ultrastructural localization of neuropeptide y y1 receptors in the rat medial nucleus tractus...
TRANSCRIPT

Ultrastructural Localization of NeuropeptideY Y1 Receptors in the Rat Medial NucleusTractus Solitarius: Relationships WithNeuropeptide Y or Catecholamine Neurons
Michael J. Glass,* June Chan, and Virginia M. PickelDepartment of Neurology and Neuroscience, Division of Neurobiology, Weill Medical College of CornellUniversity, New York, New York
Neuropeptide Y (NPY) Y1 receptor (Y1-R) agonists influ-ence cardiovascular regulation. These actions may involveNPY- and catecholamine-containing neurons in the medialnucleus of the solitary tract (mNTS), at the level of the areapostrema. The cellular sites through which Y1-R agonistsmay interact with NPY and catecholamines in the mNTS,however, are not known. To determine potential sitesof action for Y1-R agonists, and their relationship toNPY or catecholamines in the mNTS, we used electronmicroscopic immunocytochemistry for the detection ofsequence-specific antipeptide antisera against Y1-R aloneor in combination with antisera against NPY or thecatecholamine-synthesizing enzyme tyrosine hydroxylase(TH). Analyses were conducted in the rat mNTS, at the levelof the area postrema. Y1-R was found mainly in smallunmyelinated axons and axon terminals but also in somesomata and dendrites as well as a small number of glia.Within axon terminals, labeling for Y1-R was often presenton dense core vesicles and small synaptic vesicles as wellas extrasynaptic areas of the plasmalemma. Some Y1-R-labeled terminals also contained NPY or TH, suggestingthat agonists of Y1-R may influence the release of NPY orcatecholamines in the mNTS. In addition, Y1-R was foundin dendrites that received asymmetric excitatory-type syn-apses from unlabeled axon terminals. Some of these den-drites contained NPY or TH, which indicates that Y1-R maybe targeted for functional activation within NPY- orcatecholamine-expressing neurons in the mNTS. These re-sults demonstrate that Y1-R is a presynaptic receptor inNPY- or catecholamine-containing axon terminals withinthe mNTS as well as a postsynaptic receptor on NPY- orcatecholamine-containing neurons that are contacted byaxon terminals that likely contain excitatory amino acidtransmitters. Agonists of Y1-R in the mNTS may thus affectcardiovascular regulation by modulating NPY, catechol-amine, and excitatory amino acid transmission.© 2002 Wiley-Liss, Inc.
Key words: autonomic nervous system; blood pressure;excitatory amino acids; tyrosine hydroxylase; heart rate
The medial nucleus of the solitary tract (mNTS), atthe level of the area postrema, is an important anatomical
substrate for cardiovascular control (Chan et al., 2000;Deuchars et al., 2000; Gozal et al., 2000). This NTSsubregion contains the first central synapses of barorecep-tive afferents (Aicher et al., 2000b), whose cell bodiesreside in the nodose ganglia (Aicher et al., 1999) and thatmay contain the neuroactive peptide transmitter neu-ropeptide Y (NPY; Zhang et al., 1996). Other brainstemregions, such as the ventrolateral medulla, are also in-volved in cardiovascular control (Aicher et al., 2000b),contain NPY-expressing neurons (Migita et al., 2001), andinnervate the mNTS (Palkovits et al., 1995). Thus, fibersderived from the peripheral and central nervous systemsmay influence autonomic control by releasing NPY in themNTS. In addition to being present in afferent fibers,NPY is expressed within numerous somata (Chronwall etal., 1985) in the mNTS. Some of these neurons are con-tacted by �-aminobutyric acid (GABA)-ergic axon termi-nals (Pickel et al., 1989) and express the immediate earlygene c-Fos in response to changes in blood pressure(McLean et al., 1999). In summary, these findings suggestthat NPY-containing neurons in the mNTS participate inblood pressure regulation.
Complementing this anatomical evidence, it has alsobeen shown that exogenous administration of NPY intothe mNTS results in hypotension and bradycardia (Yang etal., 1997). Some of the actions of NPY on autonomicfunctions may be mediated by the neuropeptide Y Y1receptor (Y1-R; Yang et al., 1993). Cell bodies within themNTS express Y1-R mRNA (Parker and Herzog, 1999),protein (Migita et al., 2001), and ligand binding sites
Contract grant sponsor: NIH; Contract grant number: HL18974; Contractgrant sponsor: NIMH; Contract grant number: MH48776; Contract grantnumber: MH00078; Contract grant sponsor: NIDA; Contract grant num-ber: DA04600; Contract grant number: T32DA07274.
*Correspondence to: Dr. Michael J. Glass, Department of Neurology andNeuroscience, Weill Medical College of Cornell University, 410 E. 69thSt., KB410, New York, NY 10021.E-mail: [email protected]
Received 29 August 2001; Revised 27 November 2001; Accepted 6December 2001
Published online 15 February 2002
Journal of Neuroscience Research 67:753–765 (2002)DOI 10.1002/jnr.10185
© 2002 Wiley-Liss, Inc.

(Yang et al., 1996b). In addition to being present onsomata, NPY binding sites have been found on fibers thatinnervate the mNTS (McLean et al., 1996). This evidencesuggests that Y1-R agonists may modulate autonomicfunctions through pre- and postsynaptic Y1-R sites in-volving neurons that may also express NPY.
Many Y1-R (Yang et al., 1996a)- or NPY (Harf-strand et al., 1987)-containing neurons in the mNTS alsoexpress catecholamines, suggesting that both cat-echolamines and NPY may be similarly synthesized incommon neurons that also contain Y1-R. In addition,NPY, Y1-R agonists, and catecholamines may have sim-ilar actions on autonomic functions within the mNTS. Forexample, intra-mNTS microinjection of NPY or the cat-echolamine norepinephrine produces hypotension andbradycardia (Zandberg and De Jong, 1977; Yang et al.,1997). The hypotensive actions of NPY injections into themNTS are inhibited by coinjection of yohimbine, anantagonist of the alpha-2A-adrenergic receptor, one of themajor catecholamine receptors in the mNTS (Morris etal., 1997). Thus, NPY and catecholamines in the NTSmay jointly modulate cardiovascular functions, possibly byactions involving Y1-R (Yang et al., 1996b).
The cellular sites through which Y1-R agonistsmodulate the activity of mNTS neurons, particularly thosethat contain NPY or catecholamines, are unclear. Vagalafferents innervate the mNTS and may express NPY orY1-R (Zhang et al., 1996; Migita et al., 2001), suggestingthat Y1-R may modulate the release of NPY from theseafferent terminals. However, there is no evidence thatY1-R and NPY are present on the same axon terminals inthe NTS. In addition, catecholaminergic fibers derivedfrom other brainstem nuclei innervate the mNTS (Thorand Helke, 1988) and are believed to influence autonomicfunctions (Aicher et al., 2000b). Many of these cat-echolaminergic cell populations also express Y1-R (Yanget al., 1996a), suggesting that agonists of this receptor mayinfluence catecholamine release from axon terminals in themNTS. There is no evidence, however, that catechol-amine terminals in the mNTS express Y1-R.
