the influence of ige-enhancing and ige-suppressive t local ige levels (1). ige abs are also induced...
TRANSCRIPT

of July 13, 2018.This information is current as
Exposure to Inhaled Ovalbumin T Cells Changes withδγIgE-Suppressive
The Influence of IgE-Enhancing and
Willi K. BornandAydintug, J. M. Wands, Hua Huang, Rebecca L. O'Brien
Yafei Huang, Niyun Jin, Christina L. Roark, M. Kemal
http://www.jimmunol.org/content/183/2/849doi: 10.4049/jimmunol.08041042009;
2009; 183:849-855; Prepublished online 19 JuneJ Immunol
Referenceshttp://www.jimmunol.org/content/183/2/849.full#ref-list-1
, 15 of which you can access for free at: cites 32 articlesThis article
average*
4 weeks from acceptance to publicationFast Publication! •
Every submission reviewed by practicing scientistsNo Triage! •
from submission to initial decisionRapid Reviews! 30 days* •
Submit online. ?The JIWhy
Subscriptionhttp://jimmunol.org/subscription
is online at: The Journal of ImmunologyInformation about subscribing to
Permissionshttp://www.aai.org/About/Publications/JI/copyright.htmlSubmit copyright permission requests at:
Email Alertshttp://jimmunol.org/alertsReceive free email-alerts when new articles cite this article. Sign up at:
Print ISSN: 0022-1767 Online ISSN: 1550-6606. Immunologists, Inc. All rights reserved.Copyright © 2009 by The American Association of1451 Rockville Pike, Suite 650, Rockville, MD 20852The American Association of Immunologists, Inc.,
is published twice each month byThe Journal of Immunology
by guest on July 13, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
by guest on July 13, 2018
http://ww
w.jim
munol.org/
Dow
nloaded from

The Influence of IgE-Enhancing and IgE-Suppressive �� TCells Changes with Exposure to Inhaled Ovalbumin1
Yafei Huang,*† Niyun Jin,*† Christina L. Roark,*† M. Kemal Aydintug,*† J. M. Wands,*†
Hua Huang,*‡ Rebecca L. O’Brien,*† and Willi K. Born2*†
It has been reported that the IgE response to allergens is influenced by �� T cells. Intrigued by a study showing that airwaychallenge of mice with OVA induces in the spleen the development of �� T cells that suppress the primary IgE response toi.p.-injected OVA-alum, we investigated the �� T cells involved. We found that the induced IgE suppressors are contained withinthe V�4� subset of �� T cells of the spleen, that they express V�5 and CD8, and that they depend on IFN-� for their function.However, we also found that normal nonchallenged mice harbor IgE-enhancing �� T cells, which are contained within the largerV�1� subset of the spleen. In cell transfer experiments, airway challenge of the donors was required to induce the IgE suppressorsamong the V�4� cells. Moreover, this challenge simultaneously turned off the IgE enhancers among the V�1� cells. Thus, airwayallergen challenge differentially affects two distinct subsets of �� T cells with nonoverlapping functional potentials, and theoutcome is IgE suppression. The Journal of Immunology, 2009, 183: 849–855.
A ntibodies of the IgE class are prominent in the host re-sponse to parasitic infections and in allergic responses tomany nonpathogenic Ags (1). The interest in IgE has
heightened as ever-increasing proportions of the world’s popula-tion suffer from allergies (2). In healthy mammals, IgE Abs ac-quired via the gastrointestinal tract by the newborn may serve as afirst line of defense (3). IgE is synthesized and functions in thenormal adult largely in the mucosal tissues where the IgE concen-trations are high, whereas concentrations of IgE in the circulationremain low by comparison with other Igs (1). Mechanisms re-sponsible for this biased anatomical distribution include the dis-tribution and longevity of cells that express the receptors forIgE, as most of the IgE in the tissues is cell bound and thusprotected from degradation, and local IgE synthesis, which isfavored by the Th2 environment of the mucosal tissues thatmaintains local IgE levels (1).
IgE Abs are also induced during vaccination. Aluminum adju-vants (alum), currently the most widely used adjuvants in humanand animal vaccines, stimulate the innate system and can favorTh2-biased reactivity (4). Immunization of previously untreatedlaboratory animals with soluble inert protein Ags using alum typ-ically elicits Th2-type responses, accompanied by the developmentof IgE Abs. However, the outcome of this type of immunizationalso depends on prior exposure. Mucosal exposure to the same Agmay result in nonresponsiveness and the failure of the immuniza-tion to elicit Th2 reactivity (5). For example, repeated airway
challenge of rodents with OVA without adjuvant leads to non-responsiveness to a subsequent i.p. injection of OVA-alum,which otherwise would induce Th2 reactivity and a strong OVA-specific IgE response (6). How airway exposure alters the outcomeof OVA-alum immunization is not yet fully understood (7).
Several groups have provided evidence that �� T cells can mod-ulate the OVA-alum-induced IgE response. Investigating the de-velopment of tolerance to inhaled OVA, McMenamin et al. (8)found that �� T cells from tolerized mice efficiently and selectivelysuppressed primary OVA-alum-induced IgE responses. In appar-ent contrast, others reported that �� T cells are required for thedevelopment of IgE responses to OVA and other Ags (9, 10). Theunderlying mechanisms responsible for these apparently opposingobservations remained unresolved.
That �� T cells can exert both Th1-like and Th2-like effectson the immune responses to pathogens has been recognized(11), and later studies revealed the surprising circumstance thatthese different and sometimes opposed functional effects on thehost responses segregate with TCR-V�-definable subsets of ��
T cells, such as V�4� and V�1� �� T cells (12, 13). Specificfunctional contributions of these and other TCR-defined subsetssuggested that the �� TCR determines not only ligand speci-ficity of �� T cells but also their functional potential (14). Con-sistent with this concept, a recent study showed that the abilityof �� T cells with specificity for the T22 molecule to expressIL-17 and IFN-� depends on TCR-ligand interactions duringtheir development (15).
