this month in gastroenterology
TRANSCRIPT

This Month in GastroenterologyBy Eugene B. Chang
Figure 1. NOD2 protein is expressed by Paneth cells in smallintestinal crypts. (A) CD-affected terminal ileum; positive immu-nohistochemical staining of NOD2 protein is seen in the basaland apical cytoplasm of Paneth cells (thick arrow) and in amonocyte in the lamina propria (thin arrow). (B–D) Positive, butweaker immunohistochemical staining in Paneth cells (aster-isks) of histologically (B) normal terminal ileal, (C) jejunal, and(D) duodenal tissue.
Paneth Cells Abundantly Express NOD2, a Genein Which Mutations Have Been Associated Withthe Development of Crohn’s DiseaseSeveral years ago, mutations in the NOD2/CARD15 gene were foundto be associated with increased susceptibility of developing ilealCrohn’s disease in certain patient populations. NOD2 is located onchromosome 16 and encodes a protein that contains N-terminalcaspase recruitment domain (CARD) motifs, a nucleotide-bindingdomain (NBD), and a carboxy-terminal region of leucine-rich repeats(LRR). While its functions are still unclear, NOD2 appears to beactivated by muramyl dipeptide, the minimal bioactive component ofbacterial proteoglycan. Because NOD2 is highly expressed in innateimmune cells such as monocytes and macrophages, it has beenproposed as an important sensor molecule of the innate immunesystem. NOD2 may also participate in apoptosis through interactionwith proteins that can activate caspases. The study by Lala et al. nowreports that NOD2 is abundantly expressed in Paneth cells of theterminal ileum and in Crohn’s disease–affected intestinal tissue (Fig-ure 1), where TNF-� is also strongly expressed. NOD2 was alsoreadily detected in monocytes, but not in mature macrophages in thelamina propria or within granulomas, and levels declined as mono-cytes differentiated into macrophages in vitro. These findings areinteresting and may have clinical relevance. Paneth cells are special-ized intestinal epithelial cells adjacent to the stem cell zone in thebase of the crypts of Lieberkuhn in the small intestine (most abun-dantly found in the terminal ileum). In contrast, they are not found innormal colon and rectum, except in inflammatory diseases of thebowel. Paneth cells have an important role in host defense, particu-larly against enteric infection. They produce proinflammatory medi-ators such as TNF-� and antimicrobial defensins. Thus, expression ofmutated NOD2 genes in Paneth cells may have a critical role in thepathogenesis of Crohn’s ileitis, perhaps in inducing or sustaining anaberrant host response to products derived from enteric flora.
See page 47
