trasporto di membrana - genopomii.unina.it 2.pdf · relazione tra il potenziale chimico µ e il...
TRANSCRIPT

TRASPORTO DI MEMBRANA
movimento di molecole e ioni tra i diversi scomparti presenti nei sistemi biologici è noto come TRASPORTO, esso può essere
considerato fra singole membrane cellulari, fra una pianta e il suo ambiente circostante o fra una
parte e l’altra della pianta

Il movimento di molecole per diffusione è definito TRASPORTO PASSIVO
Il movimento di sostanze contro un gradiente
di concentrazione, o chimico, è definito TRASPORTO ATTIVO, non è spontaneo e necessita che venga compiuto del lavoro sul
sistema tramite l’applicazione di energia cellulare.
Un modo per ottenere questo risultato è di accoppiare il trasporto con l’idrolisi di ATP

membrana cellulare
Permette o impedisce il movimento di una sostanza (permeabilità di membrana)
garantisce un ambiente fisico-chimico
altamente stabile condizioni indispensabile al regolare svolgimento di attività metaboliche,
riproduttive e dello sviluppo
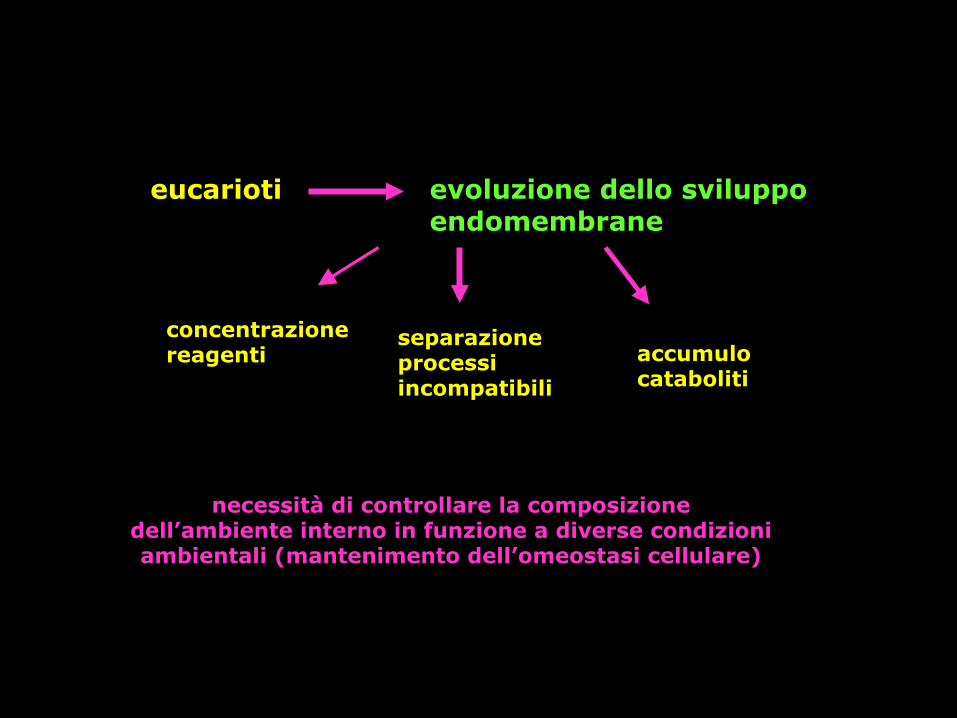
eucarioti evoluzione dello sviluppo endomembrane
concentrazione reagenti
separazione processi incompatibili
accumulo cataboliti
necessità di controllare la composizione dell’ambiente interno in funzione a diverse condizioni ambientali (mantenimento dell’omeostasi cellulare)

PIANTE
il trasporto di membrana è alla base di numerosi processi biologici essenziali
La genesi del turgore cellulare
L’acquisizione di nutrienti
La secrezione di prodotti di rifiuto del metabolismo
Distribuzione di metaboliti
La compartimentalizzazione dei metaboliti
La trasduzione dell’energia
La trasduzione del segnale

generazione del turgore. La presenza della parete cellulare permette alle cellule di generare turgore (pressione positiva)
accumulo K+ nel citoplasma e nel vacuolo
In base al principio dell’elettroneutralità delle soluzioni i cationi devono essere controbilanciati da una corrispondente concentrazioni di anioni; gli anioni principali che si accumulano nel vacuolo sono il Cl- ( in funzione alla sua disponibilità ) o il malato
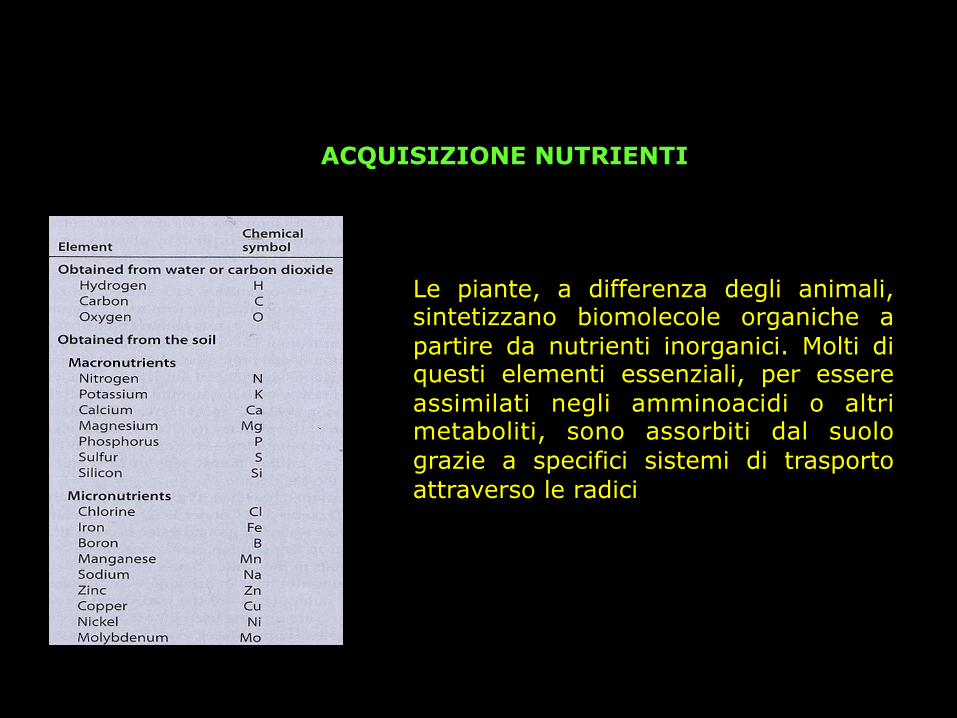
ACQUISIZIONE NUTRIENTI
Le piante, a differenza degli animali, sintetizzano biomolecole organiche a partire da nutrienti inorganici. Molti di questi elementi essenziali, per essere assimilati negli amminoacidi o altri metaboliti, sono assorbiti dal suolo grazie a specifici sistemi di trasporto attraverso le radici

ESCREZIONE PRODOTTI DI SCARTO
Il metabolismo genera inevitabilmente molecole di scarto inutilizzabili che devono
essere rimosse dal citosol.
L’H+ è il principale prodotto generato dal metabolismo
Per espellere i protoni dal citosol sulla
membrana plasmatica e su quella vacuolare delle cellule vegetali si sono evolute le
cosiddette pompe protoniche

DISTRIBUZIONE METABOLITI
Ripartizione prodotti della fotosintesi dai siti di sintesi a quelli di utilizzo e di accumulo

COMPARTIMENTALIZZAZIONE DEI METABOLITI
Alcuni processi biochimici sono incompatibili tra loro
→ necessità di evitare cicli futili
(ex. sintesi amido e glicolisi)
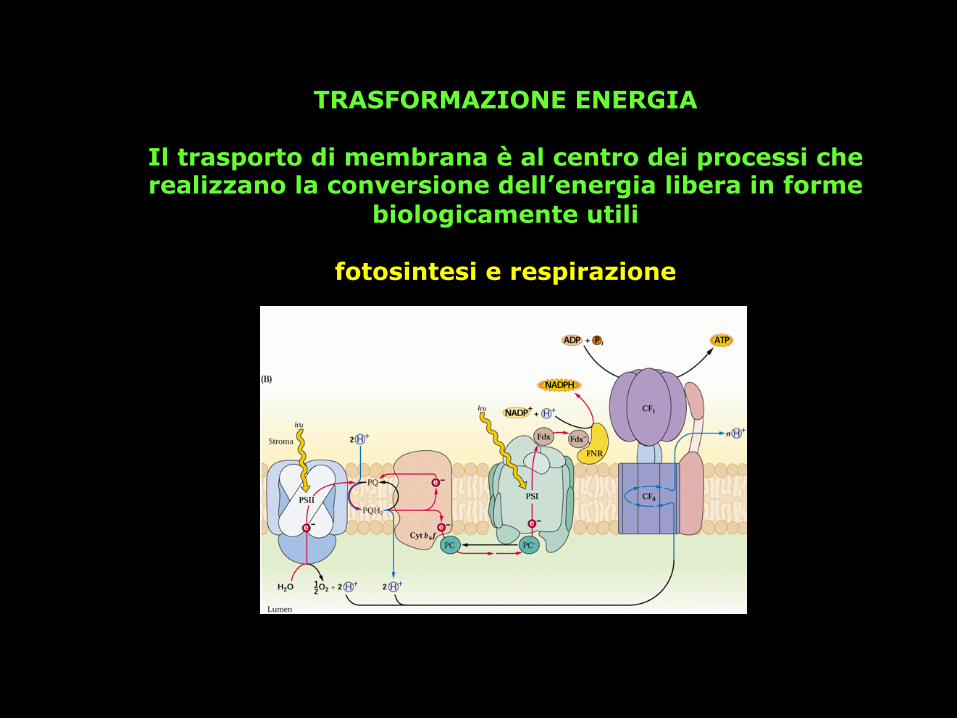
TRASFORMAZIONE ENERGIA
Il trasporto di membrana è al centro dei processi che realizzano la conversione dell’energia libera in forme
biologicamente utili
fotosintesi e respirazione