Given the divergent observations regarding the role of �� T cellsin the IgE response, we were interested in determining whetherdifferent TCR-V�-definable subsets of �� T cells also exert op-posed effects on IgE production. In the current study, we tookadvantage of the observation that normal mice immunized with asingle i.p injection of OVA-alum make a primary IgE response toOVA. The results of these experiments indicate that V�1� �� Tcells are able to enhance the primary IgE response induced byOVA-alum, whereas V�4� cells in contrast are able to suppress it.Moreover, we found that in addition to their different functionalpotentials, the overall effect of the IgE-modulating �� T cells crit-ically depends on the exposure history of the animal. Whereas the
*Integrated Department of Immunology, National Jewish Health, Denver, CO 80206;†University of Colorado, Denver, CO 80206; and ‡Department of Medicine, NationalJewish Health, Denver, CO 80206
Received for publication December 9, 2008. Accepted for publication May 6, 2009.
The costs of publication of this article were defrayed in part by the payment of pagecharges. This article must therefore be hereby marked advertisement in accordancewith 18 U.S.C. Section 1734 solely to indicate this fact.1 This work was supported by National Institutes of Health Grants AI40611 andHL65410 (to W.K.B.) and AI44920 and AI063400 (to R.L.O.).2 Address correspondence and reprint requests to Dr. Willi K. Born, Integrated De-partment of Immunology, National Jewish Health, 1400 Jackson Street, GB K409,Denver, CO 80206. E-mail address: [email protected]
Copyright © 2009 by The American Association of Immunologists, Inc. 0022-1767/09/$2.00
The Journal of Immunology
www.jimmunol.org/cgi/doi/10.4049/jimmunol.0804104
by guest on July 13, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

IgE-enhancing �� T cells lose this ability upon airway allergen-exposure, the IgE-suppressive �� T cells gain theirs under thesame circumstance.
Materials and MethodsAnimals
Female C57BL/6 mice and several mutant strains of the same genetic back-ground (B6.TCR-��/�, B6.TCR-��/�, B6.TCR-��/���/�) were obtainedfrom The Jackson Laboratory. TCR-V�4�/�6�/� mice deficient in V�4�
and V�6� T cells were a gift from Dr. K. Ikuta (Kyoto University, Kyoto,Japan). They were backcrossed to the C57BL/6 genetic background andused after 11 backcross generations. B6.TCR-��/�IFN-��/� mice weregenerated by crossing the single mutants and breeding double mutantsidentified in the F2 generation. B6.TCR-V�1-transgenic mice were a giftfrom Dr. Pablo Pereira (Institut Pasteur, Paris, France). All mice were 8–12wk old at the time of the experiments. Mice were maintained on an OVA-free diet and were cared for at National Jewish Health (Denver, CO), fol-lowing guidelines for immune deficient animals. All experiments were con-ducted under a protocol approved by the Institutional Animal Care and UseCommittee.
Ag exposure and immunization
The animals were exposed to 1% OVA (w/v) (5� crystalline; EMD Bio-sciences) in saline aerosol inhalation for 30 min daily, 5 days/wk for up to2 wk, and subsequently once per wk (designated 10N in this paper), fol-lowing a method described by others (8). To induce OVA-specific IgE,mice were immunized by i.p. injection of 10 �g of OVA in aluminumhydroxide (AlumImject; Pierce Scientific) (8).
Treatment with Abs against the TCR
Hamster pan anti-TCR-C� mAbs (clone GL3), anti-V�4 mAb (clone UC3),and anti-V�1 mAb (clone 2.11) were purified from hybridoma culture su-pernatants using a protein G-Sepharose affinity column (Amersham Phar-macia Biotech). T cells were targeted by injection of 200 �g of hamsteranti-TCR-�, anti-V�4 or anti-V�1 mAbs into the tail veins of mice 4 daysbefore the i.p. immunization with OVA. The effect of these treatments onthe targeted T cells was monitored as previously described (13). This ap-proach, which is based on staining with non-cross-blocking anti-TCRmAbs, allows an assessment of the effect of treatment on TCR expression,but it does not assess the fate of the targeted T cells. The antibody treat-ments transiently reduce TCR expression by �90%. Sham Ab treatmentswere performed with nonspecific hamster IgG (The Jackson Laboratory).Throughout this article, we use the nomenclature for murine TCR-V�genes introduced by Heilig and Tonegawa (16).
T cell purification from spleen and lung
Total spleens and lungs were harvested from naive or 10N mice at the timeof the experiments. Lungs were dissected into small pieces and exposed toan enzymatic digestion mixture containing 0.125% dispase II (Roche),0.2% collagenase II (Sigma-Aldrich), and 0.2% collagenase IV (Sigma-Aldrich) for 90 min at 37°C. After enzymatic digestion, a single-cell sus-pension was prepared by pushing the lung tissue fragments through a 70-�m-diameter nylon mesh (BD Falcon). A suspension of splenocytes wasprepared by mechanical dispersion. Cell suspensions were treated withGey’s RBC lysis solution and passed through nylon wool columns to ob-tain T lymphocyte-enriched cell preparations containing �75% T cells aspreviously described (13). Total cell counts were determined using aCoulter counter.