Differences Among Enteric Flora Species: Some Capableof Inducing Whereas Other Inhibiting ColitisNormal intestinal microflora play a critical physiological role in providing benefitand protection to the host and include organisms such Lactobacillus spp. andBifidobacterium spp. Several mechanisms have been proposed for their actions,including competition for adhesion sites with enteropathogenic bacteria, elabo-ration of antimicrobial compounds, and induction of intestinal mucosal hostprocesses that deter action or colonization by pathogenic organisms. The devel-opment and use of probiotic agents, i.e., microorganisms that have health benefitsbeyond their intrinsic nutritive value, have largely stemmed from these observa-tions. For example, Escherichia coli strain Nissle 1917 has been shown to beeffective in maintaining remission in ulcerative colitis, although the mechanismsof its action are unknown. However, the presence of enteric flora also appear tobe essential for the development of experimental colitis and probably play a rolein Crohn’s disease, suggesting certain species are capable of inciting an aberranthost response. The study by Waidmann et al. was aimed at investigating thecolitis-inducing potential of various bacterial strains indigenous to the
GASTROENTEROLOGY 2003;125:1–3
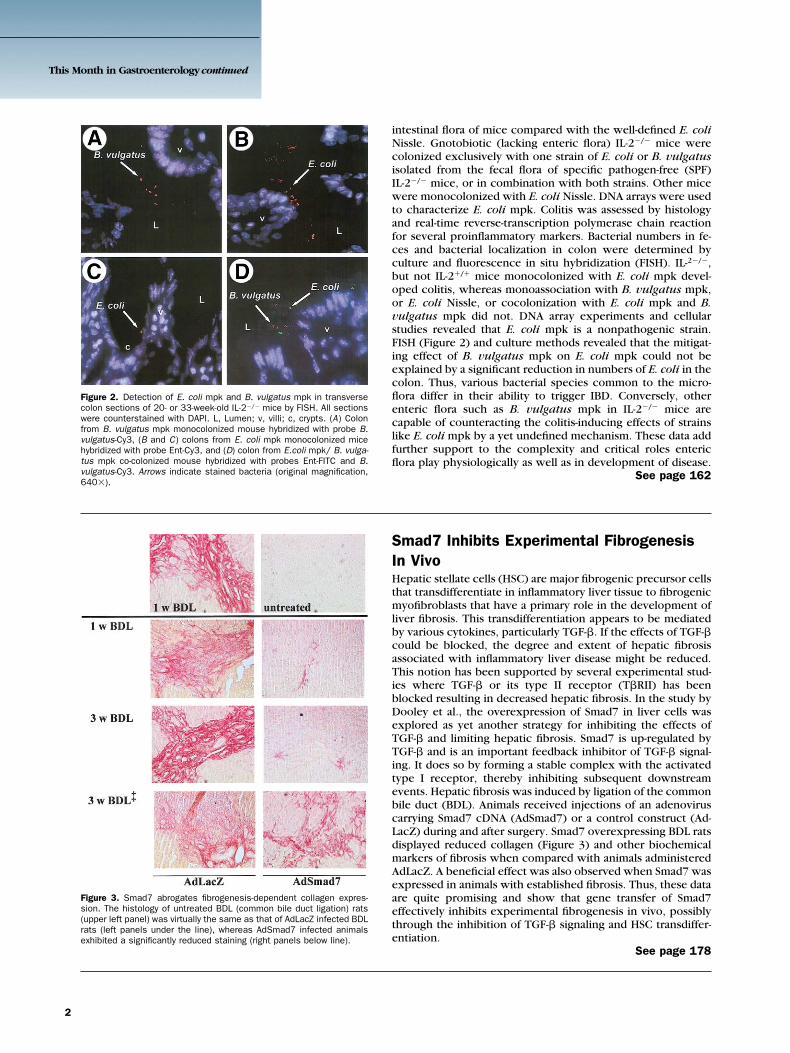
Figure 2. Detection of E. coli mpk and B. vulgatus mpk in transversecolon sections of 20- or 33-week-old IL-2�/� mice by FISH. All sectionswere counterstained with DAPI. L, Lumen; v, villi; c, crypts. (A) Colonfrom B. vulgatus mpk monocolonized mouse hybridized with probe B.vulgatus-Cy3, (B and C) colons from E. coli mpk monocolonized micehybridized with probe Ent-Cy3, and (D) colon from E.coli mpk/ B. vulga-tus mpk co-colonized mouse hybridized with probes Ent-FITC and B.vulgatus-Cy3. Arrows indicate stained bacteria (original magnification,640�).
intestinal flora of mice compared with the well-defined E. coliNissle. Gnotobiotic (lacking enteric flora) IL-2�/� mice werecolonized exclusively with one strain of E. coli or B. vulgatusisolated from the fecal flora of specific pathogen-free (SPF)IL-2�/� mice, or in combination with both strains. Other micewere monocolonized with E. coli Nissle. DNA arrays were usedto characterize E. coli mpk. Colitis was assessed by histologyand real-time reverse-transcription polymerase chain reactionfor several proinflammatory markers. Bacterial numbers in fe-ces and bacterial localization in colon were determined byculture and fluorescence in situ hybridization (FISH). IL-2�/�,but not IL-2�/� mice monocolonized with E. coli mpk devel-oped colitis, whereas monoassociation with B. vulgatus mpk,or E. coli Nissle, or cocolonization with E. coli mpk and B.vulgatus mpk did not. DNA array experiments and cellularstudies revealed that E. coli mpk is a nonpathogenic strain.FISH (Figure 2) and culture methods revealed that the mitigat-ing effect of B. vulgatus mpk on E. coli mpk could not beexplained by a significant reduction in numbers of E. coli in thecolon. Thus, various bacterial species common to the micro-flora differ in their ability to trigger IBD. Conversely, otherenteric flora such as B. vulgatus mpk in IL-2�/� mice arecapable of counteracting the colitis-inducing effects of strainslike E. coli mpk by a yet undefined mechanism. These data addfurther support to the complexity and critical roles entericflora play physiologically as well as in development of disease.
