vascular medicine - circulationcirc.ahajournals.org/content/124/19/2129.full.pdf · vascular...
TRANSCRIPT

Vascular Medicine
Intravital Microscopy on Atherosclerosis in ApolipoproteinE–Deficient Mice Establishes Microvessels as Major Entry
Pathways for Leukocytes to Advanced LesionsEinar E. Eriksson, MD, PhD
Background—There has been considerable speculation about the role of lesion microvessels in the accumulation ofleukocytes in atherosclerosis. However, direct study of microvascular recruitment of leukocytes in lesions has not beenperformed, and the quantitative role for this route of entry is unclear.
Methods and Results—Here, microvascular recruitment of leukocytes was studied in advanced lesions in 12- to24-month-old apolipoprotein E–deficient (ApoE�/�) mice. Histology and transmission electron microscopy demon-strated the presence of mainly adventitial, but also intimal, microvessels. Interactions between leukocytes andendothelium occurred in lesion venules. Leukocyte rolling was largely P-selectin dependent; however, residual rollingwas mediated by L-selectin and endothelial P-selectin glycoprotein ligand 1. Leukocyte adhesion was significant andwas attenuated in mice treated with antibodies against P-selectin, CD18, or both before preparation for intravitalmicroscopy, suggesting acute activation of these 2 molecules by surgical trauma. Nonetheless, the density of firmlyarrested leukocytes was 100-fold higher in lesion venules compared with the arterial lumen even in mice pretreated withantibodies against P-selectin and CD18, indicating strong recruitment of cells from venules that is unrelated toexperimental manipulation. Fluorescent myelomonocytic cells in ApoE�/� mice carrying a knock-in mutation forenhanced green fluorescent protein (EGFP) in the lysozyme M locus (ApoE�/�/lysMEGFP/EGFP mice) were distributedspecifically around lesion venules, but not around arterioles or capillaries, further indicating ongoing extravasation fromvenules into plaque tissue.
Conclusions—These findings provide strong data for microvascular recruitment of leukocytes in atherosclerosis andindicate roles for L-selectin and P-selectin glycoprotein ligand 1 in this process. (Circulation. 2011;124:2129-2138.)
Key Words: atherosclerosis � cell adhesion molecules � inflammation � macrophages � microcirculation � vasa vasorum
Leukocytes are key players in the development of athero-sclerotic lesions. Inhibition of recruitment of inflamma-
tory cells to plaque introduces an option for treatment ofatherosclerosis and its complications.1,2 Leukocyte invasioninto lesions has traditionally been considered to take placefrom the arterial lumen.3 Consequently, numerous studieshave investigated the activation of arterial endothelium inresponse to atherogenic stimuli, expression of cell adhesionmolecules (CAMs) and chemokines in arteries, and interac-tions between leukocytes and arterial endothelium.4–7 How-ever, although many leukocytes undoubtedly invade lesionsby the arterial route, arterial endothelium does not respondstrongly to inflammatory stimulation, and thus does not favorrecruitment of leukocytes.8,9 In contrast, endothelium on thevenous side of the vascular tree is inflammatory responsiveand provides an efficient recruitment pathway for leukocytesto sites of injury or inflammation.10 Apparently, a venous site
of entry for leukocytes in atherosclerosis might mediate moreefficient trafficking of inflammatory cells.
Clinical Perspective on p 2138As atherosclerotic lesions increase in thickness, a network
of microvessels develops inside lesions.11,12 These vessels areknown to increase in numbers in vulnerable stages of thedisease, and their density has been associated with the onsetof plaque rupture and clinical manifestations of the dis-ease.13,14 However, although inhibition of the buildup ofmicrovessels by treatment with substances blocking angio-genesis limits the development of atherosclerosis in mice,15,16
the causal roles of microvessels in atherogenesis are unclear.It has been suggested that vasa vasori open pathways fortransport of solutes to the vessel wall in coronary arteries,17
and that intraplaque hemorrhage, an event associated withrapid lesion growth and rupture, may be mediated not only bybleeding from the arterial lumen but also through loss of
Received March 8, 2011; accepted September 9, 2011.From the Department of Molecular Medicine and Surgery, Karolinska Institute, Karolinska Hospital, Stockholm, Sweden.The online-only Data Supplement is available with this article at http://circ.ahajournals.org/lookup/suppl/doi:10.1161/CIRCULATIONAHA.
111.030627/-/DC1.Correspondence to Einar Eriksson, MD, PhD, Department of Molecular Medicine and Surgery, Karolinska Institutet CMM L8:03, Karolinska Hospital,
S-171 76 Stockholm, Sweden. E-mail [email protected]© 2011 American Heart Association, Inc.
Circulation is available at http://circ.ahajournals.org DOI: 10.1161/CIRCULATIONAHA.111.030627
2129
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from