TRASDUZIONE DEL SEGNALE
Molti segnali biotici o abiotici implicati nello sviluppo e nella crescita inducono nelle piante
risposte cellulari innescate da aumenti transienti della concentrazione citosolica del Ca2
2+ libero
stimolo ambientale → ≠ [Ca22+] → risposta
fisiologica

DIFFUSIONE spostamento delle molecole secondo un gradiente di concentrazione dovuto ad
agitazione termica casuale

Il verso del movimento netto di un soluto attraverso una membrana è stabilito dal verso della forza motrice che agisce sul sistema di trasporto. Le forze motrici possono essere quantificate in termini di relazioni di energia libera connessa ai potenziali chimici del soluto

µj = µ*j + RTln Cj + zjFE + VjP
Il potenziale chimico di un soluto non carico, j, presente in un mezzo può essere definito dall’equazione:

µj = µ*j + RTln Cj + zjFE + VjP
potenziale chimico di j in condizioni
standard

µj = µ*j + RTln Cj + zjFE + VjP
componente della concentrazione

µj = µ*j + RTln Cj + zjFE + VjP
componente della carica elettrostatica

µj = µ*j + RTln Cj + zjFE + VjP
componente della pressione
idrostatica

µj = µ*j + RTln Cj + zjFE + VjP
costante universale dei
gas

µj = µ*j + RTln Cj + zjFE + VjP
temperatura assoluta

µj = µ*j + RTln Cj + zjFE + VjP
concentrazione di j
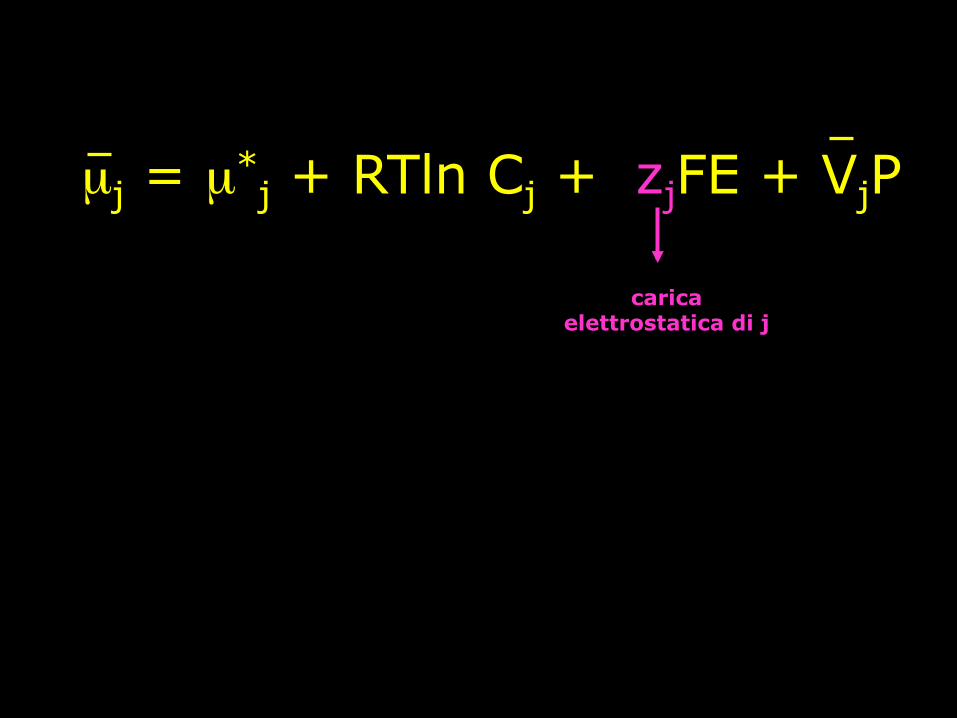
µj = µ*j + RTln Cj + zjFE + VjP
carica elettrostatica di j

µj = µ*j + RTln Cj + zjFE + VjP
costante di Faraday

µj = µ*j + RTln Cj + zjFE + VjP
potenziale elettrico generale
della soluzione
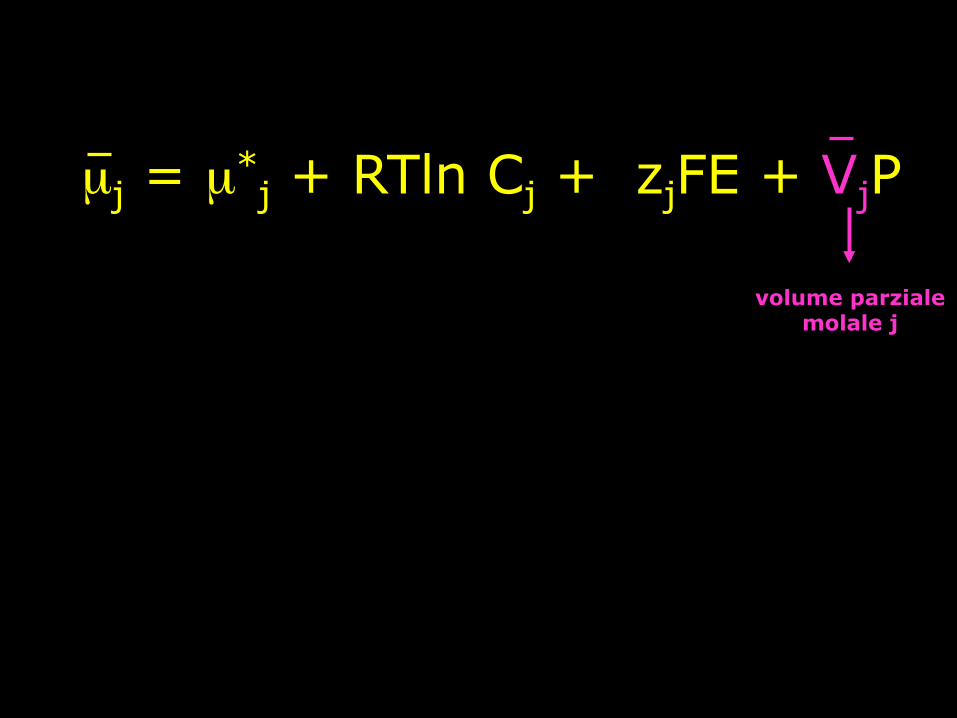
µj = µ*j + RTln Cj + zjFE + VjP
volume parziale molale j

µj = µ*j + RTln Cj + zjFE + VjP
pressione

µj = µ*j + RTln Cj + zjFE + VjP X
significativo esclusivamente nei movimenti osmotici
dell’acqua

“L’importanza del concetto del potenziale chimico è che somma tutte le forze che possono agire su una molecola e che portano al trasporto netto” In generale, la diffusione ( o trasporto passivo) muove sempre le molecole da zone a potenziale chimico superiore verso aree con potenziale chimico minore. Il movimento contro gradiente di potenziale chimico indica un trasporto attivo

µis = µ*
s + RTln Cis
µos = µ*
s + RTln Cos
saccarosio
i = interno O = esterno

Possiamo calcolare la differenza del potenziale chimico del saccarosio fra le soluzioni all’interno e all’esterno della c e l l u l a i n d i p e n d e n t e m e n t e d a l meccanismo di trasporto

Cos
Cis
= RT ln
Forza motrice Δµs = µos - µi
s =
= (µ*s + RTln Co
s) - (µ*s + RTln Ci
s) =
= RT(ln Cos- ln Ci
s) =
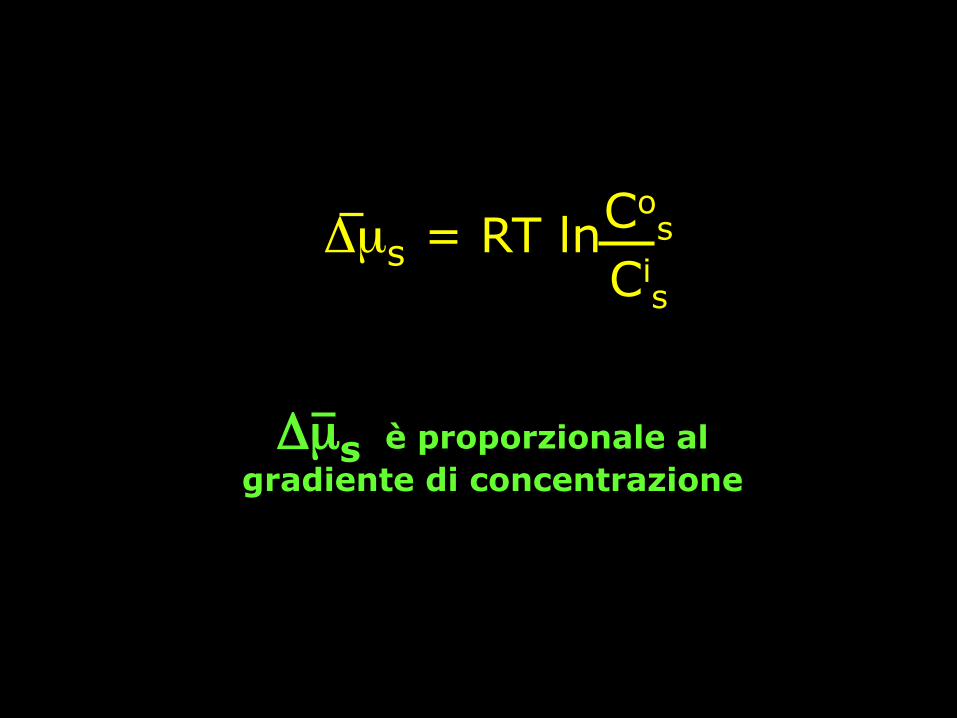
Δµs è proporzionale al gradiente di concentrazione
Cos
Cis
Δµs = RT ln