Adoptive transfer of �� T cells
Splenic nylon wool-nonadherent (NAD) cells from naive and 10N mice(B6.TCR-��/�, B6.TCR-��/�IFN-��/�) were incubated with biotinylatedanti-V�4 mAb (clone UC3) or anti-V�1 mAb (clone 2.11) for 15 min at4°C, washed, and incubated with streptavidin-conjugated magnetic beads(Streptavidin Microbeads; Miltenyi Biotec) for 15 min at 4°C and passedthrough magnetic columns to purify V�4� or V�1� cells as previouslydescribed in detail (17). This produced a cell population containing �90%V�4� or V�1� viable cells as determined by two-color staining with anti-TCR-� and anti-V�4 or anti-V�1 mAbs. These splenic V�4� or V�1�
cells were washed in PBS and resuspended to a concentration of 1.5 � 105
cells/ml PBS, and 3 � 104 cells/mouse were injected in 200 �l of PBSvia the tail vein into B6.TCR-��/� mice directly before the OVAimmunization.
In some experiments, subpopulations of V�4� or V�1� cells were pu-rified using the MoFlo cell sorter. NAD cells were incubated with FITC-
conjugated anti-V�4 mAb (clone UC3) or FITC-conjugated anti-V�1 mAb(clone 2.11) and PE-conjugated anti-CD8� (clone 53-6.7; BD Pharmingen)or biotinylated anti-V�5 (clone F45-152), followed by PE-conjugatedstreptavidin (20 min at 4°C), and then washed. Cells were next sorted basedon their expression of V� chain and V�5 or CD8� using a MoFlo cellsorter (Dako Cytomation). Purified cells were washed in PBS and resus-pended to a concentration of 1.5 � 105 cells/ml of PBS, and 3 � 104
cells/mouse injected in 200 �l of PBS via the tail vein into B6.TCR-��/�
mice directly before the OVA immunization. In general, the cell sorter-purified cells were less effective than those prepared by magnetic beadselection and therefore have only been compared with each other.
Flow cytometric analysis
For flow cytometric analyses, NAD cells (2 � 105/well) in 96-well plates(Falcon; BD Biosciences) were stained for TCR expression using a PE-labeledpan C� Ab (clone GL3) and FITC-conjugated anti-V�1 (clone 2.11), or anti-V�4 (clone UC3) Ab followed by biotinylated anti-V� mAbs: anti-V�4 (cloneGL-2), anti-V�5 (clone F45-152), anti-V� 6.3 (clone 17-C), anti-V�6�12(clone F4.22) and anti V�8 (clone B20.1), plus streptavidin-allophycocyanin.All samples were analyzed on a FACScan flow cytometer (BD Biosciences)counting a minimum of 25,000 events per gated region, and the data wereprocessed using FlowJo 6.4.1 software (Tree Star).
Determination of serum IgE levels
Sera were harvested on day 14 after the i.p. immunization with OVA-alum.For OVA-specific serum IgE determinations, plates were coated overnightat 4°C with 2 �g/ml rat anti-mouse IgE Ab (clone R35-72; BD Bio-sciences). The serum samples were then added, and biotin-labeled OVAsubsequently added to the wells. Before biotinylation, OVA was first dia-lyzed at 4°C overnight against 0.1 M borate buffer (pH 8.4). BiotinylatedOVA was then prepared by reacting 1 ml of OVA in PBS (1 mg/ml) with150 �l of N-hydroxysuccinimidobiotin in DMSO (1 mg/ml) for 4 h at roomtemperature, followed by overnight dialysis against PBS at 4°C. The boundOVA-biotin was detected with streptavidin-conjugated HRP (BD Bio-sciences) followed by 100 �l/well of TMB substrate solution. OVA-spe-cific IgE levels in the samples were compared with an internal standardobtained from pooled sera of hyperimmunized BALB/c mice, which wasarbitrarily assigned to equal 1000 ELISA U. Total IgE levels were mea-sured by sandwich ELISA using rat anti-mouse IgE at 2 �g/ml (cloneR35-72; BD Biosciences) as a capture Ab followed by biotinylated ratanti-mouse IgE H chain mAb (clone R35-118; BD Biosciences) at 2 �g/ml,and detected as described above. All samples were read using a VERSA-max tunable microplate reader and processed using a SoftMax Pro 4.7.1software.
Statistical analysis
Data are presented as means � SEM. The unpaired t test was used for twogroup comparisons, and ANOVA was used for analysis of differences inthree or more groups. Statistically significant levels are indicated as fol-lows: �, p � 0.05; ��, p � 0.01; ���, p � 0.001; NS, not significant.