To determine potential sites of action for Y1-Ragonists, particularly in relation to NPY and cat-echolaminergic axon terminals, we used single- anddouble-labeling electron microscopic immunocytochem-istry for the detection of sequence-specific antipeptideantisera against Y1-R alone or in combination with NPYor tyrosine hydroxylase (TH), the rate-limiting enzyme incatecholamine biosynthesis (Zigmond et al., 1989). Anal-yses were performed in sections taken from the rat inter-mediate mNTS, corresponding to the level of the areapostrema.
MATERIALS AND METHODS
Animals and Tissue Preparation
Experimental protocols involving animals and their carewere approved by the Institutional Animal Care and Use Com-mittee at Weill Medical College of Cornell University andconformed to NIH guidelines. Two adult male Sprague-Dawley
rats (Taconic Farms, Germantown, NY) were anesthetized withpentobarbital (150 mg/kg, i.p.), and their brains were fixed byaortic arch perfusion sequentially with the following: 1) 15 ml ofnormal saline (0.9%), containing 1,000 units/ml of heparin; 2)50 ml of 3.75% acrolein in 2% paraformaldehyde in 0.1 Mphosphate buffer (PB; pH 7.4); and 3) 200 ml of 2% parafor-maldehyde in PB, all delivered at a flow rate of 100 ml/min. Thebrains were removed and postfixed for 30 min in 2% parafor-maldehyde in PB. Coronal sections 40 �m thick were cut witha vibrating microtome from the intermediate medial NTS at thelevel of the area postrema according to the atlas of Paxinos andWatson (1986). This area is known to contain a large populationof noradrenergic neurons, in addition to smaller groups ofepinephrine- and dopamine-containing cells. Tissue sectionswere then placed in 1.0% sodium borohydride in PB for 30 minand washed in PB. Sections were then immersed in a cryopro-tectant solution (25% sucrose and 2.5% glycerol in 0.05M PB)for 15 min. To enhance tissue permeability, sections werefreeze-thawed in liquid freon and liquid nitrogen. Sections werenext rinsed in 0.1 M Tris-buffered saline (TBS; pH 7.6) andthen incubated for 30 min in 0.1% bovine serum albumin (BSA)in TBS to minimize nonspecific labeling.
Antisera Used for Immunocytochemistry
A rabbit polyclonal antibody raised against amino acids282–299 in the second part of the third extracellular loop (E3/2)of Y1-R was used in this study. The specificity of this antiserahas been previously demonstrated using several methods, in-cluding immunobloting, enzyme-linked immunosorbent assay(ELISA), and preadsorption with undiluted antiserum (Pickel etal., 1998).
For dual labeling with the rabbit Y1-R antiserum, weused a goat antiserum that was raised against the carboxy-terminal hexapeptide of NPY 13–36 (provided by the laboratoryof the late Dr. T.L. Donohue). This antiserum has been shownto recognize human NPY 13–36 and pancreatic PYY but nothuman NPY 1–24 or desamino NPY (Pickel et al., 1998). Acommercial mouse monoclonal TH antibody (Incstar, Stillwa-ter, MN) was used in dual-labeling experiments with Y1-R.This antibody has been well characterized and extensively usedfor immunocytochemistry (Glass et al., 2001).
Immunocytochemical Procedures
Single Labeling. The Y1-R antiserum was processedfor immunogold or immunoperoxidase labeling (Chan et al.,1990). For immunogold labeling, tissue sections were incubatedin primary Y1-R (gold: 1:2,000; peroxidase: 1:4,000) antiserumin TBS for 48 hr. Next, sections were rinsed in TBS and thenin 0.01 M PBS (pH 7.4) and blocked for 10 min in 0.5% BSAand 0.1% gelatin in PBS to reduce nonspecific binding of goldparticles. Sections were then incubated for 2 hr in anti-rabbitIgG conjugated with 1 nm gold particles (1:50; AuroProbeOne;Amersham, Arlington Heights, IL). The sections were rinsed inPBS and incubated for 10 min in 2% glutaraldehyde. The boundgold particles were enlarged by a 5–10 min silver intensificationusing an IntenSE-M kit (Amersham). The tissue was then post-fixed in 2% osmium tetroxide in PB for 1 hr and dehydrated ina series of alcohols, through propylene oxide, and embedded in
754 Glass et al.

EM BED 812 (EMS, Fort Washington, PA) between two sheetsof Aclar plastic.
For immunoperoxidase labeling, sections were washed inTBS and then incubated in anti-rabbit IgG conjugated to biotin,rinsed in TBS, and incubated for 30 min in avidin-biotin-peroxidase complex (1:100; Vectastain Elite Kit; Vector Labo-ratories, Burlingame, CA) in TBS. The bound peroxidase wasvisualized by reaction for 6 min in a 0.2% solution of 3,3�-diaminobenzidine (DAB) and 0.003% hydrogen peroxide inTBS.
Double Labeling. Preparation of tissue sections fordual labeling was identical to that for single labeling, except thatthe sections were incubated in a solution containing both anti-Y1-R and anti-NPY (gold: 1:2,000; peroxidase: 1:4,000) oranti-TH (gold: 1:2,500; peroxidase: 1:20,000) antisera. After aTBS wash, sections were prepared first for peroxidase identifi-cation of antisera. Sections were incubated in secondary anti-body conjugated to biotin, rinsed in TBS, and then incubatedfor 30 min in avidin-biotin-peroxidase complex (1:100; Vec-tastain Elite Kit; Vector Laboratories) in TBS. The bound per-oxidase was visualized by reaction for 6 min in a 0.2% solutionof 3,3�-DAB and 0.003% hydrogen peroxide in TBS. For theimmunogold detection of antisera, sections were prepared asdescribed above, except that secondary antibody conjugated togold particles was used. The tissue was then postfixed, dehy-drated, and embedded in Aclar plastic as described above. Goldand peroxidase markers were reversed in alternate sections.Control experiments included processing the tissue for duallabeling by omission of one of the primary antisera in each set tocontrol for possible cross-reactivity, which upon subsequentanalysis was not encountered.
Tissue Preparation for Microscopy
Ultrathin sections from the surface of the tissue were cutwith a diamond knife using an ultramicrotome (Ultratome;Nova LKB, Bromma, Sweden). These sections were collected
on grids and then counterstained with Reynold’s lead citrate anduranyl acetate and examined with a Philips 201 electron micro-scope.
Ultrastructural Analysis
In the dual-labeling experiment, three vibratome sectionsfrom each of two animals were sectioned on an ultratome. Onethin section from each of six blocks corresponding to eachlabeling condition showing optimal labeling and morphologywas chosen for analysis. Because of the different sensitivities ofthe immunoperoxidase and immunogold methods, samplingwas performed only on thin sections taken from the plastic–tissue interface. Whole thin sections were scanned at low power(7,000�) until labeled neuropil was detected. All potentiallylabeled profiles were then photographed at higher magnification(20,000�) for subsequent identification according to the criterialisted below. Only the area that included at least one labeledprofile was photographed and counted as sampled. In total,570 �m2 of tissue was sampled in the single-labeling experi-ment, 310 �m2 of tissue was sampled from the Y1-R and NPYexperiment, and 298 �m2 of tissue was sampled from the Y1-Rand TH experiment.
