transfer reactions of acetyl phosphate catalyzed · transfer reactions of acetyl phosphate...
TRANSCRIPT

TRANSFER REACTIONS OF ACETYL PHOSPHATE CATALYZED BY GLYCERALDEHYDE-3-PHOSPHATE DEHYDROGENASE*
BY JANE HARTINGt AND SIDNEY F. VELICK
(Prom the Department of Biological Chemistry, Washington University School of Medicine, St. Louis, Missouri)
(Received for publication, July 20, 1953)
Glyceraldehyde-3-phosphate dehydrogenase catalyzes the oxidative phos- phorylation of acetaldehyde (1) and a number of other aliphatic aldehydes (2, 3) and also has a transacetylating activity (4). In the present paper the reactions of acetyl phosphate which characterize the transacetylating activity of the enzyme are presented. These reactions include (a) the phos- phate exchange between acetyl phosphate and radioact,ive inorganic phos- phate, (b) t.he arsenolysis of acetyl phosphate, and (c) the acetylation of the thiol groups of coenzyme A (CoA) and glutathione to form X-acetyl CoA and S-acetylglutathione.
Although transfer reactions with acetyl phosphate as substrate have not previously been observed to be catalyzed by enzymes from mammalian tissues or yeast, numerous species of bacteria contain a phosphotransacet- ylase which catalyzes similar reactions (5, 6). The crystalline dehydro- genase differs from the bacterial enzyme in that it cat,alyzes oxidative as well as transfer reactions. Both of the activities of the dehydrogenase may be described by a single reaction mechanism in which an acetyl-enzyme com- pound is the common intermediate.
Methods
The preparations of the crystalline dehydrogenase from rabbit muscle and yeast were obtained as previously indicated (2). The pigeon liver enzyme used in the acetylation of sulfanilamide was fractionated by the method of Chou et al. (7). We are indebted to Dr. Joseph Stern and Dr. Sever0 Ochoa for samples of the crystalline condensing enzyme (8).
Diphosphopyridine nucleot~ide (DPN) was purified by ion exchange chro-
* This work was supported in part by a grant from the American Cancer Society on the recommendat.ion of the Committee on Growth of the National Research Council.
The material in this paper is taken from the dissertation submitted by Jane Harting in partial fulfilment of the requirements for the degree of Doctor of Philoso- phy in Biological Chemistry, Washington University, XIny, 1952.
t Predoctoral Fellow of the National Institutes of Health, United States Public Health Service, and Baxter Fellow. Present address, Biochemical Research Labo- ratory, Massachusetts General Hospital, Boston.
867
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

868 TRANSFER REACTIONS OF ACETYL PHOSPHATE
matography on Dowex 50 according to an unpublished method of Dr. A. Kornberg and Dr. B. Horecker.’ Samples of coenzyme A with activities of 170 and 330 units per mg. mere the gifts of Dr. F. Lipmann, Dr. J. Gregory, and Dr. G. Novelli. A CoA unit is defined in terms of the assay system used by Kaplan and Lipmann (9). Lithium monoacetyl phosphate was prepared by the method of Stadtman and Lipmann (10). Acetyl CoA, 40 per cent pure, was furnished by Dr. C. Martius. Oxalacetate was prepared by the method of Krampitz and Werkman (11). The analytical methods are described in the text and in the preceding paper (2).
TABLE I
Exchange between Inorganic Phosphate and Acetyl Phosphate Catalyzed by Glyceraldehyde-S-phosphate Dehydrogenase from Rabbit Muscle
Each reaction mixture contained 21 PM of acetyl phosphate and 23 .UM of Pss-la- beled inorganic phosphate (specific activity about 3100 c.p.m. per y of P), 37 MM of sodium Veronal, pH 8.1, 2.2 .UM of L-cysteine in a total volume of 2.2 ml. Tubes I to IV contained 0.056 PM of glyceraldehyde dehydrogenase in the following forms: Tube I, 2 moles of bound DPN per mole of enzyme; Tube II, bound DPN removed from enzyme by adsorption on charcoal; Tube III, DPN restored to the charcoal- treated enzyme (enzyme-DPN ratio 1:3); Tube IV, TPN added to an enzyme from which bound DPN had been removed (enzyme-TPN ratio 1:3). After incubation for 15 minutes the inorganic phosphate was separated from the acetyl phosphate by calcium precipitation. Phosphate determinations and radioactivity measurements were made on suitable aliquots of the two fractions from the calcium precipitation.
