the recovery of 5-ht immunoreactivity in lumbosacral spinal cord and locomotor function after...
TRANSCRIPT

The Recovery of 5-HT Immunoreactivity in Lumbosacral Spinal Cordand Locomotor Function after Thoracic Hemisection
YASUO SARUHASHI,1 WISE YOUNG,2 AND RICHARD PERKINSDepartment of Neurosurgery, New York University Medical Center, 550 First Avenue, New York, New York 10016
To determine the role of serotonin (5-HT) in recoveryfrom spinal cord injury, we examined spinal cord 5-HTimmunohistologically and assessed locomotor recov-ery after thoracic (T8) spinal cord hemisection in 68rats. Forty eight rats had laminectomy and hemisec-tion, while the remaining 20 rats received laminec-tomy only.All rats were evaluated every other day for 4weeks, using a 0–14 point scale openfield test. Hemisec-tion markedly reduced mean hindlimbs scores from 14to 1.5 6 0.32 and 5.6 6 0.31 (mean 6 standard error ofmean) in the ipsilateral and contralateral side, respec-tively. The rats all recovered apparently normal walk-ing by 4 weeks. The 5-HT immunohistological studyrevealed amarked reduction of 5-HT-containing termi-nals in the ipsilateral but not the contralateral lumbo-sacral cord by 1 week after hemisection. By 4 weeksafter hemisection, 5-HT immunoreactive fibers andterminals returned to the ipsilateral lumbosacral cord,with many 5-HT fibers crossing over the central canalat thoracic level. We estimated the recovery of 5-HTneural elements in lumbosacral ventral horn by rank-ing 5-HT staining intensity and counting 5-HT termi-nals. The return of 5-HT immunoreactivity of the lum-bosacral ventral horn correlated with locomotorrecovery. Locomotory recovery invariably occurredwhen the density of 5-HT terminals approached 20% ofcontrol values. These results indicate that return of5-HT fibers and terminals predict the time course andextent of locomotory recovery after thoracic spinalcordhemisection. r 1996 Academic Press, Inc.
INTRODUCTION
Serotonin (5-HT) has unique roles in both acute andchronic spinal cord injury. During the acute phase ofinjury, 5-HT may accelerate secondary damage since itis released at the injury site (38, 54, 55), 5-HT receptors
are present on axons (50), and the 5-HT antagonistmianserin has protective effects in acute spinal cordmodels (41, 43, 44, 46, 47). During the chronic phase ofspinal cord injury, 5-HT may contribute to recoverygiven the strong correlation between locomotor recov-ery and lumbosacral spinal 5-HT in rats after injury(22, 23, 41–47).We recently showed that mianserin (a 5-HT2A and
5-HT2C receptor antagonist) depressed locomotor recov-ery in lateral hemisected rats (51). Hemisection of thethoracic spinal cord eliminated descending spinal fiberson one side of the spinal cord. After hemisection, ratstypically became hemiplegic (29) on the hemisectedside and showed some contralateral weakness as well.The rats, however, recovered normal locomotor scoreson both sides over 2–4 weeks. Because 5-HT fibers inspinal cord originate almost exclusively from supraspi-nal neurons (3) and descend to lumbosacral cord,thoracic cord hemisection should reduce 5-HT elementson one side of the lumbosacral cord.We consequently evaluated 5-HT immunoreactivity
of lumbosacral ventral gray matter and used an openfield test to assess motor function after thoracic spinalcord hemisection.
MATERIALS AND METHODS
Experimental Procedures
We studied a total of 68 male adult Long Evanshooded rats, weighing 464–594 g. The rats were housedin cages under constant environmental conditions (72–78°F and 50–60% humidity) and fed ad libitum. Afteranesthesia with 45 mg/kg i.p. sodium pentobarbital,the thoracic spinal cord was exposed by laminectomy.All surgery was carried out aseptically. In 48 of the rats,the thoracic (T8) spinal cord was hemisected on the leftside; 20 rats had laminectomy but no hemisection. Thespinal cord was hemisected with a scalpel under adissecting microscope. The knife was inserted into thespinal cord with the tip touching midline until the leftside of the cord was completely divided. Hemorrhagewas controlled by gentle pressure with gelfoam pads.Skin was closed with stainless steel surgical clips.
1 Present address: Department of Orthopedic Surgery, Shiga Uni-versity of Medical Science, Otsu, Shiga 520-21, Japan.
2 To whom correspondence should be addressed. Fax: (212) 689-0334.
EXPERIMENTAL NEUROLOGY 139, 203–213 (1996)ARTICLE NO. 0094
203 0014-4886/96 $18.00Copyright r 1996 by Academic Press, Inc.
All rights of reproduction in any form reserved.

Ampicillin (50 mg/kg/day, Marsam Pharmaceuticals)was given daily for 3 days after injury. In addition, thehemisected rats received daily bladder compressions ifnecessary to express urine and reduce the incidence ofbladder infections.
Locomotor Evaluation
To assess locomotor recovery, we scored the ratsevery other day using an open field test (OFT) oflocomotor function (6–8), modified and expanded tohave 0–14 points (51). The original scoring test had ascale of 0 to 5 with some plus and minus categories. Wemodified these scores, listed in Table 1, by expandingplus and minus categories into an ordinal 0 to 14 scale.This scoring system was based on the characteristicsequence of locomotor recovery in rats after spinal cordinjury. The rats were individually taken out of their
boxes and placed on a platform where they wereencouraged to move around and explore. Observationsbegan 5 min after the rats were placed on the platformsand continued for about 10 min per rat. Care was takennot to stimulate the rats for several minutes before theobservations.
Immunohistological Studies
Ten rats from the hemisected group were perfused forimmunohistological studies (31, 58) every week for 4weeks after hemisection. Five control rats receivingonly laminectomy were sacrificed every week afteroperation. The rats were anesthetized with pentobarbi-tal (45 mg/kg i.p.) and perfused through the heart with50 ml of phosphate-buffered saline (0.9% NaCl in 0.1Mphosphate buffer, pH 7.4) to wash out the blood,followed by 300 ml of cold (4°C) solutions containing 4%paraformaldehyde (FA), 0.5% glutaraldehyde, and 0.2%picric acid (PA) in 0.1M phosphate buffer at pH 7.4. Thespinal cords were further fixed by immersion for 2 daysin solutions containing 4% FA and 0.2% PA in 0.1 Mphosphate buffer at 4°C. The specimens were thensoaked in 15% sucrose solutions for 2 weeks, mountedon cardboard, frozen at 215°C, and cut into threepieces: T7, T8–13, and L1. The T7 and L1 pieces wereused for coronal sections while the T8–13 pieces werecut into horizontal 40-µm-thick serial sections with acryostat. Several sections were then stainedwith hema-toxylin and eosin (HE).To stain 5-HT, we applied rabbit polyclonal anti-
serotonin antibodies (ICN ImmunoBiologicals), diluted1:2000 to the sections for 72 h at 4°C. The sections werethen rinsed in phosphate-buffered solution (PBS) for 30min and exposed for 3 h to goat antibody against rabbitIgG (Vector, diluted 1:1000) at 4°C. The sections werewashed in PBS for 30 min, incubated for an hour inavidin–biodin–peroxidase solution, and rinsed withPBS again. The immunoprecipitates were stained byapplying 0.02% diaminobenzidine containing 0.3%nickel ammonium sulfate and 0.003% H2O2 dilutedwith 0.05MTris–HCl buffer. The sectionsweremountedon chrome–alum–gelatin-coated glass slides, air-dried,covered with plastic (Crystal Mount, Biomedia Corp.),and coverslipped.The slides were examined with a light microscope
(Axiophot, Zeiss) and representative sections were pho-tographed with special attention paid to three areas.First, at the hemisection site, we determined whetherthe lesion extended up to or across the midline. Second,we examined the horizontal sections for the presence of5-HT fibers that crossed the midline. Third, we exam-ined the lumbosacral spinal cords for the 5-HT termi-nals, comparing ipsilateral and contralateral sides withcontrol spinal cords.