The classification of labeled and unlabeled neurons wasbased on descriptions by Peters and coworkers (1991). Immu-noreactive processes were identified as axon terminals if theymeasured at least 0.2 �m in diameter and contained numeroussynaptic vesicles. Most dendrites were identified by the presenceof postsynaptic densities as well as ribosomes and both rough andsmooth endoplasmic reticulum. However, profiles were alsoconsidered dendritic whenever postsynaptic densities were ob-served, independent of endoplasmic reticulum. Somata weredistinguished by the presence of a nucleus. Astrocytic processeswere identified by their irregular shape and filamentous mem-branes that often surrounded both axons and dendrites. Synapseswere defined as either symmetric or asymmetric according tothe presence of either thin or thick postsynaptic specializations,respectively. Appositions were distinguished by closely spacedplasma membranes that lacked recognizable specializations,without the presence of interposing astrocytic processes.
To be considered labeled, profiles had to contain at leasttwo gold particles. Single gold particles, however, were some-times found on the plasma membrane in small profiles. Becauseof the strong correlation between profile size and the number ofgold particles it contains, the presence of a single plasmalemmal
TABLE I. Distribution of Y1-R-Labeled Profiles in Single-Labeling Experiments*
Axons/axon terminals (%) Dendrites (%) Glia (%) Somata (%)
53 25 20 2
*Based on 106 Y1-R-labeled profiles.
TABLE II. Percentage of Single- and Double-Labeled Profiles in Double-Labeling Experiments
Y1-Ra (%) NPYb (%) Y1 � NPYc (%) Y1-Rd (%) THe (%) Y1 � THf (%)
Axons/axon terminals 29 58 78 25 60 35Dendrites 41 33 11 63 40 53Glia 18 7 0 13 0 0Somata 12 2 11 0 0 12aBased on 17 single-labeled Y1-R profiles.bBased on 45 single-labeled NPY profiles.cBased on nine dual-labeled profiles.dBased on eight single-labeled Y1-R profiles.eBased on 25 single-labeled TH profiles.fBased on 17 dual-labeled profiles.
Y1 Receptors in mNTS 755

Figure 1.

gold particle marking Y1-R is likely to represent specific label-ing in small structures, which has been shown for other surfaceantigens (Garzon et al., 1999). In addition, background goldlabeling was established to be low as evidenced by infrequentobservation of particles over myelin or other structures notexpected to show Y1-R immunoreactivity. Thus, small pro-cesses with a single gold particle on the plasma membrane wereanalyzed in serial sections to confirm the specificity of singleplasmalemmal gold particles in these processes, which nonethe-less represented only a small number of all labeled profiles (n � 3).
Electron micrographs used for illustrations were scannedusing an AGFA Arcus II scanner (Agfa-Gevaert NV Belgium)connected to a power Macintosh 8500/150 computer (AppleComputer Inc., Cupertino, CA). These images were then im-ported into Adobe Photoshop (Adobe Systems Inc., MountainView, CA). Figures were prepared using QuarkXPress (QuarkInc., Denver, CO) with Adobe Illustrator (Adobe Systems Inc.).
RESULTSIn mNTS sections prepared for single labeling, Y1-R
was found mainly in axonal, dendritic, and glial profiles(Table I). The percentage of Y1-R-labeled profiles dif-fered in single- and double-labeling experiments in whichthe profiles were selected exclusively in regions containingthe secondary markers NPY or TH. Dual labeling forY1-R and NPY showed that many axons and axon ter-minals, as well as some somata and dendrites, also con-tained both markers (Table II). Appositions betweenY1-R- and NPY-labeled profiles were also detected.Fifty-six percent (5/9) of all appositions involved NPY-labeled terminals and Y1-R-labeled dendrites, whereas33% (3/9) involved NPY-labeled axons terminals andY1-R-labeled glia. In sampled tissue sections prepared fordual labeling of Y1-R and TH antisera, both markers weremost often found in dendrites, axons, and axon terminals(Table II). Of all appositions between TH- and Y1-R-labeled profiles, 80% (4/5) were between TH-labeledaxon terminals and Y1-R-labeled dendrites.
Y1-Labeled Axon Terminals: Association WithNPY or TH
Many axon terminals and small unmyelinated axonscontained labeling for Y1-R. Immunoreactivity for Y1-R
was most prominent in axon terminals that sometimesformed asymmetric synapses, with marked perforations ofthe postsynaptic density, onto unlabeled dendrites (Fig.1A,B). Labeling for Y1-R was occasionally detected as asingle gold particle (Fig. 2E) or as an aggregate of immu-noperoxidase reaction product (Fig. 1A,B) on the plasma-lemma and surrounding cytosol. Within axon terminals,labeling for Y1-R was also often associated with largedense core vesicles and small synaptic vesicles. These densecore vesicles, which were often scattered within the ter-minal or near extrasynaptic portions of the plasmalemma(Fig. 1C), contained peroxidase mainly on the vesicularmembrane, although the central lumens were also fre-quently obscured by the dense accumulation of immuno-reactivity (Fig. 1C). Peroxidase reaction product for Y1-Rwas also seen to rim the outer membrane of small synapticvesicles (Fig. 2A), some of which were located near seg-ments of the plasma membrane that also appeared tocontain receptor immunoreactivity. Labeling for Y1-Rwas frequently found in small unmyelinated axons thatwere apposed to unlabeled neurons (Fig. 1D).
In sections prepared for dual labeling, Y1-R-immunoreactive axons and axon terminals were found tocontain NPY or TH, although many of the Y1-R-labeledaxon terminals were without detectable labeling for theseantigens. The distribution of Y1-R in dual-labeled axonterminals did not differ from those that were singly la-beled. Immunoreactivity for NPY was most often seen interminals containing numerous small synaptic and largedense core vesicles (Fig. 2A). Immunolabeling for NPYwas detected in the cytosol and in association with smallclear or large dense core vesicles near the plasma mem-brane. Many NPY-containing terminals—some of whichformed asymmetric synapses—apposed unlabeled den-drites or were surrounded by unlabeled glia (data notshown). In Y1-R-labeled axons or axon terminals thatcontained TH, receptor labeling was found near synapticvesicles or the plasmalemma, whereas the catecholamine-synthesizing enzyme was diffusely distributed within thecytosol (Fig. 2E,F).
Presence of Y1-R in Somata and Dendrites:Relationship to NPY or TH
Within somata, Y1-R immunoreactivity was foundmainly in the cytoplasma, as well as near Golgi complexesand vesicular organelles, but was rarely observed on theplasma membrane. Immunogold particles for Y1-R wereoften widely dispersed throughout the soma and werefound in the cytosol (Fig. 3A) and on the outer mitochon-drial membrane or nearby tubulovesicles (Fig. 3A) as wellas rough endoplasmic reticulum (Fig. 3C). Similarly, im-munoperoxidase reaction product for Y1-R was typicallydetected near the outer mitochondrial membrane or tu-bulovesicular organelles (Fig. 3B).