relazione tra il potenziale chimico µ e il trasporto di molecole attraverso una membrana

ione K+
(Bisogna considerare la componente elettrica del potenziale chimico)
CoK
CiK
ΔµK = RT ln + zF(Eo-Ei)
dato che la carica elettrostatica del K+ z=+1
CoK
CiK
ΔµK = RT ln + F(Eo-Ei)
Potenziale elettrico

gli ioni diffondono sia in risposta ai loro gradienti di concentrazione
che a qualsiasi differenza di potenziale elettrico, se viene
applicato un voltaggio (un campo elettrico) fra i due scomparti gli
ioni possono essere condotti contro un gradiente di
concentrazione

cosa succede se una membrana ha una permeabilità maggiore per il K+ che per il Cl-?
I due i on i d i f f onde ranno attraverso la membrana a velocità diversa. Il risultato sarà una debole separazione di c a r i che che gene re rà un potenziale elettrico attraverso la membrana. Il potenziale di membrana che si forma è detto potenziale di diffusione
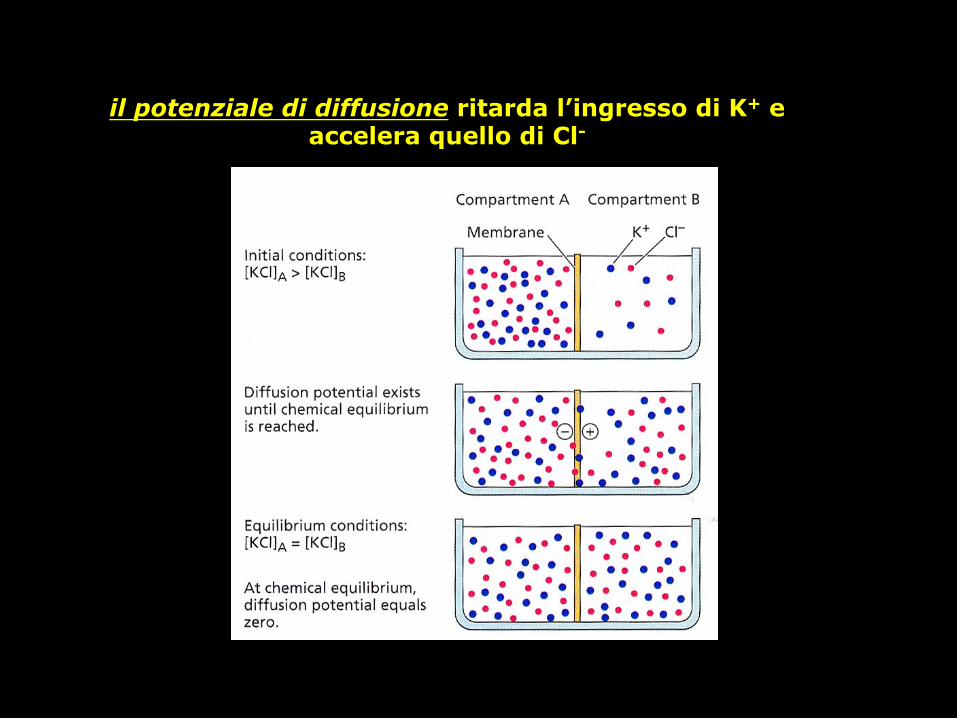
il potenziale di diffusione ritarda l’ingresso di K+ e accelera quello di Cl-

la presenza di un potenziale di membrana indica una distribuzione disuguale degli ioni attraverso la
membrana
trascurabile in termini chimici
-100 mV → un anione in più su 100.000
PRINCIPIO DELL’ELETTRONEUTRALITÀ
una soluzione contiene sempre un numero uguale di anioni e cationi

cosa succede se la membrana è permeabile solo al K+?
all’equilibrio [K+]B > [Cl-]B
all’equilibrio si genera un:
POTENZIALE DI MEMBRANA che bilancia il
GRADIENTE DI CONCENTRAZIONE

quando la distribuzione di un soluto attraverso la membrana raggiunge l’equilibrio, il flusso
passivo (J) (cioè la quantità di soluto che attraversa un’unità di area di membrana
nell’unità di tempo) è lo stesso in entrambe le direzioni
Jo→i = Ji→o

Jo→i = Ji→o
Ei-Eo = RT zjF
ln Cj
o
Cji
equazione di Nerst
quindi per ogni ione
µ*j + RTln Cj
o + zjFEo = µ*j + RTln Cj
i + zjFEi risolvendo l’equazione si ottiene la differenza del potenziale elettrico fra i due scomparti
i potenziali elettrochimici saranno uguali µjo = µj
i

ΔEn = Ei-Eo
potenziale di Nerst
ΔEn = 2.3RT zjF
log Cj
o
Cji
per uno ione univalente a 25°C
ΔEn = 59 log Cj
o
Cji
La differenza di potenziale elettrico è conosciuta come il

all’equilibrio, la differenza di concentrazione di uno ione tra due scomparti è bilanciata dalla
differenza di voltaggio tra gli scomparti
ΔEn=59 log Cj
o
Cji
stabilendo un ΔE di 59 mV mantiene una differenza di
concentrazione di 10 volte per uno ione che viene trasportato
tramite diffusione passiva Co/Ci=10/1
log 10=1

tutte le cellule possiedono un potenziale di membrana, dovuto alla distribuzione asimmetrica tra l’interno e l’esterno della cellula. Questi potenziali di membrana possono essere facilmente determinati inserendo un
microelettrodo in una cellula e misurando la differenza di voltaggio fra l’interno della cellula e il mezzo esterno
in cui la cellula è immersa
-240 mV < ΔEn > -60 mV

Trasporto passivo = frecce tratteggiate Trasporto attivo = frecce intere

EQUAZIONE DI GOLDMAN
Numerosi ioni permeano simultaneamente le membrane cellulari; quindi il potenziale di Nerst per ogni singola specie ionica descrive raramente il potenziale di diffusione della membrana. Invece tutti i potenziali di diffusione attraverso la membrana sono descritti dall’equazione di Goldman

ΔE con equazione di Goldman = -50/-40 mV
misura sperimentale ΔE = -240/-60 mV
Nelle piante i potenziali di membrana misurati sperimentalmente sono spesso molto più negativi di quelli calcolati dall’equazione di Goldman
Oltre al potenziale di diffusione, esiste una seconda componente del potenziale di membrana. L’eccesso di voltaggio è dato dalla H+-ATPasi elettrogenica della membrana plasmatica. Si genera un voltaggio attraverso la membrana ogni qualvolta uno ione si sposta dentro o fuori dalla cellula senza essere controbilanciato da uno ione di carica opposta. Tale meccanismo di trasporto attivo, definito POMPA ELETTROGENICA, che tenderà a spostare il potenziale di membrana lontano rispetto al valore previsto dall’equazione di Goldman
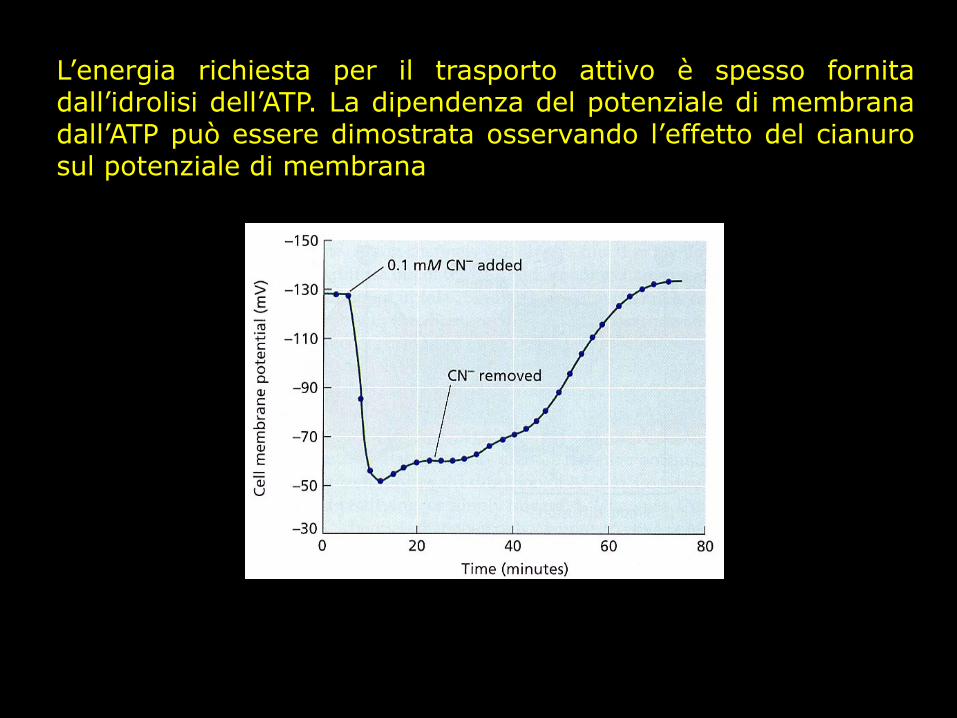
L’energia richiesta per il trasporto attivo è spesso fornita dall’idrolisi dell’ATP. La dipendenza del potenziale di membrana dall’ATP può essere dimostrata osservando l’effetto del cianuro sul potenziale di membrana

