the male gametophyte of flowering plants - plant · pdf filethe male gametophyte of flowering...
TRANSCRIPT

The Plant Cell, Vol. 1, 657-664, July 1989 O 1989 American Society of Plant Physiologists
The Male Gametophyte of Flowering Plants
Joseph P. Mascarenhas
Department of Biological Sciences, State University of New York at Albany, Albany, New York 12222
REVIEW
INTRODUCTION AND OVERVIEW OF POLLEN DEVELOPMENT
Angiosperm plants produce flowers, very beautiful and intricate structures, within which their reproductive devel- opment takes place. In flowering plants, as in other groups of plants, a diploid, spore-producing generation (sporo- phyte) alternates with a haploid, gamete-producing gen- eration (gametophyte). Unlike some of the evolutionarily more primitive plants, the male and female gametophytes of angiosperms are reduced to microscopic structures that are dependent on the tissues of the sporophyte for their development. The flower contains specialized structures, the anthers and the pistil or gynoecium, in which the male and female gametophytes, respectively, are formed. The functions of the gametophytes are the production of the 'sperm cells and the female cells, and their union in fertilization. In flowering plants, the pollen grain is the male gametophyte and the embryo sac is the female gametoph yte.
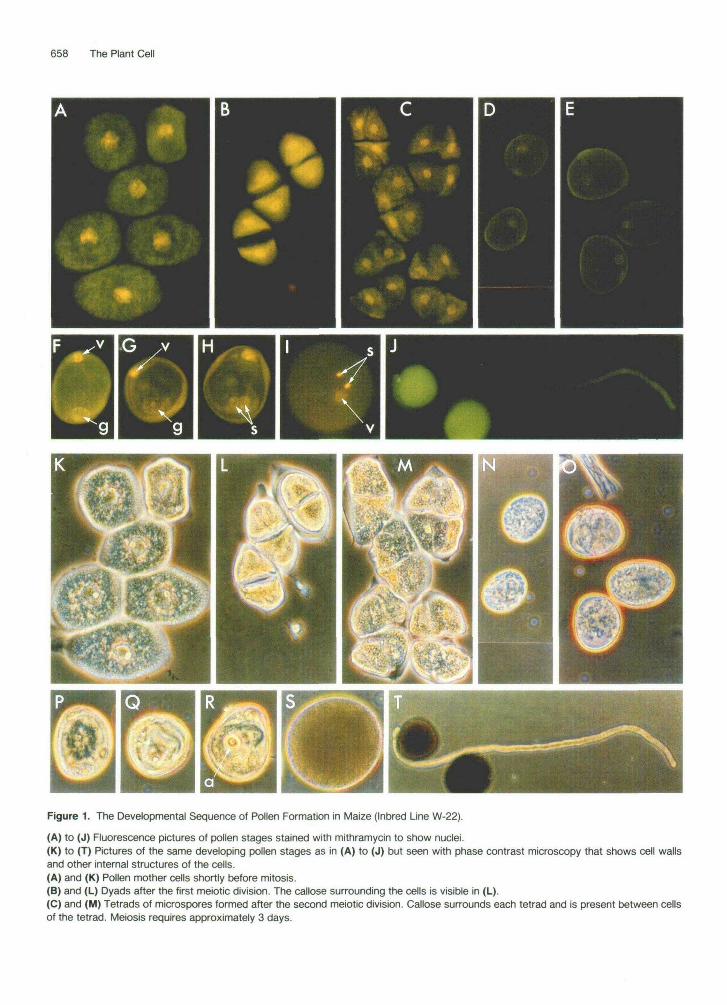
The male gametophyte completes its early development within the anther. The sequential stages of pollen devel- opment are shown in Figure 1. Microsporocytes or pollen mother cells (Figures 1 A and 1 K) are produced in the sporogenous tissue within the anther. The two divisions of meiosis transform these cells into haploid microspores, each pollen mother cell producing first a dyad (Figures 1 B and 1 L) and, after the second meiotic division, a tetrad of microspores (Figures 1C and 1M). The tetrad and each microspore within the tetrad are surrounded by a callose (1,3-P-glucan) wall (Figure 1M). Upon release from the tetrad, the microspores increase rapidly in volume and undergo a change in shape (Figures 1 D, 1 E, 1 N, and 1 O). This is followed by a period of slower growth until the maximum volume of the pollen grains is reached before anthesis. Following release of microspores from the te- trads, there is an extended interphase period that termi- nates with a very unequal division of the microspore (mi- crospore mitosis), forming a vegetative cell and a genera- tive cell, both of which are included within the confines of the cell wall of the original microspore. The vegetative cell constitutes the bulk of the young pollen grain, while the
generative cell, which inherits a very small amount of the microspore cytoplasm, lies within the vegetative cell (Fig- ures 1F and 1G). In severa1 plants, such as corn, the generative cell undergoes a mitotic division within the pollen grain, forming two sperm cells (Figures 1 H and 1 R). In most pollens, however, the generative cell completes its division during the growth of the pollen tube in the style. At maturity, the male gametophyte consists of three cells, the vegetative cell and the two sperm cells, which lie within the cytoplasm of the vegetative cell (Figures 1 I and 1 S).