microvascular integrity in lesions.18,19 In addition, certainstudies have investigated the possibility that lesion microves-sels could be involved in the delivery of leukocytes toplaque.20 However, the functional characteristics of mi-crovessels in atherosclerotic lesions, including their inflam-matory properties, are unknown, and direct evidence for theroles of microvessels in the accumulation of leukocytes inlesions is missing.
In the present study, various microscopic techniqueswere used to investigate the characteristics of microvesselsin advanced atherosclerotic lesions in apolipoproteinE– deficient (ApoE�/�) mice. The data reveal that venulesin atherosclerosis represent an important entry pathway forleukocytes to plaque. This study establishes a majorrole for lesion microvessels in leukocyte trafficking inatherosclerosis.
MethodsAnimalsOne hundred sixty-eight (male, n�93; female, n�75) ApoE�/� micewere obtained from M&B, Ry, Denmark. ApoE�/�/lysMEGFP/EGFP
mice (EGFP indicates enhanced green fluorescent protein) werecreated as previously described.21 Beginning at 8 weeks of age, theApoE�/� mice were fed a Western diet (Analyzen, Odal, Sweden)based on cornstarch, sucrose, glucose, cocoa butter, cellulose,minerals, and a vitamin mix. The diet contained 0.15% choles-terol and 21% (wt/wt) total fat. Ten C57Bl/6 mice were fedstandard chow. Water was provided ad libitum. The experimentswere approved by the regional ethics committee foranimal experimentation.
Intravital MicroscopyUnder isoflurane anesthesia, catheters were placed in the left carotidartery and the left jugular vein. The abdomen was opened and theaorta was exposed. If atherosclerotic plaque containing microvesselswere found, lesions were exposed for microscopic observation. Theoutmost care was taken not to injure vessels feeding or draining theplaque microvascular network, and if any signs of impaired bloodflow were noticed, the animal was excluded from the study. Theexposed tissue was superfused by buffered saline at 37°C. A pressuretransducer connected to the carotid catheter monitored blood pres-sure. All mice had a mean blood pressure �60 mm Hg with anaverage of �90 mm Hg. Rectal temperature was kept at 37°C withan infrared heat lamp. Blood samples (10 �L) were taken from thetail vein and analyzed for white blood count in a Burker chamber.Microscopic observations were made by use of a Leitz Biomedmicroscope with Nikon WI10 or Leitz SW25 water immersionobjectives. Epi-illumination fluorescence microscopy was startedafter injection of fluorescent beads, the plasma tracer molecule FITCdextran (250 mg/mL), after labeling of circulating leukocytes with anintravenous injection of rhodamine 6G (0.3 mg/mL, 0.67 mg/kg), orafter no treatment in ApoE�/�/lysMEGFP/EGFP mice in which neu-trophils and monocytes are endogenously fluorescent.22 Images wererecorded with a VNC-703 charge-coupled device video camera foroffline analysis. Rolling leukocyte flux was determined as thenumber of leukocytes passing a reference line perpendicular to bloodflow. Leukocyte adhesion was determined as the number of leuko-cytes stably arrested for �60 seconds in individual vessel segments.
In Vivo Detection of CAMsFluorescent beads coated with protein G (2 �m in diameter,Polysciences Inc) were coupled to antibodies by incubating 50 �Lbead solution with 50 �g antibody (1 mg/mL) for 30 minutes at roomtemperature. The solution was washed, resuspended in 1.0 mL Hankbuffered salt solution, and sonicated. Mice were given 300 �L of thebead solution through the carotid artery catheter, and binding of
beads to the endothelium was analyzed after 5 minutes. All bindingof beads to endothelium was compared with beads coated with anirrelevant isotype control antibody in each animal. As for staining ofPSGL-1, binding of beads was measured in the central 100 �m ofvenules in atherosclerotic lesions in which P-selectin–independentrolling was detected. The number of beads was compared with thatof all other observed atherosclerotic lesional venules of similardiameter in the same animal.
Scanning Electron MicroscopyMice were perfused with 2.5% glutaraldehyde through the leftventricle (100 mm Hg for 20 minutes) with outflow through the rightatrium. The aorta was mounted en face on cork and dehydrated inethanol and by critical point dehydration in CO2. After gold sputtercoating, the vessels were examined in a Philips 515 scanningelectron microscopy.
Histology and Transmission Electron MicroscopyAortic segments from perfusion-fixed animals were prepared bystandard protocols described fully in the online-only Data Supple-ment. Specimens were examined in a Philips CM120Twin electronmicroscope at 80 kV. For toluidine blue staining, 1-�m-thicksections were cut, stained, and observed in a Nikon Optiphot-2microscope with Plan 20�/0.50 or 40�/0.85 objectives.
Antibodies and ReagentsMonoclonal antibodies (mAbs) RB40.34 against mouse P-selectinand 4RA10 against mouse PSGL-1 were kindly provided by DietmarVestweber, University of Munster, Germany. The hybridoma formAb GAME46 against mouse CD18 was kindly provided by EdRoos, Netherlands Cancer Institute, Amsterdam, the Netherlands,and antibody was prepared from cell supernatants on a protein Lcolumn. Rhodamine 6G and FITC dextran came from Sigma.FITC-labeled microspheres were obtained from Molecular Probes,and beads coated with protein G came from Polysciences Inc.
Statistical AnalysisThe data represent the mean�SEM of measurements obtained in theindicated number of experiments. Statistical comparison betweenindependent groups was performed with the Student t test or theMann-Whitney rank-sum test; comparison before and after antibodyblockage of CAMs was performed with a paired t test or Wilcoxonsigned-rank test for paired samples. Multiple comparisons wereperformed by 1-way ANOVA or the Kruskal-Wallis 1-way ANOVAon ranks adjusted for multiple comparisons with the Dunn test. Dataon leukocyte rolling before and after antibody treatment wereanalyzed by use of a 2-level conditional hierarchical linear modelwith fixed group effect in which the fixed-effect parameters corre-spond to the means of the groups and the random effects are thelevels of the random effect nested within the levels of the fixed effectaccording to the following equation:
Responseijk�Groupi�Animalj(i)�Segmentk(ij)
The model has 2 variance component parameters to be estimated:the animal-to-animal variability and the segment-to-segment vari-ability within an animal. Data on leukocyte adhesion were calculatedboth by analysis of pooled data on adhesion per 1-mm2 endothelialarea and by a model similar to that described for leukocyte rolling.Post hoc analyses were performed by the Dunnett test. Distributionof these variables was positively skewed, and before the formalanalyses, the variables were log transformed. Statistical significancewas set at P�0.05.
ResultsMicrovessels in atherosclerotic arteries were studied in 12- to24-month-old ApoE�/� mice fed a Western diet from 8weeks of age. Macroscopically visible lesions were present inthe abdominal aorta in 126 of 168 mice, a few of which
2130 Circulation November 8, 2011
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from

carried �1 abdominal plaque. Microvessels were regularlyfound adjacent to plaque (Figure 1A). On gentle dissection,most of these vessels were mobile in relation to the lesion;however, certain more deeply located vessels were immobile,suggesting that they were located closer to or within theplaque tissue. Such vessels, regarded as lesional vessels, werepresent in 109 of 141 observed abdominal plaque (77%). Ingeneral, lesional vessels were found on larger, thicker lesions,whereas small plaque carried few or no microvessels.
Characteristics of Microvessels in AtheroscleroticLesions in ApoE�/� MiceTo study the detailed structure of plaque microvessels inApoE�/� mice, light microscopy and transmission electron
microscopy were performed on control C57Bl/6 mice and on thefirst 39 lesion specimens in which microvessels had beenobserved by stereomicroscopy (Figure 1B through 1E). Therewere no microvessels in the healthy mouse aorta of C57Bl/6mice. Adventitial microvessels were detected in sections from13 of 39 of the studied advanced atherosclerotic lesions. Incontrast, intimal microvessels were found in only 2 of 39 studiedadvanced lesions; in both, maximal intimal thickness exceeded500 �m (Figure IA in the online-only Data Supplement). Thenumber of adventitial vessels increased with lesion progression(Figure IB and IC in the online-only Data Supplement). Notably,the advanced lesions observed could extend for several millime-ters in the abdominal aorta, and thin sections for transmissionelectron microscopy were prepared from only a small area of theplaque. Thus, microvessels located outside the sectioned areawere not visualized, explaining at least some of the differences
Figure 2. Photomontages of images taken from intravital micros-copy experiments in ApoE�/� mice with (A and B) or without (C)treatment with the plasma tracer FITC dextran. Bars�200 �m. InA, a large section of an atherosclerotic lesion in the abdominalaorta containing microvessels is displayed. The lesion, which fullycovered the aorta, is outlined with dashed lines. Note the differ-ence in microvessel density between different parts of the lesion.B, Microvessel sprout in an atherosclerotic lesion (circled). C,Interconnecting loop and vessel dilatation in lesion microves-sels. This image was taken in an ApoE�/� mouse carrying aknock-in mutation for enhanced green fluorescent protein in thelysozyme M locus, rendering monocytes and neutrophils endog-enously fluorescent.10
Figure 1. A, Microvessels in an advanced atherosclerotic lesion inthe abdominal aorta in an ApoE�/� mouse. Most microvessels visi-ble in the image were mobile in relation to the lesion, whereassome were immobile and regarded as true lesional vessels.Bar�250 �m. B, Light microscopy image of a 1-�m section of anormal abdominal aorta in an ApoE�/� mouse stained with tolu-idine blue. The media contains regular layers of smooth musclecells and elastic lamellae. The adventitia is somewhat poorlydefined, and there are no microvessels. Bar�50 �m. C, Intimalmicrovessel in an advanced atherosclerotic lesion. The vessellumen is outlined by an endothelial monolayer. The aortic lumen isseen on the lower right. Bar�50 �m. D, Severe lesion in theabdominal aorta. The aortic lumen is in the upper right (not visible).A mass of necrotic core fills out the lumen. Several microvessels(circled) are present in a densely cellular adventitia. The dashedline indicates the destructed media in which the elastic lamellaeare degraded. Bar�50 �m. E, Electron microscopy image of alesional microvessel. Red cells are visible in the lumen. A pericyte(PC) supports the endothelial cells (ECs). Bar�10 �m.
Eriksson Leukocyte Recruitment From Lesion Microvessels 2131
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from