Tube No. Type of enzyme Inorganic phosphate Acetyl phosphate
I II
III IV
Enzyme-(DPN)2 complex DPN removed
“ restored “ removed, TPN added
c.g.m. per y P c.p.m. fW y P
1910 1090 3130 43 2575 700 3210 22
EXPERIMENTAL
Phosphate Exchange between Inorganic Phosphate and Ace&l Phosphate- When crystalline glyceraldehyde-3-phosphate dehydrogenase is incubated with inorganic phosphate labeled with P32 and unlabeled acetyl phosphate, the organic phosphate becomes labeled and the specific activity of the in- organic phosphate decreases. An exchange of phosphate therefore occurs which, in the experiments described in Table I, is 70 per cent complete in 15 minutes. Cysteine is required for maximal rates under these conditions.
The dehydrogenase crystallized from rabbit muscle contains 2 equivalents of bound DPN per mole of enzyme (12, 13). When the bound nucleotide
i Personal communication.
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

J. HARTING AND S. F. VELICK 869
is removed by adsorption on charcoal, the enzyme can no longer catalyze the phosphate exchange reaction (Table I). The small amount of P32 which appears in the acetyl phosphate fraction probably arises from contamina- tion with inorganic phosphate. When DPN is added back to the enzyme in a molar ratio of 3 : 1, an exchange of phosphate again takes place but only at half the rate obtained with the untreated enzyme-(DPN)z complex. Triphosphopyridine nucleotide (TPN) cannot be used in place of DPN to restore the exchange activity.
An analogous experiment was carried out with the glyceraldehyde-3-phos- phate dehydrogenase crystallized from yeast. After a few recrystalliza- tions this enzyme contains no bound DPN. As shown in Table II, the radioactivity which appears in the acetyl phosphate fraction is about the
TABLE II
Exchange between Inorganic Phosphate and Acetyl Phosphate Catalyzed by Glyceraldehyde-3-phosphate Dehydrogenase from Yeast
The experimental conditions were almost identical with those described in Table I. Tubes II and III contained 0.048 PM of glyceraldehyde phosphate dehydrogenase; 0.19 PM of DPN was added to Tube III. The ratio of enzyme to DPN is 1:4. 25 minute incubation. The phosphate fractionation and analytical determinations were carried out as previously described in Table I.
Tube No.
I II
III
Type of enzyme
No enzyme Enzyme; no DPN Enzyme; DPN added
Inorganic phosphate
c.p.m. gcr y P
1060 1049 804
-
Acetyl phosphate
c.g.m. jer y P
58 47
412
same in the presence and absence of the enzyme. However, when DPN is added to the yeast dehydrogenase, a rapid exchange of phosphate occurs which is about 60 per cent complete after 25 minutes incubation. Thus both the yeast and the mammalian enzymes require DPN for the phosphate exchange reaction.
Arsenolysis oj Acetyl Phosphate-Glyceraldehyde-3-phosphate dehydro- genase also catalyzes the arsenolysis of acetyl phosphate. The accelerated decomposition of acetyl phosphate in the presence of arsenate (Table III) was followed by analyses for acetyl phosphate by the hydroxamic acid method (14). The arsenolysis is inhibited by inorganic phosphate. Ace- tate, however, has no effect. As in the case of the phosphate exchange re- action DPN is required for the arsenolysis (15).