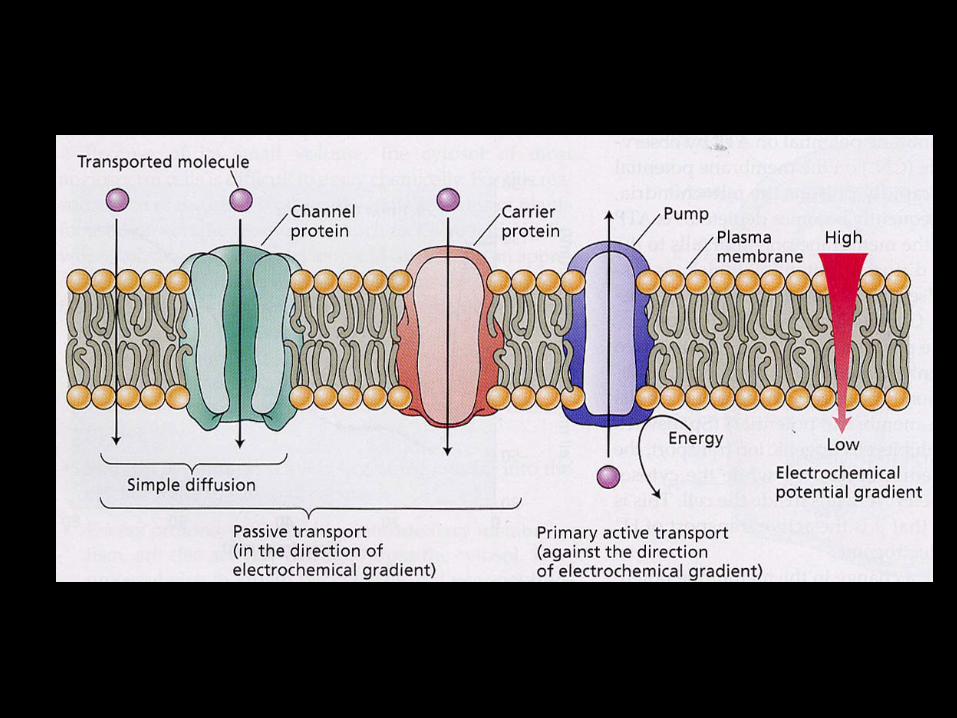
Due tipi di trasportatori aumentano il movimento di soluti attraverso la membrana: Canali sono delle proteine transmembrana che fungono da pori selettivi nella membrana Carrier la sostanza che viene trasportata è inizialmente legata da un sito attivo posto sul carrier proteico. Il legame porta a un cambiamento conformazionale della proteina, che espone la sostanza alla soluzione dalla parte opposta della membrana.
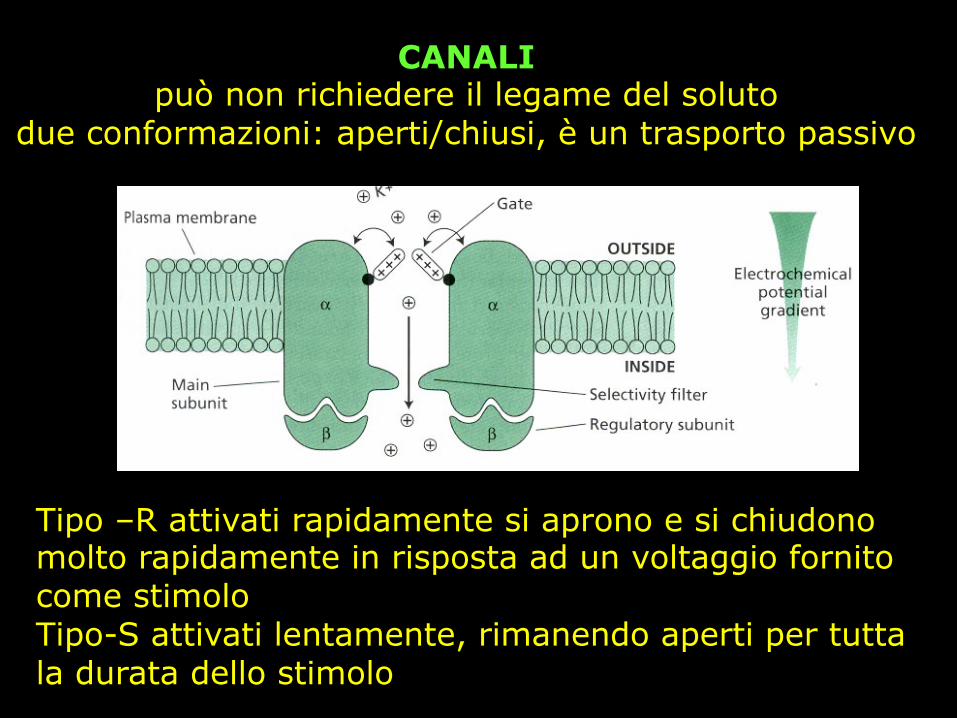
CANALI può non richiedere il legame del soluto
due conformazioni: aperti/chiusi, è un trasporto passivo
Tipo –R attivati rapidamente si aprono e si chiudono molto rapidamente in risposta ad un voltaggio fornito come stimolo Tipo-S attivati lentamente, rimanendo aperti per tutta la durata dello stimolo
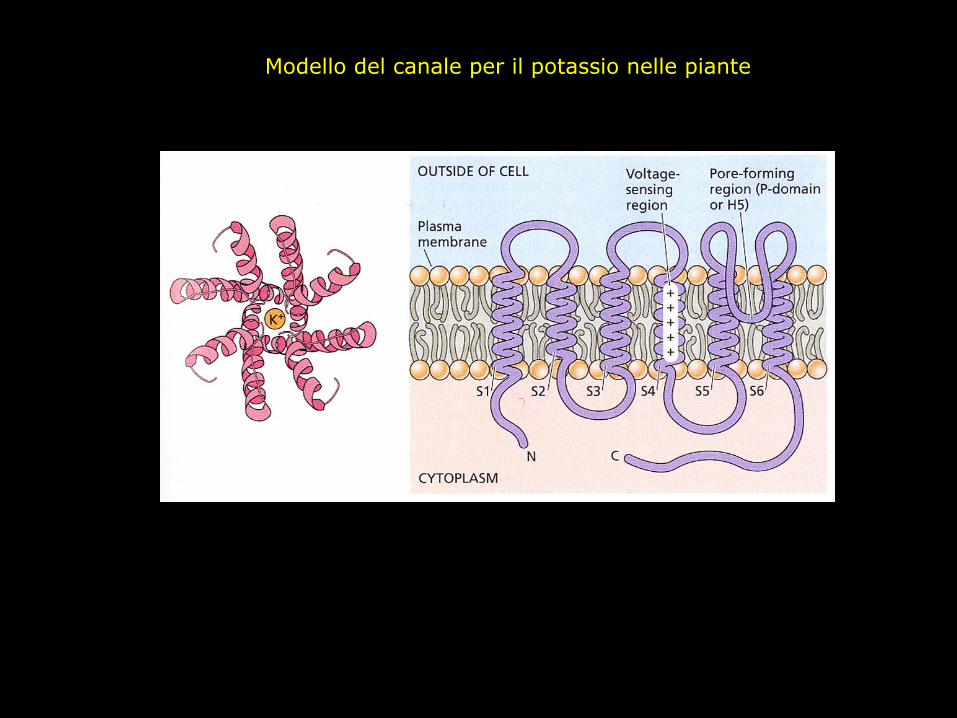
Modello del canale per il potassio nelle piante
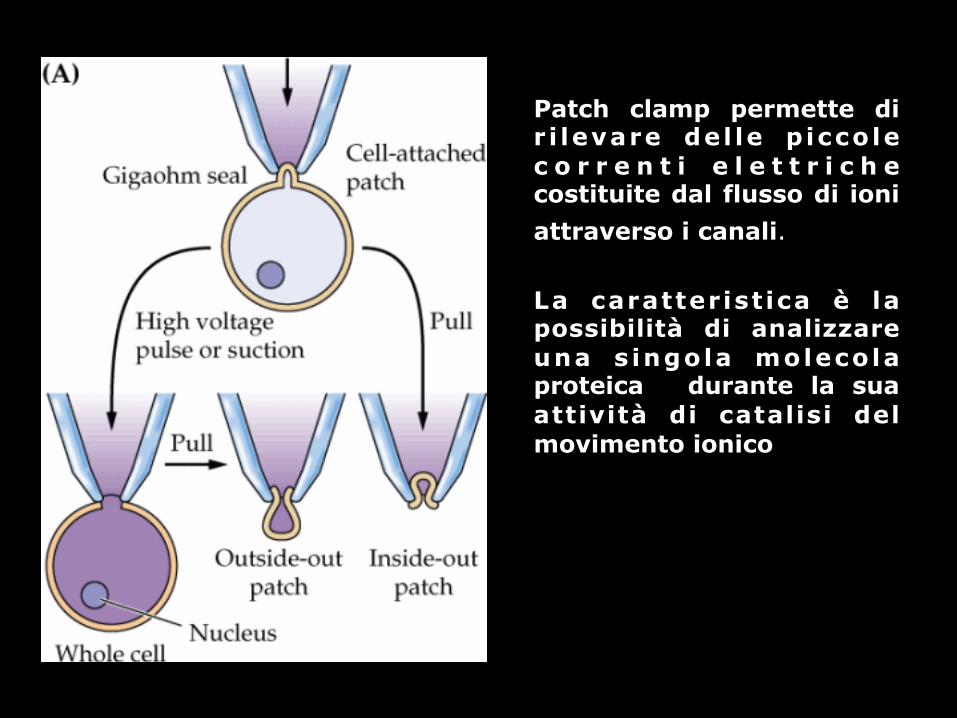
Patch clamp permette di r i levare del le p iccole c o r r e n t i e l e t t r i c h e costituite dal flusso di ioni attraverso i canali. La caratterist ica è la possibilità di analizzare una s ingola molecola proteica durante la sua attività di catalisi del movimento ionico

