valproate prevents mk801-induced changes in brain-derived neurotrophic factor mrna in the rat brain
TRANSCRIPT

European Journal of Pharmacology 545 (2006) 142–146www.elsevier.com/locate/ejphar
Short communication
Valproate prevents MK801-induced changes in brain-derivedneurotrophic factor mRNA in the rat brain
Gye Sun Jeon a,1, Sang-Ha Park a,1, Kuem-Ju Lee a, Min-Soo Lee b,Boe-Gwun Chun a, Kyung-Ho Shin a,⁎
a Department of Pharmacology, Korea University College of Medicine, #5 Anam-dong, Seoul 136-705, South Koreab Department of Psychiatry, Korea University College of Medicine, #5 Anam-dong, Seoul 136-705, South Korea
Received 23 November 2005; received in revised form 20 June 2006; accepted 22 June 2006Available online 28 June 2006
Abstract
To investigate whether the anticonvulsant valproate influences the changes in brain-derived neurotrophic factor (BDNF) mRNA expressioninduced by MK801 in rat brain, we injected valproate prior to MK801 and observed the changes in the BDNF expression 3 h later. MK801significantly increased BDNF expression in the retrosplenial and entorhinal cortex, and these increases were prevented by valproate pretreatment.Valproate pretreatment significantly blocked the MK801-induced increase of BDNF expression in retrosplenial cortex at 3 h, 6 h, and 9 h afterMK801 injection, suggesting that valproate pretreatment did not delay the MK801-induced increase of BDNF expression. However, MK801significantly decreased BDNF expression in the granule cell layer of hippocampus, and valproate pretreatment before MK801 potentiated theMK801-induced decrease in BDNF expression in granule cell layer. These results indicate that valproate pretreatment differentially affects theMK801-induced changes in BDNF expression in a region-selective manner.© 2006 Elsevier B.V. All rights reserved.
Keywords: BDNF (brain-derived neurotrophic factor); MK801; Valproate; Retrosplenial cortex
1. Introduction
MK801 (dizocilpine) is a noncompetitive antagonist of the N-methyl-D-aspartate (NMDA) receptor and has been shown toreduce neuronal injury associated with excitotoxicity (Choi,1988). However, the administration of MK801 producesneurotoxic effects in selective neurons in the retrosplenial andcingulate cortices of the rat brain (Fix et al., 1993). In addition toits neurotoxic effects, MK801 also induces the expression of avariety of genes, such as heat shock protein genes, the brain-derived neurotrophic factor (BDNF) gene, immediate early genes,and transcription factor genes in rat brains (Castren et al., 1993;Gass et al., 1993; Sharp et al., 1991). For example,MK801 causesa marked increase in BDNF mRNA levels in the rat retrosplenialcortex (Castren et al., 1993; Hughes et al., 1993).
⁎ Corresponding author. Tel.: +82 2 920 6195; fax: +82 2 927 0824.E-mail address: [email protected] (K.-H. Shin).
1 GSJ and SHP equally contributed to this article.
0014-2999/$ - see front matter © 2006 Elsevier B.V. All rights reserved.doi:10.1016/j.ejphar.2006.06.046
It was reported that several antiepileptic drugs preventMK801-induced neurotoxicity in the retrosplenial cortex, which may becorrelated with their antiepileptic efficacy (Farber et al., 2002).Among the antiepileptic drugs, valproate potentiates gamma-aminobutyric acid (GABA)-mediated neuronal responses, and alsoregulates excitatory amino acid neurotransmission (Loscher, 1993)by attenuating NMDA receptor-mediated excitation (Zeise et al.,1991). Although valproate is known to prevent MK801-inducedneurotoxicity in the retrosplenial cortex, it is not clear whethervalproate influences the MK801-induced changes in BDNFmRNA levels. Therefore, we injected valproate prior to theinjection of MK801 in rats and observed the changes in BDNFmRNA levels in rat brain areas, including the retrosplenial cortex.