The possibility was considered that either the enzyme or the DPN might contain traces of DPNH which could initiate the reversible reduction of acetyl phosphate to acetaldehyde and thereby account for the P32 exchange
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

870 TR.\NSFER RE.kCTIONS OF .lCETYL PHOSPH.\TE
and arsenolysis. However, this explanation of the transfer reactions ap- pears very unlikely in view of the following observations: (a) The rates of phosphate exchange and arsenolysis are both greatly in excess of the rate of acetaldehyde oxidation under comparable conditions. (6) The exchange reaction can be further separated from the oxidation by lowering the pH from 8.1 to 7.0. This reduces the rate of exchange by 50 per cent and the rate of oxidation of acetaldehyde by about 95 per cent. (c) In those cases in which DPN was added to the DPN-free enzyme, the coenzyme was pur- ified chromatographically under acid conditions which destroy DPXH.
Formation of Acetyl CoA-Since the preceding reactions appeared some- what analogous to those catalyzed by the bacterial phosphotransacetylase
TABLE III
Arsenolysis of Acetyl Phosphate Catalyzed by Glyceraldehyde-3-phosphate Dehydrogenase
The complete system for the arsenolysis reaction contained 4 PM of acetyl phos- phate, 4 PM of sodium arsenate, 40 PM of sodium Verona1 buffer, pH 8.1, 1.5 PM of L-cysteine, 0.028 PM of glyceraldehyde-3-phosphate dehydrogenase. Total volume 2.0 ml. Temperature 23”. Incubation times as indicated. The reaction mixtures were analyzed for acetyl phosphate by the hydroxamic acid method (14). The results are given in micromoles per 2.0 ml.
Components Acetyl phosphate
0 min. I 25 min. I 36 min.
Complete ....................... ..i 4.0 2.7 1.7 No arsenate .................... .../ 4.0 3.7
I 3.5
(16), it was decided to determine whether the dehydrogenase could also cat- alyze the transfer of the acetyl group of acetyl phosphate to CoA. The formation of acetyl CoA was demonstrated by coupling the dehydrogenase reaction with a system known to require acetyl CoA, namely, the acetyl- ation of sulfanilamide by the pigeon liver enzyme (17). The reaction se- quence may be formulated as follows:
(a) Acetyl phosphate + CoA dehydrogenasz acetyl CoA + phosphate
(b) Acetyl CoA + sulfanilamidc pigeon liver acetyl sulfanilamide + CoA
enzyme
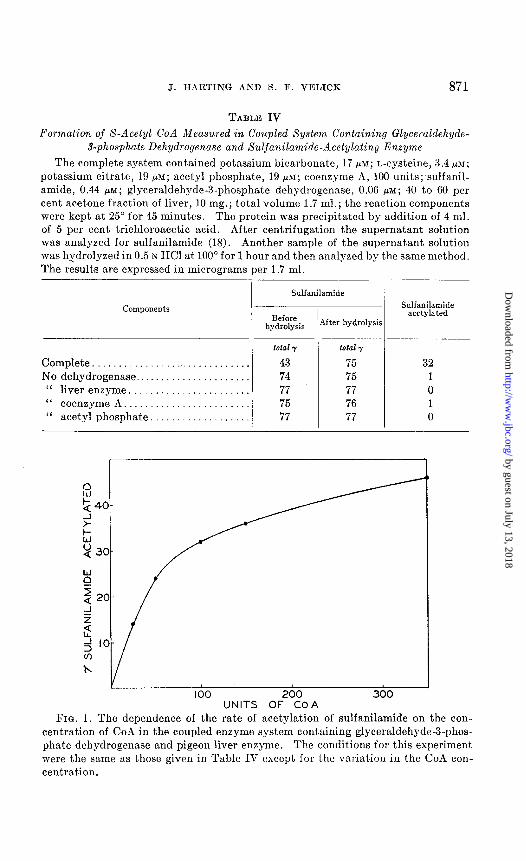
The results obtained with this coupled system are presented in Table IV. It may be seen that the system was inactive when the dehydrogenase, pigeon liver enzyme, CoA, or acetyl phosphate was omitted. Acetate could not replace acetyl phosphate. The rate of acetylation was a function of the CoA concentration as shown in Fig. 1.