voltage-clamp misura la corrente elettrica e i potenziali di membrana che vengono applicati dall’esterno ad una membrana e quindi all’apertura di un numero crescente di canali in risposta ai potenziali applicati
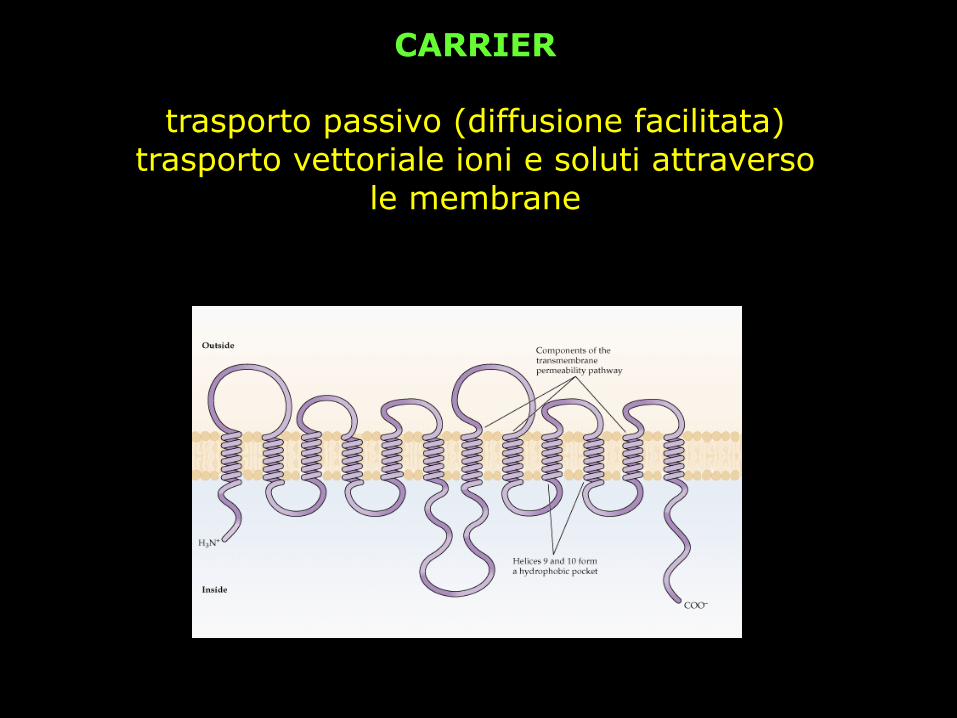
CARRIER
trasporto passivo (diffusione facilitata) trasporto vettoriale ioni e soluti attraverso
le membrane

la velocità di trasporto è più lenta rispetto ai canali (Vcanale = 106 Vcarrier)
Il meccanismo proposto non prevede un movimento dei siti di legame del soluto da un lato all’altro della membrana, ma piuttosto che la reazione di trasporto sia conseguente a variazioni conformazionali del carrier

le cinetiche di attività dei carrier sono descrivibili con l’equazione di
Michaelis-Menten
La concentrazione di substrato che determina una velocità di trasporto corrisponde alla metà della velocità massima rappresenta la costante di Michaelis-Menten

PRINCIPALI IONI E SOLUTI TRASPORTATI DA CARRIER NH4
+, NO3-, Pi, SO4
2-, zuccheri, amminoacidi, basi azotate
alta selettività per la molecola da trasportare

TRASPORTO ATTIVO PRIMARIO il trasporto è accoppiato a una fonte di energia metabolica idrolisi di ATP una reazione di ossido-riduzione
POMPE elettrogeniche → H+-ATPasi è riferito al trasporto di ioni che coinvolge il movimento netto di carica attraverso la membrana elettroneutre → H+/K+-ATPasi (animali)
→ Na+/K+-ATPasi (animali)
non coinvolge il movimento netto di carica
Le proteine di membrana responsabili del trasporto attivo primario sono definite pompe

principali pompe elettrogeniche H+-ATPasi di plasmalemma H+-ATPasi vacuolare H+-pirofosfatasi (vacuolo e golgi)
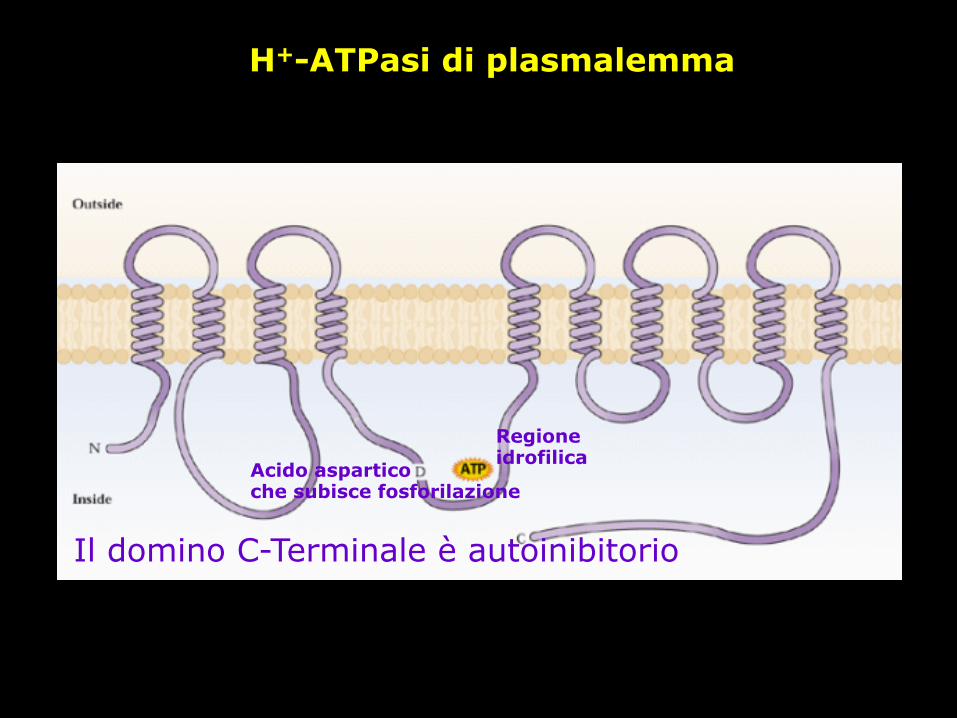
H+-ATPasi di plasmalemma
Il domino C-Terminale è autoinibitorio
Regione idrofilica Acido aspartico
che subisce fosforilazione
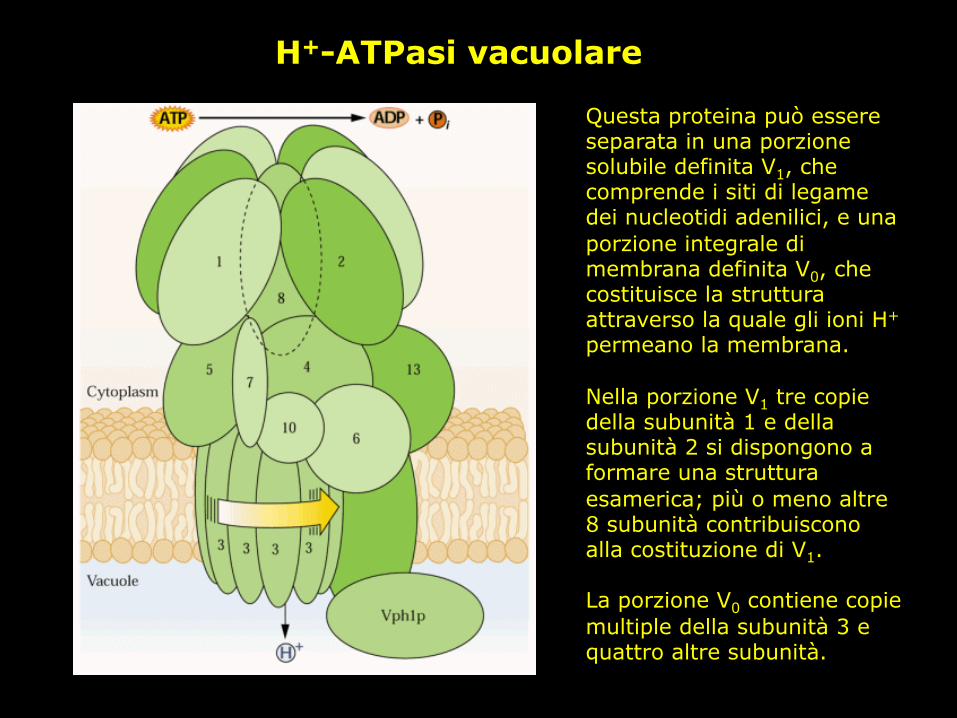
H+-ATPasi vacuolare
Questa proteina può essere separata in una porzione solubile definita V1, che comprende i siti di legame dei nucleotidi adenilici, e una porzione integrale di membrana definita V0, che costituisce la struttura attraverso la quale gli ioni H+ permeano la membrana. Nella porzione V1 tre copie della subunità 1 e della subunità 2 si dispongono a formare una struttura esamerica; più o meno altre 8 subunità contribuiscono alla costituzione di V1. La porzione V0 contiene copie multiple della subunità 3 e quattro altre subunità.
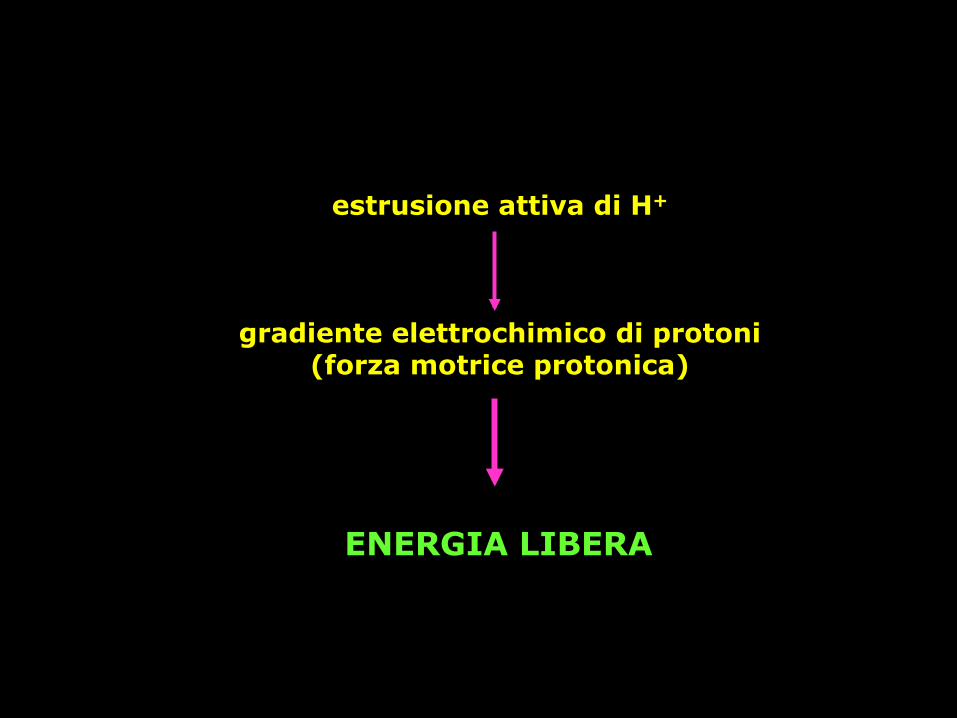
estrusione attiva di H+
gradiente elettrochimico di protoni (forza motrice protonica)
ENERGIA LIBERA