in the abundance of microvessels observed by stereomicroscopyand transmission electron microscopy. Nonetheless, adventitialmicrovessels seem to dominate over intimal microvessels inadvanced lesions in mice.
Study of the architecture of lesion microvascular networkswas performed by intravital microscopy after injection of theplasma tracer molecule FITC dextran. As shown previ-ously,15,23 microvessel development ranged from isolatedvessels to extensive networks (Figure 2A and Figure IIA andIIB and Video I in the online-only Data Supplement).Lesional vessels showed frequent changes in direction, withvessels running in loops and with abundant intersections(Figure 2A). Moreover, sprouting and vessel dilation, featurescharacteristic of immature or angiogenic vessels,24 were alsoobserved (Figure 2B and 2C).
Rolling of Leukocytes in Plaque Venules IsMediated by P-Selectin, L-Selectin, and PSGL-1We used intravital microscopy to study recruitment of leuko-cytes to atherosclerotic lesions. There were no microvessels inthe macroscopically healthy aortic wall in C57Bl/6 mice or inApoE�/� mice (n�10�22 mice). In parallel, no microvesselswere present in small and thin early lesions (n�23). Conse-quently, no rolling or recruitment occurred from vasa vasori inthese locations. Nonetheless, rolling of leukocytes occurredregularly on aortic endothelium in these plaques. Thus, recruit-ment of leukocytes to early lesions appears to take place throughthe arterial lumen. Data on leukocyte rolling on luminal endo-thelium in lesions were described in detail previously.5
In larger, thicker, and thus more advanced lesions, mi-crovessels were regularly found. Under the intravital micro-scope, there was a difference in appearance between mi-
crovessels that were mobile or immobile in relation to thelesion on surgical manipulation. The immobile vessels werelocated deeper and were found in the same focal plane as theplaque tissue, supporting that they were indeed integrated intolesions. In these vessels, no interactions between leukocytesand endothelium occurred in feeding arterial vessels or incapillaries. However, rolling and adhesion of leukocytes wereabundant in draining venules (Video II in the online-onlyData Supplement). Blockage of P-selectin with mAbRB40.34 reduced rolling to 14�5.4% of rolling beforeinhibition (rolling mean estimate, 14%; 95% confidenceinterval [CI], 2.6–25, representing a significance of P�0.05according to the mixed statistical procedure described inMethods; Figure 3A), suggesting a major role for P-selectinin rolling in plaque microvessels. Residual rolling occurredin a minority of venules and was unaffected by blockage ofE-selectin with mAb 9A9 (Figure 3B; n�5 mice, n�8venules; mean, 110%; 95% CI, 43–283; P�NS). However,inhibition of L-selectin strongly reduced P-selectin–indepen-dent rolling (n�9 mice, n�12 venules; mean, 1.2%; 95% CI,0.19–3.2; P�0.05). P-selectin–independent rolling was notinfluenced by blockage of known lymphoid L-selectin li-gands such as the peripheral node addressin (mAb MECA-79;n�5 mice, n�6 venules; mean, 93%; 95% CI, 38–222;P�NS) or MAdCAM-1 (mAb HECA-367; n�8 mice, n�8venules; mean, 103%; 95% CI, 58–148; P�NS). However,function inhibition of PSGL-1 with mAb 4RA10 abolishedP-selectin–independent rolling, suggesting that PSGL-1 actsas an L-selectin ligand in lesion microvessels (n�12 mice,n�14 venules; mean, 0.26; 95% CI, �0.29–1.24; P�0.05).Importantly, P-selectin–independent rolling occurred in theabsence of leukocytes adherent to the endothelium, suggest-
Figure 3. Leukocyte rolling in venules in atherosclerotic lesions. A, Observation of rolling leukocytes performed before and after intravenousinjection of an antibody against P-selectin (Sel). n�19 animals, n�49 venules. B, Identification of molecules mediating P-selectin–indepen-dent rolling. Shown are data for rolling in venules in which rolling was observed after blockage of P-selectin. Rolling in venules in response toadditional inhibition of the indicated molecules is displayed. Anti–L-selectin, n�9 mice, n�12 venules; anti–peripheral node addressin (PNAd),n�5 mice, n�6 venules; anti-MAdCAM-1, n�8 mice, n�8 venules; anti–P-selectin glycoprotein ligand 1 (PSGL-1), n�12 mice, n�14venules. In A and B, statistical analysis was performed with a 2-level conditional hierarchical linear model with a fixed group effect on dataobtained before and after antibody blockage. For details, see Methods. *The equivalent of P�0.05.
2132 Circulation November 8, 2011
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from

ing that capture and rolling were mediated by interactionsbetween leukocytes and endothelium and not by interactionsbetween leukocytes.
Because P-selectin stored in endothelial cells may berapidly upregulated in response to surgical trauma,25 theimportant role of P-selectin–dependent rolling in plaquemicrovessels exposed to intravital microscopy may not rep-resent a similar importance in leukocyte accumulation inatherosclerosis. To study whether P-selectin was induced bythe surgical procedure, we stained P-selectin before and aftersurgery by fluorescent beads coated with mAb RB40.34.Intravenous injection of coated beads before surgery did notresult in strong binding of beads (Figure 4A1). However,when RB40.34-coated beads were introduced after the tissuehad been prepared for microscopy, beads bound the endothe-lium of drainage venules in high numbers, supporting the ideathat rolling mediated by P-selectin was due partly to experi-mental manipulation (Figure 4A2). Binding of beads coatedwith an irrelevant control antibody was very low (not shown).Importantly, binding of beads occurred only in draining butnot feeding vessels or capillaries in a way similar to that
demonstrated in systemic microvessels (Figure 4B), support-ing that microvascular networks in lesions are built up byarterioles, venules, and capillaries with respective functionssimilar to those seen in other vascular beds.
Expression of PSGL-1 has been demonstrated not only onleukocytes but also on chronically inflamed intestinal endo-thelium. Because blockage of PSGL-1 was found to inhibitL-selectin–dependent rolling in plaque venules, we alsostained for this molecule in lesion venules. Indeed, binding ofmAb 2PH1-coated beads was more prominent in segments ofvenules in which P-selectin–independent rolling was ob-served than in the segments in which no such interactionstook place (2.1�0.35 versus 0.27�0.82 beads per 100-�mvenular length; P�0.05 by t test; n�5 mice). Thus, the datasupport that PSGL-1 acts as an endothelial ligand forL-selectin in microvessels in lesions.
Firm Adhesion of Leukocytes in Lesion VenulesWe subsequently sought to investigate whether firm adhesionoccurs in venules and whether adhesion, like rolling, wasstimulated by mechanisms induced by the surgical procedure.
Figure 4. Staining of P-selectin inmicrovessels in atherosclerotic lesions.Staining was performed by injection offluorescent beads coated with the anti-body RB40.34 against mouse P-selectin.A1, Injection of beads was performed 15minutes before surgical manipulation.Bar�200 �m. A2, Staining was repeatedin the same preparation as in A1 but thistime 15 minutes after surgical exposure ofthe abdominal aorta. Images in A1 andA2 are representative of 5 individualexperiments. B, Staining of a separatelesion was performed after manipulation.A indicates arterioles; C, capillaries; andV, venules. White arrows indicate arterialflow; black arrows, venular flow. Becauseof the 3-dimensional structure of theaorta, some vessels are somewhat out offocus. Regardless, the difference inexpression of P-selectin in distinct typesof microvessels is apparent.Bar�200 �m.
Eriksson Leukocyte Recruitment From Lesion Microvessels 2133
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from