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from
J. HARTING AND R. F. VELICK 871
TABLE IV
Formation of S-Acetyl CoA Measured in Coupled System Containing Glyceraldehyde- J-phosphate Dehydrogenase and Suljanilamide-Acetylating Enzyme
The complete system contained potassium bicarbonate, 17 p&r; L-cysteine, 3.4 PY; potassium citrate, 19 pu; acetyl phosphate, 19 ~11; coenzyme A, 100 units; sulfanil- amide, 0.44 PM; glyceraldehyde-3-phosphate dehydrogenase, 0.06 PM; 40 to 60 per cent acetone fraction of liver, 10 mg.; total volume 1.7 ml.; the reaction components were kept at 25” for 45 minutes. The protein was precipitated by addition of 4 ml. of 5 per cent trichloroacetic acid. After centrifugation the supernatant solution was analyzed for sulfanilamide (18). Another sample of the supernatant solution was hydrolyzed in 0.5 N HCl at 100” for 1 hour and then analyzed by the same method. The results are expressed in micrograms per 1.7 ml.
components
Complete .............................. No dehydrogenase ...................... I‘ liver enzyme ....................... “ coenzyme A ........................ “ acetyl phosphate. ..................
-
-
Sulfanilamide
Before hydrolysis A .fter hydrolysi!
tofd y fatal y
43 75 74 75 77 77 75 76 77 77
Sulfanilamide acetylated
32 1 0 1 0
100 200 300 UNITS OF COA
FIG. 1. The dependence of the rate of acetylation of sulfanilamide on the con- centration of CoA in the coupled enzyme system containing glyceraldehyde-3-phos- phate dehydrogenase and pigeon liver enzyme. The conditions for this experiment
were the same as those given in Table IV except for the variation in the CoA con- centration.
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

872 TRANSFER REACTIONS OF ACETYL PHOSPHATE
The above results were confirmed in another coupled system in which the crystalline condensing enzyme (8) was used to measure the acetyl CoA formed in the transfer reaction
(a) Acetyl phosphate + CoA dehydrogenase acetyl CoA + phosphate
@) Acetyl CoA + oxalacetate condensing enzym5 citrate + CoA
TABLE V
Formation of S-Acetyl CoA from Acetyl Phosphate Catalyzed by Glyceraldehyde-3-phosphate Dehydrogenase in Presence and
Absence of Glutathione
Reaction System a contained 70 PM of potassium bicarbonate, 30 pM of glutathione adjusted to pH 7.5, 4 PM of MgC12, 15 PM of acetyl phosphate, and 25 pM of oxalace- tate; CoA concentration varied as indicated; 100 units of condensing enzyme; 0.033 PM of glyceraldehyde&phosphate dehydrogenase. Final volume 1.7 ml. In Re- action System b, the total micromoles of potassium bicarbonate, MgClz, oxalacetate, and acetyl phosphate were the same as those in Reaction System a. In addition, Reaction System b contained 66 units of condensing enzyme, 0.058 PM of glyceralde- hyde dehydrogenase (preparation recrystallized in the presence of 0.001 M Versene and in the absence of glutathione or cysteine), and CoA as indicated. Total volume 2.1 ml. The CoA in Tests I to VII had an activity of 170 units per mg. and in Test VIII an activity of 330 units per mg. In the absence of glutathione, the CoA of Tests VII and VIII was converted to the reduced form with hydrogen sulfide. The coenzyme in Test VI was not treated with hydrogen sulfide and was present as oxi- dized CoA. All samples were incubated for 1 hour and then analyzed for citric acid (19).
Test No.