ResultsAltered IgE response in �� T cell genetically modified mice andin normal mice treated with Abs against the �� TCR
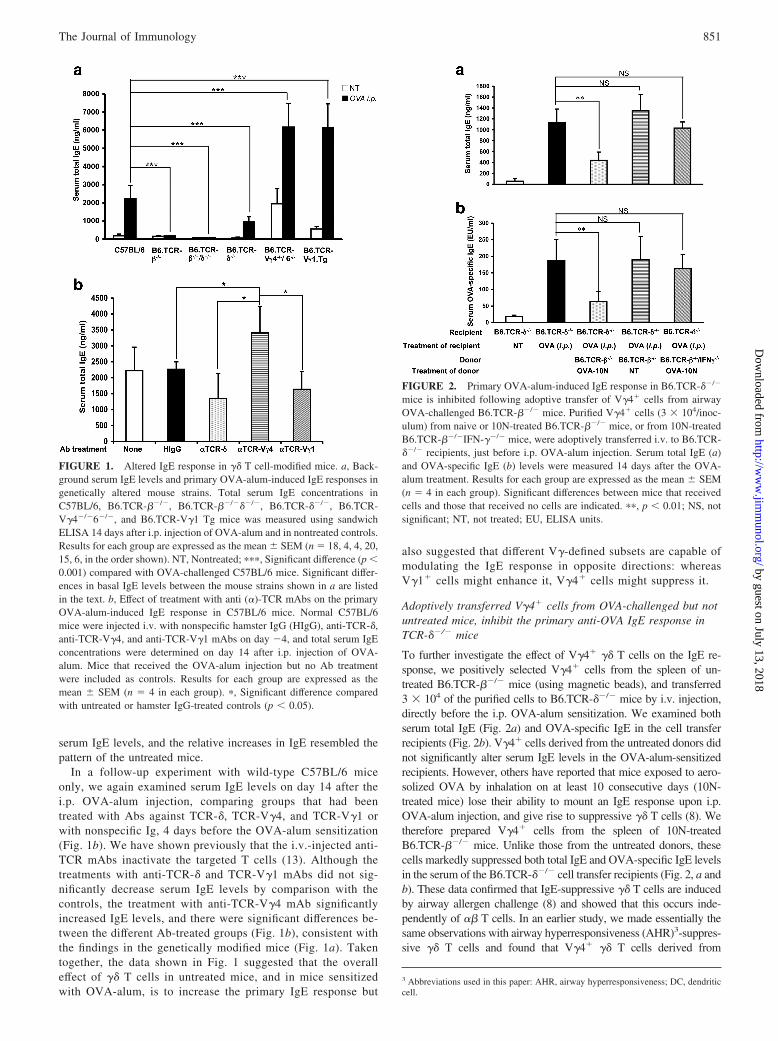
To test the concept that �� T cells are capable of modulating theIgE response, we initially examined genetically modified mice,including mice lacking all T cells (B6.TCR-��/���/�), all �� Tcells (B6.TCR-��/�), V�4� and V�6� �� T cells only (B6.TCR-V�4�/�6�/�), and mice expressing a rearranged V�1J�4C�4transgene (B6.TCR-V�1 Tg), plus wild-type controls (C57BL/6).We also examined mice lacking all �� T cells (B6.TCR-��/�).Total serum IgE levels were measured by ELISA, in adult age- andsex-matched untreated mice, and in mice injected i.p. with OVA-alum, 14 days after the injection (Fig. 1a). Without treatment,basal serum IgE levels were low in comparison with wild-typecontrols in mice lacking all �� T cells (B6.TCR-��/� vs C57BL/6,p � 0.01), but high in mice deficient in V�4� and V�6� �� T cells(B6.TCR-V�4�/�6�/� vs C57BL/6, p � 0.0001) and in mice ex-pressing the V�1J�4C�4 transgene (B6.TCR-V�1 transgenic vsC57BL/6, p � 0.001). Mice lacking �� T cells did not express IgEat significant levels. At day 14 after OVA-alum injection, all miceexcept those lacking �� T cells exhibited substantially increased
850 IgE REGULATION BY �� T CELLS
by guest on July 13, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
serum IgE levels, and the relative increases in IgE resembled thepattern of the untreated mice.
In a follow-up experiment with wild-type C57BL/6 miceonly, we again examined serum IgE levels on day 14 after thei.p. OVA-alum injection, comparing groups that had beentreated with Abs against TCR-�, TCR-V�4, and TCR-V�1 orwith nonspecific Ig, 4 days before the OVA-alum sensitization(Fig. 1b). We have shown previously that the i.v.-injected anti-TCR mAbs inactivate the targeted T cells (13). Although thetreatments with anti-TCR-� and TCR-V�1 mAbs did not sig-nificantly decrease serum IgE levels by comparison with thecontrols, the treatment with anti-TCR-V�4 mAb significantlyincreased IgE levels, and there were significant differences be-tween the different Ab-treated groups (Fig. 1b), consistent withthe findings in the genetically modified mice (Fig. 1a). Takentogether, the data shown in Fig. 1 suggested that the overalleffect of �� T cells in untreated mice, and in mice sensitizedwith OVA-alum, is to increase the primary IgE response but
also suggested that different V�-defined subsets are capable ofmodulating the IgE response in opposite directions: whereasV�1� cells might enhance it, V�4� cells might suppress it.
Adoptively transferred V�4� cells from OVA-challenged but notuntreated mice, inhibit the primary anti-OVA IgE response inTCR-��/� mice
To further investigate the effect of V�4� �� T cells on the IgE re-sponse, we positively selected V�4� cells from the spleen of un-treated B6.TCR-��/� mice (using magnetic beads), and transferred3 � 104 of the purified cells to B6.TCR-��/� mice by i.v. injection,directly before the i.p. OVA-alum sensitization. We examined bothserum total IgE (Fig. 2a) and OVA-specific IgE in the cell transferrecipients (Fig. 2b). V�4� cells derived from the untreated donors didnot significantly alter serum IgE levels in the OVA-alum-sensitizedrecipients. However, others have reported that mice exposed to aero-solized OVA by inhalation on at least 10 consecutive days (10N-treated mice) lose their ability to mount an IgE response upon i.p.OVA-alum injection, and give rise to suppressive �� T cells (8). Wetherefore prepared V�4� cells from the spleen of 10N-treatedB6.TCR-��/� mice. Unlike those from the untreated donors, thesecells markedly suppressed both total IgE and OVA-specific IgE levelsin the serum of the B6.TCR-��/� cell transfer recipients (Fig. 2, a andb). These data confirmed that IgE-suppressive �� T cells are inducedby airway allergen challenge (8) and showed that this occurs inde-pendently of �� T cells. In an earlier study, we made essentially thesame observations with airway hyperresponsiveness (AHR)3-suppres-sive �� T cells and found that V�4� �� T cells derived from
