1385-1101/$ – see front matter © 2010 Elsevier B.V. Adoi:10.1016/j.seares.2010.02.005
a b s t r a c t
a r t i c l e i n f oArticle history:Received 27 September 2009Received in revised form 24 January 2010Accepted 9 February 2010Available online 16 February 2010
Keywords:Aurelia labiataBehaviourHydrozoaSyphozoaTidesVertical Migration
The vertical distribution of the jellyfish, Aurelia labiata, was assessed during flood and ebb tides in April, May,June, and August 2009 in Roscoe Bay (British Columbia, Canada). Assessments were made at locations alongthe entire length of the bay. The average tide change was 3.7 m. The vertical position of medusae (in 2 mintervals) did not differ significantly during ebb (N=17) and flood tides (N=18) in 6 and 8m watercolumns. These results indicate that A. labiata do not use tidally synchronized vertical migration to maintaintheir location or presence in Roscoe Bay.
ll rights reserved.
© 2010 Elsevier B.V. All rights reserved.
1. Introduction
Behaviour as well as tidal currents influence the horizontaldistribution of Aurelia sp jellyfish (Hamner, et al., 1994; Toyokawa et al.,1997; Purcell, et al., 2000; RakowandGraham, 2006;Magomeet al., 2007;Lo and Chen, 2008; Albert, 2007, 2009). Aurelia sp also space themselvesvertically (Papathanassiou et al., 1987; Rakow and Graham, 2006; Malej,et al., 2007;HanandUye, 2009;Albert, 2009). ShanksandGraham(1987),Zavodnik (1987), Malej (1989), and Dawson and Hamner (2003) haveprovided similar evidence for the scyphozoans Stomopolus meleagris L.agassiz, Pelagia noctiluca, andMastigias, respectively.
In the Wadden Sea, the hydrozoans Rathkea octopunctata, Sarsiatubulosa, Bougainvillia ramosa, and Eucheilota maculata, and thectenophore Pleurobrachia pileus remained in the inner part of anestuary by migrating to the moving surface water during flood tidesand to the still water near the bottom during ebb tides (van der Veerand Sadée, 1984; Kopacz, 1994). Some copepods and mysids behavein a similar way (Trinast, 1975; Wooldridge and Erasmus, 1980; forreviews see Creutzberg, 1975; Naylor, 2006). Vertical migrationsenable these species to drift to the inner part of the estuary on floodtides and to avoid drifting out of the estuary on ebb tides.
Aurelia labiata are a resident population in Roscoe Bay. When thepopulation is small, medusae usually accumulate at the end of the bayfarthest from the bay's entrance (Albert, 2009). Further, A. labiataswim down when they are swept over a gravel bar and out of the bayon ebb tides and swim up when the tide turns to flood (Albert, 2007).
These observations raise the possibility that A. labiata are usingvertically synchronized tidal migration to stay in Roscoe Bay just assome hydrozoans use it to stay in the Wadden Sea (van der Veer andSadée, 1984; Kopacz, 1994). The validity of this line of reasoning issuspect since A. labiata spread across the entire length of RoscoeBay when the population is large (Albert, 2007, 2009). Further,preliminary observations by Verwey (1966) on Chrysaora hysoscellaand Rhizostoma pulmo suggest that these scyphozoans do not displaytidally synchronized vertical migration in the Wadden Sea.
The present observations examined the possibility that A. labiata areusing tidally synchronized vertical migration to assist them in remainingin Roscoe Bay. Assessments of the vertical location ofmedusaeweremadealong the entire length of the bay during the time of year when large tidechanges would make the use of vertical migration most advantageous.
2. Method
2.1. Vertical distribution of medusae
Lift nets were used to assess the position of medusae in the watercolumn. The lift nets were in the shape of an open cone (0.70 mdiameter at the large end; 0.75 m deep, 0.30 m dia. at the small end,3 mm mesh). The mesh was attached to a ring of 1.6 cm dia. plasticpipe at the top and a ring of 1 cm dia. polyethylene tubing at thebottom. Both ringswere filledwith sand. Each net wasweighted at thebottom with a 19 cm length of 1.5 cm dia. copper pipe. Nets werereleased into the water with the plastic rings in a vertical orientation.The open cone shape allowed medusae to be easily spilled from thenet and the large mesh facilitated water flow.