See page 162
Figure 3. Smad7 abrogates fibrogenesis-dependent collagen expres-sion. The histology of untreated BDL (common bile duct ligation) rats(upper left panel) was virtually the same as that of AdLacZ infected BDLrats (left panels under the line), whereas AdSmad7 infected animalsexhibited a significantly reduced staining (right panels below line).
Smad7 Inhibits Experimental FibrogenesisIn VivoHepatic stellate cells (HSC) are major fibrogenic precursor cellsthat transdifferentiate in inflammatory liver tissue to fibrogenicmyofibroblasts that have a primary role in the development ofliver fibrosis. This transdifferentiation appears to be mediatedby various cytokines, particularly TGF-�. If the effects of TGF-�could be blocked, the degree and extent of hepatic fibrosisassociated with inflammatory liver disease might be reduced.This notion has been supported by several experimental stud-ies where TGF-� or its type II receptor (T�RII) has beenblocked resulting in decreased hepatic fibrosis. In the study byDooley et al., the overexpression of Smad7 in liver cells wasexplored as yet another strategy for inhibiting the effects ofTGF-� and limiting hepatic fibrosis. Smad7 is up-regulated byTGF-� and is an important feedback inhibitor of TGF-� signal-ing. It does so by forming a stable complex with the activatedtype I receptor, thereby inhibiting subsequent downstreamevents. Hepatic fibrosis was induced by ligation of the commonbile duct (BDL). Animals received injections of an adenoviruscarrying Smad7 cDNA (AdSmad7) or a control construct (Ad-LacZ) during and after surgery. Smad7 overexpressing BDL ratsdisplayed reduced collagen (Figure 3) and other biochemicalmarkers of fibrosis when compared with animals administeredAdLacZ. A beneficial effect was also observed when Smad7 wasexpressed in animals with established fibrosis. Thus, these dataare quite promising and show that gene transfer of Smad7effectively inhibits experimental fibrogenesis in vivo, possiblythrough the inhibition of TGF-� signaling and HSC transdiffer-entiation.
See page 178
This Month in Gastroenterology continued
2

Figure 4. TUNEL staining, a measure of apoptosis, in liversections 6 hours after challenge with Con A in mice pretreatedwith CT-1 or saline. Decreased fluorescence in CT-1–treatedlivers (right panel) indicates increased survival.
Cardiotrophin-1 is Up-Regulated in RegeneratingLiver and Is CytoprotectiveCardiotrophin-1 (CT-1) is a member of the interleukin-6 (IL-6) familyof cytokines, which is able to support survival and proliferation ofimmature cardiomyocytes and developing motoneurons and to pro-tect cardiac cells against thermal and ischemic insults. Its actions arebelieved to be mediated by several signaling pathways including Jak(Janus activated kinase)/Stat (signal transducer and activator of tran-scription)-3 pathway, Erk 1/2 (extracellular regulated kinases), andPI3K (phosphatidylinositol 3-OH kinase)/Akt pathway. CT-1 also in-duces acute phase protein gene expression in human and rat hepa-toma cells and in primary rat hepatocytes and increases the size of theliver when administered chronically to mice. Thus, agents like CT-1may be useful in treating acute liver failure where effective medicaltherapy is lacking and which has been associated with poor survivalwhen emergency liver transplantation is not feasible. In the study byBustos et al., the expression of CT-1 by liver cells and its possiblehepatoprotective properties are examined. In animals subjected tosubtotal hepatectomy, CT-1 is up-regulated during liver regenerationand has potent antiapoptotic effects. Moreover, hepatocytes culturedunder serum starvation or stimulated with the pro-apoptotic cytokineTGF-� produce CT-1, which serves as a survival factor through anautocrine/paracrine manner. In addition, adenoviral gene delivery ofCT-1 to the liver resulted in significantly reduced mortality in ratsdeveloping fulminant liver failure following subtotal hepatectomy(91% vs. 54% in control and treated animals, respectively). There wasan associated decrease in caspase-3 activity and activation of theantiapoptotic signaling cascades Stat-3, Erk1/2, and Akt in the rem-nant liver. Gene transfer of or intravenously administered recombi-nant CT-1 to the liver also reduced Con-A liver injury and activatedantiapoptotic pathways in the hepatic tissue (Figure 4). Thus, CT-1appears to have promise as a hepatocyte survival factor, whichefficiently reduces hepatocellular damage in animal models of acuteliver injury. Whether these actions hold true clinically remains to bedetermined.
See page 192
This Month in Gastroenterology continued
3