TABLE 1
Open Field Test Scores
OSU NYU
Paralyzed
0 0The rats shows no nonreflexivehindlimb movements
Active hip and/or knee movements12 1 Slight movement at the hip onlya1 2 Active movement at the hip or knee
11 3Active movement at hip and knee,no ankle movement
Active hip, knee, and ankle movement22 4 Weak movement of ankle joint
2 5Active movement in hip, knee, andankle joints
21 6 Attempts at support are seenActive support with stance or uncoordinated gait
32 7 Support in stance only3 8 Active support, uncoordinated gaitb
31 9Occasional bouts of coordinatedgait
Coordination of forelimb and hindlimb in gait42 10 Lack of control at foot and anklec
4 11Coordinated fore- and hindlimbgait, wide support based
41 12Narrowing support base, noabdomen drag, some toe drags
Normal or almost-normal gait
52 13Occasional toe drags, slightunsteadiness on full speed turns
5 14Steady gait with no toe drags, fullspeed turns
a The rat must not have been stimulated for several minutes beforeobservation.
b Coordinated gait refers to coordination between fore- and hind-limb movement.
c Walking on metatarsal phalangeal joints or on medial surface offoot.
d Hindlimbs are spread farther than normal and the rat may dragits abdomen.
204 SARUHASHI, YOUNG, AND PERKINS
As described above, groups of 10 rats were eutha-nized at 1, 2, 3, and 4 weeks. We chose 40 rats thatshowed precise hemisections that included the midlineconfirmed on HE-stained sections of the lesion. Afterinspecting the immunohistochemical stains, we tookblack and white photographs of the ventral gray matterin the lumbosacral spinal cord (L1 spinal level) of allrats. To assess the intensity of 5-HT immunoreactivity,we selected 90 typical pictures of ventral horns. Theseincluded 80 pictures from both sides of the hemisectedgroup and 10 from the left side of the control group. Weassessed the 5-HT immunoreactivity in the ventralhorn by two methods as followed. First, we ranked the90 pictures in accordance to the density and presence ofimmunostained processes. The ipsilateral and contralat-eral sides were separately ranked in the hemisectedrats by two different investigators (YS and WY). Asimilar ranking approach was used by Faden et al. andSharma et al. (23, 53) to assess the density of 5-HT-containing terminal and fibers in the ventral horn. Thenormal rat characteristically showed heavy serotoner-gic staining in the ventral cord, typically coveringmotoneuron surfaces varicose terminals. Hemisectedrats showed reduced distribution of 5-HT fibers andterminals, both in terms of fiber number and intensityof 5-HT immunoreactivity. We ranked all 90 picturesfrom 1st (lowest immunointensity) to 90th (highestimmunointensity). Second, we counted all the 5-HTimmunoreactive terminals in ventral horn of all 90pictures. The surgical history of the rats was maskedduring the ranking and counting.
Data Analysis
All 6 values indicate standard errors of means. Toascertain the statistical significance of 5-HT stainingand OFT score changes, we used analysis of variance
(ANOVA) to compare control and hemisected groups, aswell as linear regression.
RESULTS
Open Field Test
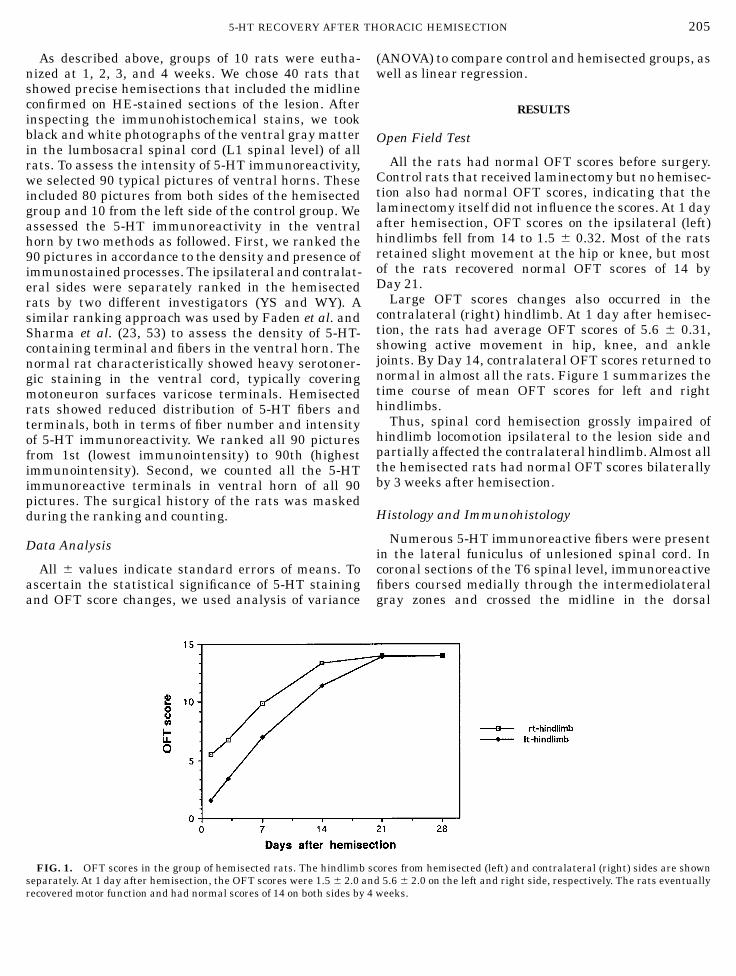
All the rats had normal OFT scores before surgery.Control rats that received laminectomy but no hemisec-tion also had normal OFT scores, indicating that thelaminectomy itself did not influence the scores.At 1 dayafter hemisection, OFT scores on the ipsilateral (left)hindlimbs fell from 14 to 1.5 6 0.32. Most of the ratsretained slight movement at the hip or knee, but mostof the rats recovered normal OFT scores of 14 byDay 21.Large OFT scores changes also occurred in the
contralateral (right) hindlimb. At 1 day after hemisec-tion, the rats had average OFT scores of 5.6 6 0.31,showing active movement in hip, knee, and anklejoints. By Day 14, contralateral OFT scores returned tonormal in almost all the rats. Figure 1 summarizes thetime course of mean OFT scores for left and righthindlimbs.Thus, spinal cord hemisection grossly impaired of
hindlimb locomotion ipsilateral to the lesion side andpartially affected the contralateral hindlimb.Almost allthe hemisected rats had normal OFT scores bilaterallyby 3 weeks after hemisection.