il movimento di protoni secondo il loro gradiente elettrochimico può essere accoppiato al lavoro cellulare
ΔµH+ = FΔE + 2.3RT log [H+]i
[H+]o
Δp = = ΔE + 2.3RT log [H+]i
[H+]o
ΔµH+
F F

poiché pH = -log[H+]
log [H+]i
[H+]o =- (pHi-pHo)= ΔpH
Δp = ΔE - 2.3RT F
ΔpH
Δp = ΔE -59 ΔpH Forza proton motrice è l ’energia l ibera accumulata sotto forma di gradiente di H+

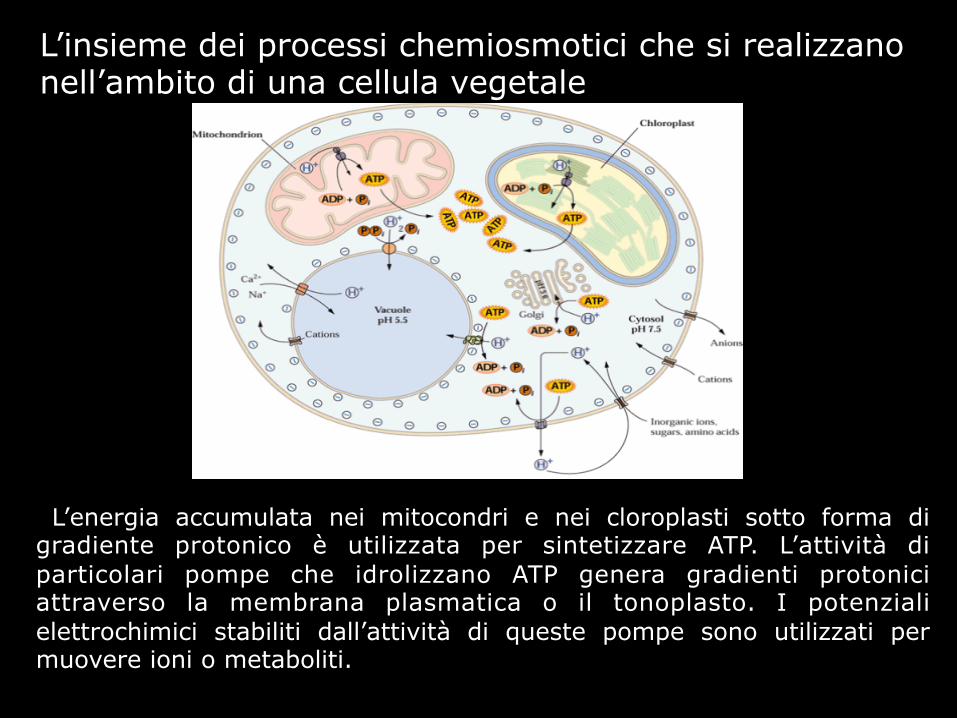
L’energia accumulata nei mitocondri e nei cloroplasti sotto forma di gradiente protonico è utilizzata per sintetizzare ATP. L’attività di particolari pompe che idrolizzano ATP genera gradienti protonici attraverso la membrana plasmatica o il tonoplasto. I potenziali elettrochimici stabiliti dall’attività di queste pompe sono utilizzati per muovere ioni o metaboliti.
L’insieme dei processi chemiosmotici che si realizzano nell’ambito di una cellula vegetale

TRASPORTO ATTIVO SECONDARIO
la forza proton motrice può guidare il trasporto di altre sostanze contro il gradiente elettrochimico
SIMPORTO protone e molecola si muovono nella stessa direzione ANTIPORTO protone e molecola si muovono in direzioni opposte


L’energia che garantisce lo svolgimento del processo è accumulata tramite il gradiente di protoni viene utilizzata per assorbire il substrato contro il suo gradiente di concentrazione


simporto glucosio-H+
L’aumento osservabile di pH indica che i protoni sono scomparsi dal liquido e che allo stesso tempo diminuisce il potenziale di membrana della cellula

Le proprietà di trasporto di un soluto possono cambiare a diverse concentrazioni del soluto A b a s s e c o n c e n t r a z i o n i d i s a c c a r o s i o l a v e l o c i t à d i assorbimento del saccarosio mostra cinetiche di saturazione t i p i c h e d i u n a m o l e c o l a trasportatrice simporto H+-saccarosio
alte concentrazioni saccarosio, la velocità di assorbimento aumenta linearmente trasporto passivo

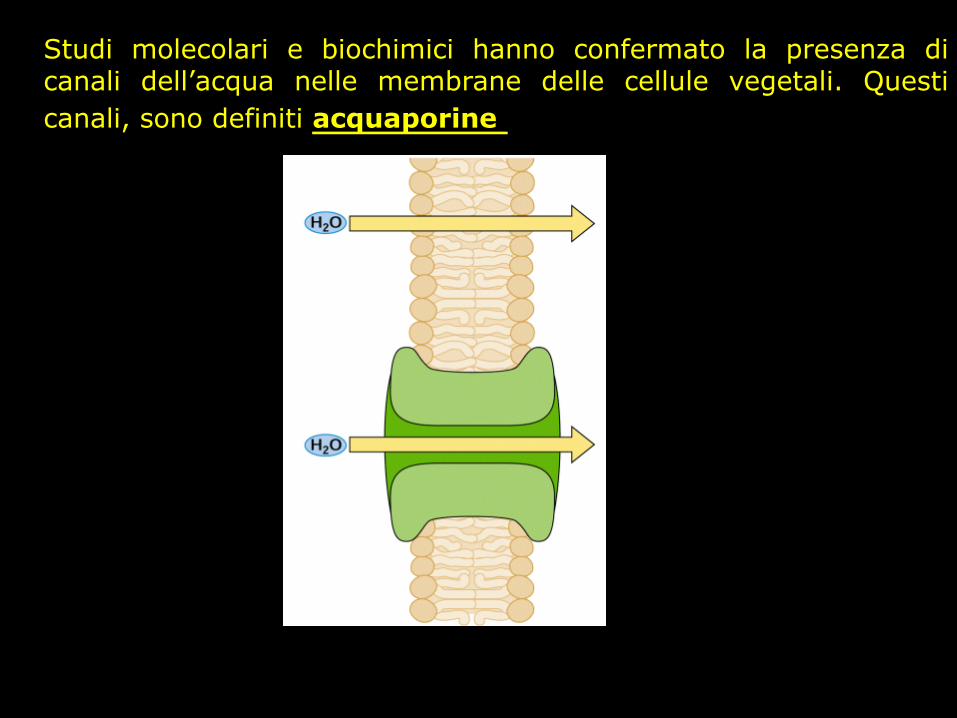
Studi molecolari e biochimici hanno confermato la presenza di canali dell’acqua nelle membrane delle cellule vegetali. Questi canali, sono definiti acquaporine

Le acquaporine
Sono membri della famiglia dei canali transmembrana conosciuta come major intrinsic protein (MIP) hanno dimensioni piuttosto limitate (25-30 kDa), presentano 6 regioni transmembrana e possiedono una sequenza interna ripetuta indicativa della loro origine da un gene che ha subito fenomeni di duplicazione e
fusione. Le acquaporine sono caratterizzate anche da residui NPA (Asn-Pro-
Ala) altamente conservati sia nella metà N- sia in quella C-terminale della proteina. Nelle piante sono state identificate
acquaporine sulle membrane vacuolari e su quelle plasmatiche ed è quindi possibile definire due subfamiglie:quella delle proteine
integrali del tonoplasto (TIP) e quella delle proteine integrali della membrana plasmatica (PIP)
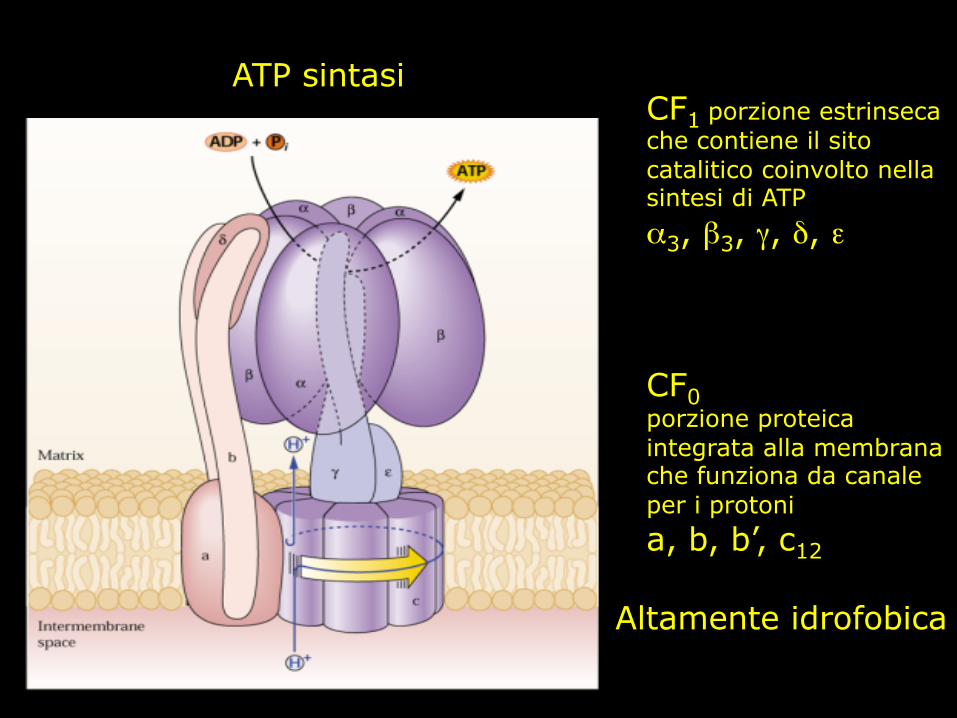
CF1 porzione estrinseca che contiene il sito catalitico coinvolto nella sintesi di ATP α3, β3, γ, δ, ε CF0 porzione proteica integrata alla membrana che funziona da canale per i protoni a, b, b’, c12
ATP sintasi
Altamente idrofobica