2. Materials and methods
2.1. Animals and drug treatments
Adult male Sprague–Dawley rats (280–300 g; n=5 or 6 pergroup; Orient Co., Gyunggi-do, Korea) were used. The rats were
143G.S. Jeon et al. / European Journal of Pharmacology 545 (2006) 142–146
brought into the laboratory 1week before the experiment andwerehoused, three per cage, under a 12-h light–dark cycle (lights on at06:00 h). Food and water were available ad libitum. The animalswere administered either valproate (400 or 800 mg/kg, i.p.,dissolved in sterile water) or vehicle (sterile water), followed byeither MK801 (2 mg/kg, i.p., dissolved in 0.9% sterile saline) or0.9% sterile saline 20 min later. Rats were deeply anesthetizedwith sodium pentobarbital (100 mg/kg i.p.) 3 h after the saline orMK801 injection. MK801 and valproate were purchased fromSigma (St. Louis, MO, USA).
2.2. Experimental groups
The animals were divided into five experimental groups.Experimental group 1, which served as a control, wasadministered vehicle prior to a saline injection (Veh+Sal,n=5). The animals in group 2, which served as an additionalcontrol, were administered valproate (800 mg/kg) prior to asaline injection (VPA+Sal, n=5). The rats in experimentalgroup 3 received vehicle prior to an MK801 injection (Veh+MK801, n=6), and the rats in experimental group 4 or 5received 400 or 800mg valproate per kg, respectively, prior to anMK801 injection (VPA400+MK801 and VPA800+MK801,respectively; n=5 for each group). To measure BDNF mRNAlevels in the entorhinal cortex, we conducted an additionalexperiment, in which animals (n=3 per group) received vehicle
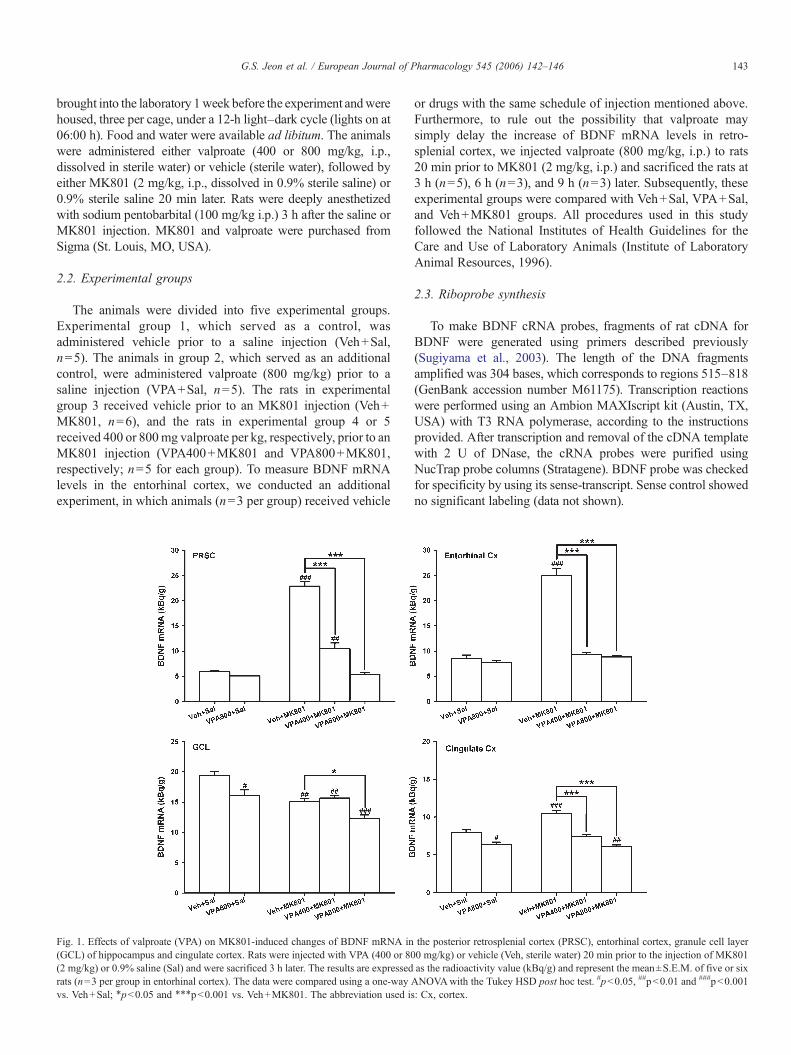
Fig. 1. Effects of valproate (VPA) on MK801-induced changes of BDNF mRNA in(GCL) of hippocampus and cingulate cortex. Rats were injected with VPA (400 or 80(2 mg/kg) or 0.9% saline (Sal) and were sacrificed 3 h later. The results are expressedrats (n=3 per group in entorhinal cortex). The data were compared using a one-wayvs. Veh+Sal; ⁎p<0.05 and ⁎⁎⁎p<0.001 vs. Veh+MK801. The abbreviation used is
or drugs with the same schedule of injection mentioned above.Furthermore, to rule out the possibility that valproate maysimply delay the increase of BDNF mRNA levels in retro-splenial cortex, we injected valproate (800 mg/kg, i.p.) to rats20 min prior to MK801 (2 mg/kg, i.p.) and sacrificed the rats at3 h (n=5), 6 h (n=3), and 9 h (n=3) later. Subsequently, theseexperimental groups were compared with Veh+Sal, VPA+Sal,and Veh+MK801 groups. All procedures used in this studyfollowed the National Institutes of Health Guidelines for theCare and Use of Laboratory Animals (Institute of LaboratoryAnimal Resources, 1996).