Indeed, firmly arrested cells were visible in venules in plaque.Moreover, antibody blockage of P-selectin with RB40.34reduced adhesion compared with when this mAb was givenduring microscopy (P�0.05; statistical significance was ob-tained with both the mixed-model approach outlined in theMethods section and the t test on pooled data from controland antibody-treated mice; Figure 5). These data indicate thatfirm adhesion was increased by surgical manipulation. Asimilar tendency was observed after treatment with mAbGAME-46 against the common �2 integrin subunit CD18;however this was not fully significant (t test pooled data,P�0.032; mixed model raw, P�0.027; mixed model adjustedby the Dunnett test, P�0.078). Combined blockage of bothmolecules resulted in no additional inhibition of adhesion,indicating that P-selectin and CD18 mediate adhesion of the
same population of cells (adhesion, P�0.05 versus control).Nonetheless, a significant number of adherent leukocyteswere observed in plaque venules even in experiments inwhich P-selectin and CD18 were blocked before surgery.Because no leukocytes initiated firm arrest without priorrolling and because firm arrest after P-selectin–independentrolling was rarely observed during the time of microscopicobservation, the data indicate that at least a majority of firmlyarrested leukocytes observed in these experiments had ad-hered before inhibition of P-selectin and CD18 and thusbefore experimental manipulation. Thus, leukocyte adhesionto venular endothelium in mice receiving antibodies againstP-selectin and CD18 before surgery represents an ongoingrecruitment of leukocytes to the atherosclerotic vascular wall.
Venules in Atherosclerosis Are Major EntryPathways for Leukocytes to LesionsGiven the above findings, it is plausible that leukocyteadhesion in venules in atherosclerotic lesions seen whenP-selectin and/or CD18 were blocked before experimentalmanipulation represents an ongoing recruitment of cells. Tostudy whether this recruitment makes a quantitatively impor-tant contribution to the overall recruitment of leukocytes, wecompared adhesion in lesional venules detected by intravitalmicroscopy with adhesion to arterial endothelium coveringplaque as studied by scanning electron microscopy. In ourintravital microscopy experiments, endothelial surface area ofplaque venules was 0.22�0.055 mm2, whereas aortic luminalarea was 0.88�0.077 mm2 in the same plaque specimensprepared for scanning electron microscopy, thus representinga roughly 4-fold-larger luminal endothelial area (Figure 6A).As demonstrated above, adhesion after blockage of P-selectinand CD18 before surgery likely represents ongoing recruit-ment of leukocytes during the course of atherogenesis.Interestingly, in a comparison of the number of adherentleukocytes per 1-mm2 surface area after this treatment,leukocyte adhesion was markedly (�100-fold) higher inlesion venules compared with the arterial lumen. Thus, thedata indicate a strong potential for recruitment from venulesin plaque (Figure 6B).
To investigate whether adhesion of leukocytes in lesionvenules is followed by invasion into plaque tissue, weanalyzed extravasation of EGFP-labeled myelomonocytic
Figure 5. Leukocyte adhesion in lesion venules. Inhibition ofP-selectin or CD18 was performed by intravenous injection ofthe function-blocking antibodies RB40.34 (P-selectin) orGAME46 (CD18) 15 minutes before surgical manipulation. Con-trol, n�19 mice, n�74 venules; P-selectin blockage, n�12mice, n�102 venules; CD18 blockage, n�6 mice, n�34venules; P-selectin�CD18 blockage, n�5 mice, n�21 venules.Statistical analysis was performed by comparing mean adhesiondensity in individual animals by use of the t test and a mixed-effect model similar to that used in Figure 3. *Adjusted P�0.05by the Dunnett test. Raw and adjusted P values for GAME-46with the mixed effect model were 0.027 and 0.078, respectively.
Figure 6. Endothelial surface area andleukocyte adhesion in atherosclerosis. A,Lesion endothelial surface area in the ar-terial lumen (n�30 plaque) and in plaquevenules (n�16 plaque) in individuallesions. B, Data on leukocyte adhesionper 1-mm2 endothelial surface in the arte-rial lumen as measured by scanning elec-tron microscopy and in venules measuredby intravital microscopy (P�0.01 by t test;only 1 lesion in each animal was studied).
2134 Circulation November 8, 2011
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from