In contrast to somata, immunoperoxidase or immu-nogold labeling for Y1-R in dendrites was often foundnear the plasma membrane (Fig. 4A–C) as well as in thecytosol (Fig. 4B,C) and near intracellular organelles (datanot shown). Immunoperoxidase for Y1-R was often
Š
Fig. 1. In sections prepared for single labeling, Y1-R immunoreactivityis found in the cytosol, on the plasma membrane, and near synapticvesicles in axon terminals or unmyelinated axons in the mNTS. A,B:Two ultrathin serial sections through an axon terminal (Y1-t) withimmunoperoxidase for Y1-R. Dense aggregates of peroxidase reactionproduct for Y1-R are shown in the cytosol and near the plasmamembrane in proximity to mitochondria (m). This axon terminalmakes an asymmetric synapse (arrows), with an apparent perforation,onto an unlabeled dendrite (ud). C: Immunoperoxidase for Y1 isassociated with membranes of several large dense core vesicles (dcv) ina Y1-R-labeled axon terminal (Y1-t). Many unlabeled small synapticvesicles (ssv) are widely dispersed throughout the cytoplasm. D: Anunmyelinated axon (Y1-a) contains peroxidase for Y1-R. ua, Unmy-elinated axon; ut, unlabeled axon terminal. Scale bars � 0.5 �m.
Y1 Receptors in mNTS 757

Figure 2.
758 Glass et al.

found on the postsynaptic density of asymmetric synapsesmade by unlabeled axon terminals (Fig. 4A,C), some ofwhich displayed pronounced perforations of the postsyn-aptic density (Fig. 4A). Most of the afferent terminals weredensely filled with small synaptic vesicles that were nearthe active zone (Fig. 4C,D). Large dense core vesicleswere detected in these axon terminals, but these were onlyoccasionally seen near the synapse (Fig. 4A).
In sections prepared for dual labeling, some Y1-R-containing somata and dendrites were shown to containNPY (Fig. 3B) or TH (Figs. 3C, 4B,D). The subcellulardistribution of Y1-R did not differ in single- or double-labeled profiles. In addition immunoperoxidase or immu-nogold Y1-R-labeled dendrites were occasionally con-tacted by NPY (Fig. 5A)- or TH (Fig. 5B)-containingaxon terminals. In some instances, dendrites contained adiscrete puncta of peroxidase for Y1-R on the plasma mem-brane apposed to an NPY-labeled axon terminal (Fig. 5A).NPY-labeled terminals contacting Y1-R-immunoreactivedendrites often contained dense core vesicles, many ofwhich showed a single gold particle for the peptide on theouter vesicular membrane. Y1-R-containing dendritesalso received asymmetric synapses from TH-labeled axonterminals (Fig. 5B).
Y1-R in GliaOccasionally, Y1-R-containing glial processes were
observed. Y1-R labeling was usually detected within thecytosol in large fibrous astrocytes or near the plasma mem-brane and within the cytosol in thin elongated processes(Fig. 4A). Many Y1-R-containing glia partially or com-pletely surrounded labeled or unlabeled profiles, appearingto isolate these profiles from nearby neuronal processes,most typically unmyelinated axons, or axon terminals. Lessfrequently, Y1-R-labeled glia apposed Y1-R-containing
dendritic profiles. No Y1-R-expressing glial processes haddetectable labeling for TH or NPY.
DISCUSSIONThis study provides qualitative evidence that Y1-R
is present in small unmyelinated axons and axon terminalsas well as some somata, dendrites, and glia in the interme-diate mNTS. In addition, some Y1-R-labeled axons andaxon terminals in the mNTS were found to contain NPYor TH. Together, these results suggest that Y1-R is in-volved in the presynaptic release of NPY or cat-echolamines within the mNTS. In addition, Y1-R wasfound in a select population of dendrites, many of whichreceived asymmetric excitatory-type synapses from unla-beled axon terminals. Numerous Y1-R-labeled dendritesalso were shown to contain NPY or TH or apposedterminals that contained one of these antigens. Theseresults suggest that Y1-R agonists may affect the activity ofNPYergic or catecholaminergic neurons by modulatingexcitatory postsynaptic signaling. Activation of Y1-R mayhave numerous actions involving the release of NPYand/or catecholamines from axon terminals, or thepostsynaptic responses to glutamate on apposed dendrites,which are likely to include cardiovascular regulation,which is believed to be affected by NPY or cat-echolamines.
Y1-R Labeling Is Found on the Plasmalemma andNear Synaptic Vesicles in Axon Terminals
Numerous axon terminals were shown to containY1-R in mNTS sections prepared for gold or peroxidaselabeling. Y1-R-labeled axon terminals were usually ofmedium size and contained dense core vesicles and were,thus, similar to NPY-containing axon terminals in themNTS as identified in this and previous studies (Pickel etal., 1989). Within axon terminals, Y1-R labeling wasoften seen on the extrasynaptic plasmalemma, indicatingthat these receptors are positioned for functional activa-tion. Y1-R is coupled to inhibitory G-proteins (Cabreleand Beck-Sickinger, 2000); therefore, agonist activationwould be expected to result in inhibition of transmitterrelease. This suggestion is consistent with physiologicalanalyses in other brain regions, such as the hippocampus,showing that Y1-R agonists influence the release of trans-mitters such as glutamate (Biello et al., 1997; Patrylo et al.,1999).
Within Y1-R-containing axon terminals, immuno-labeling was also often seen on large dense core vesiclesand sometimes small synaptic vesicles as well. Severalreports of immunolabeling for peptide receptors on largedense core vesicles exist and include the � (Cheng et al.,1995)- and � (Drake et al., 1996)-opioid receptors. Thepotential functional significance of the affiliation of Y1-Rwith large dense core synaptic vesicles is unclear but mayindicate that the plasmalemmal availability of the receptoris determined by recycling of these vesicles followingphysiological activation. A similar mechanism has beenproposed for the �-opioid receptor in vasopressin-containing cells of the hypothalamus (Shuster et al., 1999).
Š
Fig. 2. Y1-R is present in axon terminals containing NPY or TH inmNTS sections prepared for dual labeling. A: An axon terminal con-tains labeling for both Y1-R and NPY (Y1&NPY-t). Immunoperox-idase for Y1-R (arrowhead) is found in the cytosol near the plasmamembrane and synaptic vesicles. Gold particles for NPY (arrows) arewidely dispersed in the cytosol. B: A small unmyelinated axon(Y1&NPY-a) shows peroxidase immunoreactivity for Y1-R on a dis-crete portion of the outer membrane of a mitochondrion (m). Goldparticles for NPY are associated with vesicles near the plasmalemma. Asingle NPY immunogold-labeled axon (NPY-a) and unlabeled (ua)axons are found in the adjacent neuropil. C,D: In two ultrathin serialsections, exclusively Y1-R- or TH-labeled profiles are present in theneuropil. A dendrite (Y1-d) shows gold particles on the plasma mem-brane. An axon terminal (TH-t) with diffuse immunoperoxidase forTH is present nearby. E,F: Serial sections display labeling for Y1-R andTH in an axon terminal. Single immunogold particles for Y1-R (ar-rowheads) can be seen on the plasmalemma (E) or in the cytosol (F) ofa small axon terminal (Y1&TH-t) that also shows diffuse labeling forTH. In direct apposition to this terminal is a dendrite (Y1-d) thatcontains two gold particles for Y1-R in E but not in F, which wascollected at a greater depth from the surface of the tissue. ssv, Smallsynaptic vesicles; ud, unlabeled dendrite; ut, unlabeled axon terminal.Scale bars � 0.5 �m.