I II
III IV
V VI
VII VIII
a “ “
“ (no condensing enzyme) “ ( “ dehydrogenase) b “ C‘
Reaction system ( Xutathione CoA Citrate formed
I- PM units
30 200 30 100 30 None 30 200 30 200 None 500 (oxidized CoA)
“ 500 (reduced “ ) “ 350 “ “
NM per hr.
1.10 0.55 0.03 0.01 0.04 0.00 1.04 0.51
The experimental conditions for this system are shown in Table V. It can be seen (Tests I, II, and III) that the synthesis of citrate is a linear func- tion of the CoA concentration. The system was inactive in the absence of glyceraldehyde-3-phosphate dehydrogenase or condensing enzyme.
Since glutathione was present in the above experiments, two alternative pathways may be postulated for the formation of acetyl CoA: (1) The acetyl group of acetyl phosphate may be transferred directly to CoA. (2) The dehydrogenase may first catalyze the formation of acetylglutathione, as will
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

J. HARTING AND S. F. VELICK 873
be described in the next section, and the acetyl group could then be trans- ferred non-enzymatically to CoA (20). The intermediate formation of free acetylglutathione appears unlikely, however, from the data of Table V, Re- action System b. In this system the reaction proceeds in the presence of highly purified CoA (Test VIII) without added glutathione.
An active acetyl group for the acetylation of CoA may also be produced from the oxidation of acetaldehyde by the dehydrogenase in the absence of any detectable inorganic phosphate. The reaction was carried out in a coupled enzyme system with the crystalline condensing enzyme as pre- viously described. The dehydrogenase was repeatedly crystallized to re- move inorganic phosphate and was maintained in the active form with 0.001 M Versene. The results shown in Table VI indicate that the acetyl
TABLE VI
Synthesis of S-Acetyl CoA with Acetyl Groups Formed in Oxidation of Acetaldehyde The same concentrations of potassium bicarbonate, MgCl,, and oxalacetate
were used as those given in Table V. 0.1 pM of glyceraldehyde-3-phosphate dehydro- genase (preparation recrystallized in 0.001 M Versene); 66 units of condensing en- zyme dialyzed free of phosphate; acetaldehyde, 178 PM; DPN, 2.7 PM; CoA as indi- cated; total volume 2.0 ml. Temperature 29”. Incubation 1 hour.
Components
No CoA .................... Complete ..................
“ ..................
I CoA
Activity
units ger 2 ml.
170 300
Units added
None 500 300
Citrate formed in 1 hr.
PM
0.00
0.63 0.28
group formed in the oxidation of acetaldehyde may be transferred directly to CoA without the intermediary formation of acetyl phosphate.
The above reaction has been shown to be reversible. When acetyl CoA is incubated with DPNH and glyceraldehyde dehydrogenase, a slow oxidation of DPNH can be observed as measured by the decrease in optical density at 340 mp. The acetaldehyde produced in the reaction has been identified with alcohol dehydrogenase.
Acetylation of Glufathione and Cysteine-It has been found that glyceral- dehyde&phosphate dehydrogenase catalyzes a transfer of acetyl groups from acetyl phosphate to glutathione to form X-acetylglutathione. In the experiments shown in Table VII acetyl phosphate, glutathione, and crystal- line dehydrogenase were incubated together for 1 hour. The product, X- acetylglutathione, could be quantitatively determined, since it gives a hy- droxamic acid test and can be distinguished from acetyl phosphate by its heat stability (20). At the end of the incubation, therefore, the reaction
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

874 TRANSFER RErZCTIONS OF ACETYL PHOSPHATE
mixture was heated to 100” for 9 minutes at pH 4.0 in order to hydrolyze any residual acetyl phosphate. The formation of the heat-stable thiol ester was then measured by the hydroxamic acid procedure (14). It can be seen (Table VII) that the acetylglutathione formed was roughly proportional to the glutathione concentration. In the absence of the enzyme or acetyl phosphate no thiol ester was formed. The total hydroxamic acid, which is a measure of both acetylglutathione and acetyl phosphate, remained essen- tially unchanged in the reaction mixtures during the course of incubation.