2.3. Riboprobe synthesis
To make BDNF cRNA probes, fragments of rat cDNA forBDNF were generated using primers described previously(Sugiyama et al., 2003). The length of the DNA fragmentsamplified was 304 bases, which corresponds to regions 515–818(GenBank accession number M61175). Transcription reactionswere performed using an Ambion MAXIscript kit (Austin, TX,USA) with T3 RNA polymerase, according to the instructionsprovided. After transcription and removal of the cDNA templatewith 2 U of DNase, the cRNA probes were purified usingNucTrap probe columns (Stratagene). BDNF probe was checkedfor specificity by using its sense-transcript. Sense control showedno significant labeling (data not shown).
the posterior retrosplenial cortex (PRSC), entorhinal cortex, granule cell layer0 mg/kg) or vehicle (Veh, sterile water) 20 min prior to the injection of MK801as the radioactivity value (kBq/g) and represent the mean±S.E.M. of five or sixANOVAwith the Tukey HSD post hoc test. #p<0.05, ##p<0.01 and ###p<0.001: Cx, cortex.
144 G.S. Jeon et al. / European Journal of Pharmacology 545 (2006) 142–146
2.4. In situ hybridization
After anesthesia, the rats were perfused with 4% paraformal-dehyde in 0.1 M sodium phosphate buffer (PPB), and the brainswere removed from the skull, post-fixed for 2 h in 4% PPB, andcryoprotected in 20% sucrose in PPB. Serial coronal sections(30 μm) of the whole brain were obtained using a freezingmicrotome. In addition, serial horizontal sections (30 μm) wereobtained tomeasure BDNFmRNA levels in the entorhinal cortex.The sections were permeabilized with proteinase K (1 μg/ml,37 °C, 30 min), treated with acetic anhydride in 0.1 Mtriethanolamine (pH 8.0), and then hybridized for 18 h at 55 °C.After hybridization, sections were treated with RNase A (20 μg/ml, 30 min, 37 °C), washed to a stringency of 0.1× sodium citratebuffer (1× sodium citrate buffer=0.15 M sodium chloride and15 mM sodium citrate, pH 7.0), mounted onto gelatin-coatedmicroscope slides, air-dried overnight, and then exposed toHyperfilm β-max (Amersham, Arlington Heights, IL, USA) for3 days. To correct for nonlinearity, [14C]-labeled radioactivestandards (Amersham) ranging from 1.15 to 32.7 kBq/g wereused for calibration.