3 Abbreviations used in this paper: AHR, airway hyperresponsiveness; DC, dendriticcell.
FIGURE 1. Altered IgE response in �� T cell-modified mice. a, Back-ground serum IgE levels and primary OVA-alum-induced IgE responses ingenetically altered mouse strains. Total serum IgE concentrations inC57BL/6, B6.TCR-��/�, B6.TCR-��/���/�, B6.TCR-��/�, B6.TCR-V�4�/�6�/�, and B6.TCR-V�1 Tg mice was measured using sandwichELISA 14 days after i.p. injection of OVA-alum and in nontreated controls.Results for each group are expressed as the mean � SEM (n � 18, 4, 4, 20,15, 6, in the order shown). NT, Nontreated; ���, Significant difference (p �0.001) compared with OVA-challenged C57BL/6 mice. Significant differ-ences in basal IgE levels between the mouse strains shown in a are listedin the text. b, Effect of treatment with anti (�)-TCR mAbs on the primaryOVA-alum-induced IgE response in C57BL/6 mice. Normal C57BL/6mice were injected i.v. with nonspecific hamster IgG (HIgG), anti-TCR-�,anti-TCR-V�4, and anti-TCR-V�1 mAbs on day �4, and total serum IgEconcentrations were determined on day 14 after i.p. injection of OVA-alum. Mice that received the OVA-alum injection but no Ab treatmentwere included as controls. Results for each group are expressed as themean � SEM (n � 4 in each group). �, Significant difference comparedwith untreated or hamster IgG-treated controls (p � 0.05).
FIGURE 2. Primary OVA-alum-induced IgE response in B6.TCR-��/�
mice is inhibited following adoptive transfer of V�4� cells from airwayOVA-challenged B6.TCR-��/� mice. Purified V�4� cells (3 � 104/inoc-ulum) from naive or 10N-treated B6.TCR-��/� mice, or from 10N-treatedB6.TCR-��/�IFN-��/� mice, were adoptively transferred i.v. to B6.TCR-��/� recipients, just before i.p. OVA-alum injection. Serum total IgE (a)and OVA-specific IgE (b) levels were measured 14 days after the OVA-alum treatment. Results for each group are expressed as the mean � SEM(n � 4 in each group). Significant differences between mice that receivedcells and those that received no cells are indicated. ��, p � 0.01; NS, notsignificant; NT, not treated; EU, ELISA units.
851The Journal of Immunology
by guest on July 13, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
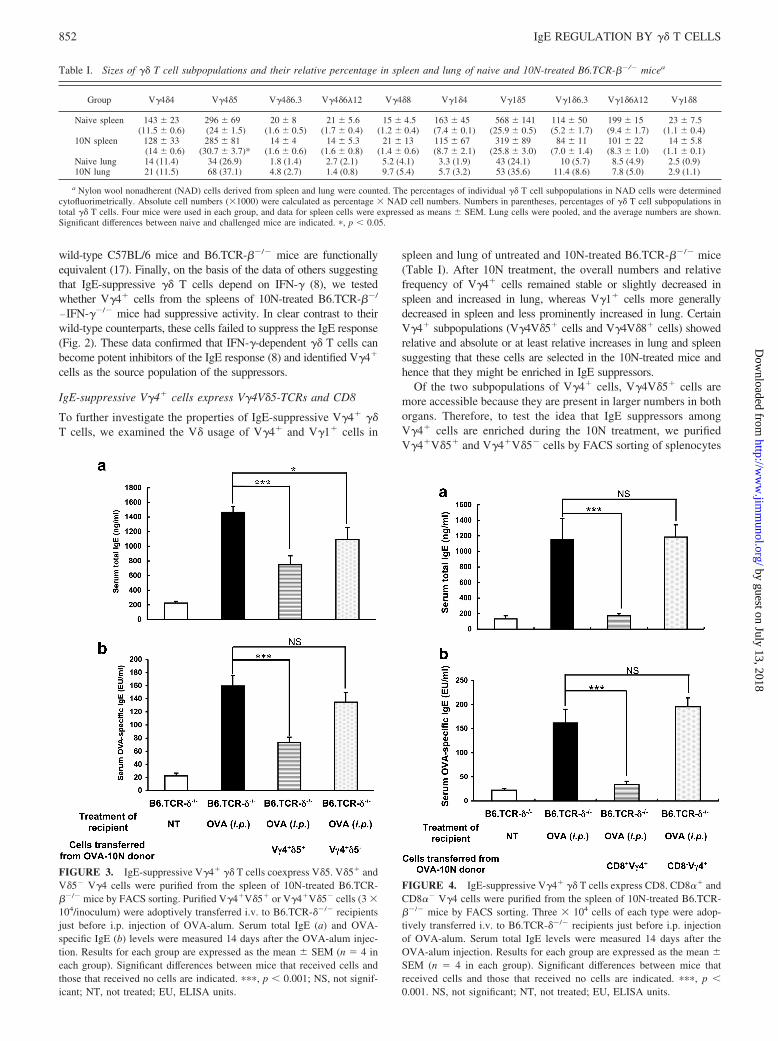
wild-type C57BL/6 mice and B6.TCR-��/� mice are functionallyequivalent (17). Finally, on the basis of the data of others suggestingthat IgE-suppressive �� T cells depend on IFN-� (8), we testedwhether V�4� cells from the spleens of 10N-treated B6.TCR-��/
�IFN-��/� mice had suppressive activity. In clear contrast to theirwild-type counterparts, these cells failed to suppress the IgE response(Fig. 2). These data confirmed that IFN-�-dependent �� T cells canbecome potent inhibitors of the IgE response (8) and identified V�4�
cells as the source population of the suppressors.