For a short while following anthesis (the rupture of the anther and release of pollen), the mature pollen grain exists as a free organism until it is transported by wind, insects, or other agents to the stigma of an appropriate pistil. It then begins another phase of its life and development. Each pollen grain germinates by the extrusion of a tube through a germ pore in the pollen wall (Figures 1 J and 1T). The tube then grows down into the style and the vegetative nucleus, and, depending on the plant species, the gener- ative cell or sperm cells move out of the pollen grain and into the tube. Germination and pollen tube growth are relatively rapid events in most plants, the period from pollination to fertilization ranging from 1 hr to around 48 hr. The rate of pollen tube growth varies in different plant species; extremely rapid rates as high as 35 mm/hr have been reported for some plants. The pollen tube grows through the transmitting tissue of the style, enters the micropyle of the ovule, and reaches the embryo sac. It penetrates one of the synergids of the embryo sac (nor- mally the one that has begun to degenerate), the tube ruptures, and the sperm cells together with some of the other tube contents are discharged into the synergid. The two sperm move by a presently unknown mechanism, one fusing with the egg cell to form the diploid zygote. The other sperm fuses with the normally diploid central cell, giving rise to the primary endosperm nucleus. This com- pletes the process of double fertilization. A major recent advance is the observation that the two sperm cells in a pollen grain are often morphologically different (Russell,
658 The Plant Cell
Figure 1. The Developmental Sequence of Pollen Formation in Maize (Inbred Line W-22).
(A) to (J) Fluorescence pictures of pollen stages stained with mithramycin to show nuclei.(K) to (T) Pictures of the same developing pollen stages as in (A) to (J) but seen with phase contrast microscopy that shows cell wallsand other internal structures of the cells.(A) and (K) Pollen mother cells shortly before mitosis.(B) and (L) Dyads after the first meiotic division. The callose surrounding the cells is visible in (L).(C) and (M) Tetrads of microspores formed after the second meiotic division. Callose surrounds each tetrad and is present between cellsof the tetrad. Meiosis requires approximately 3 days.

Male Gametophyte of Flowering Plants 659
1985; McConchie et al., 1987), and which of the two fuses with the egg and which with the central cell is predeter- mined (Russell, 1985). Although extremely reduced in mor- phological complexity, the haploid male plant (gameto- phyte) undergoes a number of discrete differentiation events, and it has several very specialized functions.
This review will concentrate on the current status of our knowledge of the genes expressed in the developing male gametophyte. For a more extensive review of gene expres- sion during anther and pollen development, see Mascar- enhas (1988). Because of time and space constraints, the role of the tapetum and other anther tissues in pollen development has not been included in this review. The emerging area of study of flowering plant sperm cells has also not been covered. A recent publication edited by Wilms and Keijzer (1988) describes the current status of research on plant sperm.
TRANSCRIPTION ANO TRANSLATION DURING POLLEN MATURATION
Transcription and translation occur in the microspore after its formation and during its subsequent development. Con- clusive evidence that transcription and translation from the haploid genome occur during pollen development has been obtained from studies of several dimeric enzymes. The Adh-7 (alcohol dehydrogenase) gene in maize specifies a dimeric enzyme that is responsible for the ADH activity of the pollen grain (Felder, Scandalios, and Liu, 1973; Freeling and Schwartz, 1973). The enzyme is the product of the activation of the Adh-7 gene in the male gametophyte. Pollen extracts from plants heterozygous for two Adh-7 electrophoretic variants, fast (F) and slow (S), show only the two homodimeric enzymes (FF and SS) and no heter- odimers (FS) (Schwartz, 1971). Since two alleles are pres- ent at each locus in somatic tissues, but only one or the other allele is present in individual pollen grains, these
results would only be expected if haploid transcription and translation occurred, and if the dimer in extracts did not dissociate and reassociate to form hybrids. If, however, the enzyme in the pollen grains were synthesized prior to meiosis, all three enzymes, FF, FS, and SS, would be present in each haploid pollen grain, as is found in sporo- phytic tissue. Heterodimers were found when the individual pollen grains carried two Adh-7 alleles, as was the case with the Adh-FCM duplication. These results indicate that the synthesis of the ADH enzyme in pollen depends solely on the genotype of the pollen nuclei and is not influenced by the genotype of the diploid plant (Schwartz, 1971; 1981). Evidence for haploid transcription has been ob- tained for other enzymes in corn (Brink, and MacGillivray, 1924; Demerec, 1924; Nelson, 1958; Sari-Gorla et al., 1986), and in other plants (Weeden, and Gottlieb, 1979; Tanksley, Zamir, and Rick, 1981; Miller and Mulcahy, 1983; Singh, O’Neill, and Knox, 1985).