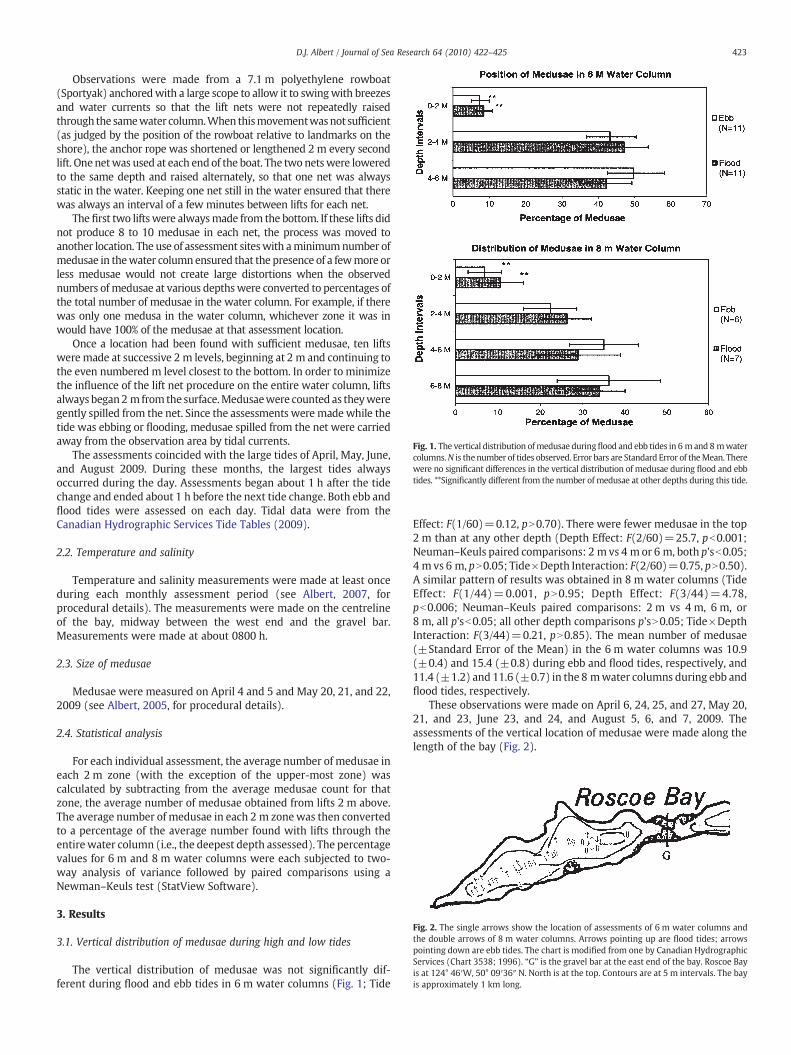
Fig. 1. The vertical distribution ofmedusae duringflood and ebb tides in 6 m and 8 mwatercolumns.N is thenumber of tides observed. Error bars are Standard Error of theMean. Therewere no significant differences in the vertical distribution of medusae during flood and ebbtides. **Significantly different from the number of medusae at other depths during this tide.
Fig. 2. The single arrows show the location of assessments of 6 m water columns andthe double arrows of 8 m water columns. Arrows pointing up are flood tides; arrowspointing down are ebb tides. The chart is modified from one by Canadian HydrographicServices (Chart 3538; 1996). “G” is the gravel bar at the east end of the bay. Roscoe Bayis at 124° 46′W, 50° 09′36″ N. North is at the top. Contours are at 5 m intervals. The bayis approximately 1 km long.
423D.J. Albert / Journal of Sea Research 64 (2010) 422–425
Observations were made from a 7.1 m polyethylene rowboat(Sportyak) anchoredwith a large scope to allow it to swingwith breezesand water currents so that the lift nets were not repeatedly raisedthrough the samewater column.When thismovementwasnot sufficient(as judged by the position of the rowboat relative to landmarks on theshore), the anchor rope was shortened or lengthened 2 m every secondlift. Onenetwas used at each endof the boat. The two netswere loweredto the same depth and raised alternately, so that one net was alwaysstatic in the water. Keeping one net still in the water ensured that therewas always an interval of a few minutes between lifts for each net.
The first two lifts were alwaysmade from the bottom. If these lifts didnot produce 8 to 10 medusae in each net, the process was moved toanother location. The use of assessment siteswith aminimumnumber ofmedusae in thewater columnensured that the presence of a fewmore orless medusae would not create large distortions when the observednumbers of medusae at various depthswere converted to percentages ofthe total number of medusae in the water column. For example, if therewas only one medusa in the water column, whichever zone it was inwould have 100% of the medusae at that assessment location.
Once a location had been found with sufficient medusae, ten liftsweremade at successive 2 m levels, beginning at 2 m and continuing tothe even numbered m level closest to the bottom. In order to minimizethe influence of the lift net procedure on the entire water column, liftsalways began2 m from the surface.Medusaewere counted as theyweregently spilled from the net. Since the assessments were made while thetide was ebbing or flooding, medusae spilled from the net were carriedaway from the observation area by tidal currents.