IgE-suppressive V�4� cells express V�4V�5-TCRs and CD8
To further investigate the properties of IgE-suppressive V�4� ��T cells, we examined the V� usage of V�4� and V�1� cells in
spleen and lung of untreated and 10N-treated B6.TCR-��/� mice(Table I). After 10N treatment, the overall numbers and relativefrequency of V�4� cells remained stable or slightly decreased inspleen and increased in lung, whereas V�1� cells more generallydecreased in spleen and less prominently increased in lung. CertainV�4� subpopulations (V�4V�5� cells and V�4V�8� cells) showedrelative and absolute or at least relative increases in lung and spleensuggesting that these cells are selected in the 10N-treated mice andhence that they might be enriched in IgE suppressors.
Of the two subpopulations of V�4� cells, V�4V�5� cells aremore accessible because they are present in larger numbers in bothorgans. Therefore, to test the idea that IgE suppressors amongV�4� cells are enriched during the 10N treatment, we purifiedV�4�V�5� and V�4�V�5� cells by FACS sorting of splenocytes
FIGURE 3. IgE-suppressive V�4� �� T cells coexpress V�5. V�5� andV�5� V�4 cells were purified from the spleen of 10N-treated B6.TCR-��/� mice by FACS sorting. Purified V�4�V�5� or V�4�V�5� cells (3 �104/inoculum) were adoptively transferred i.v. to B6.TCR-��/� recipientsjust before i.p. injection of OVA-alum. Serum total IgE (a) and OVA-specific IgE (b) levels were measured 14 days after the OVA-alum injec-tion. Results for each group are expressed as the mean � SEM (n � 4 ineach group). Significant differences between mice that received cells andthose that received no cells are indicated. ���, p � 0.001; NS, not signif-icant; NT, not treated; EU, ELISA units.
FIGURE 4. IgE-suppressive V�4� �� T cells express CD8. CD8�� andCD8�� V�4 cells were purified from the spleen of 10N-treated B6.TCR-��/� mice by FACS sorting. Three � 104 cells of each type were adop-tively transferred i.v. to B6.TCR-��/� recipients just before i.p. injectionof OVA-alum. Serum total IgE levels were measured 14 days after theOVA-alum injection. Results for each group are expressed as the mean �SEM (n � 4 in each group). Significant differences between mice thatreceived cells and those that received no cells are indicated. ���, p �0.001. NS, not significant; NT, not treated; EU, ELISA units.
Table I. Sizes of �� T cell subpopulations and their relative percentage in spleen and lung of naive and 10N-treated B6.TCR-��/� micea
Group V�4�4 V�4�5 V�4�6.3 V�4�6�12 V�4�8 V�1�4 V�1�5 V�1�6.3 V�1�6�12 V�1�8
Naive spleen 143 � 23 296 � 69 20 � 8 21 � 5.6 15 � 4.5 163 � 45 568 � 141 114 � 50 199 � 15 23 � 7.5(11.5 � 0.6) (24 � 1.5) (1.6 � 0.5) (1.7 � 0.4) (1.2 � 0.4) (7.4 � 0.1) (25.9 � 0.5) (5.2 � 1.7) (9.4 � 1.7) (1.1 � 0.4)
10N spleen 128 � 33 285 � 81 14 � 4 14 � 5.3 21 � 13 115 � 67 319 � 89 84 � 11 101 � 22 14 � 5.8(14 � 0.6) (30.7 � 3.7)* (1.6 � 0.6) (1.6 � 0.8) (1.4 � 0.6) (8.7 � 2.1) (25.8 � 3.0) (7.0 � 1.4) (8.3 � 1.0) (1.1 � 0.1)
Naive lung 14 (11.4) 34 (26.9) 1.8 (1.4) 2.7 (2.1) 5.2 (4.1) 3.3 (1.9) 43 (24.1) 10 (5.7) 8.5 (4.9) 2.5 (0.9)10N lung 21 (11.5) 68 (37.1) 4.8 (2.7) 1.4 (0.8) 9.7 (5.4) 5.7 (3.2) 53 (35.6) 11.4 (8.6) 7.8 (5.0) 2.9 (1.1)
a Nylon wool nonadherent (NAD) cells derived from spleen and lung were counted. The percentages of individual �� T cell subpopulations in NAD cells were determinedcytofluorimetrically. Absolute cell numbers (�1000) were calculated as percentage � NAD cell numbers. Numbers in parentheses, percentages of �� T cell subpopulations intotal �� T cells. Four mice were used in each group, and data for spleen cells were expressed as means � SEM. Lung cells were pooled, and the average numbers are shown.Significant differences between naive and challenged mice are indicated. �, p � 0.05.