Y1 Receptors in mNTS 759

Fig. 3. Y1-R is found in somata of the mNTS that sometimes expressNPY or TH. A: In a section processed for single labeling, immunogoldparticles (arrowheads) for Y1-R are found in the cytosol and near theouter membrane of a mitochondria (m) of a soma (Y1-s). B: In asection processed for dual labeling, immunoperoxidase reaction prod-uct for Y1 (arrowheads) is found near a mitochondria (m) and sur-rounding the outer membrane of a vesicular organelle (vb) in a soma(Y1&NPY-s) that also contains numerous gold particles (arrows) for
NPY. A cluster of gold particles for NPY is seen in a multivesicularbody (mvb). C: In a section processed for dual labeling, immunogoldfor Y1-R (arrowheads) is found in a soma (Y1&TH-s) that also containsdiffuse immunoperoxidase labeling for TH. Single gold particles forY1-R are scattered in the cytoplasma or on rough endoplasmic retic-ulum (rER). nuc, Nucleus; ua, unlabeled axons; ud, unlabeled dendrite;us, unlabeled soma; asterisks, unlabeled glia. Scale bars � 0.5 �m.

Y1-R Labeling Is Present in Axon TerminalsContaining NPY or TH
In sections processed for dual labeling, many Y1-R-containing axon terminals also expressed NPY or TH.These results indicate that Y1-R agonists can modulate thelocal release of NPY or catecholamines in the mNTS.Occasionally, labeling for Y1-R and NPY was localized tothe same dense core vesicles, which are known to storeand release both neuropeptides and monoamine transmit-ters (DeCamilli and Jahn, 1990). The dual-labeled axonterminals resembled those that have previously beenshown to contain the vesicular monoamine transporter 2(VMAT2) in the mNTS (Nirenberg et al., 1995). VMAT2is responsible for the uptake of monoamines into vesiclesin the central and peripheral nervous systems (Schuldiner,1994) and the release of monoamines from large densecore vesicles (Waites et al., 2001). Axon terminals thatexpress VMAT2 in the mNTS are thought to containmainly norepinephrine and epinephrine (Pickel, 2000) andare, thus, likely derived from catecholaminergic brainstemcell groups that innervate the mNTS, including the rostralventrolateral medulla and locus coeruleus (Thor andHelke, 1988). The present results thus suggest that Y1-Ris an important presynaptic receptor in axon terminals thatstore and release catecholamines and NPY within themNTS.
Y1-R Shows Qualitative Differences in SubcellularDistribution in Somata and Dendrites
Y1-R was found in somata within the mNTS at thelevel of the area postrema. When detected with either goldor peroxidase markers, Y1-R labeling was found mostoften in the cytosol, in vesicular organelles, and on smoothor rough endoplasmic reticula and Golgi complexes. Thelatter structures are sites of protein synthesis, trafficking, ordegradation (Peters et al., 1991); thus, these results areconsistent with molecular evidence (Parker and Herzog,1999) that Y1-R is expressed within mNTS neurons.Y1-R immunoreactivity, however, was rarely present onthe plasmalemma. This may reflect limited targeting ofY1-R to the somatic plasmalemma or a high degree ofreceptor endocytosis (Parker et al., 2001) from the somaticplasmalemma because of tonic receptor activation by en-dogenous NPY (Shih et al., 1992). The intracellular dis-tribution of Y1-R immunoreactivity in the present studyis similar to that seen in spiny and aspiny neurons in thenucleus accumbens (Pickel et al., 1998).
Y1-R labeling was often detected on the plasmamembrane of dendrites, in distinction from somata, al-though labeling was also found in the cytoplasma and nearcytoplasmic organelles. When Y1-R labeling was foundon the plasmalemma of dendrites, it was often present atextrasynaptic sites and occasionally near the postsynapticdensity formed by axon terminals, relatively few of whichcontained NPY. Although the differences in cytoplasmicand plasmalemmal distribution in somata and dendritesrequire further quantitative verification, the qualitative
observations are consistent with active trafficking of Y1-Rto postsynaptic sites in dendrites in mNTS.
Many Y1-R-labeled dendrites received asymmetricsynapses that exhibited perforated postsynaptic densities.Perforated synapses have been hypothesized to reflect anearly stage in synaptic remodeling (Nieto-Sampedro et al.,1982), including formation of new synapses or redistribu-tion of receptors (Toni et al., 1999). The significance ofperforated synapses in the mNTS is unclear, but it mayrepresent basal remodeling of tonically active synapses thatmediate cardiovascular or respiratory function. The majorsources of excitatory fibers in the mNTS include afferentsof the vagus nerve (Aicher et al., 1999, 2000a) as well asinsular (Torrealba and Muller, 1996) and infralimbic (Tor-realba and Muller, 1999) cortices. Many of the axonterminals that contacted Y1-R-labeled dendrite were ofmedium size, were ovoid, and made single synaptic con-tacts and are characteristic of NPY-containing terminals(Pickel et al., 1989) but not vagal afferents (Sumal et al.,1983). The insular and infralimbic cortices also providemajor sources of excitatory afferents into the mNTS andcontain neurons that express NPY. Unlike the NPY-containing axon terminals detected in the present study,however, axon terminals derived from these cortical areasare devoid of dense core vesicles (Torrealba and Muller,1996, 1999). It seems unlikely that many of the axonterminals making excitatory-type synapses onto Y1-Rdendrites are derived from vagal or cortical sources ofglutamate in the mNTS. We postulate that at least some ofthese terminals may be derived from collaterals of gluta-matergic or catecholamine-containing neurons within themNTS or from other catecholamine-containing cellgroups such as the locus coeruleus or ventrolateral me-dulla. The latter speculation is based on the finding thatsome catecholaminergic terminals form asymmetric synapseswithin the mNTS (Pickel et al., 1996).