2.5. Data analysis and statistics
The sections for analysis were selected through the cingulateand piriform cortex (0.60–1.44 mm from bregma), anteriorretrospenial cortex (−3.12 to −3.84 mm from bregma), posteriorretrosplenial cortex (−5.40 to −6.0 mm from bregma), hippo-campus (−3.60 to −3.80 mm from bregma) and entorhinal cortex(−5.3 to −6.6 mm in horizontal plane) according to Paxinos andWatson (1998). The level of BDNF mRNA was analyzed by
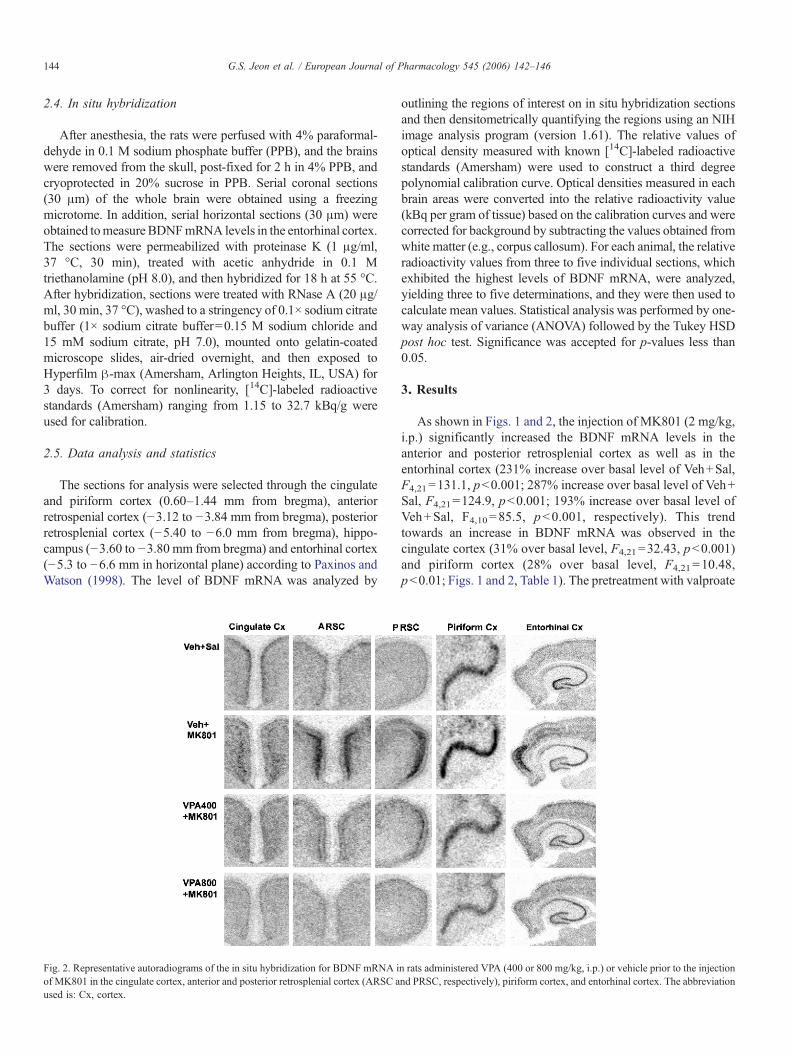
Fig. 2. Representative autoradiograms of the in situ hybridization for BDNF mRNA iof MK801 in the cingulate cortex, anterior and posterior retrosplenial cortex (ARSC aused is: Cx, cortex.
outlining the regions of interest on in situ hybridization sectionsand then densitometrically quantifying the regions using an NIHimage analysis program (version 1.61). The relative values ofoptical density measured with known [14C]-labeled radioactivestandards (Amersham) were used to construct a third degreepolynomial calibration curve. Optical densities measured in eachbrain areas were converted into the relative radioactivity value(kBq per gram of tissue) based on the calibration curves and werecorrected for background by subtracting the values obtained fromwhite matter (e.g., corpus callosum). For each animal, the relativeradioactivity values from three to five individual sections, whichexhibited the highest levels of BDNF mRNA, were analyzed,yielding three to five determinations, and they were then used tocalculate mean values. Statistical analysis was performed by one-way analysis of variance (ANOVA) followed by the Tukey HSDpost hoc test. Significance was accepted for p-values less than0.05.