Both in lily and in Tfadescantia, large amounts of ribo- soma1 RNA are synthesized prior to microspore mitosis. Following this mitosis, a sharp decrease occurs in rRNA synthesis, and these genes become transcriptionally in- active during the terminal stages of pollen maturation and during subsequent pollen tube growth (Steffensen, 1966; Mascarenhas and Bell, 1970; Peddada and Mascarenhas, 1975; see also review in Mascarenhas, 1975). The tran- scription of the transfer RNA genes also follows a similar pattern (see review in Mascarenhas, 1975). However, pro- teins are synthesized during pollen germination and tube growth utilizing the ribosomes and tRNAs made and ac- cumulated during pollen maturation. Many pollens contain all the proteins, mRNAs, ribosomes, etc., to enable rapid germination and initial tube growth on the stigma. The environment of the pollen grain from the time of its release from the anther to deposition on the stigma surface is quite inhospitable. The trend in evolution has, accordingly, been toward the pollen grain containing all that is neces- sary to germinate rapidly to enable the tube to penetrate the stigma where conditions are favorable for its further development (see review in Mascarenhas, 1975).
Figure 1 (continued).
(D) and (N) Early uninucleate microspores. (E) and (O) Late uninucleate microspores. Seven days to 9 days elapse between release of microspores from tetrads and the start of microspore mitosis, which extends over a period of 2 days. (F) and (P) Binucleate pollen grain after microspore mitosis. (G) and (a) Late binucleate pollen grain with the generative cell in late prophase of mitosis. (H) and (R) Trinucleate pollen grain, with the generative cell having just completed mitosis, forming two sperm cells. Generative cell division takes approximately 2 days. (I) and (S) Mature pollen grain showing two sperm cells and the nucleus of the vegetative cell. Anthesis occurs 6 days to 7 days after sperm cell formation. (J) and (T) Germinating pollen tube. The magnification in all the figures is the same with the exception of (J) and (T), which are at half the magnification. Relative size differences are thus evident. The diameter of the mature pollen grain in (S) is 95 pm. g, nucleus of the generative cell; s, nucleus of a sperm cell; v, nucleus of the vegetative cell; a, aperture or germ pore in the pollen wall.

660 The Plant Cell
ESTIMATES OF THE NUMBER OF GENES EXPRESSED IN THE MALE GAMETOPHYTE
The mature pollen grain at the time of anthesis contains a store of mRNAs that were synthesized during maturation (see review in Mascarenhas, 1988). From the kinetics of hybridization of 3H-cDNA with poly(A) RNA in excess, the mRNAs in mature Tradescantia paludosa pollen have been found to consist of three abundance classes with com- plexities of 5.2 x 1 04, I .6 x 1 06, and 2.1 x 1 O7 nucleotides (Willing and Mascarenhas, 1984). Approximately 15% of the mRNA is very abundant and consists of about 40 different sequences, each present on an average in 26,000 copies per pollen grain. The bulk of the mRNA (60%) comprises about 1400 diverse sequences, each present in about 3400 copies per pollen grain. The least abundant fraction (24%) consists of 18,000 different sequences, each present on an average in 1 O0 copies per pollen grain. In pollen, about 75% of the mRNAs occur in the two more abundant frequency classes, whereas only 35% of the shoot mRNAs are abundant. The least abundant class of shoot mRNAs (64%) consists of 29,000 different se- quences, each present in 5 copies to 1 O copies per cell. In the least abundant fraction in pollen, the sequences are much more abundant (about 1 O0 copies) than in the cor- responding fraction in shoots. The total complexity of Tradescantia pollen poly(A) RNA is 2.3 X 1 O7 nucleotides, which is equivalent to about 20,000 different transcripts, whereas, in shoots, it is 3.4 x 1 O7 nucleotides, correspond- ing to about 30,000 different sequences.
The complexity of maize pollen poIy(A) RNAs is very similar to that found for Tradescantia pollen (Willing, Bashe, and Mascarenhas, 1988). The total complexity of corn pollen poly(A) RNA is 2.4 x 107 nucleotides, whereas, in the seedling shoot, it is 4.0 X 107 nucleotides. This is equivalent to about 24,000 different mRNAs in mature pollen and about 31,000 different mRNAs in shoots. The mRNAs in maize pollen fall into three abundance classes. The most abundant class comprises 35% of the mRNAs with about 240 sequences each present in about 32,000 copies per pollen grain. The middle abundance class, which makes up the major fraction of the mRNA (49%), consists of about 6000 different sequences present on average in 1700 copies per pollen grain. About 17,000 different se- quences, each with about 200 copies per grain, make up the least abundant fraction, which is 15% of the total mRNA of the mature pollen grain.
The hybridization studies with both corn and Tradescan- tia show that a large number of genes are transcribed in the male gametophyte. One should bear in mind that the 20,000 to 24,000 differeni mRNAs are the sequences found in mature pollen and mostly represent genes that are activated late in pollen development. It is likely that additional genes might be active immediately after meiosis and during the early stages of pollen development. At
present, we have no estimates of their numbers or similar- ity to the genes expressed later in pollen development. The complexity of mRNA sequences in the mature pollen grains of both Tradescantia and corn are about 60% those of shoot mRNAs, i.e., approximately 60% as many genes are expressed in pollen as are expressed in the vegetative shoot (Willing and Mascarenhas, 1984; Willing, Bashe, and Mascarenhas, 1988).