Dendrites in mNTS Express Y1-R and ContainNPY or TH
In sections prepared for dual labeling, Y1-R wasoften found on the plasmalemma of dendrites that expressNPY or TH. The presence of functionally positionedY1-R in dendrites that express NPY or TH indicates thatagonist-induced activation of this receptor may affect thephysiological activity of catecholamine- and/or NPY-containing neurons. These actions may affect not onlycardiovascular functions but also the autonomic effects ofcertain drugs of abuse. Accumulating evidence suggeststhat NPY (Thiele et al., 1998) and the catecholaminenorepinephrine (Mao and Abdel-Rahman, 1996) have animportant role in ethanol consumption. Some of theseactions may involve neurons in the mNTS, at the level ofthe area postrema, which contain NPY and norepineph-rine, the major catecholamine expressed by cells in this brainregion (Kalia et al., 1985). For example, norepinephrine-and NPY-containing neurons in the mNTS show cFOSexpression in response to ethanol ingestion (Thiele et al.,2000). In addition, the hypothalamus (Sawchenko et al.,1985) and amygdala (Riche et al., 1990; Zardetto-Smith
Y1 Receptors in mNTS 761
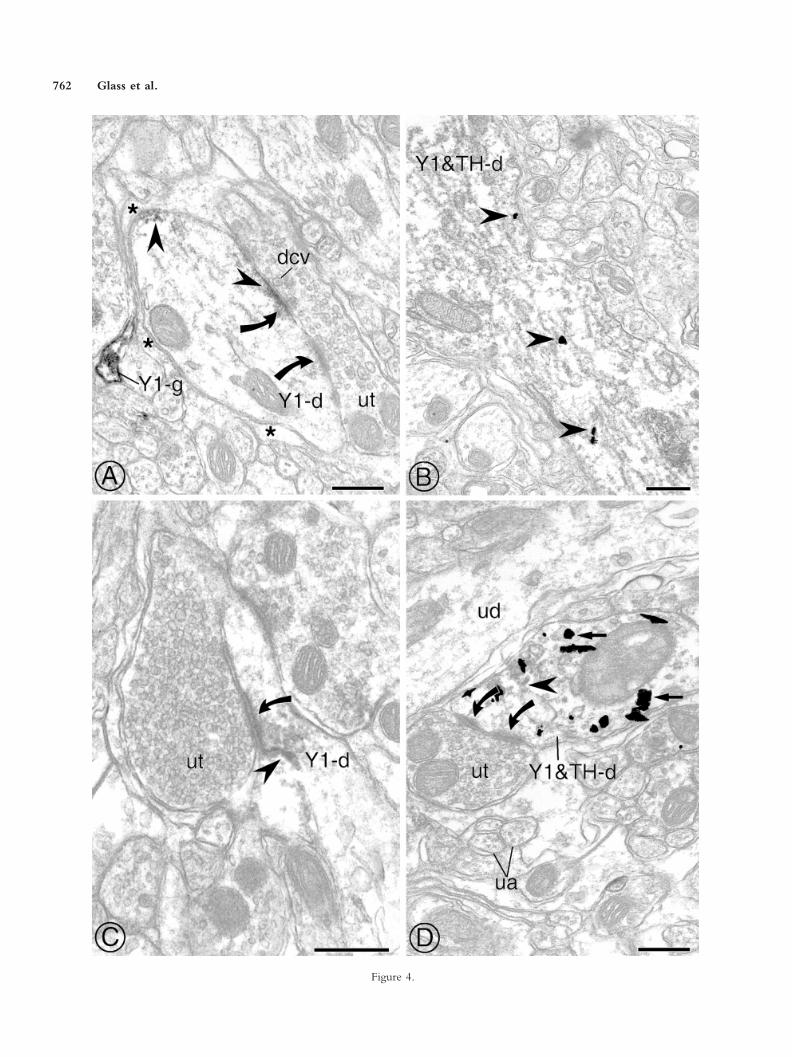
Figure 4.
762 Glass et al.

and Gray, 1990), which are major targets of mNTSneurons, have been implicated in mediating the actionsof norepinephrine (Hodge et al., 1996) and NPY(Ehlers et al., 1998; Hwang et al., 1999) on ethanolingestion. Thus, activation of Y1-R-containing neu-rons in the mNTS may affect release of NPY andcatecholamines in the amygdala or hypothalamus,which may have significant effects on autonomic pro-cesses affected by alcohol consumption.
In addition to the presence of Y1-R and NPY orcatecholamines in the same neurons, some dendrites ex-clusively labeled for Y1-R were contacted by NPY- orTH-containing axon terminals. NPY labeling in these andall other NPY-containing terminals was associated withdense core vesicles and suggests that these vesicles aremajor sites for storage and release of NPY in the mNTS(Pickel et al., 1989). NPY-labeled terminals were oftenseen to form asymmetric synapses onto Y1-R or unlabeleddendrites. Because asymmetric synapses are associated withexcitatory synaptic transmission, dendritic Y1-R, activatedby NPY released from apposed axon terminals, is posi-tioned for involvement in the modulation of postsynapticexcitation.
Fig. 5. Y1-R immunoreactivity is found in dendrites contacted by NPY-or TH-containing axon terminals in mNTS sections processed for duallabeling. A: A dense puncta of peroxidase reaction product for Y1-R(arrowhead) is seen on the plasma membrane of a small dendrite (Y1-d)apposed to an axon terminal (NPY-t), with gold particles for NPY (arrows)
that are widely distributed throughout the axon terminal. B: Immunogoldparticles for Y1-R are present in the cytosol of a dendrite (Y1-d) receivingan asymetric synapse (arrow) from a TH-labeled axon terminal (TH-t).This axon terminal contains numerous small synaptic vesicles (ssv) and adense core vesicle (dcv). ua, Unlabeled axon. Scale bars � 0.5 �m.
Š
Fig. 4. Y1-R is present in the cytoplasma and near the plasmalemma ofsingle-labeled dendrites or glia and in TH-containing dendrites in themNTS. A: In a section processed for single labeling, immunoperoxidasereaction product for Y1-R (arrowhead) is found on a portion of thepostsynaptic density of a dendrite (Y1-d). This dendrite receives a perfo-rated asymmetric synapse (arrows) from an unlabeled axon terminal (ut)that contains dense core vesicles (dcv), which can be seen near the activezone. A segment of unlabeled postsynaptic density (arrow curved down-ward) can also be seen adjacent to the perforation. A glial process (Y1-g)with peroxidase labeling for Y1-R is also present in the neuropil. B: In asection processed for dual labeling, immunogold particles for Y1-R (ar-rowheads) are seen near the plasmalemma as well as dispersed in the cytosolof a longitudinally sectioned dendrite that also contains diffuse peroxidaselabeling for TH (Y1&TH-d). C: In a section processed for single labeling,immunoperoxidase for Y1-R (arrowhead) is found in a dendrite (Y1-d)that receives an asymmetric synapse (arrow) from an unlabeled axonterminal (ut). Peroxidase reaction product for Y1-R is seen near thepostsynaptic density and in the cytosol. D: In a section processed for duallabeling, Y1-R and TH are present in a coronally sectioned dendrite(Y1&TH-d). A discrete puncta of peroxidase for Y1-R (arrowhead) ispresent in the cytoplasm and near the plasmalemma. Gold particles for THcan be seen overlapping the peroxidase reaction product as well as areas ofthe cytoplasm devoid of Y1-R labeling. This dual-labeled dendrite receivesa perforated asymmetric synapse (arrows) from an unlabeled axon terminal.ua, Unlabeled axon; ud, unlabeled dendrite; asterisks, unlabeled glial pro-cess. Scale bars � 0.5 �m.