3. Results
As shown in Figs. 1 and 2, the injection of MK801 (2 mg/kg,i.p.) significantly increased the BDNF mRNA levels in theanterior and posterior retrosplenial cortex as well as in theentorhinal cortex (231% increase over basal level of Veh+Sal,F4,21=131.1, p<0.001; 287% increase over basal level of Veh+Sal, F4,21=124.9, p<0.001; 193% increase over basal level ofVeh+Sal, F4,10=85.5, p<0.001, respectively). This trendtowards an increase in BDNF mRNA was observed in thecingulate cortex (31% over basal level, F4,21=32.43, p<0.001)and piriform cortex (28% over basal level, F4,21=10.48,p<0.01; Figs. 1 and 2, Table 1). The pretreatment with valproate
n rats administered VPA (400 or 800 mg/kg, i.p.) or vehicle prior to the injectionnd PRSC, respectively), piriform cortex, and entorhinal cortex. The abbreviation

Table 1Effects of valproate (VPA) on the MK801-induced changes of BDNF mRNA in the piriform cortex and subfields of hippocampus
Veh+Sal VPA800+Sal Veh+MK801 VPA400+MK801 VPA800+MK801
Piriform cortex 19.1±1.0a 18.9±0.2a 24.3±1.1c 20.1±0.8a 17.7±0.3b
CA1 area 11.4±0.4 10.5±0.5 10.0±0.3 10.9±0.4 8.4±0.5c
CA3 area 23.9±0.5 21.7±0.7 23.5±0.8 22.6±0.6 17.0±0.7b,d
The results are expressed as the kBq/g value and represented as the mean±S.E.M. from five or six rats. The data were compared using a one-way ANOVAwith theTukey HSD post hoc test. aP<0.01 and bP<0.001 compared to Veh+MK801; cP<0.01 and dP<0.001 compared to Veh+Sal.
145G.S. Jeon et al. / European Journal of Pharmacology 545 (2006) 142–146
prevented the MK801-induced increase in BDNF mRNA in theanterior and posterior retrosplenial cortex in a dose-dependentmanner, but the high dose of valproate (800 mg/kg) adminis-tration prior to the saline did not change the BDNFmRNA levelsin these areas (Figs. 1 and 2). Interestingly, pretreatment of ratswith high dose of valproate significantly blocked the MK801-induced increase of BDNF mRNA levels in retrosplenial cortexat 3 h, 6 h, and 9 h after MK801 injection (data not shown). Thisresult suggests that valproate pretreatment significantly blocked,but not delayed, the MK801-induced increase of BDNF mRNAlevels in retrosplenial cortex. In contrast, the MK801-inducedincrease of BDNF mRNA levels in the entorhinal, cingulate andpiriform cortex was completely prevented by pretreatment withthe low dose of valproate (400 mg/kg, Figs. 1 and 2, Table 1).Notably, MK801 significantly decreased the BDNF mRNAlevels in the granule cell layer of hippocampus (Fig. 1). Thepretreatment with high dose of valproate prior to MK801administration significantly potentiated the MK801-induceddecrease in BDNF expression in the granule cell layer ofhippocampus (Fig. 1) and other subfields of hippocampus (CA1and CA3 area, Table 1). Furthermore, pretreatment with the highdose of valproate prior to the saline injection significantlydecreased the basal levels of BDNF mRNA in the granule celllayer of hippocampus and cingulate cortex (Fig. 1).
4. Discussion
In the present study,MK801 significantly increased the BDNFmRNA levels in the retrosplenial and entorhinal cortex, as shownby the previous studies (Castren et al., 1993; Hughes et al., 1993).However, treatment with valproate prior to the injection ofMK801 significantly prevented the increase in BDNF mRNAlevels in these areas. Previously, Farber (2003) hypothesized thatNMDA receptors are located on GABAergic interneurons, whichinhibit excitatory projections to the retrosplenial cortex, and thatNMDA receptor antagonists disinhibit the GABAergic interneur-ons, resulting in increased release of glutamate and acetylcholinein the retrosplenial cortex. Thus, an MK801-induced increase inglutamate and acetylcholine may lead to the increase in BDNFmRNA levels in the retrosplenial cortex. We demonstrated thatvalproate provided dose-dependent inhibition of the MK801-induced increase of BDNF mRNA levels in the retrosplenialcortex, although themechanism is not clear. One possibility is thatvalproate may inhibit voltage-dependent sodium channels andthereby interfere with neurotransmitter release, which would inturn lead to a reduction in the MK801-induced release ofglutamate and acetylcholine (Falqueto et al., 2002). Alternatively,valproate may enhance inhibitory GABAergic activity, either by
increasing GABA levels and its subsequent release or byenhancing postsynaptic GABA responses (Motohashi, 1992).This possibility is supported by the fact that pentobarbital, whichpotentiates the activation of GABAergic receptors, also com-pletely blocked the effect of MK801 on the BDNF mRNA levelsin the retrosplenial cortex (Castren et al., 1993). Interestingly,valproate pretreatment was reported to prevent MK801-inducedneuronal damage in retrosplenial cortex (Farber et al., 2002).