In general, the mRNAs in pollen are much more prevalent than those in the shoot. Neither maize nor Tradescantia shoots contain mRNAs present in excess of 25,000 copies per cell, and the most abundant fraction in shoots contains mRNAs that are present in only a few hundred copies per cell. Even the least abundant fraction in pollen includes sequences that are much more abundant (100 copies to 200 copies) than in the corresponding fraction in shoots (5 copies to 1 O copies). This abundance of mRNAs in mature póllen would seem to suggest a requirement for the rapid synthesis of a large quantity of protein product during the terminal stages of pollen maturation, and/or germination and early pollen tube growth.
OVERLAP OF THE GENETIC PROGRAMS UTlLlZED IN THE SPOROPHYTE AND GAMETOPHYTE
Are specific sets of genes transcribed during male game- tophyte development or is there an overlap of the genetic programs of the sporophyte and the gametophyte? Iso- zymes for nine dimeric enzyme systems encoded in the sporophyte by 28 genes were compared in pollen and in diploid vegetative tissues of tomato (Tanksley, Zamir, and Rick, 1981). Sixty percent of the isozymes found in roots, leaves, or developing and mature seeds are also found in pollen, indicating that 60% of the structural genes coding for these enzymes in the sporophyte are expressed in the male gametophyte. In addition, 18 of 19 pollen-expressed isozymes (95%) are expressed in one or more vegetative tissues. A similar analysis in maize found that 72% of the isozymes studied are expressed in both the sporophyte and the male gametophyte, whereas only 6% of the iso- zymes are specific to pollen (Sari-Gorla et al., 1986). A similar overlap between isozyme profiles in pollen and sporophyte tissues has been reported for three Populus species (Rajora and Zsuffa, 1986). Between 74% and 80% of the isozymes in the sporophyte are expressed in the male gametophyte. Similarly in barley, 60% of the iso- zymes studied are expressed in both the male gameto- phyte and sporophyte tissues; 30% are sporophyte-spe- cific and 10% are male gametophyte-specific (Pedersen, Simonsen, and Loeschcke, 1987). An extensive overlap of gene expression between the endosperm and the male gametophyte also occurs (Ottaviano, Petroni, and Pe, 1988).

Male Gametophyte of Flowering Plants 661
Heterologous hybridizations of pollen cDNA with shoot poly(A) RNA and shoot cDNA to pollen poly(A) RNA indi- cate that no more than 60% of the shoot mRNAs are represented in pollen, whereas a minimum of 64% of the pollen mRNA mass is found in shoot RNA (Willing and Mascarenhas, 1984). A minimum of 65% of the mRNAs in maize pollen are similar to those found in shoots. Because of various constraints in this type of analysis, the estimate of pollen sequences shared with the sporophyte could be in excess of 90% (Willing, Bashe, and Mascarenhas, 1988).
It has been estimated, based on colony hybridizations with cDNA libraries made to pollen poly(A) RNA and hy- bridized with 32P-cDNAs from pollen and vegetative tis- sues, that about 10% and 20% of the total sequences expressed in maize and Jradescantia pollen, respectively, are pollen-specific (Stinson et al., 1987). From the evidence discussed, it is apparent that the genetic program ex- pressed during pollen development is extensive and that a substantial overlap occurs between genes active in the sporophyte and in the male gametophyte. Thus, it should be possible to use pollen to screen for desirable traits in the sporophyte, and such studies have been initiated (Mul- cahy, 1979; Mulcahy and Bergamini-Mulcahy, 1987; see review in Mascarenhas, 1988).
In addition to a possible significant sharing of genes in pollen and sporophyte, there is evidence that some en- zymes in pollen are coded for by genes that are different from those coding for the Same enzyme activity in sporo- phytic tissues, i.e., the genes code for nonallelic isozymes. ADP-glucose pyrophosphorylase is possibly coded in pol- len and the endosperm by nonallelic genes (Bryce and Nelson, 1979). P-Glucosidase in corn is another example of an enzyme that is encoded by different genes in pollen and sporophyte (Frova, Binelli, and Ottaviano, 1987).