The X-acetylglutathione formed in the reaction was identified by paper chromatography in a phenol-water-isopropyl alcohol solvent (21). The
TABLE VII
Acetylation of Glutathione by Acetyl Phosphate
The complete system contained 66 PM of sodium Verona], pH 8.0, 8 MM of acetyl phosphate, 0.047 PM of glyceraldehyde-3.phosphate dehydrogenase, and glutathione, neutralized to pH 7.0, in various concentrations as indicated. Total volume 4.0 ml. After incubation for 50 minutes an aliquot was analyzed for total hydroxamic acid by the method of Lipmann and Tuttle. Another aliquot was adjusted to pH 4.0 with acetic acid, heated for 9 minutes at loo”, and then analyzed for heat-stable hydroxamic acid.
Components
Complete...................... “ “
No enzyme..
Glutathione
@f
80 40
4 80
Hydroxamic acid analyses
Total acetyl Heat-stable acetyl
PM PM
6.9 1.5 6.7 1.1 6.6 0.1 7.8 0.0
bands were located by spraying with ninhydrin. In addition to the bands for oxidized and reduced glutathione (RF 0.14 and 0.37, respectively) a third rapidly moving band appeared with an RF of 0.46. The fast moving band did not give a nitroprusside reaction when tested with Reagent I of Toennies and Kolb (21). After treatment with an alcoholic solution of sodium hy- droxide, which hydrolyzes the X-acetyl bond, the nitroprusside reaction be- came positive.2 The formation of X-acetylglutathione by the dehydro- genase was found also by Racker and Krimsky (15). Considerably more thiol ester was synthesized in their system, presumably because of the higher concentrations of enzyme and substrate and a longer incubation at a higher temperature.
Presumptive evidence has been obtained that glyceraldehyde dehydro-
2 We are indebted to Dr. Earl Stadtman for his suggestions for modifications in the chromatographic methods.
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

J. HARTING AND S. F. VELICK 875
genase catalyzes the acetylation of cysteine. The product of the reaction, S-acetylcysteine, is unstable because an S-acetyl bond is formed adjacent to a free amino group and decomposes rapidly and spontaneously (20). Since no analytical test has been devised as yet for the decomposition product, the reaction was followed by the disappearance of acctyl phosphate from the re- action mixture. Table VIII shows that the rate of disappearance of acetyl phosphate in a system containing cysteine and the dehydrogenase is a func- tion of the cysteine concentration between 0.00025 and 0.01 M. Cysteine alone has no effect on the stability of acetyl phosphate. On the other hand, the enzyme without cysteine catalyzes a very rapid disappearance of sub-
TABLE VIII
Disappearance of Acetyl Phosphate, As Function oj Cysteine Concentration, Catalyzed by Glyceraldehyde-d-phosphate Dehydrogenase
2.0 ml. of the complete reaction mixture contained 40 FM of sodium Verona1 buffer, p1-I 8.0, 4 PM of acetyl phosphate, 0.039 pM of glgceraldehyde phosphate dehydro- genase, and varying concentrations of cysteine. Incubation time 25 minutes. The disappearance of acetyl phosphate was determined as a decrease in the hydroxamic acid derivative (14).
Components
No enzyme. . I‘ “
Complete. “ “ ‘< . “
, --
-
Cyst&e concentration
PM 20.0
2.0 0.5 2.0 6.0
20.0 0.0
c -
-
Acetyl phosphate lisappearing in 25 min.
FM 0.1 0.1 0.4 0.7 1.0 1.5 2.0
strate. This is due to an acetyl phosphatase activity of the enzyme which will be reported in detail in a subsequent publication. The dehydrogenase can only function as a phosphatase when it is in the “oxidized” st*ate, that is, in the absence of reducing agents such as cysteine or glutathione. As indicated in Table VIII, the phosphatase activity is strongly inhibited by low concentrations of cysteinc, and, as the concentration is increased, cys- teine itself becomes acetylated.