Consistent with the previous study (Castren et al., 1993),MK801 significantly increased BDNF mRNA levels in theentorhinal cortex. Compared to the retrosplenial cortex, the lowdose of valproate completely blocked the MK801-inducedincrease of BDNF mRNA in the entorhinal cortex (Fig. 1). Asvalproate reduces glutamate release and induces a potentiation ofthe postsynaptic GABAA receptor in rat entorhinal cortex(Cunningham et al., 2003), it is likely that valproate pretreatmentblocked the MK801-induced increase of BDNF mRNA levels inthe entorhinal cortex. Similar patterns of increased BDNFmRNAlevels induced by MK801 were also observed in the cingulatecortex and piriform cortex, and these increases were prevented bypretreatment with valproate. Valproate pretreatment in the saline-treated control induced a small but significant decrease in BDNFmRNA in the cingulate cortex, although the underlyingmechanism is not clear.
MK801 significantly decreased BDNF mRNA levels in thegranule cell layer of hippocampus. This MK801-induceddecrease in BDNF mRNA in the hippocampus was similar tothe previous finding (Zafra et al., 1991). The high dose ofvalproate pretreatment prior to saline injection significantlydecreased BDNF mRNA levels in the granule cell layer ofhippocampus. Moreover, the high dose of valproate pretreatmentprior to MK801 administration significantly potentiated theMK801-induced decrease in BDNFmRNA levels in the subfieldsof the hippocampus (CA1, CA3 and granule cell layer area).Considering that valproate enhances basal extracellular GABAconcentration in the rat hippocampus (Ahmad et al., 2005), it ispossible that valproate pretreatment prior to MK801 maypotentiate the decrease in BDNF mRNA produced by MK801in hippocampus. Alternatively, it is possible that valproatepretreatment prior to MK801 may activate 5-hydroxytryptamine(5-HT)2A receptors in the hippocampus, which in turn leads todecrease in the BDNFmRNA levels. This possibility is supportedby the fact that 5-HT2A receptors are expressed on GABAergicinterneurons as well as in the granule cell layer of thehippocampus (Wright et al., 1995), and that the activation of 5-HT2A receptors reduces BDNF mRNA levels in the granule celllayer of hippocampus (Vaidya et al., 1997). Interestingly, it wasreported that valproate significantly increases serotonin release in

146 G.S. Jeon et al. / European Journal of Pharmacology 545 (2006) 142–146
the hippocampus (Ahmad et al., 2005). Clearly, further work isneeded to define the changes in serotonin release with thecombined valproate and MK801 treatment.
Taken together, the present study demonstrated that MK801significantly increased BDNF expression in the retrosplenial andentorhinal cortex and that these increases were prevented byvalproate pretreatment. However, MK801 significantly decreasedBDNF expression in the granule cell layer of hippocampus, andvalproate pretreatment before MK801 significantly potentiatedthe MK801-induced decrease in BDNF mRNA levels in thesubfields of the hippocampus (CA1, CA3 and granule cell layer ofhippocampus). These results indicate that valproate pretreatmentdifferentially affects the MK801-induced changes in BDNFexpression in a region-selective manner.
Acknowledgements
This research was supported by a grant from the BrainResearch Center of the 21st Century Frontier Research Program,funded by the Korean Ministry of Science and Technology(M103KV010007-03K2201-00720), Brain Korea 21 Project in2006 and Korea Health 21 R&D Project (KPGRN-R-04-04) bythe Ministry of Health and Welfare, Republic of Korea.
References
Ahmad, S., Fowler, L.J., Whitton, P.S., 2005. Effects of combined lamotrigineand valproate on basal and stimulated extracellular amino acids andmonoamines in the hippocampus of freely moving rats. Naunyn-Schmie-deberg's Arch. Pharmacol. 371, 1–8.
Castren, E., da Penha Berzaghi, M., Lindholm, D., Thoenen, H., 1993.Differential effects of MK-801 on brain-derived neurotrophic factor mRNAlevels in different regions of the rat brain. Exp. Neurol. 122, 244–252.