852 IgE REGULATION BY �� T CELLS
by guest on July 13, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
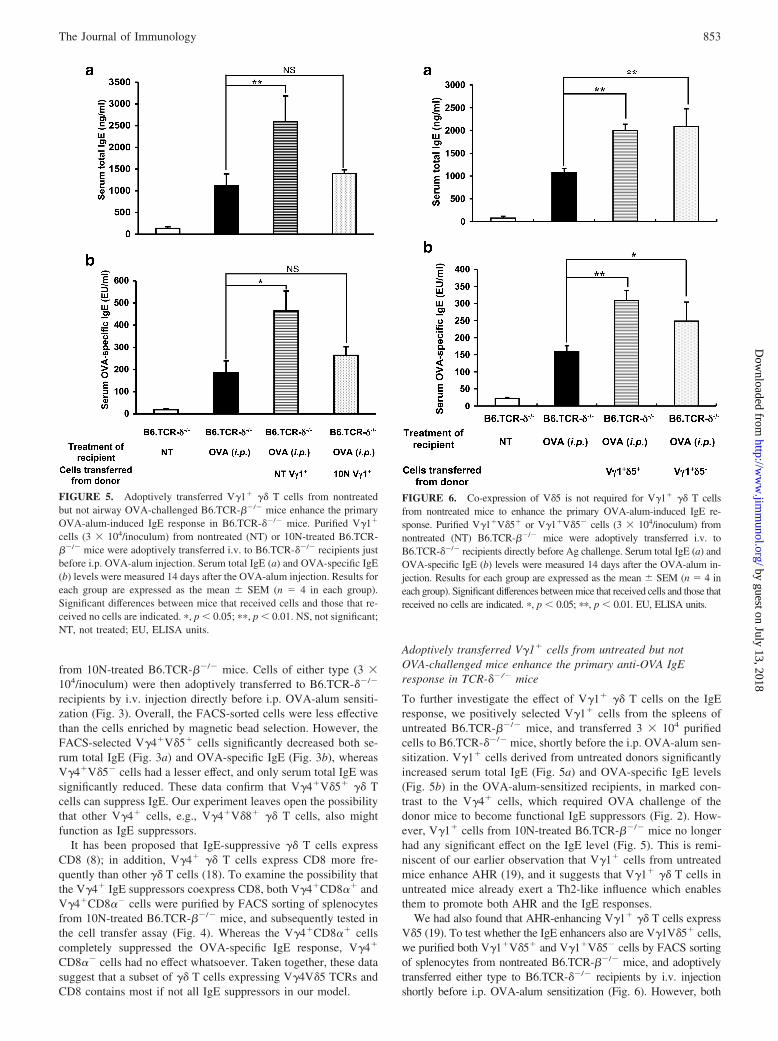
from 10N-treated B6.TCR-��/� mice. Cells of either type (3 �104/inoculum) were then adoptively transferred to B6.TCR-��/�
recipients by i.v. injection directly before i.p. OVA-alum sensiti-zation (Fig. 3). Overall, the FACS-sorted cells were less effectivethan the cells enriched by magnetic bead selection. However, theFACS-selected V�4�V�5� cells significantly decreased both se-rum total IgE (Fig. 3a) and OVA-specific IgE (Fig. 3b), whereasV�4�V�5� cells had a lesser effect, and only serum total IgE wassignificantly reduced. These data confirm that V�4�V�5� �� Tcells can suppress IgE. Our experiment leaves open the possibilitythat other V�4� cells, e.g., V�4�V�8� �� T cells, also mightfunction as IgE suppressors.
It has been proposed that IgE-suppressive �� T cells expressCD8 (8); in addition, V�4� �� T cells express CD8 more fre-quently than other �� T cells (18). To examine the possibility thatthe V�4� IgE suppressors coexpress CD8, both V�4�CD8�� andV�4�CD8�� cells were purified by FACS sorting of splenocytesfrom 10N-treated B6.TCR-��/� mice, and subsequently tested inthe cell transfer assay (Fig. 4). Whereas the V�4�CD8�� cellscompletely suppressed the OVA-specific IgE response, V�4�
CD8�� cells had no effect whatsoever. Taken together, these datasuggest that a subset of �� T cells expressing V�4V�5 TCRs andCD8 contains most if not all IgE suppressors in our model.
Adoptively transferred V�1� cells from untreated but notOVA-challenged mice enhance the primary anti-OVA IgEresponse in TCR-��/� mice
To further investigate the effect of V�1� �� T cells on the IgEresponse, we positively selected V�1� cells from the spleens ofuntreated B6.TCR-��/� mice, and transferred 3 � 104 purifiedcells to B6.TCR-��/� mice, shortly before the i.p. OVA-alum sen-sitization. V�1� cells derived from untreated donors significantlyincreased serum total IgE (Fig. 5a) and OVA-specific IgE levels(Fig. 5b) in the OVA-alum-sensitized recipients, in marked con-trast to the V�4� cells, which required OVA challenge of thedonor mice to become functional IgE suppressors (Fig. 2). How-ever, V�1� cells from 10N-treated B6.TCR-��/� mice no longerhad any significant effect on the IgE level (Fig. 5). This is remi-niscent of our earlier observation that V�1� cells from untreatedmice enhance AHR (19), and it suggests that V�1� �� T cells inuntreated mice already exert a Th2-like influence which enablesthem to promote both AHR and the IgE responses.
We had also found that AHR-enhancing V�1� �� T cells expressV�5 (19). To test whether the IgE enhancers also are V�1V�5� cells,we purified both V�1�V�5� and V�1�V�5� cells by FACS sortingof splenocytes from nontreated B6.TCR-��/� mice, and adoptivelytransferred either type to B6.TCR-��/� recipients by i.v. injectionshortly before i.p. OVA-alum sensitization (Fig. 6). However, both
FIGURE 5. Adoptively transferred V�1� �� T cells from nontreatedbut not airway OVA-challenged B6.TCR-��/� mice enhance the primaryOVA-alum-induced IgE response in B6.TCR-��/� mice. Purified V�1�
cells (3 � 104/inoculum) from nontreated (NT) or 10N-treated B6.TCR-��/� mice were adoptively transferred i.v. to B6.TCR-��/� recipients justbefore i.p. OVA-alum injection. Serum total IgE (a) and OVA-specific IgE(b) levels were measured 14 days after the OVA-alum injection. Results foreach group are expressed as the mean � SEM (n � 4 in each group).Significant differences between mice that received cells and those that re-ceived no cells are indicated. �, p � 0.05; ��, p � 0.01. NS, not significant;NT, not treated; EU, ELISA units.
FIGURE 6. Co-expression of V�5 is not required for V�1� �� T cellsfrom nontreated mice to enhance the primary OVA-alum-induced IgE re-sponse. Purified V�1�V�5� or V�1�V�5� cells (3 � 104/inoculum) fromnontreated (NT) B6.TCR-��/� mice were adoptively transferred i.v. toB6.TCR-��/� recipients directly before Ag challenge. Serum total IgE (a) andOVA-specific IgE (b) levels were measured 14 days after the OVA-alum in-jection. Results for each group are expressed as the mean � SEM (n � 4 ineach group). Significant differences between mice that received cells and those thatreceived no cells are indicated. �, p � 0.05; ��, p � 0.01. EU, ELISA units.