By immunoblotting two-dimensional gels of various tis- sue extracts with anti-tubulin monoclonal antibodies, six well-defined, P-tubulin isotypes were found to be ex- pressed differentially within the carrot plant (Hussey, Lloyd, and Gull, 1988). The p5- and P6-tubulin isotypes are absent from reproductive organs. P,-Tubulin is found in stamens and is the most abundant P-tubulin in the mature pollen grain. The pl- and Ps-tubulins are expressed in all the organs of the floret except the pollen grain. These results would seem to suggest that, in carrot, different P-tubulin genes might be expressed in pollen and vegetative tissues, although it is possible that post-translational modifications could produce the multiple P-tubulins. Ludwig et al. (1988) have used 3’-noncoding, gene-specific probes with RNA blot hybridizations to study al- and a,-tubulin gene expres- sion in various tissues of Arabidopsis thaliana. The O(,-
tubulin transcripts were found in roots, leaves, and flowers. The al-tubulin gene-specific probe hybridized very weakly to RNA from roots and leaves, but strongly to RNA from flowers. Of several stages in flower development, the a, transcript is most abundant in flowers actively shedding pollen. The results seem to suggest a preferential expres-
sion of the a1 gene in pollen, but, unfortunately, in this study, the levels of the O ( , transcripts were not determined in pollen.
ISOLATION OF GENES EXPRESSED IN POLLEN AND THE PATTERN OF TRANSCRIPTION AND ACCUMULATION OF POLLEN-SPECIFIC mRNAs
Severa1 laboratories have recently constructed cDNA li- braries to poly(A) RNA from mature pollen. These include libraries to pollen sequences from Tradescantia paludosa and maize (Stinson et al., 1987), from tomato (Twell et ai, 1989), and from Oenothera organensis (Brown, 1988; S.M.G. Brown and M.L. Crouch, manuscript in prepara- tion). The libraries from Tradescantia and maize contain several clones that are specific to pollen and other clones that are expressed in pollen and in vegetative tissues (Stinson et al., 1987). DNA gel blot hybridizations indicate that the pollen-expressed genes that have been analyzed are present in one or a very few copies in the maize genome (Stinson et al., 1987). A tomato gene, LAT52, that is expressed in abundance in pollen and anthers and weakly in petals, appears to be present in one copy in the genome (Twell et al., 1989). The six analyzed, pollen- expressed cDNA clones in Oenothera (Brown, 1988) in- clude single copy genes and members of relatively small families (showing 3 bands to 12 bands on the DNA hybrid- ization blots).
RNA blot hybridizations using pollen-specific cDNA clones as probes have shown in both Tradescantia and maize that the mRNAs are first detectable in the young pollen grain after microspore mitosis. They continue to accumulate thereafter, reaching their maximum concentra- tion in the mature pollen grain. Using an actin clone as a probe, actin mRNA was first detectable in Tradescantia during the microspore interphase after release from the tetrads. It accumulated thereafter, reaching a maximum concentration at late pollen interphase before decreasing substantially in the mature pollen grain (Stinson et al., 1987). These results indicate that there are at least two sets of genes with respect to the timing of their activity during pollen development. The first set of genes repre- sented by the pollen-specific cDNA clones in the libraries are activated after microspore mitosis, and the mRNAs increase in content up to maturity. This pattern of accu- mulation would suggest a major function for these mRNAs during germination and early tube growth in addition to possible functions during the latter part of pollen matura- tion. The second group, which includes genes like actin, are active soon after meiosis is completed. The mRNAs reach their maximum accumulation by late pollen inter- phase and then decrease substantially by anthesis (Stin- son et al., 1987). ADH and p-galactosidase are probably other examples of genes that are expressed early after

662 The Plant Cell
meiosis in a manner similar to that of actin. The appearance and pattern of increase of ADH enzyme activity in maize (Stinson and Mascarenhas, 1985) and p-galactosidase in Brassica campestris (Singh, O’Neill, and Knox, 1985) are what would be expected if their mRNAs were synthesized in a fashion similar to that of actin in Tradescantia. Glu- tamic-oxaloacetic transaminase and p-glucosidase in maize are other examples of genes that appear to be activated early in pollen development (Frova, Binelli, and Ottaviano, 1987).
Mutants of A. thaliana deficient in adenine phosphori- bosyltransferase activity (conversion of adenine and 5- phosphoribosyl pyrophosphate to adenylic acid) were iden- tified by selecting for seed germination on a medium containing 2,6-diaminopurine (Moffatt and Somerville, 1988). The diaminopurine tolerance is due to a recessive nuclear mutation at the apt locus. The mutants are male- sterile, with pollen abortion occurring after release of the microspores from the tetrads. The tapetum in the mutant anthers is morphologically normal. This is probably the first known nuclear male-sterile mutant in which the biochemi- cal basis of the sterility has been determined. How a defect in purine salvage causes male sterility is not clear, but it has been suggested that the defect may be due to a change in the cytokinin metabolism.
The LAT52 mRNA from tomato shows a similar accu- mulation profile, being first detectable in immature anthers and increasing progressively until anthesis (Twell et al., 1989). In Oenothera, the mRNAs for three pollen-specific clones, P1, P2, and P3, seem to follow a very similar pattern of accumulation, being first detectable in immature pollen and reaching their maximum levels in mature pollen (Brown, 1988). The transcripts for one other pollen-specific clone, P6, are not detectable until fairly late in pollen development. The mRNAs for two clones, P4 and P5, which are expressed in pollen and in leaves and ovaries, are present at all stages of pollen development (Brown, 1988).