Choi, D.W., 1988. Glutamate neurotoxicity and diseases of the nervous system.Neuron 1, 623–634.
Cunningham, M.O., Woodhall, G.L., Jones, R.S., 2003. Valproate modifiesspontaneous excitation and inhibition at cortical synapses in vitro. Neurophar-macology 45, 907–917.
Falqueto, E.B., Massensini, A.R., Moraes-Santos, T., Gomez, M.V., Romano-Silva,M.A., 2002. Modulation of Na+-channels by neurotoxins produces differenteffects on [3H]ACh release with mobilization of distinct Ca2+-channels. Cell.Mol. Neurobiol. 22, 819–826.
Farber, N.B., 2003. The NMDA receptor hypofunction model of psychosis.Ann. N. Y. Acad. Sci. 1003, 119–130.
Farber, N.B., Jiang, X.P., Heinkel, C., Nemmers, B., 2002. Antiepileptic drugsand agents that inhibit voltage-gated sodium channels prevent NMDAantagonist neurotoxicity. Mol. Psychiatry 7, 726–733.
Fix, A.S., Horn, J.W., Wightman, K.A., Johnson, C.A., Long, G.G., Storts,R.W., Farber, N., Wozniak, D.F., Olney, J.W., 1993. Neuronal vacuoliza-tion and necrosis induced by the noncompetetive N-methyl-D-aspartate(NMDA) antagonist MK(+)801 (dizocilpine maleate): a light and electronmicroscopic evaluation of the rat retrosplenial cortex. Exp. Neurol. 123,204–215.
Gass, P., Herdegen, T., Bravo, R., Kiessling, M., 1993. Spatiotemporal inductionof immediate early genes in the rat brain after limbic seizures: effects ofNMDA receptor antagonist MK-801. Eur. J. Neurosci. 5, 933–943.
Hughes, P., Dragunow, M., Beilharz, E., Lawlor, P., Gluckman, P., 1993.MK801 induces immediate-early gene proteins and BDNF mRNA in ratcerebrocortical neurones. Neuroreport 4, 183–186.
Loscher, W., 1993. Effects of the antiepileptic drug valproate on metabolism andfunction of inhibitory and excitatory amino acids in the brain. Neurochem.Res. 18, 485–502.
Motohashi, N., 1992. GABA receptor alterations after chronic lithiumadministration. Comparison with carbamazepine and sodium valproate.Prog. Neuro-Psychopharmacol. Biol. Psychiatry 16, 571–579.
Paxinos, G., Watson, C., 1998. The Rat Brain in Stereotaxic Coordinates, 4th ed.Academic Press, San Diego.
Sharp, F.R., Jasper, P., Hall, J., Noble, L., Sagar, S.M., 1991. MK-801 andketamine induce heat shock protein HSP72 in injured neurons in posteriorcingulate and retrosplenial cortex. Ann. Neurol. 30, 801–809.
Sugiyama, N., Kanba, S., Arita, J., 2003. Temporal changes in the expression ofbrain-derived neurotrophic factor mRNA in the ventromedial nucleus of thehypothalamus of the developing rat brain. Brain Res. Mol. Brain Res. 115,69–77.
Vaidya, V.A., Marek, G.J., Aghajanian, G.K., Duman, R.S., 1997. 5-HT2A
receptor-mediated regulation of brain-derived neurotrophic factor mRNA inthe hippocampus and the neocortex. J. Neurosci. 17, 2785–2795.
Wright, D.E., Seroogy, K.B., Lundgren, K.H., Davis, B.M., Jennes, L., 1995.Comparative localization of serotonin1A, 1C, and 2 receptor subtypemRNAs in rat brain. J. Comp. Neurol. 351, 357–373.
Zafra, F., Castren, E., Thoenen, H., Lindholm, D., 1991. Interplay betweenglutamate and gamma-aminobutyric acid transmitter systems in the physio-logical regulation of brain-derived neurotrophic factor and nerve growth factorsynthesis in hippocampal neurons. Proc. Natl. Acad. Sci. U. S. A. 88,10037–10041.
Zeise, M.L., Kasparow, S., Zieglgansberger, W., 1991. Valproate suppresses N-methyl-D-aspartate-evoked, transient depolarizations in the rat neocortex invitro. Brain Res. 544, 345–348.