leukocytes in 12- to 24-month-old ApoE�/�/lysMEGFP/EGFP
mice26 by studying the spatial distribution of leukocytes inrelation to individual microvessels. To minimize bias byextravasation from adjacent vessels, we selected those thatwere as isolated from other vessels as possible. As shown inFigure 7A through 7C, extravasated leukocytes were locatedpredominantly around plaque venules but not around arteri-oles or capillaries. Hence, the data indicate that lesion venulesrepresent a major recruitment pathway for leukocytes tolesions.
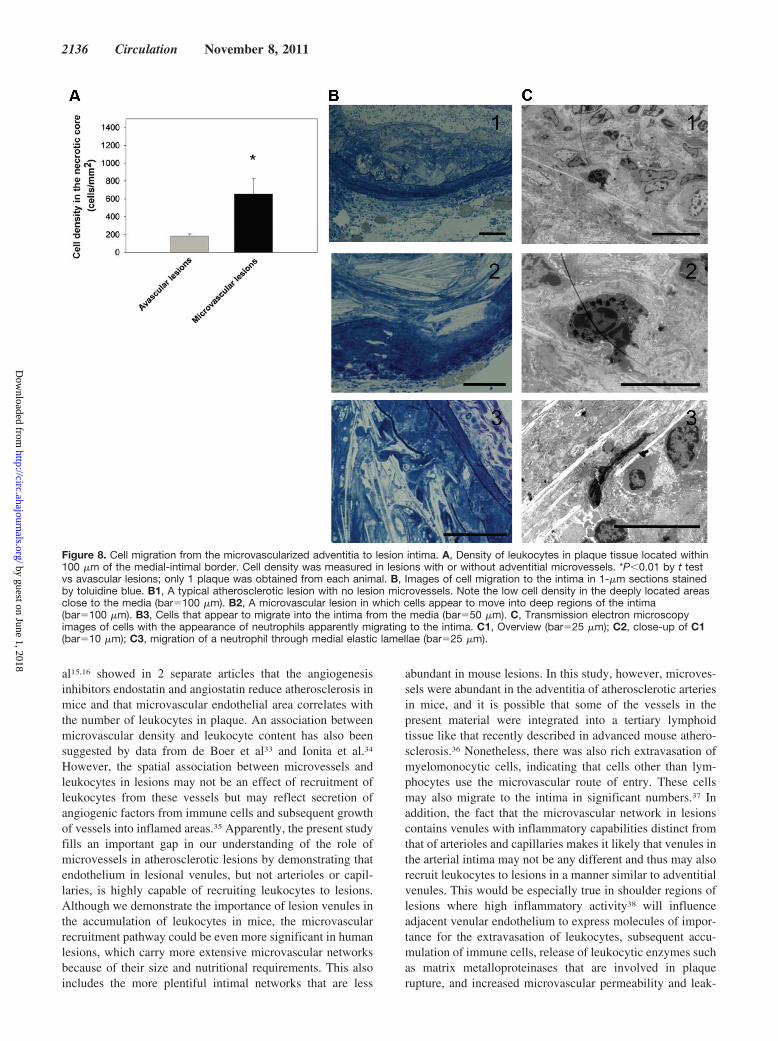
To investigate whether adventitial microvessels couldserve as a conduit for leukocytes to the intima of atheroscle-rotic lesions, we analyzed the density of leukocytes in lesionsin 1-�m sections stained with toluidine blue and by transmis-sion electron microscopy. In these plaques, all of whichexceeded �200 �m in thickness, cellular density in theintima was low in deeply located regions of the necrotic corewithin 100 �m of the intimal/medial border (n�39; P�0.05).In lesions that were lacking microvessels, the average cellulardensity in these areas was 183�26 cells per 1 mm2 (26 mice).However, cellular density in lesions carrying microvesselswas significantly higher (654�176 cells per 1 mm2; 13 mice;P�0.05; Figure 8A), indicating that the presence of adventi-tial microvessels open a gateway for leukocytes to plaqueintima. Images of migrating cells on the adventitial/intimalborder are shown in Figure 8B and 8C.
DiscussionInvasion of leukocytes in the arterial wall is a key mechanismin the development of atherosclerotic lesions and in thetransition from stable to vulnerable plaque. Over the years,there has been considerable speculation about the role forleukocyte entry from lesional microvessels.27 However, al-though such vessels have been demonstrated in associationwith inflammation in lesions, conclusive evidence for theimportance of the microvascular network in the invasion ofleukocytes is missing.
In the present study, light, fluorescence, and electron andintravital microscopy were used to study microvessels inadvanced atherosclerotic lesions in ApoE�/� mice. The datashow that lesion vasa vasori make up complete microvascularnetworks with arterioles, capillaries, and venules and thatnetworks are somewhat irregular, with some vessel featurestypical of ongoing angiogenesis. In contrast, less advancedlesions carry no microvessels. Nonetheless, interactions be-tween leukocytes and endothelium occur in the arteriallumen.5 Combined, these data indicate that, at least in thismouse model, leukocytes gain entry from the arterial endo-thelium during early stages of the disease. However, asmicrovascular networks develop, rolling, adhesion and ex-travasation will start to occur spontaneously in lesion venules.Strikingly, venules in atherosclerosis represent a recruitmentpathway that, in our experiments, was �100-fold moreefficient per endothelial area than invasion from the arteriallumen. Hence, once microvascular networks have developedinside lesions, leukocytes will likely use venules as theirprimary site of entry.
Neovessels in atherosclerotic lesions have received in-creasing interest with regard to their potential roles inatherogenesis. The mechanisms by which microvessels havebeen hypothesized to influence atherogenesis include deliv-ery of proatherogenic solutes and lipoproteins,28 contributionto plaque hemorrhage,29 and recruitment of inflammatorycells to plaque.30 Studies investigating a possible role forlesional microvessels in the invasion of leukocytes have beenpresented previously. O’Brien et al31,32 showed that neovesselendothelium in lesions from human coronary arteries ex-presses high levels of intercellular adhesion molecule-1,vascular cell adhesion molecule-1, and E-selectin. Moulton et
Figure 7. Leukocyte adhesion and extravasation in microvesselsin ApoE/lysMEGFP/EGFP mice. Fluorescent monocytes and neu-trophils adherent to endothelium and extravasated into plaquetissue were visualized by intravital microscopy. The number ofcells located within and at various distances from microvesselswas calculated in 100-�m vessel segments. A through C, Datafrom arterioles (n�22), capillaries (n�22), and venules (n�50),respectively.
Eriksson Leukocyte Recruitment From Lesion Microvessels 2135
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from
al15,16 showed in 2 separate articles that the angiogenesisinhibitors endostatin and angiostatin reduce atherosclerosis inmice and that microvascular endothelial area correlates withthe number of leukocytes in plaque. An association betweenmicrovascular density and leukocyte content has also beensuggested by data from de Boer et al33 and Ionita et al.34
However, the spatial association between microvessels andleukocytes in lesions may not be an effect of recruitment ofleukocytes from these vessels but may reflect secretion ofangiogenic factors from immune cells and subsequent growthof vessels into inflamed areas.35 Apparently, the present studyfills an important gap in our understanding of the role ofmicrovessels in atherosclerotic lesions by demonstrating thatendothelium in lesional venules, but not arterioles or capil-laries, is highly capable of recruiting leukocytes to lesions.Although we demonstrate the importance of lesion venules inthe accumulation of leukocytes in mice, the microvascularrecruitment pathway could be even more significant in humanlesions, which carry more extensive microvascular networksbecause of their size and nutritional requirements. This alsoincludes the more plentiful intimal networks that are less
abundant in mouse lesions. In this study, however, microves-sels were abundant in the adventitia of atherosclerotic arteriesin mice, and it is possible that some of the vessels in thepresent material were integrated into a tertiary lymphoidtissue like that recently described in advanced mouse athero-sclerosis.36 Nonetheless, there was also rich extravasation ofmyelomonocytic cells, indicating that cells other than lym-phocytes use the microvascular route of entry. These cellsmay also migrate to the intima in significant numbers.37 Inaddition, the fact that the microvascular network in lesionscontains venules with inflammatory capabilities distinct fromthat of arterioles and capillaries makes it likely that venules inthe arterial intima may not be any different and thus may alsorecruit leukocytes to lesions in a manner similar to adventitialvenules. This would be especially true in shoulder regions oflesions where high inflammatory activity38 will influenceadjacent venular endothelium to express molecules of impor-tance for the extravasation of leukocytes, subsequent accu-mulation of immune cells, release of leukocytic enzymes suchas matrix metalloproteinases that are involved in plaquerupture, and increased microvascular permeability and leak-
Figure 8. Cell migration from the microvascularized adventitia to lesion intima. A, Density of leukocytes in plaque tissue located within100 �m of the medial-intimal border. Cell density was measured in lesions with or without adventitial microvessels. *P�0.01 by t testvs avascular lesions; only 1 plaque was obtained from each animal. B, Images of cell migration to the intima in 1-�m sections stainedby toluidine blue. B1, A typical atherosclerotic lesion with no lesion microvessels. Note the low cell density in the deeply located areasclose to the media (bar�100 �m). B2, A microvascular lesion in which cells appear to move into deep regions of the intima(bar�100 �m). B3, Cells that appear to migrate into the intima from the media (bar�50 �m). C, Transmission electron microscopyimages of cells with the appearance of neutrophils apparently migrating to the intima. C1, Overview (bar�25 �m); C2, close-up of C1(bar�10 �m); C3, migration of a neutrophil through medial elastic lamellae (bar�25 �m).
2136 Circulation November 8, 2011
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from