Histology and Immunohistology
Numerous 5-HT immunoreactive fibers were presentin the lateral funiculus of unlesioned spinal cord. Incoronal sections of the T6 spinal level, immunoreactivefibers coursed medially through the intermediolateralgray zones and crossed the midline in the dorsal
FIG. 1. OFT scores in the group of hemisected rats. The hindlimb scores from hemisected (left) and contralateral (right) sides are shownseparately. At 1 day after hemisection, the OFT scores were 1.5 6 2.0 and 5.6 6 2.0 on the left and right side, respectively. The rats eventuallyrecovered motor function and had normal scores of 14 on both sides by 4 weeks.
2055-HT RECOVERY AFTER THORACIC HEMISECTION
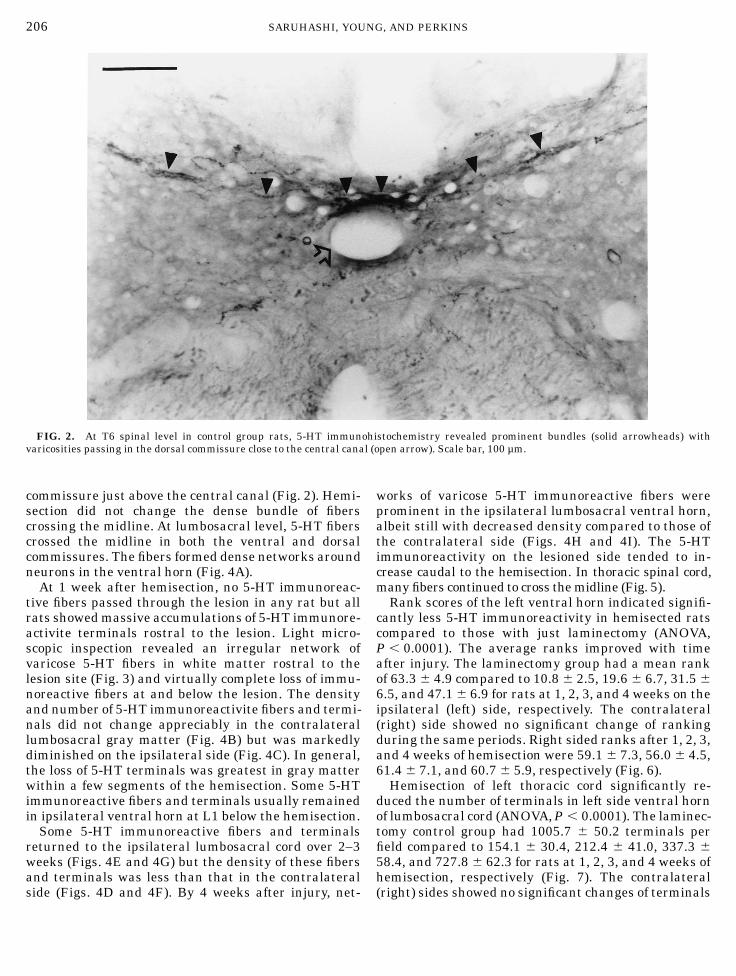
commissure just above the central canal (Fig. 2). Hemi-section did not change the dense bundle of fiberscrossing the midline. At lumbosacral level, 5-HT fiberscrossed the midline in both the ventral and dorsalcommissures. The fibers formed dense networks aroundneurons in the ventral horn (Fig. 4A).At 1 week after hemisection, no 5-HT immunoreac-
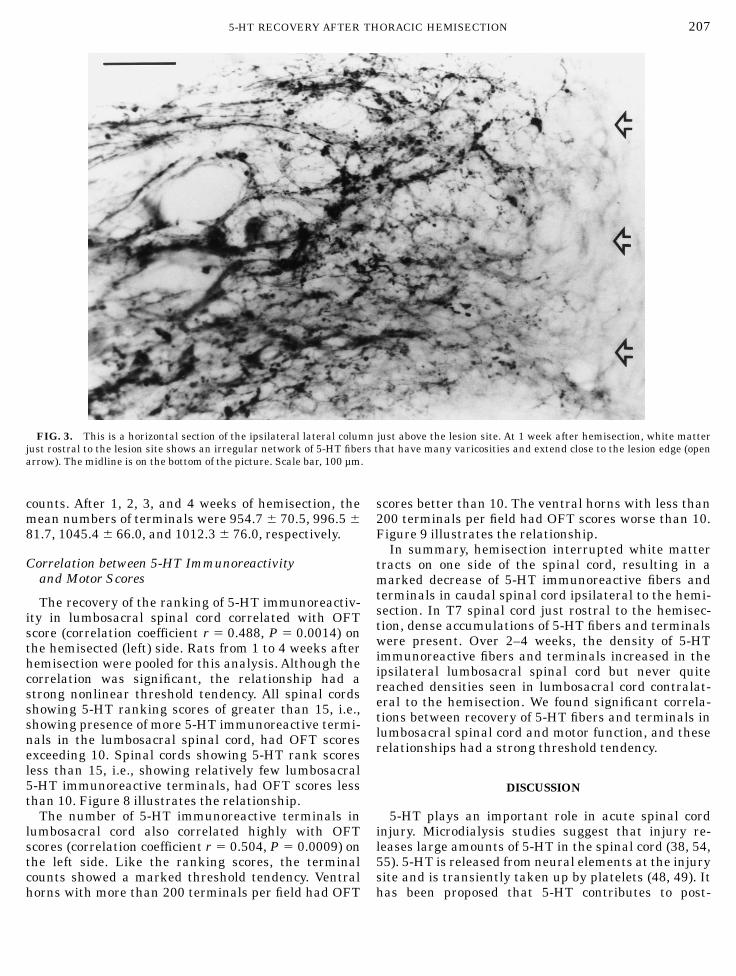
tive fibers passed through the lesion in any rat but allrats showedmassive accumulations of 5-HT immunore-activite terminals rostral to the lesion. Light micro-scopic inspection revealed an irregular network ofvaricose 5-HT fibers in white matter rostral to thelesion site (Fig. 3) and virtually complete loss of immu-noreactive fibers at and below the lesion. The densityand number of 5-HT immunoreactivite fibers and termi-nals did not change appreciably in the contralaterallumbosacral gray matter (Fig. 4B) but was markedlydiminished on the ipsilateral side (Fig. 4C). In general,the loss of 5-HT terminals was greatest in gray matterwithin a few segments of the hemisection. Some 5-HTimmunoreactive fibers and terminals usually remainedin ipsilateral ventral horn at L1 below the hemisection.Some 5-HT immunoreactive fibers and terminals
returned to the ipsilateral lumbosacral cord over 2–3weeks (Figs. 4E and 4G) but the density of these fibersand terminals was less than that in the contralateralside (Figs. 4D and 4F). By 4 weeks after injury, net-
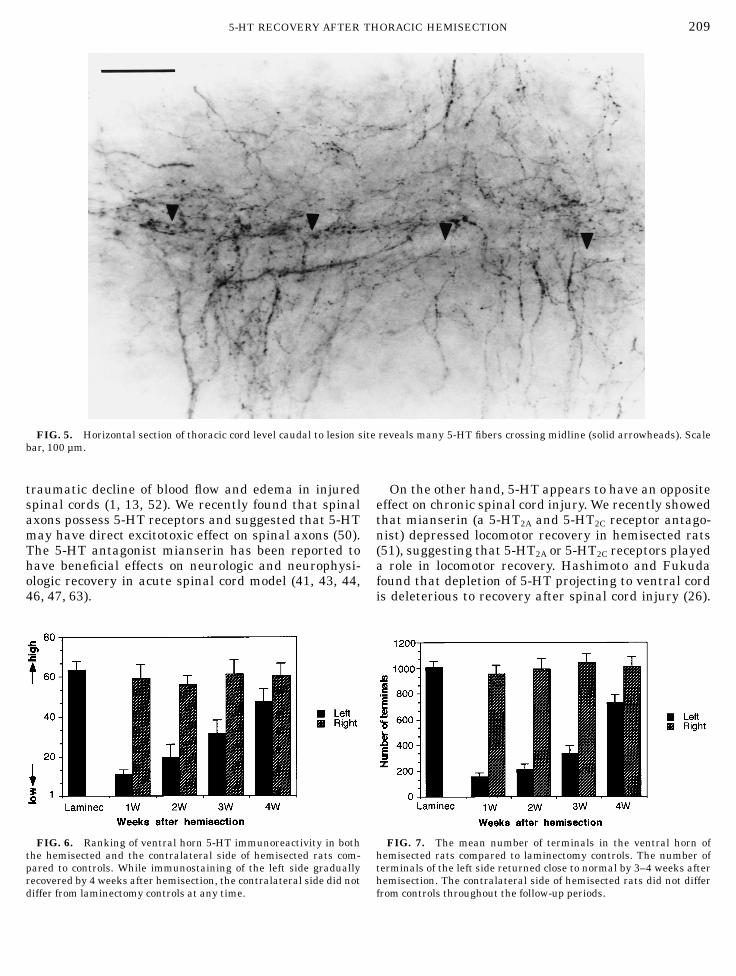
works of varicose 5-HT immunoreactive fibers wereprominent in the ipsilateral lumbosacral ventral horn,albeit still with decreased density compared to those ofthe contralateral side (Figs. 4H and 4I). The 5-HTimmunoreactivity on the lesioned side tended to in-crease caudal to the hemisection. In thoracic spinal cord,many fibers continued to cross themidline (Fig. 5).Rank scores of the left ventral horn indicated signifi-
cantly less 5-HT immunoreactivity in hemisected ratscompared to those with just laminectomy (ANOVA,P , 0.0001). The average ranks improved with timeafter injury. The laminectomy group had a mean rankof 63.3 6 4.9 compared to 10.8 6 2.5, 19.6 6 6.7, 31.5 6