The assessments coincided with the large tides of April, May, June,and August 2009. During these months, the largest tides alwaysoccurred during the day. Assessments began about 1 h after the tidechange and ended about 1 h before the next tide change. Both ebb andflood tides were assessed on each day. Tidal data were from theCanadian Hydrographic Services Tide Tables (2009).
2.2. Temperature and salinity
Temperature and salinity measurements were made at least onceduring each monthly assessment period (see Albert, 2007, forprocedural details). The measurements were made on the centrelineof the bay, midway between the west end and the gravel bar.Measurements were made at about 0800 h.
2.3. Size of medusae
Medusae were measured on April 4 and 5 and May 20, 21, and 22,2009 (see Albert, 2005, for procedural details).
2.4. Statistical analysis
For each individual assessment, the average number of medusae ineach 2 m zone (with the exception of the upper-most zone) wascalculated by subtracting from the average medusae count for thatzone, the average number of medusae obtained from lifts 2 m above.The average number of medusae in each 2 m zonewas then convertedto a percentage of the average number found with lifts through theentire water column (i.e., the deepest depth assessed). The percentagevalues for 6 m and 8 m water columns were each subjected to two-way analysis of variance followed by paired comparisons using aNewman–Keuls test (StatView Software).
3. Results
3.1. Vertical distribution of medusae during high and low tides
The vertical distribution of medusae was not significantly dif-ferent during flood and ebb tides in 6 m water columns (Fig. 1; Tide
Effect: F(1/60)=0.12, pN0.70). There were fewer medusae in the top2 m than at any other depth (Depth Effect: F(2/60)=25.7, pb0.001;Neuman–Keuls paired comparisons: 2 m vs 4 mor 6 m, both p'sb0.05;4 m vs 6 m, pN0.05; Tide×Depth Interaction: F(2/60)=0.75, pN0.50).A similar pattern of results was obtained in 8 m water columns (TideEffect: F(1/44)=0.001, pN0.95; Depth Effect: F(3/44)=4.78,pb0.006; Neuman–Keuls paired comparisons: 2 m vs 4 m, 6 m, or8 m, all p'sb0.05; all other depth comparisons p'sN0.05; Tide×DepthInteraction: F(3/44)=0.21, pN0.85). The mean number of medusae(±Standard Error of the Mean) in the 6 m water columns was 10.9(±0.4) and 15.4 (±0.8) during ebb and flood tides, respectively, and11.4 (±1.2) and 11.6 (±0.7) in the 8 mwater columns during ebb andflood tides, respectively.
These observations were made on April 6, 24, 25, and 27, May 20,21, and 23, June 23, and 24, and August 5, 6, and 7, 2009. Theassessments of the vertical location of medusae were made along thelength of the bay (Fig. 2).
Table 1Mean tide change on the days when these observations were made. Seawatertemperature and salinity were measured from the surface to the bottom in 2-mintervals.
Mean tide Seawater temperature (C) Salinity (ppt)
Change (m) Depth (m) Depth (m)
0.1 2 4 6 8 0.1 2 4 6 8
April 3.3 9.4 9.2 9.0 8.9 8.6 27.0 30.0 30.0 30.0 30.0May 3.2 14.4 13.3 12.3 11.5 11.0 27.0 28.0 29.0 30.0 30.0June 4.6 16.0 16.7 16.8 16.8 16.8 20.0 21.5 22.0 23.5 23.5August 3.1 19.5 19.7 18.9 17.5 16.7 20.0 21.5 25.0 26.5 26.5
424 D.J. Albert / Journal of Sea Research 64 (2010) 422–425
3.2. Size of tidal change, seawater temperature, salinity, and size ofmedusae
Table 1 shows the size of the tidal changes, seawater temperature,and salinity. Medusae were measured in April and again in May 2009.The average size (±SD) of medusae on April 4 and 5, 2009 was 8.3±2.0 cm (N=102). On May 20, 21, and 22, 2009, the average size was11.1±1.8 cm (N=178). These data indicate that medusae wereregaining bulk lost over the winter (t[278]=12.4, pb0.0001).Ephyrae were in the bay at the end of June and juveniles (about1 cm dia.) were present in early August. Only adults were counted inthe assessments of the vertical distribution of medusae.