age of plasma constituents such as inflammatory lipoproteins,antibodies, or complement factors. Hence, a positive feed-back loop of inflammation and degradation would develop inareas with the highest density of microvessels. Indeed, asshown previously, plaque rupture is increased in areas withabundant microvessels.39,40 In parallel, systemic inflamma-tion also triggers acute events related to atherosclerosis,41
which may be enhanced by the presence of venules withstrong inflammatory properties inside lesions. However, be-cause vasa vasori are rare in the normal mouse aorta andintimal vessels are absent in healthy human arteries, the effectof leukocyte invasion from lesion venules will probably be ofimportance primarily at later stages during the course of thedisease. Previous data indicate that there is a continuousbuildup of neovessels in advanced lesions, which is at leastpartly dependent on the hypoxia-inducible factor and vascularendothelial growth factor pathways.42,43 In the present mate-rial, the architecture of the microvascular network in mouselesions indeed revealed signs of ongoing angiogenesis such asfrequent vessel turns, vessel dilatation, arteriovenous shunts,and occasional vessel sprouts44 that are scarce in establishedtissues. Because previously presented data have shown thatmicrovessels in human plaque start to develop at a lesionthickness �500 �m regardless of disease stage45 and becauseprevious data and those presented here indicate that mi-crovessels in mouse lesions develop at intimal depths similarto those in humans,15,16 these data collectively support thatformation of lesion microvessels is driven mainly by hypoxiaand nutritional demands at significant intimal depths, notprimarily by secretion of factors by cells in plaque at a certainphase of disease progress. In fact, it is possible that growth ofmicrovessels into lesions is a prerequisite for plaque progres-sion above a certain lesion thickness, and inhibition ofmicrovessel angiogenesis may thus be a therapeutic target. Tothis end, the data in the present study indicate that thereduction in lesion progress caused by angiogenesis inhibitorsmight be dependent not only on intervention of the supply ofoxygen and nutrients to lesions but also on attenuatedaccumulation of leukocytes.
ConclusionsThis study investigates leukocyte trafficking in microvesselsin atherosclerotic lesions. The data demonstrate that venulesin atherosclerosis represent a major entry pathway for leuko-cytes to lesions that, in advanced stages of atherogenesis, mayoutnumber recruitment from the arterial lumen. These find-ings establish the development of microvessels as a keyelement in the development of atherosclerosis.
Sources of FundingThis work was supported by the Swedish Heart and Lung Founda-tion, the Swedish Research Council, the Swedish Society of Medi-cine, the Swedish Society for Medical Research, the Osterman Fund,the Tore Nilson Foundation, the Lars Hierta Memorial Fund, theAFA Health Fund, AstraZeneca, and Karolinska Institutet.
DisclosuresNone.
References1. Hansson GK. Inflammation, atherosclerosis, and coronary artery disease.
N Engl J Med. 2005;352:1685–1695.2. Weber C, Zernecke A, Libby P. The multifaceted contributions of leu-
kocyte subsets to atherosclerosis: lessons from mouse models. Nat RevImmunol. 2008;8:802–815.
3. Faggiotto A, Ross R, Harker L. Studies of hypercholesterolemia in thenonhuman primate, I: changes that lead to fatty streak formation.Arteriosclerosis. 1984;4:323–340.
4. Navab M, Berliner JA, Watson AD, Hama SY, Territo MC, Lusis AJ,Shih DM, Van Lenten BJ, Frank JS, Demer LL, Edwards PA, FogelmanAM. The yin and yang of oxidation in the development of the fatty streak:a review based on the 1994 George Lyman Duff Memorial Lecture.Arterioscler Thromb Vasc Biol. 1996;16:831–842.
5. Eriksson EE, Xie X, Werr J, Thoren P, Lindbom L. Direct viewing ofatherosclerosis in vivo: plaque invasion by leukocytes is initiated by theendothelial selectins. FASEB J. 2001;15:1149–1157.
6. Eriksson EE. Leukocyte recruitment to atherosclerotic lesions, a complexweb of dynamic cellular and molecular interactions. Curr Drug TargetsCardiovasc Haematol Disord. 2003;3:309–325.
7. Zernecke A, Shagdarsuren E, Weber C. Chemokines in atherosclerosis:an update. Arterioscler Thromb Vasc Biol. 2008;28:1897–1908.
8. Ley K, Gaehtgens P. Endothelial, not hemodynamic, differences areresponsible for preferential leukocyte rolling in rat mesenteric venules.Circ Res. 1991;69:1034–1041.
9. Kunkel EJ, Jung U, Ley K. TNF-alpha induces selectin-mediated leu-kocyte rolling in mouse cremaster muscle arterioles. Am J Physiol. 1997;272:H1391–H1400.
10. Eriksson EE, Karlof E, Lundmark K, Rotzius P, Hedin U, Xie X.Powerful inflammatory properties of large vein endothelium in vivo.Arterioscler Thromb Vasc Biol. 2005;25:723–728.
11. Langheinrich AC, Michniewicz A, Bohle RM, Ritman EL. Vasa vasorumneovascularization and lesion distribution among different vascular bedsin ApoE-/-/LDL-/- double knockout mice. Atherosclerosis. 2007;191:73–81.
12. Moreno PR, Purushothaman KR, Sirol M, Levy AP, Fuster V. Neovas-cularization in human atherosclerosis. Circulation. 2006;113:2245–2252.
13. Ritman EL, Lerman A. The dynamic vasa vasorum. Cardiovasc Res.2007;75:649–658.
14. Fleiner M, Kummer M, Mirlacher M, Sauter G, Cathomas G, Krapf R,Biedermann BC. Arterial neovascularization and inflammation in vul-nerable patients: early and late signs of symptomatic atherosclerosis.Circulation. 2004;110:2843–2850.
15. Moulton KS, Vakili K, Zurakowski D, Soliman M, Butterfield C, SylvinE, Lo KM, Gillies S, Javaherian K, Folkman J. Inhibition of plaqueneovascularization reduces macrophage accumulation and progression ofadvanced atherosclerosis. Proc Natl Acad Sci U S A. 2003;100:4736–4741.
16. Moulton KS, Heller E, Konerding MA, Flynn E, Palinski W, Folkman J.Angiogenesis inhibitors endostatin or TNP-470 reduce intimal neovascu-larization and plaque growth in apolipoprotein E-deficient mice.Circulation. 1999;99:1726–1732.
17. Gossl M, Beighley PE, Malyar NM, Ritman EL. Role of vasa vasorum intransendothelial solute transport in the coronary vessel wall: a study withcryostatic micro-CT. Am J Physiol Heart Circ Physiol. 2004;287:H2346–H2351.
18. Levy AP, Moreno PR. Intraplaque hemorrhage. Curr Mol Med. 2006;6:479–488.
19. Kolodgie FD, Gold HK, Burke AP, Fowler DR, Kruth HS, Weber DK,Farb A, Guerrero LJ, Hayase M, Kutys R, Narula J, Finn AV, Virmani R.Intraplaque hemorrhage and progression of coronary atheroma. N EnglJ Med. 2003;349:2316–2325.
20. Yilmaz A, Lipfert B, Cicha I, Schubert K, Klein M, Raithel D, DanielWG, Garlichs CD. Accumulation of immune cells and high expression ofchemokines/chemokine receptors in the upstream shoulder of athero-sclerotic carotid plaques. Exp Mol Pathol. 2007;82:245–255.
21. Rotzius P, Soehnlein O, Kenne E, Lindbom L, Nystrom K, Thams S,Eriksson EE. ApoE(-/-)/lysozyme M(EGFP/EGFP) mice as a versatilemodel to study monocyte and neutrophil trafficking in atherosclerosis.Atherosclerosis. 2009;202:111–118.
22. Faust N, Varas F, Kelly LM, Heck S, Graf T. Insertion of enhanced greenfluorescent protein into the lysozyme gene creates mice with greenfluorescent granulocytes and macrophages. Blood. 2000;96:719–726.
23. Langheinrich AC, Michniewicz A, Sedding DG, Walker G, Beighley PE,Rau WS, Bohle RM, Ritman EL. Correlation of vasa vasorum neovas-
Eriksson Leukocyte Recruitment From Lesion Microvessels 2137
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from