DISCUSSION
The transacetylase activity of the dehydrogenase differs from that of the purified bacterial phosphotransacetylase (22) in several respects. (a) The mammalian enzyme does not require Coh for the catalyses of phosphate ex- change, arsenolysis, or acetylation of glutathione; however, CoA is a nec- essary cofactor for the bacterial enzyme. (b) The CoA requirement in the
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

876 TRANSFER REACTIONS OF ACETYL PHOSPHATE!
coupled systems for acetylation of sulfanilamide and citrate formation is greatly increased when acetyl CoA is generated with the dehydrogenase in- stead of phosphotransacetylase. (c) Acetaldehyde can serve as a substrate for the dehydrogenase, but not for the phosphotransacetylase, in the for- mation of active acetyl groups. (d) With both the yeast and mammalian dehydrogenases bound DPN is required for the transfer reactions with acetyl phosphate. Such a requirement for DPN has not been shown and would not be expected for a phosphotransacetylase.
The differences between the two enzymes exclude the possibility that the transfer reactions of the glyceraldehyde dehydrogenase are due to bacterial contamination. It was also considered that the transfer activity of the de- hydrogenase might be accounted for by an enzymatic impurity from the muscle itself. However, this possibility is rather remote, since the specific activity for both the oxidation of acetaldehyde and the formation of acetyl CoA is essentially unchanged after three and eight recrystallizations. Moreover, the crystalline dehydrogenase from yeast also catalyzes the P32 exchange and formation of acetyl CoA.
Animal tissues and yeast had not previously been found to form acetyl phosphate or to utilize acetyl phosphate for the production of acetyl CoA. There is, as yet, no evidence that the transfer reactions of the glyceralde- hyde dehydrogenase constitute steps in the major metabolic pathways.
The oxidative phosphorylation of acetaldehyde and the various acetyl transfer reactions catalyzed by the dehydrogenase may be formulated as occurring in the accompanying reaction scheme with an acetyl-enzyme com- pound as the common intermediate.
Step I Step II
Acetaldehyde DPN
--- acetyl-enzyme &PO4
DPNH ---- acetyl phosphate
-7 Lr, Hz0 &As01 fglutathione ztCoA
I 1 1 1 acetate acetyl As04 acetylglutathione acetyl CoA
1 acetate
As required by this mechanism acetaldehyde has been shown to be oxi- dized in the absence of added phosphate. The acetyl group formed in this type of oxidation may be transferred to water, arsenate, glutathione, or CoA. Moreover, the acetyl group of acetyl phosphate can be transferred to the various acceptors without the occurrence of the oxidation-reduction reaction. The separation of Steps I and II follows from the difference in their rates and from their different pH dependence.
We have found, in accord with the scheme, that inorganic phosphate la-
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

J. HARTING AND S. F. VELICK 877
beled with Pz2 exchanges with the phosphate of adenosinetriphosphate in the presence of Bucher’s 3-phosphoglycerokinase (23), the dehydrogenase, and 3-phosphoglyceric acid.3 When arsenate and the dehydrogenase are added to the Bucher enzyme system, a decomposition of adenosinetriphos- phate is observed (24). These observations are explained by a phosphorol- ysis or an arsenolysis of a 3-phosphoglyceryl-enzyme intermediate. The 3-phosphoglyceryl-enzyme compound is sufficiently stable to permit the di- rect observation of its formation and phosphorolysis, corresponding to Steps I and II, and the measurement of the respective equilibria (2526).
SUMMARY
1. Glyceraldehyde-3-phosphate dehydrogenase has been found to catalyze the transfer of acetyl groups of acetyl phosphate to various acceptors: inorganic phosphate, arsenate, and the thiol groups of CoA and glutnthione.
2. The dehydrogenase requires the presence of bound DPN for the trans- ferase activity, but the DPN does not function as a hydrogen donor or ac- ceptor in the transfer reactions.