SEQUENCE CHARACTERIZATION OF POLLEN cDNA AND GENOMIC CLONES
A pollen-specific cDNA clone from maize, Zmcl3, which is a full-length copy of the mRNA, has been sequenced (Hanson et al., 1989). It is 929 nucleotides in length and codes for a predicted polypeptide of 170 amino acid resi- dues with a molecular mass of 18.3 kD. The hydropathy profile suggests a possible signal sequence at the amino terminus. The mRNA contains a 5’-untranslated region of 127 nucleotides and a 3’-untranslated region of 292 nu- cleotides (to the polyadenylation site). In Situ hybridizations using RNA probes show that the mRNA is located in the cytoplasm of the vegetative cell of the pollen grain, and,
after germination, is distributed throughout the pollen tube cytoplasm.
The LAT52 anther-specific gene from tomato encodes an 800-nucleotide-long transcript that has 5’-untranslated and 3’-untranslated regions of 11 O nucleotides and ap- proximately 150 nucleotides, respectively (Twell et al., 1989). It contains a single intron with an unusual structure, being composed almost entirely of a directly repeated 46- nucleotide sequence. The cDNA clone contains an open reading frame that codes for a putative protein of 17.8 kD, which has an amino-terminal hydrophobic region with char- acteristics of a signal sequence. The predicted peptide sequence of LAT52 exhibits substantial amino acid identity with the putative protein sequence of the pollen-specific sequence from maize, Zm13 (Hanson et al., 1989), de- scribed above. Neither Zml3 nor LAT52, however, shows significant sequence homology with any known protein in DNA or protein data banks.
Two of the cDNA clones from the P1/P2 family of Oenotbera pollen cDNAs were sequenced, and putative amino acid sequences determined (Brown, 1988). Sub- stantial homology was found in the amino acid sequence between the two clones and a polygalacturonase from tomato. Antibodies prepared to a portion of the protein coding region were used in immunoblots to determine the timing of synthesis and accumulation of the protein during pollen development. The protein was found to be present in pollen at later stages of development and in pollen tubes, indicating that the mRNA is not merely stored for use during germination.
Two cDNA clones expressed in tomato pollen share similarity in amino acid sequence with pectate lyases from Erwinia (Wing et al., 1989). It is interesting that the two pollen-expressed genes that have been tentatively identi- fied with respect to function are enzymes that are involved in degradation of the middle lamella region of cell walls. Thus, they could be involved in the solubilization of cell wall material during growth of the pollen tube through the tissues of the style.
It will be interesting to discover the functions of the severa1 other pollen-specific genes that have been isolated. The primary synthetic events that occur during pollen tube growth are concerned with pollen tube wall synthesis and the synthesis of the cell membrane of the elongating tube. Accordingly, one might expect that many of the presyn- thesized mRNAs and proteins in the mature pollen grain code for enzymes required for these two processes to occur. Unfortunately, our knowledge of the enzymology of plant cell wall synthesis is very primitive. This deficiency will make it more difficult to identify the many gene prod- ucts present in the pollen grain that are probably involved with tube wall synthesis.
The genomic clone of LAT52 including 666 bp of 5‘- flanking DNA and 200 bp of 3‘-flanking DNA has been sequenced (Twell et al., 1989). Approximately 1300 bp of the 5’-flanking region of the Zm13 gene has been se-

Male Gametophyte of Flowering Plants 663
quenced (D.A. Hamilton, D.M. Bashe, J. Stinson, and J.P. Mascarenhas, manuscript in preparation). Unfortunately, there are no regions of identity in the presumptive promoter regions of the 5’-flanking sequences of these two pollen- expressed genes.
The promoter regions of two other pollen-expressed but not pollen-specific genes have been sequenced. The Adh-1 gene from maize is expressed in pollen and in other tissues of the plant. The complete sequence of the gene, including extensive flanking regions, has been determined (Sachs et al., 1986). The waxy (wx) locus of maize is another genetically well-characterized gene. It encodes the UDP-g1ucose:starch glycosyltransferase that is bound to starch granules and is expressed in pollen, in the embryo sac, and in the endosperm (Echt and Schwartz, 1981). About 900 bp of 5’-flanking sequences of the wx gene together with 3718 bp of the coding region, which is composed of 14 exons and 13 small introns, and several hundred base pairs of 3’-flanking sequences are available (Klosgen et al., 1986). A comparison of the promoter regions of the different available pollen-expressed genes might prove informative. Mutation and deletion analysis of pollen-specific promoters currently under study by several groups should provide valuable information about tran- scriptional regulatory elements in the male gametophyte.