Y1 Receptors in mNTS 763

CONCLUSIONSThe present results show that Y1-R labeling is
present on some NPY- or catecholamine-containing axonterminals and dendrites within the mNTS at the level ofthe area postrema. Thus, Y1-R is strategically positionedfor involvement in a complex set of actions, includingpresynaptic release of NPY or catecholamines as well asthe postsynaptic modulation of the effects of synapticallyreleased glutamate onto neurons that contain NPY and/orcatecholamines. These actions may influence cardiovascu-lar regulation and the autonomic effects of consumption ofdrugs of abuse, such as alcohol, which may be dependenton the activity of neurons that express NPY and cat-echolamines.
REFERENCESAgnati LF, Zoli M, Stromberg I, Fuxe K. 1995. Intercellular communica-
tion in the brain: wiring versus volume transmission. Neuroscience 69:711–726.
Aicher SA, Sharma S, Pickel VM. 1999. N-methyl-D-aspartate receptorsare present in vagal afferents and their dendritic targets in the nucleustractus solitarius. Neuroscience 91:119–132.
Aicher SA, Goldberg A, Sharma S, Pickel VM. 2000a. �-opioid receptorsare present in vagal afferents and their dendritic targets in the medialnucleus tractus solitarius. J Comp Neurol 422:181–190.
Aicher SA, Milner TA, Pickel VM, Reis DJ. 2000b. Anatomical substratesfor baroreflex sympathoinhibition in the rat. Brain Res Bull 51:107–110.
Biello SM, Golombek DA, Harrington ME. 1997. Neuropeptide Y andglutamate block each other’s phase shifts in the suprachiasmatic nucleus invitro. Neuroscience 77:1049–1057.
Cabrele C, Beck-Sickinger AG. 2000. Molecular characterization of theligand-receptor interaction of the neuropeptide Y family. J Peptide Sci6:97–122.
Chan J, Aoki C, Pickel VM. 1990. Optimization of differentialimmunogold-silver and peroxidase labeling with maintenance of ultra-structure in brain sections before plastic embedding. J Neurosci Methods33:113–127.
Chan RKW, Peto CA, Sawchenko PE. 2000. Fine structure and plasticityof barosensitive neurons in the nucleus of solitary tract. J Comp Neurol422:338–351.
Cheng PY, Svingos AL, Wang H, Clarke CL, Jenab S, Beczkowska IW,Inturrisi CE, Pickel VM. 1995. Ultrastructural immunolabeling showsprominent presynaptic vesicular localization of �-opioid receptor withinboth enkephalin- and nonenkephalin-containing axon terminals in thesuperficial layers of the rat cervical spinal cord. J Neurosci 15:5976–5988.
Chronwall BM, DiMaggio DA, Massari VJ, Pickel VM, Ruggiero DA,O’Donohue TL. 1985. The anatomy of neuropeptide Y containingneurons in the rat brain. Neuroscience 15:159–181.
DeCamilli P, Jahn R. 1990. Pathways to regulated exocytosis in neurons.Annu Rev Physiol 52:625–645.
Deuchars J, Li YW, Kasparov S, Paton JF. 2000. Morphological andelectrophysiological properties of neurons in the dorsal vagal complex ofthe rat activated by arterial baroreceptors. J Comp Neurol 417:233–249.
Ehlers CL, Li TK, Lumeng L, Hwang BH, Somes C, Jimenez P, MatheAA. 1998. Neuropeptide Y levels in ethanol-naive alcohol-preferring andnonpreferring rats and in Wistar rats after ethanol exposure. Alcohol ClinExp Res 22:1778–1782.
Garzon M, Vaughan RA, Uhl GR, Kuhar MJ, Pickel VM. 1999. Cholin-ergic axon terminals in the ventral tegmental area target a subpopulationof neurons expressing low levels of the dopamine transporter. J CompNeurol 410:197–210.
Glass MJ, Huang J, Aicher SA, Milner TA, Pickel VM. 2001. Subcellularlocalization of alpha-2A-adrenergic receptors in the rat medial nucleus
tractus solitarius: regional targeting and relationship with catecholamin-ergic neurons. J Comp Neurol 433:193–207.
Gozal D, Gozal E, Simakajornboon N. 2000. Signaling pathways of theacute hypoxic ventilatory response in the nucleus tractus solitarius. RespirPhysiol 121:209–221.
Harfstrand A, Fuxe K, Terenius L, Kalia M. 1987. NeuropeptideY-immunoreactive perikarya and nerve terminals in the rat medullaoblongata: relationship to cytoarchitecture and catecholaminergic cellgroups. J Comp Neurol 260:20–35.
Hodge CW, Slawecki CJ, Aiken AS. 1996. Norepinephrine and serotoninreceptors in the paraventricular nucleus indirectly modulate ethanol con-sumption. Alcohol Clin Exp Res 20:1669–1674.
Hwang BH, Zhang JK, Ehlers CL, Lumeng L, Li TK. 1999. Innatedifferences of neuropeptide Y (NPY) in hypothalamic nuclei and centralnucleus of the amygdala between selectively bred rats with high and lowalcohol preference. Alcohol Clin Exp Res 23:1023–1030.
Kalia M, Fuxe K, Goldstein M. 1985. Rat medulla oblongata III. Adren-ergic (C1 and C2) neurons, nerve fibers, and presumptive terminalprocesses. J Comp Neurol 233:333–349.
Mao L, Abdel-Rahman AA. 1996. Synergistic behavioral interaction be-tween ethanol and clonidine in rats: role of alpha-2 adrenoceptors. J Phar-macol Exp Ther 279:443–449.
McLean KJ, Jarrott B, Lawrence AJ. 1996. Neuropeptide Y gene expressionand receptor autoradiography in hypertensive and normotensive rat brain.Mol Brain Res 35:249–259.
McLean KJ, Jarrott B, Lawrence AJ. 1999. Hypotension activates neu-ropeptide Y-containing neurons in the rat medulla oblongata. Neuro-science 92:1377–1387.
Migita K, Loewy AD, Ramabhadran TV, Krause JE, Waters SM. 2001.Immunohistochemical localization of the neuropeptide Y Y1 receptor inrat central nervous system. Brain Res 889:23–37.
Morris MJ, Hastings JA, Pavia JM. 1997. Central interactions betweennoradrenaline and neuropeptide Y in the rat: implications for bloodpressure control. Clin Exp Hypertens 19:619–630.
Nicholson C, Chen KC, Hrabetova S, Tao L. 2000. Diffusion of moleculesin brain extracellular space: theory and experiment. Progr Brain Res125:129–154.
Nieto-Sampedro M, Hoff SF, Cotman CW. 1982. Perforated postsynapticdensities: probable intermediates in synapse turnover. Proc Natl Acad SciUSA 79:5718–5722.
Nirenberg MJ, Liu YJ, Peter D, Edwards RH, Pickel VM. 1995. Thevesicular monoamine transporter 2 is present in small synaptic vesicles andpreferentially localizes to large dense core vesicles in rat solitary tractnuclei. Proc Natl Acad Sci USA 92:8773–8777.