6.5, and 47.1 6 6.9 for rats at 1, 2, 3, and 4 weeks on theipsilateral (left) side, respectively. The contralateral(right) side showed no significant change of rankingduring the same periods. Right sided ranks after 1, 2, 3,and 4 weeks of hemisection were 59.1 6 7.3, 56.0 6 4.5,61.4 6 7.1, and 60.7 6 5.9, respectively (Fig. 6).Hemisection of left thoracic cord significantly re-
duced the number of terminals in left side ventral hornof lumbosacral cord (ANOVA,P , 0.0001). The laminec-tomy control group had 1005.7 6 50.2 terminals perfield compared to 154.1 6 30.4, 212.4 6 41.0, 337.3 6
58.4, and 727.8 6 62.3 for rats at 1, 2, 3, and 4 weeks ofhemisection, respectively (Fig. 7). The contralateral(right) sides showed no significant changes of terminals
FIG. 2. At T6 spinal level in control group rats, 5-HT immunohistochemistry revealed prominent bundles (solid arrowheads) withvaricosities passing in the dorsal commissure close to the central canal (open arrow). Scale bar, 100 µm.
206 SARUHASHI, YOUNG, AND PERKINS
counts. After 1, 2, 3, and 4 weeks of hemisection, themean numbers of terminals were 954.7 6 70.5, 996.5 6
81.7, 1045.4 6 66.0, and 1012.3 6 76.0, respectively.
Correlation between 5-HT Immunoreactivityand Motor Scores
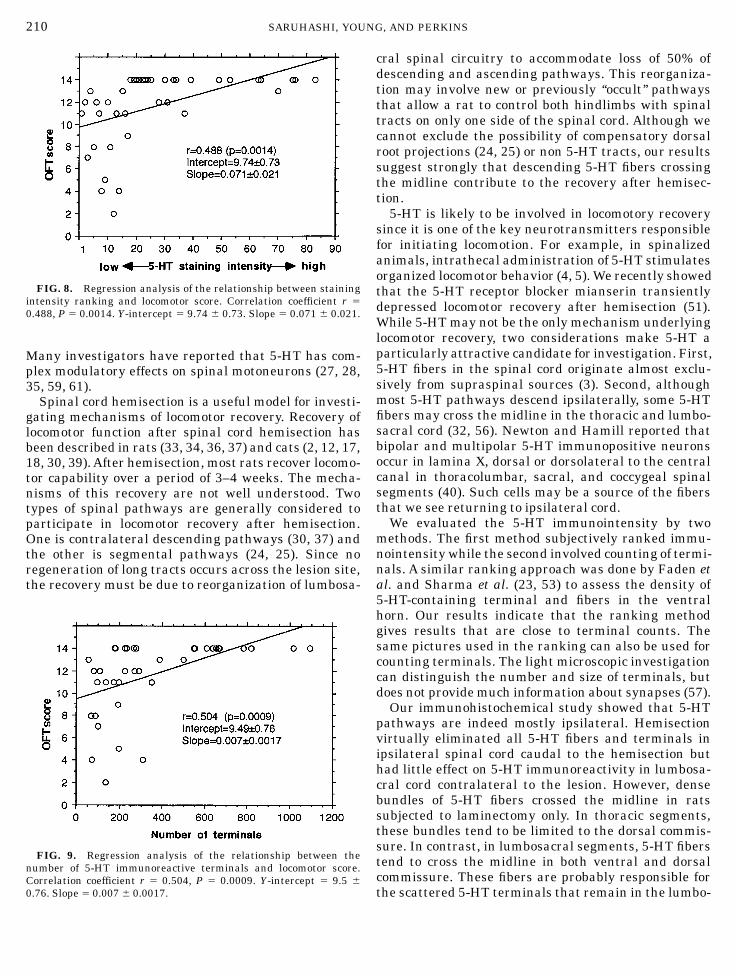
The recovery of the ranking of 5-HT immunoreactiv-ity in lumbosacral spinal cord correlated with OFTscore (correlation coefficient r 5 0.488, P 5 0.0014) onthe hemisected (left) side. Rats from 1 to 4 weeks afterhemisection were pooled for this analysis. Although thecorrelation was significant, the relationship had astrong nonlinear threshold tendency. All spinal cordsshowing 5-HT ranking scores of greater than 15, i.e.,showing presence of more 5-HT immunoreactive termi-nals in the lumbosacral spinal cord, had OFT scoresexceeding 10. Spinal cords showing 5-HT rank scoresless than 15, i.e., showing relatively few lumbosacral5-HT immunoreactive terminals, had OFT scores lessthan 10. Figure 8 illustrates the relationship.The number of 5-HT immunoreactive terminals in
lumbosacral cord also correlated highly with OFTscores (correlation coefficient r 5 0.504, P 5 0.0009) onthe left side. Like the ranking scores, the terminalcounts showed a marked threshold tendency. Ventralhorns with more than 200 terminals per field had OFT
scores better than 10. The ventral horns with less than200 terminals per field had OFT scores worse than 10.Figure 9 illustrates the relationship.In summary, hemisection interrupted white matter
tracts on one side of the spinal cord, resulting in amarked decrease of 5-HT immunoreactive fibers andterminals in caudal spinal cord ipsilateral to the hemi-section. In T7 spinal cord just rostral to the hemisec-tion, dense accumulations of 5-HT fibers and terminalswere present. Over 2–4 weeks, the density of 5-HTimmunoreactive fibers and terminals increased in theipsilateral lumbosacral spinal cord but never quitereached densities seen in lumbosacral cord contralat-eral to the hemisection. We found significant correla-tions between recovery of 5-HT fibers and terminals inlumbosacral spinal cord and motor function, and theserelationships had a strong threshold tendency.