CONCLUSIONS
Although the mature male gametophyte of flowering plants is morphologically simple, during its development it under- goes a series of differentiation events. A large number of genes are required to program its entire development. Most of the genes expressed during pollen development appear also to be expressed in the sporophyte. Recent increased interest in the male gametophyte has resulted in the isolation of pollen-expressed sequences including pollen-specific cDNAs and genes from several plants. These pollen-specific genes are now being characterized with respect to promoter elements required for male ga- metophyte-specific gene regulation, and one might expect rapid progress in this work. Determining the functions of the pollen-specific genes in pollen development is a major future goal. The mRNAs tentatively identified in mature pollen seem to be metabolic genes that might be involved in pollen tube growth in the style. A greater challenge will be to identify those genes that are critical in initiating the unique developmental events during microsporogenesis.
ACKNOWLEDGMENTS
I thank Dr. Sheila McCormick for graciously making available a copy of a manuscript in press, Dr. Sherri M.G. Brown for providing
a copy of her Ph.D. thesis, David Bashe and Dr. Jeffrey L. Travis for help with the photomicroscopy, Ryland Loos for the artwork, and Dr. Gregory Smutzer for critically reading the manuscript. Work in the author’s laboratory is supported by Grant DCB88- 1 O1 65 from the National Science Foundation.
Received May 15,1989.
REFERENCES
Brink, R.A., and MacGillivray, J.H. (1 924). Segregation for the waxy character in maize pollen and differential development of the male gametophyte. Am. J. Bot. 11, 465-469.
Brown, S.M.G. (1 988). Molecular analysis of gene expression during pollen development in Oenofhera organensis. Ph.D. the- sis, Indiana University.
Bryce, W.H., and Nelson, O.E. (1979). Starch synthesizing en- zymes in the endosperm and pollen of maize. Plant Physiol. 63,
Demerec, M. (1 924). A case of pollen dimorphism in maize. Am.
Echt, C.S., and Schwartr, D. (1981). Evidence for the inclusion of controlling elements within the structural gene at the waxy locus in maize. Genetics 99, 275-284.
Felder, M.R., Scandalios, J.R., and Liu, E.H. (1 973). Purification and partia1 characterization of two genetically defined alcohol dehydrogenase isozymes in maize. Biochim. Biophys. Acta 317,
Freeling, M., and Schwartr, D. (1 973). Genetic relationships between the multiple alcohol dehydrogenases of maize. Biochem. Genet. 8,27-36.
Frova, C., Binelli, O., and Ottaviano, E. (1987). lsozyme and hsp gene expression during male gametophyte development in maize. In Isozymes: Current Topics in Biological and Medical Research, Vol. 15, Genetics, Development and Evolution, M.C. Rattazzi and J.G. Scandalios, eds (New York: Alan R. Liss), pp. 97-1 20.
Hanson, D.D., Hamilton, D.A., Travis, J.L., Bashe, D.M., and Mascarenhas, J.P. (1 989). Characterization of a pollen-specific cDNA clone from Zea mays and its expression. Plant Cell 1,
Hussey, P.J., Lloyd, C.W., and Gull, K. (1988). Differential and developmental expression of 8-tubulins in a higher plant. J. Biol. Chem. 263,5474-5479.
Klosgen, R.B., Gierl, A., Schwarz-Sommer, Z., and Saedler, H. (1986). Molecular analysis of the waxy locus of Zea mays. MOI. Gen. Genet. 203,237-244.
Ludwig, S.R., Oppenheimer, D.G., Silflow, C.D., and Snustad, D.P. (1988). The a,-tubulin gene of Arabidopsis thaliana: Pri- mary structure and preferential expression in flowers. Plant MOI. Biol. 10, 31 1-321.
Mascarenhas, J.P. (1 975). l h e biochemistry of angiosperm pollen development. Bot. Rev. 41, 259-314.
Mascarenhas, J.P. (1 988). Anther- and pollen-expressed genes.
31 2-31 7.
J. BOt. 11,461-464.
149-1 59.
173-1 79.

664 The Plant Cell
In Temporal and Spatial Regulation of Plant Genes, D.P.S. Verma and R.B. Goldberg, eds (New York: Springer-Verlag),
Mascarenhas, J.P., and Bell, E. (1970). RNA synthesis during development of the male gametophyte of Tradescantia. Dev. Biol. 21, 475-490.
McConchie, C.A., Russell, S.D., Dumas, C., Tuohy, M., and Knox, R.B. (1987). Quantitative cytology of the sperm cells of Brassica campestris and E. oleracea. Planta 170, 446-452.
Miller, J.C., and Mulcahy, D.L. (1 983). Microelectrophoresis and the study of genetic overlap. In Pollen: Biology and Applications in Plant Breeding, D.L. Mulcahy and E. Ottaviano, eds (New York: Elsevier/North Holland), pp. 31 7-321.
Moffatt, B., and Somerville, C. (1988). Positive selection for male sterile mutants of Arabidopsis lacking adenine phosphoribosyl transferase activity. Plant Physiol. 86, 11 50-1 154.
Mulcahy, D.L. (1979). The rise of the angiosperms: A genecol- ogical factor. Science 206, 20-23.
Mulcahy, D.L., and Bergamini-Mulcahy, G. (1 987). The effects of pollen competition. Am. Sci. 75, 44-50.