3. The oxidative phosphorylation of acetaldehyde and the acetyl transfer reactions are formulated as occurring through an acetyl-enzyme intermedi- ate.
We wish to acknowledge the many helpful suggestions of Dr. Robert Crane and of Dr. Carl F. Cori.
BIBLIOGRAPHY
1. Harting, J., Federation Proc., 10, 195 (1951). 2. Harting, J., and Velick, S. F., J. Biol. Chem., 207, 857 (1954). 3. Nygaard, A. P., and Sumner, J. B., Arch. Biochem. and Biophys., 39, 119 (1952). 4. Harting, J., and Velick, S., Federation Proc., 11, 226 (1952). 5. Stadtman, E. R., Novelli, G. D., and Lipmann, F., J. Biol. Chem., 191,365 (1951). 6. Korkes, S., Stern, J. S., Gunsalus, I. C., and Ochoa, S., Nature, 166, 493 (1950). 7. Chou, T. C., Novelli, G. D., Stadtman, E. R., and Lipmann, F., Federation Proc.,
9, 160 (1950). 8. Ochoa, S., Stern, J. R., and Schneider, M. C., J. Biol. Chem., 193, 691 (1951). 9. Kaplan, N. O., and Lipmann, F., J. Biol. Chem., 174, 37 (1948).
10. Stadtman, E. R., and Lipmann, F., J. Biol. Chem., 166, 549 (1950). 11. Krampitz, L. G., and Werkman, C. H., Biochem. J., 35, 595 (1941). 12. Taylor, J. F., Velick, S. F., Cori, G. T., Cori, C. F., and Slein, M. W., J. Biol.
Chem., 173, 619 (1948). 13. Velick, S. F., Hayes, J. E., Jr., and Harting, J., J. Biol. Chem., 203, 527 (1953). 14. Lipmann, F., and Tuttle, L. C., J. BioZ. Chem., 169, 21 (1945). 15. Racker, E., and Krimsky, I., Nature, 169, 1043 (1952). 16. Stadtman, E. R., and Barker, H. A., J. BioZ. Chem., 184, 769 (1950). 17. Lipmann, F., Harvey Lectures, 74, 99 (1948-49). 18. Bratton, A. C., and Marshall, E. K., Jr., J. BioZ. Chem., 128, 537 (1939).
3 Unpublished experiments carried out with Dr. Mildred Cohn.
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

878 TRANSFER REACTIONS OF ACETYL PHOSPHATE
19. Natelson, S., Pincus, J. B., and Lugovoy, J. K., J. Biol. Chem., 176, 745 (1948). 20. Stadtman, E. R., J. Biol. Chem., 196, 535 (1952). 21. Toennies, G., and Kolb, J., Anal. Chem., 23, 823 (1951). 22. Stadtman, E. R., J. Biol. Chewt., 196, 527 (1952). 23. Bucher, T., Biochinz. et biophys. acta, 1, 292 (1947). 24. Krimsky, I., and Racker, E., J. Biol. Chem., 198, 721 (1952). 25. Velick, S. F., and Hayes, J. E., Jr., J. Biol. Chem., 203, 545 (1953). 26. Segal, H. L., and Boyer, P,. D., J. Biol. Chem., 204, 265 (1953).
by guest on July 13, 2018http://w
ww
.jbc.org/D
ownloaded from

Jane Harting and Sidney F. VelickDEHYDROGENASE
GLYCERALDEHYDE-3-PHOSPHATEPHOSPHATE CATALYZED BY
TRANSFER REACTIONS OF ACETYL
1954, 207:867-878.J. Biol. Chem.
http://www.jbc.org/content/207/2/867.citation
Access the most updated version of this article at
Alerts:
When a correction for this article is posted•
When this article is cited•
alerts to choose from all of JBC's e-mailClick here
tml#ref-list-1
http://www.jbc.org/content/207/2/867.citation.full.haccessed free atThis article cites 0 references, 0 of which can be by guest on July 13, 2018
http://ww
w.jbc.org/
Dow
nloaded from