DISCUSSION
5-HT plays an important role in acute spinal cordinjury. Microdialysis studies suggest that injury re-leases large amounts of 5-HT in the spinal cord (38, 54,55). 5-HT is released from neural elements at the injurysite and is transiently taken up by platelets (48, 49). Ithas been proposed that 5-HT contributes to post-
FIG. 3. This is a horizontal section of the ipsilateral lateral column just above the lesion site. At 1 week after hemisection, white matterjust rostral to the lesion site shows an irregular network of 5-HT fibers that have many varicosities and extend close to the lesion edge (openarrow). The midline is on the bottom of the picture. Scale bar, 100 µm.
2075-HT RECOVERY AFTER THORACIC HEMISECTION
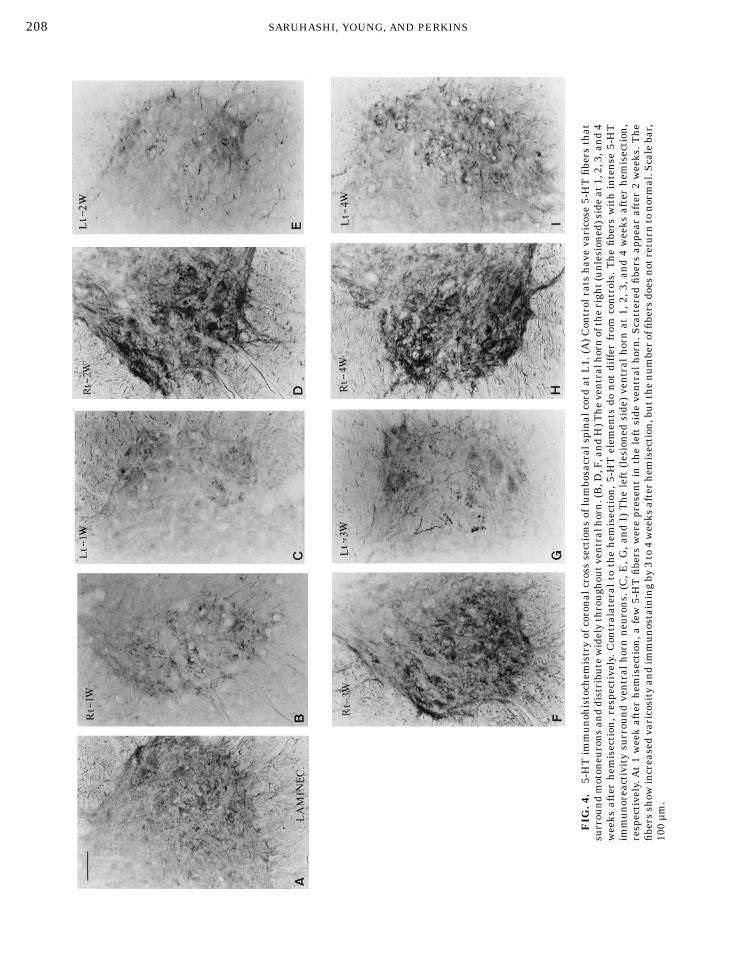
FIG
.4.
5-HTimmunohistochem
istryofcoronalcrosssectionsoflumbosacralspinalcord
atL1.(A)Controlratshavevaricose
5-HTfibersthat
surroundmotoneuronsanddistributewidelythroughoutventralhorn.(B,D
,F,andH)T
heventralhornoftheright(unlesioned)sideat1,2,3,and4
weeks
afterhem
isection,respectively.Contralateral
tothehem
isection,5-HTelem
entsdo
not
differ
from
controls.Thefibers
withintense
5-HT
immunoreactivitysurroundventralhornneurons.(C,E
,G,andI)Theleft(lesioned
side)ventralhornat
1,2,3,and4weeks
afterhem
isection,
respectively.A
t1weekafterhem
isection,a
few5-HTfibers
werepresentin
theleftside
ventralhorn.S
cattered
fibers
appear
after2weeks.T
he
fibersshow
increasedvaricosityandimmunostainingby
3to4weeks
afterhem
isection,butthenumberoffibersdoesnotreturn
tonormal.Scalebar,
100µm.
208 SARUHASHI, YOUNG, AND PERKINS
traumatic decline of blood flow and edema in injuredspinal cords (1, 13, 52). We recently found that spinalaxons possess 5-HT receptors and suggested that 5-HTmay have direct excitotoxic effect on spinal axons (50).The 5-HT antagonist mianserin has been reported tohave beneficial effects on neurologic and neurophysi-ologic recovery in acute spinal cord model (41, 43, 44,46, 47, 63).
On the other hand, 5-HT appears to have an oppositeeffect on chronic spinal cord injury. We recently showedthat mianserin (a 5-HT2A and 5-HT2C receptor antago-nist) depressed locomotor recovery in hemisected rats(51), suggesting that 5-HT2A or 5-HT2C receptors playeda role in locomotor recovery. Hashimoto and Fukudafound that depletion of 5-HT projecting to ventral cordis deleterious to recovery after spinal cord injury (26).
FIG. 5. Horizontal section of thoracic cord level caudal to lesion site reveals many 5-HT fibers crossing midline (solid arrowheads). Scalebar, 100 µm.
FIG. 6. Ranking of ventral horn 5-HT immunoreactivity in boththe hemisected and the contralateral side of hemisected rats com-pared to controls. While immunostaining of the left side graduallyrecovered by 4 weeks after hemisection, the contralateral side did notdiffer from laminectomy controls at any time.
FIG. 7. The mean number of terminals in the ventral horn ofhemisected rats compared to laminectomy controls. The number ofterminals of the left side returned close to normal by 3–4 weeks afterhemisection. The contralateral side of hemisected rats did not differfrom controls throughout the follow-up periods.