4. Discussion
A. labiata did not exhibit a tidally synchronized vertical migration(Fig. 1). Papathanassiou et al. (1987) found that medusae of differentsizes were comparably distributed throughout the water column. Thisalso appeared to be true during the present observations. Ephyraewere present in June and juveniles in early August as has beendescribed (Albert, 2005). These were always found mixed with adultsbut were easily distinguished from adults andwere not counted in theassessments. Accordingly, a non-random distribution of medusae ofdifferent sizes is not confounding the present results. The observationthat Aurelia aurita in the Wadden Sea drifted into the North Seaindicates that A. aurita in that location were also not maintaining theirposition in the estuary by tidally synchronized vertical migration (vander Veer and Oorthuysen, 1985). The scyphozoans C. hysoscella and R.pulmo also did not appear to display a vertically synchronized tidalmigration in the Wadden Sea (Verwey, 1966). The absence of verticalmigration by all four scyphozoans contrasts with observation that thehydrozoans, Rathkea octopuctat, S. tubulosa, B. ramosa and E.maculata,and the ctenophore, P. pileus, were near the surface on flood tides andnear the bottom on ebb tides in the Wadden Sea (van der Veer andSadée, 1984; Kopacz, 1994). The tidally synchronized verticalmigration by these hydrozoans and a ctenophore seemed to enablethem to remain in theWadden Sea. The contrasting observations withscyphozoans and several hydrozoans and a ctenophore raise thepossibility of behavioural differences between these cnidarians.
Vertical spacing by Aurelia sp has been noted in Elefsis Bay(Papathanassiou et al., 1987),Mljet Lakes (Malej et al., 2007), NakaumiLake (Han and Uye, 2009) as well as in Roscoe Bay (Fig. 1). In bothNakaumi Lake and Roscoe Bay (Fig. 1) relatively fewmedusae were inthe top 2 mof thewater column. Neither salinity (21.5 ppt to 30.0 ppt)nor temperature (9.2 °C to 19.7 °C) would have produced thisavoidance of surface water in Roscoe Bay (Albert, 2008). Furthermore,large numbers of A. labiata frequently came right to the surface in thelate afternoon (for similar observations, see Papathanassiou et al.,1987; Hamner et al., 1994, p 354; Albert, 2005, p. 576). The stimulusthat elicitedmoving into the surfacewaterwas not identified, but oncein the top 2 m, medusae in Roscoe Bay typically remained there forseveral hours.
While A. labiata were not using tidally synchronized verticalmigration to stay in Roscoe Bay, tidal currents do influence thehorizontal distribution of medusae in the bay. The effects of currentshave so far not been possible to predict because the current patternsare complex, vary with the size of the tide change and the time duringthe tide change, and the ebb and flood currents are not mirror images(Albert, 2009).
The only known variable that enables A. labiata to have a residentpopulation in Roscoe Bay is its behaviour of diving down into stillwater when individuals are swept over the gravel bar at the entranceto the bay and of swimming up when the current becomes slack andthen turns to flood (Albert, 2007). I have argued that medusae swimdown in response to turbulence as they drift over the gravel bar andthat they stay below the moving ebb stream until the tide changesbecause they sense the vibration from the ebb stream (Albert, 2007).This species' typical behaviour traps A. labiata in the bay. Within thebay, there is no turbulence associated with the ebb tide that causesmedusae to dive down nor is there vibration that would make themstay down in the water column. As a result, medusae in the baycontinue to occupy most of the water column throughout the ebb andflood tides (Fig. 2).
Acknowledgement
Thanks to D. O'Brien for helpful comments and encouragement andto theeditor and twoanonymous reviewers for constructive suggestions.
References
Albert, D.J., 2005. Reproduction and longevity of Aurelia labiata in Roscoe Bay, a smallbay on the Pacific Coast of Canada. J. Mar. Biol. Ass. UK 85, 575–581.
Albert, D.J., 2007. Aurelia labiata medusae (Scyphozoa) in Roscoe Bay avoid tidaldispersion by vertical migration. J. Sea Res. 57, 281–287.
Albert, D.J., 2008. Adaptive behaviours of the jellyfish Aurelia labiata in Roscoe Bay onthe west coast of Canada. J. Sea Res. 59, 198–201.
Albert, D.J., 2009. Aurelia labiata (Scyphozoa) jellyfish in Roscoe Bay: their spatialdistribution varies with population size and their behaviour changes with waterdepth. J. Sea Res. 61, 140–143.
Canadian Hydrographic Service, 2009. Canadian Tide and Current Tables. Juan de FucaStrait and Strait of Georgia, Fisheries and Oceans Canada, Ottawa, Canada.
Creutzberg, F., 1975. Orientation in space: animals. 8.1 Invertebrates. Mar. Ecol.Physiological Mechanisms. Pt. 2, vol. II. J. Wiley and Sons, New York, pp. 555–656.