cularization and plaque progression in aortas of apolipoprotein E(-/-)/low-density lipoprotein(-/-) double knockout mice. Arterioscler Thromb VascBiol. 2006;26:347–352.
24. Zacchigna S, Tasciotti E, Kusmic C, Arsic N, Sorace O, Marini C,Marzullo P, Pardini S, Petroni D, Pattarini L, Moimas S, Giacca M,Sambuceti G. In vivo imaging shows abnormal function of vascularendothelial growth factor-induced vasculature. Hum Gene Ther. 2007;18:515–524.
25. Mayadas TN, Johnson RC, Rayburn H, Hynes RO, Wagner DD. Leu-kocyte rolling and extravasation are severely compromised in P selectin-deficient mice. Cell. 1993;74:541–554.
26. Rotzius P. Thams S, Sohnlein O, Kenne E, Tseng CN, Bjorkstrom N,Malmberg KJ, Lindbom L, Eriksson EE. Distinct infiltration of neu-trophils in lesion shoulders in ApoE-/- mice. Am J Pathol. 2010;177:493–500.
27. Hayden MR, Tyagi SC. Vasa vasorum in plaque angiogenesis, metabolicsyndrome, type 2 diabetes mellitus, and atheroscleropathy: a malignanttransformation. Cardiovasc Diabetol. 2004;3:1.
28. Zhu XY, Bentley MD, Chade AR, Ritman EL, Lerman A, Lerman LO.Early changes in coronary artery wall structure detected by micro-computed tomography in experimental hypercholesterolemia. Am JPhysiol Heart Circ Physiol. 2007;293:H1997–H2003.
29. Kockx MM, Cromheeke KM, Knaapen MW, Bosmans JM, De MeyerGR, Herman AG, Bult H. Phagocytosis and macrophage activation asso-ciated with hemorrhagic microvessels in human atherosclerosis.Arterioscler Thromb Vasc Biol. 2003;23:440–446.
30. Doyle B, Caplice N. Plaque neovascularization and antiangiogenictherapy for atherosclerosis. J Am Coll Cardiol. 2007;49:2073–2080.
31. O’Brien KD, Allen MD, McDonald TO, Chait A, Harlan JM, Fishbein D,McCarty J, Ferguson M, Hudkins K, Benjamin CD, Lobb R, Alpers CE.Vascular cell adhesion molecule-1 is expressed in human coronary ath-erosclerotic plaques: implications for the mode of progression ofadvanced coronary atherosclerosis. J Clin Invest. 1993;92:945–951.
32. O’Brien KD, McDonald TO, Chait A, Allen MD, Alpers CE. Neovascularexpression of E-selectin, intercellular adhesion molecule-1, and vascularcell adhesion molecule-1 in human atherosclerosis and their relation tointimal leukocyte content. Circulation. 1996;93:672–682.
33. de Boer OJ, van der Wal AC, Teeling P, Becker AE. Leukocyterecruitment in rupture prone regions of lipid-rich plaques: a prominentrole for neovascularization? Cardiovasc Res. 1999;41:443–449.
34. Ionita MG, van den Borne P, Catanzariti LM, Moll FL, de Vries JP,Pasterkamp G, Vink A, de Kleijn DPV. High neutrophil numbers inhuman carotid atherosclerotic plaques are associated with characteristics
of rupture-prone lesions. Arterioscler Thromb Vasc Biol. 2010;30:1842–1848.
35. Moulton KS. Angiogenesis in atherosclerosis: gathering evidence beyondspeculation. Curr Opin Lipidol. 2006;17:548–555.
36. Grabner R, Lotzer K, Dopping S, Hildner M, Radke D, Beer M,Spanbroek R, Lippert B, Reardon CA, Getz GS, Fu YX, Hehlgans T,Mebius RE, van der Wall M, Kruspe D, Englert C, Lovas A, Hu D,Randolph GJ, Weih F, Habenicht AJ. Lymphotoxin (beta) receptor sig-naling promotes tertiary lymphoid organogenesis in the aorta adventitia ofaged ApoE-/- mice. J Exp Med. 2009;206:233–248.
37. van der Wal AC, Becker AE, Das PK. Medial thinning and atheroscle-rosis: evidence for involvement of a local inflammatory effect.Atherosclerosis. 1993;103:55–64.
38. Schmeisser A, Marquetant R, Illmer T, Graffy C, Garlichs CD, Bockler D,Menschikowski D, Braun-Dullaeus R, Daniel WG, Strasser RH. Theexpression of macrophage migration inhibitory factor 1alpha (MIF 1alpha) inhuman atherosclerotic plaques is induced by different proatherogenicstimuli and associated with plaque instability. Atherosclerosis. 2005;178:83–94.
39. Moreno PR, Purushothaman KR, Fuster V, Echeverri D, Truszczynska H,Sharma SK, Badimon JJ, O’Connor WN. Plaque neovascularization isincreased in ruptured atherosclerotic lesions of human aorta: implicationsfor plaque vulnerability. Circulation. 2004;110:2032–2038.
40. Virmani R, Kolodgie FD, Burke AP, Finn AV, Gold HK, Tulenko TN,Wrenn SP, Narula J. Atherosclerotic plaque progression and vulnerabilityto rupture: angiogenesis as a source of intraplaque hemorrhage.Arterioscler Thromb Vasc Biol. 2005;25:2054–2061.
41. Smeeth L, Thomas SL, Hall AJ, Hubbard R, Farrington P, Vallance P.Risk of myocardial infarction and stroke after acute infection or vacci-nation. N Engl J Med. 2004;351:2611–2618.
42. Vink A, Schoneveld AH, Lamers D, Houben AJ, van der Groep P, vanDiest PJ, Pasterkamp G. HIF-1 alpha expression is associated with anatheromatous inflammatory plaque phenotype and upregulated in acti-vated macrophages. Atherosclerosis. 2007;195:e69–e75.
43. Sluimer JC, Gasc JM, van Wanroij JL, Kisters N, Groeneweg M,Sollewijn Gelpke MD, Cleutjens JP, van den Akker LH, Corvol P,Wouters BG, Daemen MJ, Bijnens AP. Hypoxia, hypoxia-inducible tran-scription factor, and macrophages in human atherosclerotic plaques arecorrelated with intraplaque angiogenesis. J Am Coll Cardiol. 2008;51:1258–1265.
44. Gee MS, Procopio WN, Makonnen S, Feldman MD, Yeilding NM, LeeWM. Tumor vessel development and maturation impose limits on theeffectiveness of anti-vascular therapy. Am J Pathol. 2003;162:183–193.
45. Geiringer E. Intimal vascularization and atherosclerosis. J Pathol Bac-teriol. 1951;63:201–211.
CLINICAL PERSPECTIVEThe invasion of leukocytes in atherosclerosis has been thought to take place mainly from the arterial lumen. However, therehas been considerable speculation regarding the potential role of lesion microvessels in the recruitment of inflammatorycells to plaques. Development of a microvascular network in lesions has been associated with the transition of stable tovulnerable plaques and thus the onset of symptomatic disease. However, there has been no direct study of leukocyterecruitment from lesion microvessels, and the quantitative role of this route of entry is unclear. In this study, adventitialmicrovessels in advanced lesions in atherosclerotic ApoE�/� mice were studied with intravital microscopy. The data showthat there is no microvascular recruitment of leukocytes during early stages of the disease in mice. Inflammatory cells areinstead recruited from the arterial lumen. However, once microvascular networks develop inside lesions, venules have acapacity to recruit leukocytes that are 100-fold stronger per endothelial area than recruitment from the arterial lumen.Recruitment involves the cell adhesion molecules P-selectin, L-selectin, and P-selectin glycoprotein ligand 1. Furthermore,studies of leukocyte extravasation from lesion venules and comparison of leukocyte density in lesions with or withoutadventitial microvessels support that these vessels contribute to overall cell recruitment to plaques. Taken together, thesedata establish microvessels in atherosclerosis as important entry pathways for leukocytes to lesions.
2138 Circulation November 8, 2011
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from

Einar E. ErikssonMicrovessels as Major Entry Pathways for Leukocytes to Advanced Lesions
Deficient Mice Establishes−Intravital Microscopy on Atherosclerosis in Apolipoprotein E
Print ISSN: 0009-7322. Online ISSN: 1524-4539 Copyright © 2011 American Heart Association, Inc. All rights reserved.
is published by the American Heart Association, 7272 Greenville Avenue, Dallas, TX 75231Circulation doi: 10.1161/CIRCULATIONAHA.111.030627
2011;124:2129-2138; originally published online October 10, 2011;Circulation.
http://circ.ahajournals.org/content/124/19/2129World Wide Web at:
The online version of this article, along with updated information and services, is located on the
http://circ.ahajournals.org/content/suppl/2011/10/07/CIRCULATIONAHA.111.030627.DC1Data Supplement (unedited) at:
http://circ.ahajournals.org//subscriptions/
is online at: Circulation Information about subscribing to Subscriptions:
http://www.lww.com/reprints Information about reprints can be found online at: Reprints:
document. Permissions and Rights Question and Answer this process is available in the
click Request Permissions in the middle column of the Web page under Services. Further information aboutOffice. Once the online version of the published article for which permission is being requested is located,
can be obtained via RightsLink, a service of the Copyright Clearance Center, not the EditorialCirculationin Requests for permissions to reproduce figures, tables, or portions of articles originally publishedPermissions:
by guest on June 1, 2018http://circ.ahajournals.org/
Dow
nloaded from