F-ATPasi dei mitocondri e dei cloroplasti. V-ATPasi, presente nel tonoplasto. Pompa i protoni all’interno del vacuolo, generando la forza motrice per diversi sistemi di trasporto P-ATPasi, presente sulla membrana plasmatica
H+-ATPasi

RUOLO FISIOLOGICO DELL’H+-ATPasi DI PLASMALEMMA
mantenimento del pH del citoplasma
↑ pH interruzione
della dormienza dei semi
↑ pH sintesi DNA e
divisione cellulare
≠ pH risposte di difesa
a patogeni
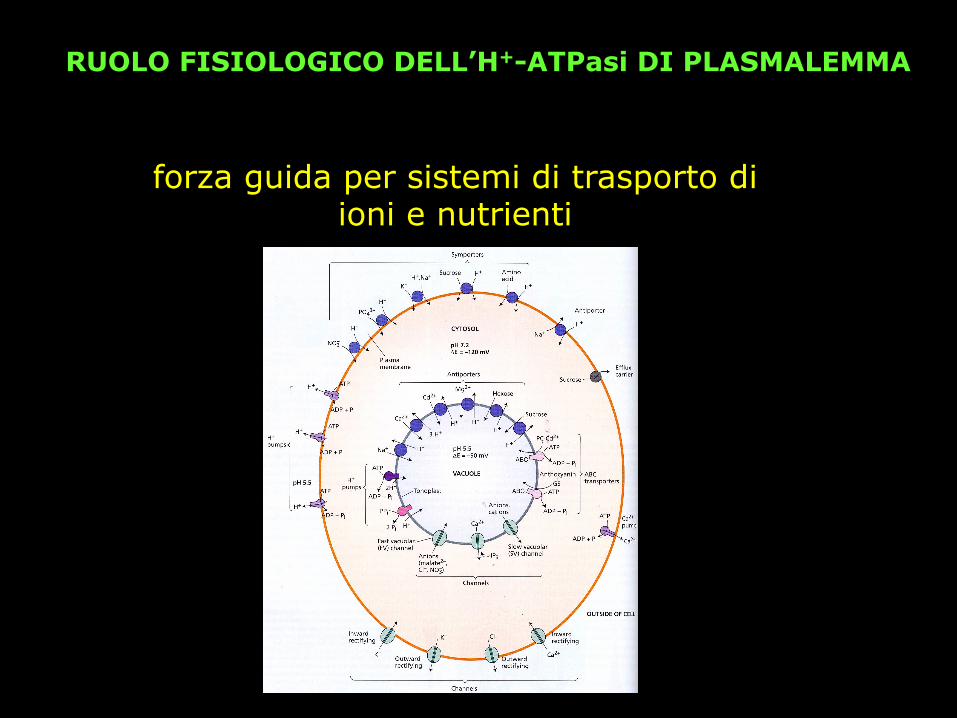
RUOLO FISIOLOGICO DELL’H+-ATPasi DI PLASMALEMMA
forza guida per sistemi di trasporto di ioni e nutrienti
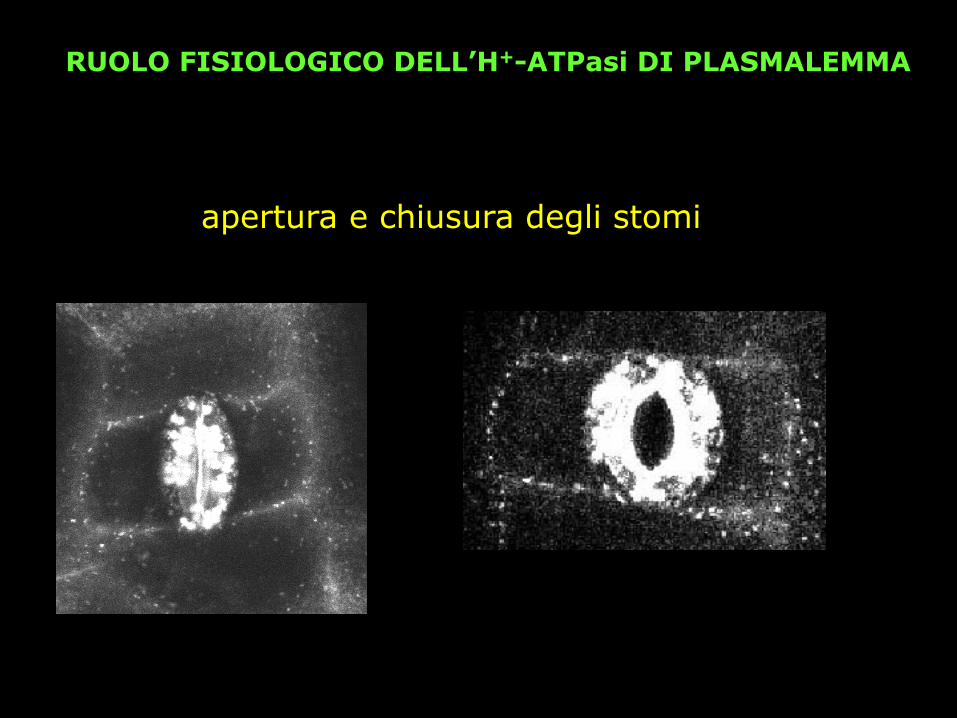
RUOLO FISIOLOGICO DELL’H+-ATPasi DI PLASMALEMMA
apertura e chiusura degli stomi
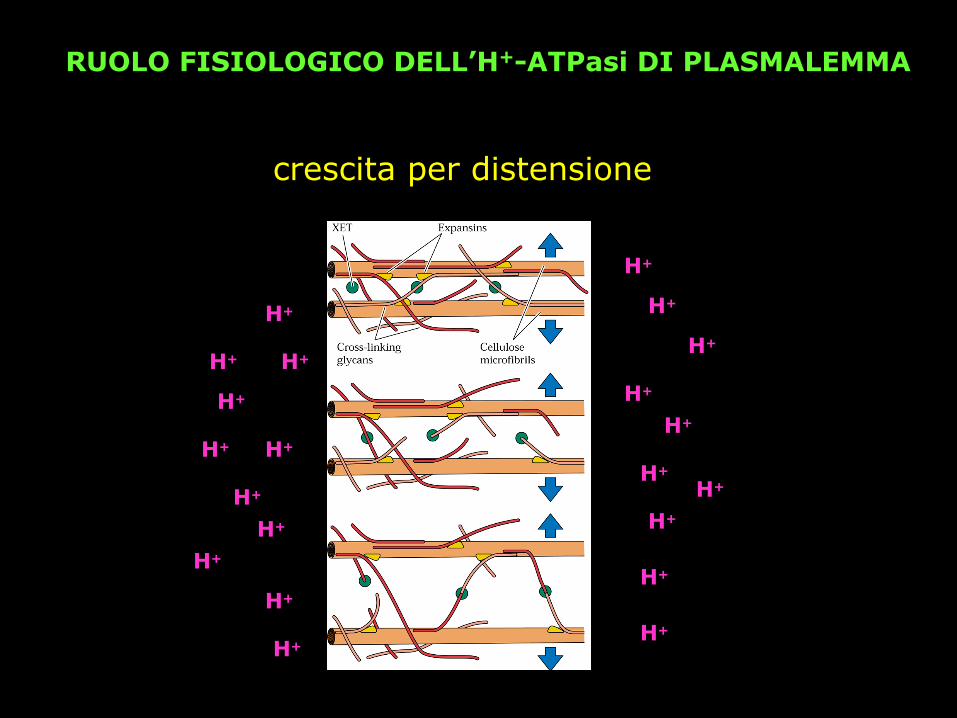
RUOLO FISIOLOGICO DELL’H+-ATPasi DI PLASMALEMMA
crescita per distensione
H+
H+
H+
H+
H+
H+
H+
H+
H+
H+
H+
H+
H+
H+
H+
H+
H+
H+
H+
H+
H+