2095-HT RECOVERY AFTER THORACIC HEMISECTION
Many investigators have reported that 5-HT has com-plex modulatory effects on spinal motoneurons (27, 28,35, 59, 61).Spinal cord hemisection is a useful model for investi-
gating mechanisms of locomotor recovery. Recovery oflocomotor function after spinal cord hemisection hasbeen described in rats (33, 34, 36, 37) and cats (2, 12, 17,18, 30, 39).After hemisection, most rats recover locomo-tor capability over a period of 3–4 weeks. The mecha-nisms of this recovery are not well understood. Twotypes of spinal pathways are generally considered toparticipate in locomotor recovery after hemisection.One is contralateral descending pathways (30, 37) andthe other is segmental pathways (24, 25). Since noregeneration of long tracts occurs across the lesion site,the recovery must be due to reorganization of lumbosa-
cral spinal circuitry to accommodate loss of 50% ofdescending and ascending pathways. This reorganiza-tion may involve new or previously ‘‘occult’’ pathwaysthat allow a rat to control both hindlimbs with spinaltracts on only one side of the spinal cord. Although wecannot exclude the possibility of compensatory dorsalroot projections (24, 25) or non 5-HT tracts, our resultssuggest strongly that descending 5-HT fibers crossingthe midline contribute to the recovery after hemisec-tion.5-HT is likely to be involved in locomotory recovery
since it is one of the key neurotransmitters responsiblefor initiating locomotion. For example, in spinalizedanimals, intrathecal administration of 5-HT stimulatesorganized locomotor behavior (4, 5).We recently showedthat the 5-HT receptor blocker mianserin transientlydepressed locomotor recovery after hemisection (51).While 5-HTmay not be the only mechanism underlyinglocomotor recovery, two considerations make 5-HT aparticularly attractive candidate for investigation. First,5-HT fibers in the spinal cord originate almost exclu-sively from supraspinal sources (3). Second, althoughmost 5-HT pathways descend ipsilaterally, some 5-HTfibers may cross the midline in the thoracic and lumbo-sacral cord (32, 56). Newton and Hamill reported thatbipolar and multipolar 5-HT immunopositive neuronsoccur in lamina X, dorsal or dorsolateral to the centralcanal in thoracolumbar, sacral, and coccygeal spinalsegments (40). Such cells may be a source of the fibersthat we see returning to ipsilateral cord.We evaluated the 5-HT immunointensity by two
methods. The first method subjectively ranked immu-nointensity while the second involved counting of termi-nals. A similar ranking approach was done by Faden etal. and Sharma et al. (23, 53) to assess the density of5-HT-containing terminal and fibers in the ventralhorn. Our results indicate that the ranking methodgives results that are close to terminal counts. Thesame pictures used in the ranking can also be used forcounting terminals. The light microscopic investigationcan distinguish the number and size of terminals, butdoes not providemuch information about synapses (57).Our immunohistochemical study showed that 5-HT
pathways are indeed mostly ipsilateral. Hemisectionvirtually eliminated all 5-HT fibers and terminals inipsilateral spinal cord caudal to the hemisection buthad little effect on 5-HT immunoreactivity in lumbosa-cral cord contralateral to the lesion. However, densebundles of 5-HT fibers crossed the midline in ratssubjected to laminectomy only. In thoracic segments,these bundles tend to be limited to the dorsal commis-sure. In contrast, in lumbosacral segments, 5-HT fiberstend to cross the midline in both ventral and dorsalcommissure. These fibers are probably responsible forthe scattered 5-HT terminals that remain in the lumbo-
FIG. 8. Regression analysis of the relationship between stainingintensity ranking and locomotor score. Correlation coefficient r 5
0.488, P 5 0.0014. Y-intercept 5 9.74 6 0.73. Slope 5 0.071 6 0.021.
FIG. 9. Regression analysis of the relationship between thenumber of 5-HT immunoreactive terminals and locomotor score.Correlation coefficient r 5 0.504, P 5 0.0009. Y-intercept 5 9.5 6
0.76. Slope 5 0.007 6 0.0017.
210 SARUHASHI, YOUNG, AND PERKINS

sacral cord after ipsilateral hemisection. Since hemisec-tion eliminates descending fibers on one side, the onlysource of 5-HT must be fibers crossing the midline fromthe other side. Our data does not allow us to distinguishbetween regeneration of new fibers crossing the mid-line or sprouting of already existing fibers crossing themidline.Many investigators have reported correlations be-
tween locomotory recovery and lumbosacral spinal5-HT after spinal cord injury (22, 23, 41–47). Ourresults show that OFT scores of the hemisected side fallfrom 14 to 1.5 (slight movement at hip or knee). Over 3weeks, most of the animals recover virtually normalcoordinated locomotion on both sides. The time courseand extent of 5-HT reinnervation predicted open fieldtest score of hindlimb ipsilateral to the lesion. Therelationship was not linear and showed a strong thresh-old tendency.Ventral horns with more than 200 terminals were
associated with OFT scores exceeding 10.An OFT scoreof .10 indicates coordinated gait between forelimb andhindlimb. Ventral horns with less than 200 terminalswere invariably associated with OFT scores less than10, suggesting that 20%, or 200 terminals of 1000counted terminals in controls, can support locomotion.A 20% threshold is similar to the amount of preservedaxons able to support locomotion in spinal-injured catsreported by Blight and Young and Blight and DeCrescito (9–11). Many investigators have reported simi-lar threshold tendency after spinal cord injury (14–21,60, 62).In conclusion, hemisection of the thoracic spinal cord
eliminated most of 5-HT fibers and terminals in theipsilateral lumbosacral spinal cord. 5-HT neural ele-ments returned to the lumbosacral site over 3–4 weeksafter thoracic hemisection. 5-HT fibers clearly crossedover from the contralateral side. Recovery of locomotorscores correlated with the return of 5-HT fibers andterminals in the lumbosacral spinal cord. The relation-ship was nonlinear with a threshold for locomotorrecovery at about 20% of normal 5-HT innervation.These results suggest that 5-HT fibers and terminals atlumbosacral ventral horn play an important role inlocomotory recovery after thoracic cord hemisection.
ACKNOWLEDGMENTS
We are grateful to Mr. BenAyala, Ms. Bo TomNg, and Ms. Mei LanChow for their technical assistance. This study was supported in partby grants from NIH P01-NS10164, R01-NS15590, andAPA.
REFERENCES
1. ABRAHAM, J., A. S. BALASUBRAMANIAN, D. R. THEODORE, S.NAGARAJAN, C. A.APTE, AND S. CHANDI. 1985. Spinal cord edema,5-hydroxytryptamine, lipid peroxidation, and lysosomal enzyme
release after acute contusion and compression injury in pri-mates. Cent. Nerv. Syst. Trauma 2(1): 45–60.
2. ALSTERMARK, B., T. ISA, A. LUNDBERG, L. G. PETTERSSON, AND B.TANTISIRA. 1989. The effect of low pyramidal lesions on forelimbmovements in the cat.Neurosci. Res. 7(1): 71–5.
3. ANDERSON, E. G. 1983. The serotonin system of the spinal cord.Pages 241–274 in R. A. Davidoff, Ed., Handbook of the SpinalCord.Dekker, NewYork.
4. BARBEAU, H., AND S. ROSSIGNOL. 1990. The effects of serotonergicdrugs on the locomotor pattern and on cutaneous reflexes of theadult chronic spinal cat. Brain Res. 514(1): 55–67.
5. BARBEAU, H., AND S. ROSSIGNOL. 1991. Initiation and modulationof the locomotor pattern in the adult chronic spinal cat bynoradrenergic, serotonergic and dopaminergic drugs.Brain Res.546(2): 250–60.