Nelson, O.E. (1 958). lntracistron recombination in the Wx/wx region of maize. Science 130, 794-795.
Ottaviano, E., Petroni, D., and Pe, M.E. (1988). Gametophytic expression of genes controlling endosperm development in maize. Theor. Appl. Genet. 74, 252-258.
Peddada, L., and Mascarenhas, J.P. (1 975). 5s ribosomal RNA synthesis during pollen development. Dev. Growth Diff. 17,
Pedersen, S., Simonsen, V., and Loeschcke, V. (1987). Overlap of gametophytic and sporophytic gene expression in barley. Theor. Appl. Genet. 75, 200-206.
Rajora, O.P., and Zsuffa, L. (1986). Sporophytic and gametophy- tic gene expression in Populus deltoides Marsh, P. nigra L, and P. maximowiczii Henry. Can. J. Genet. Cytol. 28, 476-482.
Russell, S.D. (1 985). Preferential fertilization in Plumbago: Ultra- structural evidence for gamete-leve1 recognition in an angio- sperm. Proc. Natl. Acad. Sci. USA 82,6129-6132.
Sachs, M.M., Dennis, E.S., Gerlach, W.L., and Peacock, W.J. (1986). Two alleles of maize alcohol dehydrogenase 1 have 3‘ structural and poly(A) addition polymorphisms. Genetics 113,
Sari-Gorla, M., Frova, C., Binelli, G., and Ottaviano, E. (1986). The extent of gametophytic-sporophytic gene expression in
pp. 97-1 15.
1-8.
449-467.
maize. Theor. Appl. Genet. 72, 42-47. Schwattz, D. (1 971). Genetic control of alcohol dehydrogenase-
A competition model for regulation of gene action. Genetics 67,
Schwartr, D. (1 981). Adh locus in maize for detection of mutagens in the environment. Environ. Health Perspect. 37, 75-77.
Singh, M.B., O’Neill, P., and Knox, R.B. (1985). lnitiation of postmeiotic 6-galactosidase synthesis during microsporogene- sis in oilseed rape. Plant Physiol. 77, 225-228.
Steffensen, D.M. (1 966). Synthesis of ribosomal RNA during growth and division in Lilium. Exp. Cell Res. 44, 1-12.
Stinson, J., and Mascarenhas, J.P. (1 985). Onset of alcohol dehydrogenase synthesis during microsporogenesis in maize. Plant Physiol. 77, 222-224.
Stinson, J.R., Eisenberg, A.J., Willing, R.P., Pe, M.E., Hanson, D.D., and Mascarenhas, J.P. (1987). Genes expressed in the male gametophyte of flowering plants and their isolation. Plant Physiol. 83, 442-447.
Tanksley, S.D., Zamir, D., and Rick, C.M. (1981). Evidence for extensive overlap of sporophytic and gametophytic gene expression in Lycopersicon esculenfum. Science 213, 454- 455.
Twell, D., Wing, R., Yamaguchi, J., and McCormick, S. (1989). lsolation and expression of an anther-specific gene from tomato. MOI. Gen. Genet., in press.
Weeden, F., and Gottlieb, L.D. (1 979). Distinguishing allozymes and isozymes of phosphogluco-isomerases by electrophoretic comparisons of pollen and somatic tissues. Biochem. Genet.
Willing, R.P., and Mascarenhas, J.P. (1 984). Analysis of the complexity and diversity of mRNAs from pollen and shoots of Tradescantia. Plant Physiol. 75, 865-868.
Willing, R.P., Bashe, D., and Mascarenhas, J.P. (1988). An analysis of the quantity and diversity of messenger RNAs from pollen and shoots of Zea mays. Theor. Appl. Genet. 75, 751- 753.
Wilms, H.J., and Keijrer, C.J., eds (1988). Plant Sperm Cells as Tools for Biotechnology (Wageningen, The Netherlands: Pudoc).
Wing, R., Yamaguchi, J., Larabell, S., Ursin, V., and McCormick, S. (1989). Floral specific cDNAs from tomato encode proteins homologous to pectate lyase from the plant pathogen Erwinia. J. Cell. Biochem. Suppl. 13D, p. 294 (abstract).
41 1-425.
17,287-296.

DOI 10.1105/tpc.1.7.657 1989;1;657-664Plant Cell
J. P. MascarenhasThe Male Gametophyte of Flowering Plants.
This information is current as of May 17, 2018
Permissions X
https://www.copyright.com/ccc/openurl.do?sid=pd_hw1532298X&issn=1532298X&WT.mc_id=pd_hw1532298
eTOCs http://www.plantcell.org/cgi/alerts/ctmain
Sign up for eTOCs at:
CiteTrack Alerts http://www.plantcell.org/cgi/alerts/ctmain
Sign up for CiteTrack Alerts at:
Subscription Information http://www.aspb.org/publications/subscriptions.cfm
is available at:Plant Physiology and The Plant CellSubscription Information for
ADVANCING THE SCIENCE OF PLANT BIOLOGY © American Society of Plant Biologists