possibili meccanismi che determinano un aumento di attività dell’H+-ATPasi
fattori che influenzano l’attività dell’H+-ATPasi stress salino luce auxina patogeni
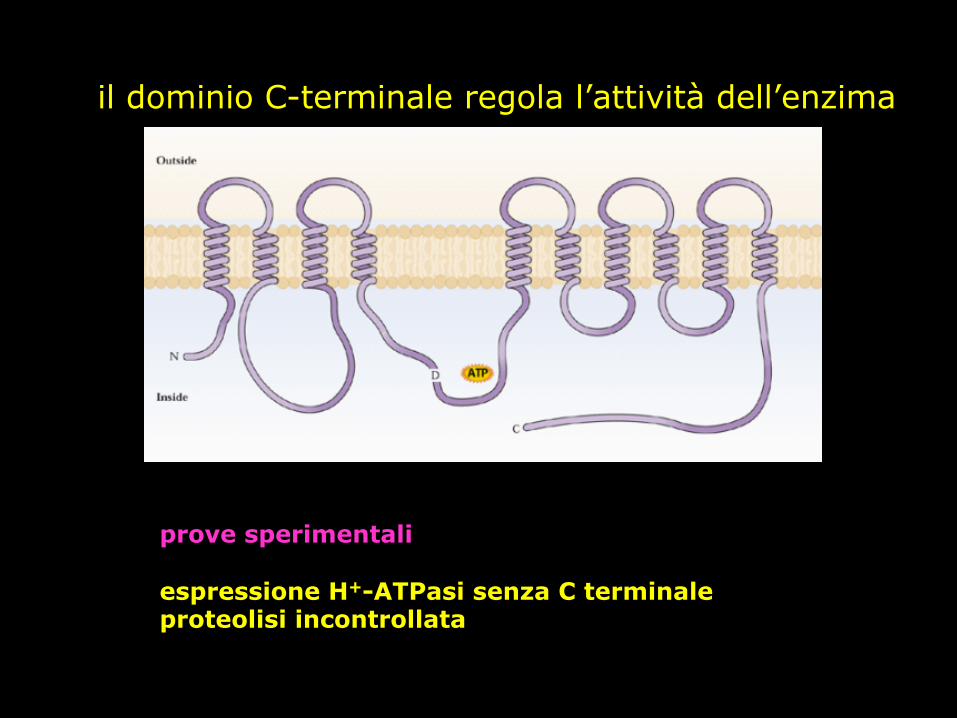
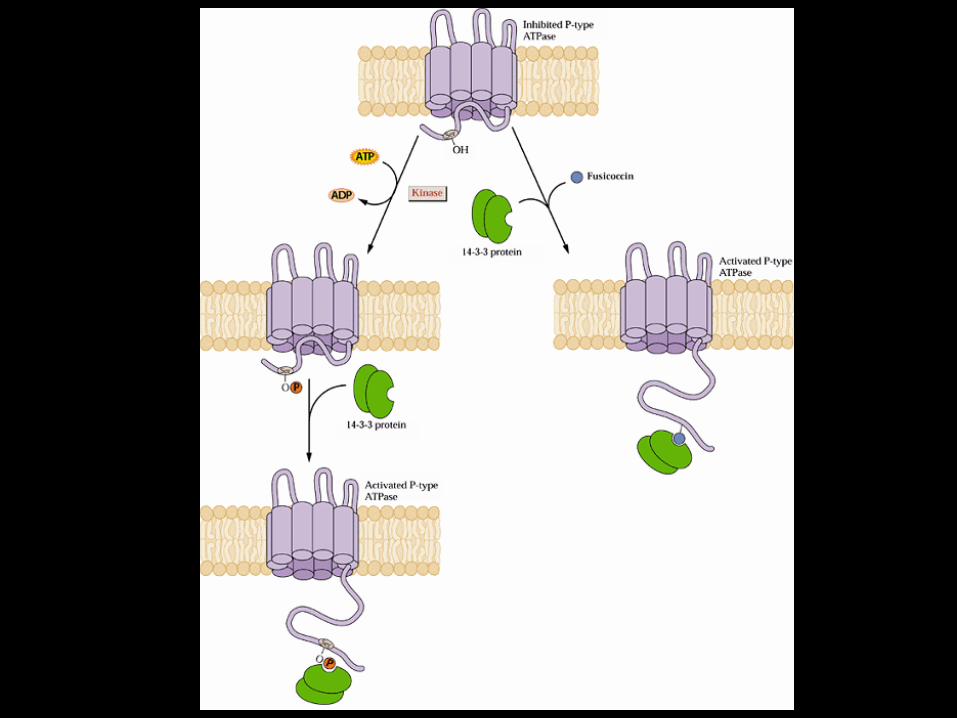
il dominio C-terminale regola l’attività dell’enzima
prove sperimentali espressione H+-ATPasi senza C terminale proteolisi incontrollata

LA FUSICOCCINA
metabolita fitotossico prodotto dal fungo Fusicoccum amygdali
l’infezione provoca una stimolazione
incontrollata dell’H+-ATPasi Il dominio C-terminale non è più in grado di controllare l’attività dell’H+
ATPasi
apertura incontrollata degli stomi
morte per avvizzimento

la fusicoccina promuove il legame di una proteina 14-3-3 sul dominio C-terminale dell’H+-ATPasi
Proteine 14-3-3 Classe di proteine regolatrici diffuse in tutti gli eucarioti Proteine dimeriche. Ogni monomero ha un PM di circa 30 kDa Interagiscono con numerose proteine bersaglio, tramite il riconoscimento di particolari sequenze fosforilate su residui di serina

Ruolo delle 14.3.3
- Regolazione dell’H+ ATPasi - Regolazione della trascrizione genica - Regolazione dell’H+ sintasi - Processo apoptotico?
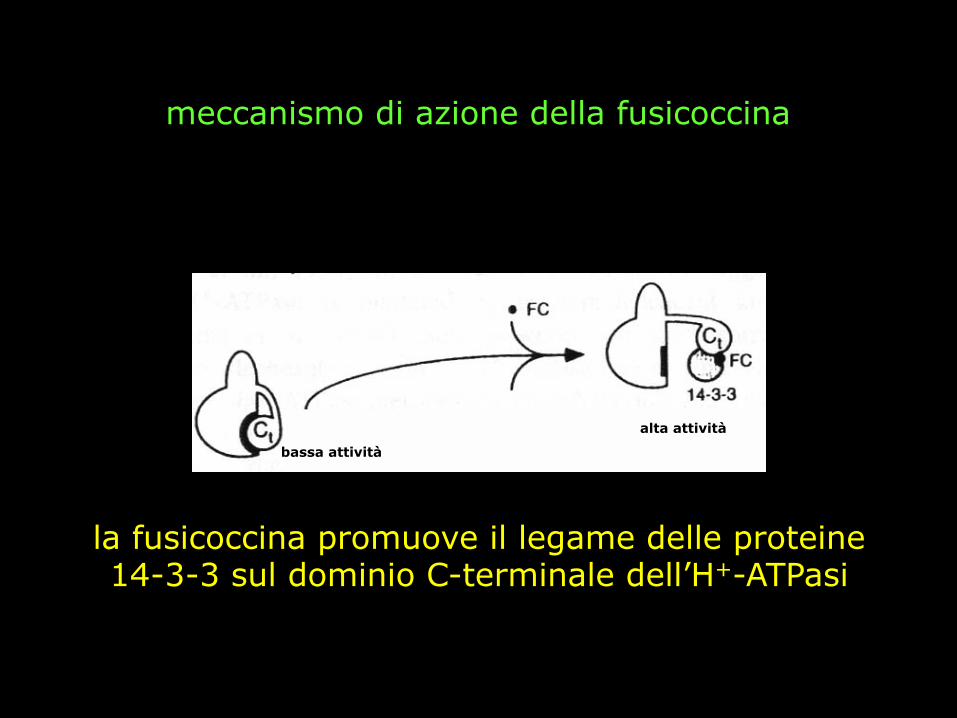
la fusicoccina promuove il legame delle proteine 14-3-3 sul dominio C-terminale dell’H+-ATPasi
meccanismo di azione della fusicoccina
bassa attività
alta attività
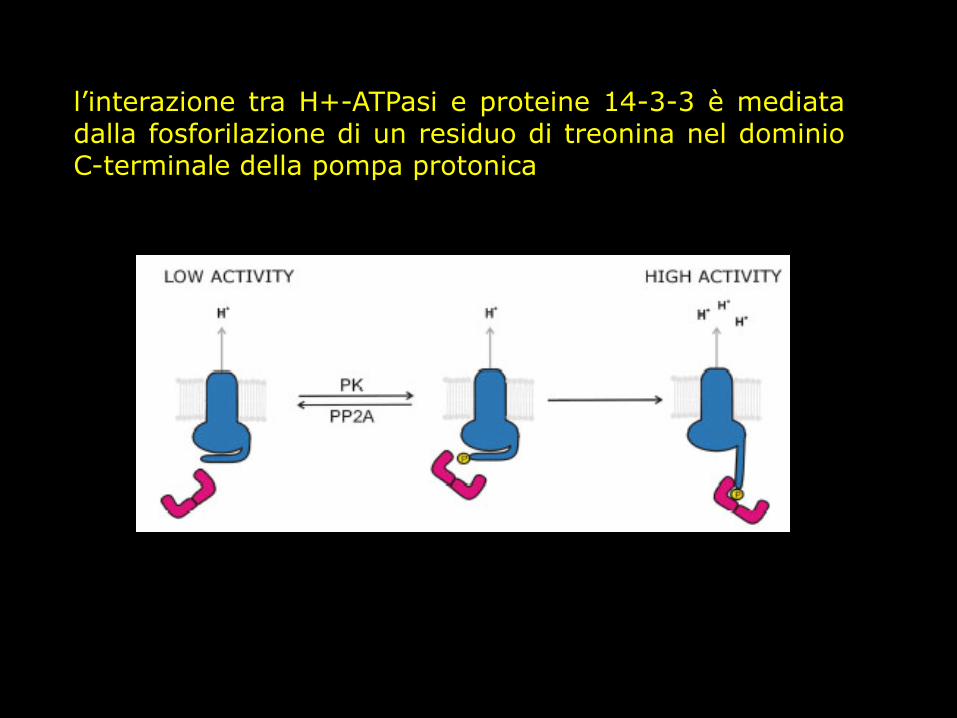
l’interazione tra H+-ATPasi e proteine 14-3-3 è mediata dalla fosforilazione di un residuo di treonina nel dominio C-terminale della pompa protonica