6. BEATTIE, M. S., AND J. C. BRESNAHAN. 1989. Longitudinalassessment of locomotor recovery in rats with spinal cordimpact lesions. Pages 16–25 in M. Brown, M. Golderger, B.Bregman, and C. Vierck, Ed., Criteria for the Assessment ofRecovery of Function: Behavioral Methods. American ParalysisAssociation, Springfield, New Jersey.
7. BEHRMANN, D. L., J. C. BRESNAHAN, AND M. S. BEATTIE. 1992. Acomparison of YM-14673, U-50488H, and nalmefene after spi-nal cord injury in the rat. Exp. Neurol. 119(2): 258–67.
8. BEHRMANN, D. L., J. C. BRESNAHAN, M. S. BEATTIE, AND B. R.SHAH. 1992. Spinal cord injury produced by consistent mechani-cal displacement of the cord in rats: Behavioral and histologicanalysis. J. Neurotrauma 9(3): 197–217.
9. BLIGHT, A., AND W. YOUNG. 1989. Central axons in injured catspinal cord recover electrophysiological function following remy-elination by Schwann cells. J. Neurol. Sci. 91: 15–34.
10. BLIGHT, A., AND W. YOUNG. 1990. Axonal morphometric corre-lates of evoked potentials in experimental spinal cord injury.Pages 87–113 in S. Salzman, Ed., Neural Monitoring. Humana,NewYork.
11. BLIGHT, A. R., AND V. DECRESCITO. 1986. Morphometric analysisof experimental spinal cord injury in the cat: The relation ofinjury intensity to survival of myelinated axons. Neuroscience19(1): 321–341.
12. BREGMAN, B. S., AND M. E. GOLDBERGER. 1982. Anatomicalplasticity and sparing of function after spinal cord damage inneonatal cats. Science 217: 553–555.
13. BRODNER, R. A., G. J. DOHRMAN, R. H. ROTH, AND R. A. RUBIN.1980. Correlation of cerebrospinal fluid serotonin and alteredspinal cord blood flow in experimental trauma. Surg. Neurol. 13:337–343.
14. DIMITRIJEVIC, M. R. 1988. Residual motor functions in spinalcord injury. Pages 139–155 in S. G. Waxman, Ed., FunctionalRecovery in Neurological Disease (Advances in Neurology).Raven, NewYork.
15. DIMITRIJEVIC, M. R., J. FAGANEL, D. LEHMKUHL, AND A. M.SHERWOOD. 1983. Motor control in man after partial or completespinal cord injury. Pages 915–926 in J. E. Desmedt, Ed., MotorControl Mechanisms in Health and Disease. Raven, NewYork.
16. EIDELBERG, E. 1981. Consequences of spinal cord lesions uponmotor function, with special reference to locomotor activity.Prog. Neurobiol. 17(3): 185–202.
17. EIDELBERG, E., L. NGUYEN, AND L. DEZA. 1986. Recovery oflocomotor function after hemisection of the spinal cord in cats.Brain Res. Bull. 16: 507–515.
18. EIDELBERG, E., J. L. STORY, J. G. WALDEN, AND B. L. MEYER. 1981.Anatomical correlates of return of locomotor function afterpartial spinal cord lesions in cats. Exp. Brain Res. 42(1): 81–88.
2115-HT RECOVERY AFTER THORACIC HEMISECTION

19. EIDELBERG, E., D. STRAEHLEY, AND R. ERSPAMER. 1977. Relation-ship between residual hindlimb assisted locomotion and surviv-ing axons after incomplete spinal cord injuries. Exp. Neurol. 56:312–322.
20. EIDELBERG, E., AND J. YU. 1981. Effects of corticospinal lesionsupon treadmill locomotion in cats. Exp. Brain Res. 43: 101–103.
21. EIDELBERG, E., AND J. YU. 1981. Effects of vestibulospinal lesionsupon locomotor function in cats. Brain Res. 22: 179–183.
22. FADEN, A. I., J. A. ELLISON, AND L. J. NOBLE. 1990. Effects ofcompetitive and non-competitive NMDAreceptor antagonists inspinal cord injury. Eur. J. Pharmacol. 175(2): 165–174.
23. FADEN, A. I., A. GANNON, AND A. I. BASBAUM. 1988. Use ofserotonin immunocytochemistry as a marker of injury severityafter experimental spinal trauma in rats. Brain Res. 450(1–2):94–100.
24. GOLDBERGER, M. E., AND M. MURRAY. 1978. Recovery of move-ment and axonal sprouting may obey some of the same laws.Pages 73–96 in C. W. Cotman, Ed., Neuronal Plasticity. Aca-demic Press, NewYork.
25. GOLDBERGER, M. E. 1988. Spared-root deafferentation of a cat’shindlimb: hierarchical regulation of pathways mediating recov-ery of motor behavior. Exp. Brain Res. 73(2): 329–342.
26. HASHIMOTO, T., AND N. FUKUDA. 1991. Contribution of serotoninneurons to the functional recovery after spinal cord injury inrats. Brain Res. 539(2): 263–270.
27. HOUNSGAARD, J., H. HULTBORN, B. JESPERSEN, AND O. KIEHN.1988. Bistability of alpha-motoneurones in the decerebrate catand in the acute spinal cat after intravenous 5-hydroxytrypto-phan. J. Physiol. 405: 345–67.
28. JACKSON, D. A., AND S. R. WHITE. 1990. Receptor subtypesmediating facilitation by serotonin of excitability of spinalmotoneurons.Neuropharmacology 29(9): 787–797.
29. KATO, M. 1985. Recovery of postural control following chronicbilateral hemisections at different spinal cord levels in adultcats. Exp. Neurol. 90: 350–364.
30. KATO, M., S. MURAKAMI, K. YASUDA, AND H. HIRAYAMA. 1984.Disruption of fore- and hindlimb coordination during over-ground locomotion in cats with bilateral serial hemisection ofthe spinal cord.Neurosci. Res. 2: 27–47.
31. KIMURA, H., P. L. MCGEER, J. H. PENG, AND E. G. MCGEER. 1989.The central cholinergic system studied by choline acetyltransfer-ase immunohistochemistry in the cat. J. Comp. Neurol. 200:151–201.
32. KRUKOFF, T. L., J. CIRIELLO, AND F. R. CALARESU. 1985. Segmen-tal distribution of peptide- and 5HT-like immunoreactivity innerve terminals and fibers of the thoracolumbar sympatheticnuclei of the cat. J. Comp. Neurol. 240: 103–116.
33. KUNKEL, B. E., AND B. S. BREGMAN. 1990. Spinal cord trans-plants enhance the recovery of locomotor function after spinalcord injury at birth. Exp. Brain Res. 81(1): 25–34.
34. KUNKEL, B. E., H. N. DAI, AND B. S. BREGMAN. 1992. Recovery offunction after spinal cord hemisection in newborn and adultrats: Differential effects on reflex and locomotor function. Exp.Neurol. 116: 40–51.
35. LAI, Y. Y., H. K. STRAHLENDORF, S. J. FUNG, AND C. D. BARNES.1989. The actions of two monoamines on spinal motoneuronsfrom stimulation of the locus coeruleus in the cat. Brain Res.484(1–2): 268–272.