Palkovits M, Mezey E, Fodor M, Ganten D, Bahner U, Geiger H, HeidlandA. 1995. Neurotransmitters and neuropeptides in the baroreceptor reflexarc: connections between the nucleus of the solitary tract and the ven-trolateral medulla oblongata in the rat. Clin Exp Hypertens 17:101–113.
Parker RMC, Herzog H. 1999. Regional distribution of Y-receptor sub-type mRNAs in rat brain. Eur J Neurosci 11:1431–1448.
Parker SL, Kane JK, Parker MS, Berglund MM, Lundell IA, Li MD. 2001.Cloned neuropeptide Y (NPY) Y1 and pancreatic polypeptide Y4 recep-tors expressed in Chinese hamster ovary cells show considerable agonist-driven internalization, in contrast to the NPY Y2 receptor. Eur J Biochem268:877–886.
Patrylo PR, van den Pol AN, Spencer DD, Williamson A. 1999. NPYinhibits glutamatergic excitation in the epileptic human dentate gyrus.J Neurophysiol 82:478–483.
Paxinos G, Watson C. 1986. The rat brain in stereotaxic coordinates. NewYork: Academic Press.
Peters A, Palay SL, Webster H. 1991. The fine structure of the nervoussystem. New York: Oxford University Press.
Pickel VM. 2000. Extrasynaptic distribution of monoamine transporters andreceptors. Progr Brain Res 125:267–276.
764 Glass et al.

Pickel VM, Chan J, Massari VJ. 1989. Neuropeptide Y-like immunoreac-tivity in neurons of the solitary tract nuclei: vesicular localization andsynaptic input from GABAergic terminals. Brain Res 476:265–278.
Pickel VM, Nirenberg MJ, Milner TA. 1996. Ultrastructural view ofcentral catecholaminergic transmission: immunocytochemical localizationof synthesizing enzymes, transporters, and receptors. J Neurocytol 25:843–856.
Pickel VM, Beck-Sickinger AG, Chan J, Wieland HA. 1998. Y1 receptorsin the nucleus accumbens: ultrastructural localization and association withneuropeptide Y. J Neurosci Res 52:54–68.
Riche D, De Pommery J, Menetrey D. 1990. Neuropeptides and cat-echolamines in efferent projections of the nuclei of the solitary tract in therat. J Comp Neurol 293:399–424.
Sawchenko PE, Swanson LW, Grzanna R, Howe PR, Bloom SR, PolakJM. 1985. Colocalization of neuropeptide Y immunoreactivity in brain-stem catecholaminergic neurons that project to the paraventricular nu-cleus of the hypothalamus. J Comp Neurol 241:138–153.
Schuldiner S. 1994. A molecular glimpse of vesicular monoamine trans-porters. J Neurochem 62:2067–2078.
Shih CD, Chan JY, Chan SH. 1992. Tonic suppression of baroreceptorreflex response by endogenous neuropeptide Y at the nucleus tractussolitarius of the rat. Neurosci Lett 148:169–172.
Shuster SJ, Riedl M, Li X, Vulchanova L, Elde R. 1999. Stimulus-dependent translocation of opioid receptors to the plasma membrane.J Neurosci 19:2658–2664.
Sumal KK, Blessing WW, Joh TH, Reis DJ, Pickel VM. 1983. Synapticinteraction of vagal afferents and catecholaminergic neurons in the ratnucleus tractus solitarius. Brain Res 277:31–40.
Thiele TE, Cubero I, van Dijk G, Mediavilla C, Bernstein IL. 2000.Ethanol-induced c-Fos expression in catecholamine- and neuropeptideY-producing neurons in rat brainstem. Alcohol Clin Exp Res 24:802–809.
Thiele TE, Marsh DJ, Marie L, Bernstein IL, Palmiter RD. 1998. Ethanolconsumption and resistance are inversely related to neuropeptide Y levels.Nature 396:366–369.
Thor KB, Helke CJ. 1988. Catecholamine-synthesizing neuronal projec-tions to the nucleus tractus solitarii of the rat. J Comp Neurol 268:264–280.
Toni N, Buchs PA, Nikonenko I, Bron CR, Muller D. 1999. LTP
promotes formation of multiple spine synapses between a single axonterminal and a dendrite. Science 402:421–425.
Torrealba F, Muller C. 1996. Glutamate immunoreactivity of insular cortexafferents to the nucleus tractus solitarius in the rat: a quantitative electronmicroscopic study. Neuroscience 71:77–87.
Torrealba F, Muller C. 1999. Ultrastructure of glutamate and GABAimmunoreactive axon terminals of the rat nucleus tractus solitarius, witha note on infralimbic cortex afferents. Brain Res 820:20–30.
Waites CL, Mehta A, Tan PK, Thomas G, Edwards RH, Krantz DE. 2001.An acidic motif retains vesicular monoamine transpoter 2 on large densecore vesicles. J Cell Biol 152:1159–1168.
Yang SN, Narvaez JA, Bjelke B, Agnati LF, Fuxe K. 1993. Microinjectionsof subpicromolar amounts of NPY(13–36) into the nucleus tractus soli-tarius of the rat counteract the vasodepressor responses of NPY(1–36) andof a NPY Y1 receptor agonist. Brain Res 621:126–132.
Yang SN, Bunnemann B, Cintra A, Fuxe K. 1996a. Localization ofneuropeptide Y Y1 receptor-like immunoreactivity in catecholaminergicneurons of the rat medulla oblongata. Neuroscience 73:519–530.
Yang SN, Finnman UB, Fuxe K. 1996b. The nonpeptide neuropeptide YY1 receptor antagonist BIBP3226 blocks the [Leu31,Pro34]neuropeptideY-induced modulation of alpha 2-adrenoceptors in the nucleus tractussolitarii of the rat. Neuroreport 7:2701–2705.
Yang SN, Fior DR, Hansson AC, Cintra A, Castellano M, Ganten U,Ganten D, Agnati LF, Fuxe K. 1997. Increased potency of neuropeptideY to antagonize alpha2-adrenoceptor function in the nucleus tractussolitarii of the spontaneously hypertensive rat. Neuroscience 78:803–813.
Zandberg P, De Jong W. 1977. Localization of catecholaminergic receptorsites in the nucleus trarctus solitarii involved in the regulation of arterialblood pressure. Progr Brain Res 47:117–122.
Zardetto-Smith AM, Gray TS. 1990. Organization of peptidergic andcatecholaminergic efferents from the nucleus of the solitary tract to the ratamygdala. Brain Res Bull 25:875–887.
Zhang X, Ji RR, Arvidsson J, Lundberg JM, Bartfai T, Bedecs K, HokfeltT. 1996. Expression of peptides, nitric oxide synthase, and NPY receptorin trigeminal and nodose ganglia after nerve lesions. Exp Brain Res111:393–404.
Zigmond RE, Schwarzschild MA, Rittenhouse AR. 1989. Acute regula-tion of tyrosine hydroxylase by nerve activity and by neurotransmitters viaphosphorylation. Annu Rev Neurosci 12:415–461.
Y1 Receptors in mNTS 765