Dawson, M.N., Hamner, W.M., 2003. Geographic variation and behavioral evolution inmarine plankton: the case of Mastigias (Scyphozoa, Rhizostomeae). Mar. Biol. 143,1161–1174.
Hamner, W.M., Hamner, P.P., Strand, S.W., 1994. Sun-compass migration by Aureliaaurita (Scyphozoa): population retention and reproduction in Saanich Inlet, BritishColumbia. Mar. Biol. 119, 347–356.
Han, C., Uye, S., 2009. Quantification of the abundance and distribution of the commonjellyfish Aurelia aurita s.l. with a Dual-frequency IDentification SONar (DIDSON).J. Plankton Res. 31, 805–814.
Kopacz, U., 1994. Evidence for tidally-induced vertical migration of some gelatinouszooplankton in the Wadden Sea area near Sylt. Helgo. Meeresunters. 48, 333–342.
Lo, W.T., Chen, I.L., 2008. Population succession and feeding of scyphomedusae, Aureliaaurita, in a eutrophic tropical lagoon in Taiwan. Estuar. Coast. Shelf Sci. 76, 227–238.
Magome, S., Yamashita, T., Kohama, T., Kenada, A., Hayami, Y., Takahashi, S., Takeoka, H.,2007. Jellyfish patch formation investigated by aerial photography and drifterexperiment. J. Oceanog. 63, 761–773.
Malej, A., 1989. Behaviour and trophic ecology of the jellyfish Pelagia noctiluca(Forsskhl, 1775). J. Exp. Mar. Biol. Ecol. 126, 259–270.
Malej, A., Turk, V., Lucic, D., Benovic, A., 2007. Direct and indirect trophic interactions ofAurelia sp. (Scyphozoa) in a stratified marine environment (Mljet Lakes, AdriaticSea). Mar. Biol. 151, 827–841.
Naylor, E., 2006. Orientation and navigation in coastal and estuarine zooplankton. Mar.Fresh. Behav. Physiol. 39, 13–24.
Papathanassiou, E., Panayotidis, P., Anagnostaki, K., 1987. Notes on the biology andecology of the jellyfish Aurelia aurita Lam. in Elefsis Bay (Saronikos Gulf, Greece).Mar. Ecol. 8, 49–58.
Purcell, J.E., Brown, E.D., Stokesbury, K.D.E., Haldorson, L.H., Shirley, T.C., 2000.Aggregations of the jellyfish Aurelia labiata: abundance, distribution, associationwith age-0 pollack, and behaviors promoting aggregation in Prince William Sound,Alaska. USA. Mar. Ecol. Prog. Ser. 195, 145–158.
Rakow, K., Graham, W.M., 2006. Orientation and swimming mechanics by thescyphomedusa Aurelia sp. in shear flow. Limnol. Oceanogr. 51, 1097–1106.
Shanks, A.L., Graham, W.M., 1987. Oriented swimming in the jellyfish Stomolopusmeleagris L. Agassiz (Schyphozoan: Rhizostomida). J. Exp. Mar. Biol. Ecol. 108,159–169.

425D.J. Albert / Journal of Sea Research 64 (2010) 422–425
Toyokawa, M., Inagaki, T., Terazaki, M., 1997. Distribution of Aurelia aurita (Linneaus,1758) in Tokyo Bay: observations with echo sounder and plankton net. Proc. 6thInternational Conference on Coelenterate Biology, 1995. NatuurhistorischMuseum,Leiden, pp. 4830–4900.
Trinast, E.M., 1975. Tidal currents and Arcartia distribution in Newport Bay, California.Estuar. Coast. Mar. Sci. 3, 165–176.
van der Veer, H.W., Oorthuysen, W., 1985. Abundance, growth and food demand of thescyphomedusa Aurelia aurita in the western Wadden Sea. Neth. J. Sea Res. 19,28–44.
van der Veer, H.W., Sadée, C.F.M., 1984. The seasonal occurrence of the ctenophorePleurobrachia pileus in the western Dutch Wadden Sea. Mar. Biol. 79, 219–227.
Verwey, J., 1966. The role of some external factors in the vertical migration of marineanimals. Netherl. J. Sea Res. 3, 245–266.
Wooldridge, T., Erasmus, T., 1980. Utilization of tidal currents by estuarine zooplankton.Estuar. Coast. Mar. Sci. 11, 107–114.
Zavodnik, D., 1987. Spatial aggregations of the swarming jellyfish Pelagia noctiluca(scyphozoa). Mar. Biol. 94, 265–269.