36. LITTLE, J. W., R. M. HARRIS, AND S. J. LERNER. 1991. Immobiliza-tion impairs recovery after spinal cord injury. Arch. Phys. Med.Rehabil. 72(6): 408–412.
37. LITTLE, J. W., R. M. HARRIS, AND R. C. SOHLBERG. 1988. Locomo-tor recovery following subtotal spinal cord lesions in a ratmodel.Neurosci. Lett. 87(1–2): 189–194.
38. LIU, D. X., V. VALADEZ, L. S. SORKIN, AND D. J. MCADOO. 1990.Norepinephrine and serotonin release upon impact injury to ratspinal cord. J. Neurotrauma 7(4): 219–227.
39. MURRAY, M., AND M. E. GOLDBERGER. 1974. Restitution of func-tion and collateral sprouting in the cat spinal cord: the partiallyhemisected animal. J. Comp. Neurol. 158: 19–36.
40. NEWTON, B. W., AND R. W. HAMILL. 1988. The morphology anddistribution of rat serotoninergic intraspinal neurons:An immu-nohistochemical study. Brain Res. Bull. 20(3): 349–360.
41. PUNIAK, M. A., G. M. FREEMAN, C. A. AGRESTA, N. L. VAN, C. A.BARONE, AND S. K. SALZMAN. 1991. Comparison of a serotoninantagonist, opioid antagonist, and TRH analog for the acutetreatment of experimental spinal trauma. J. Neurotrauma 8(3):193–203.
42. SALZMAN, S. K., K. W. DABNEY, A. A. MENDEZ, J. T. BEAUCHAMP,J. C. DALEY, G. M. FREEMAN, A. FONSECA, E. B. INGERSOLL, A. L.BECKMAN, AND W. P. BUNNELL. 1988. The somatosensory evokedpotential predicts neurologic deficits and serotonergic patho-chemistry after spinal distraction injury in experimental scolio-sis. J. Neurotrauma 5(3): 173–186.
43. SALZMAN, S. K., E. HIROFUGI, P. B. KNIGHT, E. C. LLADOS, A. L.BECKMAN, AND A. WINOKUR. 1987. Treatment of experimentalspinal trauma with thyrotropin-releasing hormone: Centralserotonergic and vascular mechanisms of action. Cent. Nerv.Syst. Trauma 4(3): 181–196.
44. SALZMAN, S. K., E. HIROFUJI, E. C. LLADOS, G. D. MACEWEN, ANDA. L. BECKMAN. 1987.Monoaminergic responses to spinal trauma.Participation of serotonin in posttraumatic progression of neu-ral damage. J. Neurosurg. 66(3): 431–439.
45. SALZMAN, S. K., P. B. KNIGHT, E. HIROFUJI, E. C. LLADOS, A. L.BECKMAN, AND A. WINOKUR. 1987. Spinal cord immunoreactiveTRH is altered after local traumatic injury. Peptides 8(2):247–250.
46. SALZMAN, S. K., A. A. MENDEZ, K. W. DABNEY, J. C. DALEY, G. M.FREEMAN, S. EL TANTAWI, A. L. BECKMAN, AND W. P. BUNNELL.1991. Serotonergic response to spinal distraction trauma inexperimental scoliosis. J. Neurotrauma 8(1): 45–54.
47. SALZMAN, S. K., M. A. PUNIAK, Z. J. LIU, H. R. P. MAITLAND, G. M.FREEMAN, AND C. A. AGRESTA. 1991. The serotonin antagonistmianserin improves functional recovery following experimentalspinal trauma. Ann. Neurol. 30(4): 533–541.
48. SARUHASHI, Y., S. HUKUDA, AND T. MAEDA. 1990. Acute aggrega-tion of serotonin-immunoreactive platelets in the injured spinalcord of rat and change of serotonin content in the neural fibers.J. Neurotrauma 7(4): 237–246.
49. SARUHASHI, Y., S. HUKUDA, AND T. MAEDA. 1991. Evidence for aneural source of acute accumulation of serotonin in platelets inthe injured spinal cord of rats. An experimental study using5,6-dihydroxytryptamine treatment. J. Neurotrauma 8(2):121–128.
50. SARUHASHI, Y., AND W. YOUNG. 1994. Opposing effects of seroton-ergic 5-HT1A and 5-HT2 receptor agonists and antagonists onspinal axon excitability.Neuroscience, in press.
51. SARUHASHI, Y., AND W. YOUNG. Role of descending serotoninergicspinal axons in locomotory recovery of rats after lateral hemisec-tion. Exp. Neurol.
52. SHARMA, H. S., Y. OLSSON, AND P. K. DEY. 1990. Early accumula-tion of serotonin in rat spinal cord subjected to traumatic injury.Relation to edema and blood flow changes. Neuroscience 36:725–730.
212 SARUHASHI, YOUNG, AND PERKINS

53. SHARMA, H. S., J. S. WESTMAN, Y. OLSSON, O. JOHANSSON, ANDP. K. DEY. 1990. Increased 5-hydroxytryptamine immunoreactiv-ity in traumatized spinal cord. An experimental study in theract. Acta Neuropathol. 80: 12–17.
54. SORKIN, L. S., M. G. HUGHES, D. LIU, W. D. J. WILLIS, AND D. J.MCADOO. 1991. Release and metabolism of 5-hydroxytrypta-mine in the cat spinal cord examined with microdialysis. J.Pharmacol. Exp. Ther. 257(1): 192–199.
55. SORKIN, L. S., J. L. STEINMAN, M. G. HUGHES, W. D. WILLIS, ANDD. J. MCADOO. 1988. Microdialysis recovery of serotonin re-leased in spinal cord dorsal horn. J. Neurosci. Methods 23:131–138.
56. STEINBUSCH, H.W.M. 1981. Distribution of serotonin-immunore-activity in the central nervous system of the rat cell bodies andterminals.Neuroscience 6: 557–618.
57. STEWARD, O. 1989. Reorganization of neuronal connectionsfollowing CNS trauma: principles and experimental paradigms.J. Neurotrauma 6(2): 99–152.
58. TAKEUCHI, Y., H. KIMURA, ANDY. SANO. 1982. Immunohistochemi-cal demonstration of serotonin neurons in brainstem of the ratand cat. Cell Tissue Res. 224: 246–267.
59. WHITE, S. R., AND S. J. FUNG. 1989. Serotonin depolarizes catspinal motoneurons in situ and decreases motoneuron afterhy-perpolarizing potentials. Brain Res. 502(2): 205–213.
60. WINDLE, W. F., J. O. SMART, AND J. J. BEERS. 1958. Residualfunction after subtotal spinal cord transection in adult cats.Neurology 8: 518–521.
61. YOMONO, H. S., H. SUZUKI, AND K. YOSHIOKA. 1992. Serotonergicfibers induce a long-lasting inhibition of monosynaptic reflex inthe neonatal rat spinal cord.Neuroscience 47(3): 521–531.
62. YU, J., AND E. EIDELBERG. 1981. Effects of vestibulospinal lesionsupon locomotor function in cats. Brain Res. 220(1): 179–183.
63. ZIVIN, J. A., AND J. A. VENDITTO. 1984. Experimental CNSischemia: Serotonin antagonists reduce or prevent damage.Neurology 34: 469–474.
2135-HT RECOVERY AFTER THORACIC HEMISECTION