Supplemental Material

Supplemental Methods
Animals: 168 male (93) and female (75) ApoE deficient mice (ApoE-/-) were obtained
from M&B, Ry, Denmark. ApoE-/-/lysMEGFP/EGFP mice were created as previously described
(21). ApoE-/- mice were from 8 weeks of age fed western diet (Analyzen, Odal, Sweden)
based on cornstarch, sucrose, glucose, cocoa butter, cellulose, minerals and a vitamin mix.
The diet contained 0.15% cholesterol and 21% (wt/wt) total fat. 10 C57Bl/6 mice were fed
standard chow. Water was provided ad libitum. The experiments were approved by the
regional ethical committee for animal experimentation.
Intravital microscopy: Under isoflurane anesthesia, catheters were placed in the left
carotid artery and the left jugular vein. The abdomen was opened and the aorta was exposed.
If atherosclerotic plaque containing microvessels were found, lesions were exposed for
microscopic observation. The outmost care was taken not to injure vessels feeding or draining
the plaque microvascular network and if any signs of impaired blood flow were noticed, the
animal was excluded from the study. The exposed tissue was superfused by buffered saline at
37°C. A pressure transducer connected to the carotid catheter monitored blood pressure. All
mice had mean blood pressure (BP) >60mmHg with an average of ∼90mmHg. Rectal
temperature was kept at 37°C with an infrared heat lamp. Blood samples (10µl) were taken
from the tail vein and analyzed for white blood count (WBC) in a Bürker chamber.
Microscopic observations were made using a Leitz Biomed microscope with Nikon WI10 or
Leitz SW25 water immersion objectives. Epi-illumination fluorescence microscopy was
started after injection of fluorescent beads, the plasma tracer molecule FITC dextran
(250mg/ml), after labeling of circulating leukocytes with an intravenous injection of
rhodamine 6G (0.3 mg/ml, 0.67 mg/kg) or following no treatment in ApoE-/-/lysMEGFP/EGFP
mice in which neutrophils and monocytes are endogenously fluorescent (22). Images were
recorded using a VNC-703 CCD video camera for offline analysis. Rolling leukocyte flux

(RLF) was determined as the number of leukocytes passing a reference line perpendicular to
blood flow. Leukocyte adhesion was determined as the number of leukocytes stably arrested
for >60 seconds in individual vessel segments.
In vivo detection of CAMs: Fluorescent beads coated with protein G (2µm in
diameter, Polysciences Inc.) were coupled to antibodies by incubating 50µl of bead solution
with 50µg of antibody (1mg/ml) for 30 minutes at room temperature. The solution was
washed, resuspended in 1.0ml of HBSS and sonicated. Mice were given 300µl of the bead
solution through the carotid artery catheter and binding of beads to the endothelium was
analyzed after 5 minutes. All binding of beads to endothelium was compared with beads
coated with an irrelevant isotype control antibody in each animal. As for staining of PSGL-1,
binding of beads was measured in the central 100µm of venules in atherosclerotic lesions in
which P-selectin-independent rolling was detected. The number of beads was compared with
that of all other observed venules of similar diameter in the same animal.
Scanning electron microscopy (SEM): Mice were perfused with 2.5% glutaraldehyde
through the left ventricle (100mmHg, 20 min) with outflow through the right atrium. The
aorta was mounted en face on cork, and dehydrated in ethanol and by critical point
dehydration in CO2. After gold sputter coating the vessels were examined in a Philips SEM
515 microscope.
Histology and transmission electron microscopy (TEM): Animals were perfusion-
fixed in 3% glutaraldehyde in phosphate buffer for 20 minutes at 100 mmHg, followed by
excision of the aorta and immersion in 3% glutaraldehyde in 0.1 M sodium cacodylate-HCl
buffer (pH 7.3) with 0.05 M sucrose (>2 hours). The specimens were then cut into small rings
with a razor blade. After rinsing, the specimens were postfixed in 1.5% osmium tetroxide in
0.1 M cacodylate buffer (pH 7.3) with 0.7% potassium ferrocyanate for 2 h at 4°C,
dehydrated in ethanol (70, 95, 100%), stained with 2% uranyl acetate in ethanol, and

embedded in Spurr low viscosity epoxy resin. For TEM, sections of uniform thickness were
cut with diamond knives on a Leica Ultracut ultramicrotome, picked up on grids, stained with
alkaline lead citrate, and examined in a Philips CM120Twin electron microscope at 80 kV.
For toluidine blue stainings, 1µm thick sections were cut, stained and observed in a Nikon
Optiphot-2 microscope with Plan 20x/0.50 or 40x/0.85 objectives and a Nikon DS-2Mv
digital camera connected to a computer with Nikon NIS Elements imaging software.
Antibodies and reagents: mAb RB40.34 against mouse P-selectin was kindly provided
by Dietmar Vestweber, University of Münster, Germany. The hybridoma for mAb GAME46
against mouse CD18 was kindly provided by Ed Roos, the Netherlands Cancer Institute,
Amsterdam, the Netherlands and antibody was prepared from cell supernatants on a protein L
column. Rhodamine 6G and FITC dextran came from Sigma. FITC-labeled microspheres
were obtained from Molecular Probes and beads coated with protein G from Polysciences Inc.
Statistical analysis: The data represent mean±SEM of measurements obtained in
the indicated number of experiments. Statistical comparison between independent groups was
performed using the Student t-test or the Mann-Whitney rank sum test, whereas comparison
before and after antibody-blockage of CAMs were performed using paired t-test or Wilcoxon
signed rank test for paired samples. Multiple comparisons were performed by one way
ANOVA or Kruskal-Wallis one way analysis of variance on ranks adjusted for multiple
comparisons using Dunn´s test. Data on leukocyte rolling before and after antibody treatment
(figure 3) were analyzed using a two-level conditional hierarchical linear model with fixed
group effect where the fixed effect parameters correspond to the means of the groups and the
random effects are the levels of the random effect nested within the levels of the fixed effect
according to:
Responseijk = Group i + Animal j(i) + Segment k(ij)

The model has two variance component parameters to be estimated, the animal-to-animal
variability and the segment-to-segment variability within an animal. Data on leukocyte
adhesion was calculated using both analysis on pooled data on adhesion per mm2 endothelial
area as well as by a similar model as that described for leukocyte rolling. Post-hoc analyses
were performed by Dunnett’s test. Distribution of these variables was positively skewed and
before the formal analyses the variables were log-transformed. Statistical significance was set
at p<0.05.

Supplemental Figure 1
Intimal thickness (µm)
0 200 400 600
Nu
mb
er
of
inti
ma
l v
es
se
ls
0
1
2
3
4
Total plaque thickness (µm)
0 200 400 600 800
Nu
mb
er
of
ad
ven
titi
al
vessels
(per
1000µ
m)
0
5
10
15
20
25
30
Nu
mb
er
of
ad
ven
titi
al
vessels
(p
er
1000µ
m)
0
2
4
6
8
10
12
14
Inta
ct m
edia
Dam
aged
med
ia
Des
truct
ed m
edia
*
A
B
C

Supplemental figure 2A

Supplemental figure 2B

Supplemental Figure Legends
Supplemental figure 1
Analysis of the presence of lesion microvessels in 1μm sections stained with toluidine blue.
(A) Number of intimal microvessels vs lesion thickness. (B) Number of adventitial vessels per
1000 μm vessel circumference vs lesion thickness. (C) Number of adventitial vessels vs level
of medial destruction. Intact media was defined as media with no injury to the elastic
lamellae. Damaged media had incomplete damage to medial elastin fibers whereas destructed
media had areas of total destruction of elastin lamellae with an open connection between the
intima and adventitia. Data are expressed as mean±SEM, n=39 for all graphs. Statistical
analysis was performed using Kruskal-Wallis one-way analysis of variance on Ranks adjusted
for multiple comparisons by Dunn’s method. Only one lesion per mouse was analyzed. *)
represents significant difference compared to intact media (p<0.05).
Supplemental figure 2
(A and B) Photomontage of intravital microscopy images on an atherosclerotic lesion after
labelling of microvessels with an intravenous injection of FITC dextran. In B, arterioles,
capillaries and venules are depicted with red, green or blue lines, respectively. The same
lesion is shown in supplemental video 1. Bar = 400μm.

Supplemental Video Legends
Supplemental video 1
Intravital microscopy on an advanced atherosclerotic lesion in an ApoE-/- mouse. The clip
shows the microvascular network during an intravenous injection of the plasma tracer FITC
dextran. Previously injected fluorescent beads (∅1µm) are visible in microvessels.
Supplemental video 2
Venular rolling and adhesion of leukocytes in an atherosclerotic lesion in an ApoE-/- mouse.
Flow is from upper right to lower left. Rolling and adherent leukocytes were visualized by
fluorescence microscopy following intravenous injection of rhodamine 6G.