853The Journal of Immunology
by guest on July 13, 2018http://w
ww
.jimm
unol.org/D
ownloaded from
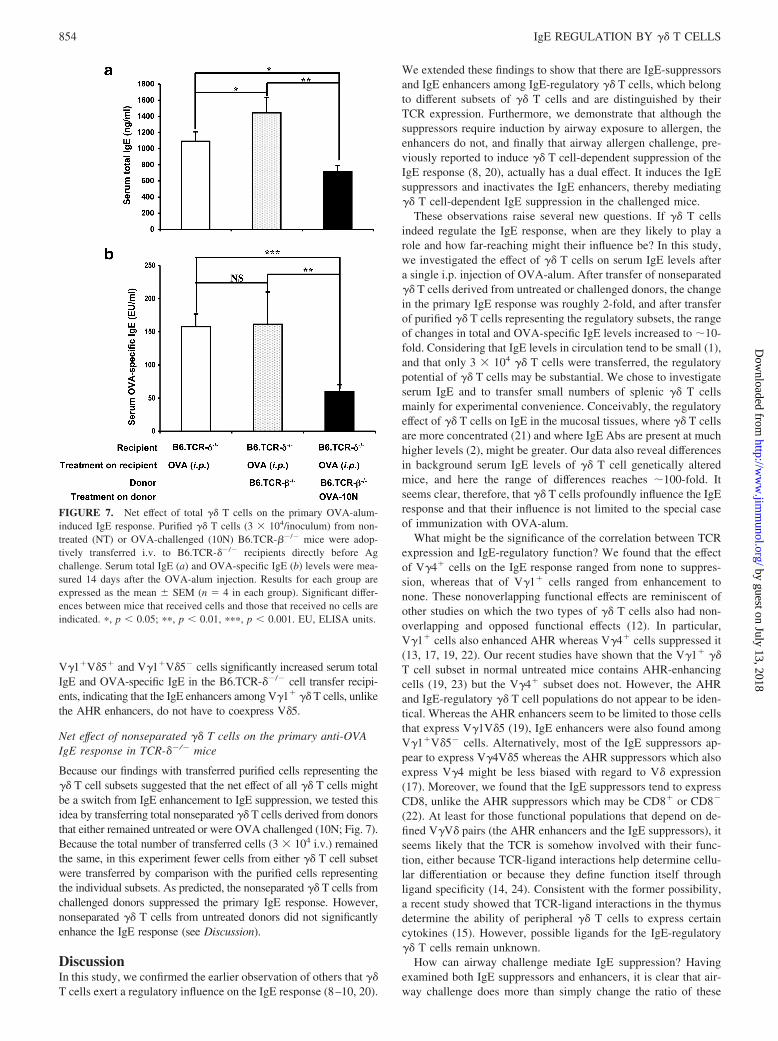
V�1�V�5� and V�1�V�5� cells significantly increased serum totalIgE and OVA-specific IgE in the B6.TCR-��/� cell transfer recipi-ents, indicating that the IgE enhancers among V�1� �� T cells, unlikethe AHR enhancers, do not have to coexpress V�5.
Net effect of nonseparated �� T cells on the primary anti-OVAIgE response in TCR-��/� mice
Because our findings with transferred purified cells representing the�� T cell subsets suggested that the net effect of all �� T cells mightbe a switch from IgE enhancement to IgE suppression, we tested thisidea by transferring total nonseparated �� T cells derived from donorsthat either remained untreated or were OVA challenged (10N; Fig. 7).Because the total number of transferred cells (3 � 104 i.v.) remainedthe same, in this experiment fewer cells from either �� T cell subsetwere transferred by comparison with the purified cells representingthe individual subsets. As predicted, the nonseparated �� T cells fromchallenged donors suppressed the primary IgE response. However,nonseparated �� T cells from untreated donors did not significantlyenhance the IgE response (see Discussion).
DiscussionIn this study, we confirmed the earlier observation of others that ��T cells exert a regulatory influence on the IgE response (8–10, 20).
We extended these findings to show that there are IgE-suppressorsand IgE enhancers among IgE-regulatory �� T cells, which belongto different subsets of �� T cells and are distinguished by theirTCR expression. Furthermore, we demonstrate that although thesuppressors require induction by airway exposure to allergen, theenhancers do not, and finally that airway allergen challenge, pre-viously reported to induce �� T cell-dependent suppression of theIgE response (8, 20), actually has a dual effect. It induces the IgEsuppressors and inactivates the IgE enhancers, thereby mediating�� T cell-dependent IgE suppression in the challenged mice.
These observations raise several new questions. If �� T cellsindeed regulate the IgE response, when are they likely to play arole and how far-reaching might their influence be? In this study,we investigated the effect of �� T cells on serum IgE levels aftera single i.p. injection of OVA-alum. After transfer of nonseparated�� T cells derived from untreated or challenged donors, the changein the primary IgE response was roughly 2-fold, and after transferof purified �� T cells representing the regulatory subsets, the rangeof changes in total and OVA-specific IgE levels increased to �10-fold. Considering that IgE levels in circulation tend to be small (1),and that only 3 � 104 �� T cells were transferred, the regulatorypotential of �� T cells may be substantial. We chose to investigateserum IgE and to transfer small numbers of splenic �� T cellsmainly for experimental convenience. Conceivably, the regulatoryeffect of �� T cells on IgE in the mucosal tissues, where �� T cellsare more concentrated (21) and where IgE Abs are present at muchhigher levels (2), might be greater. Our data also reveal differencesin background serum IgE levels of �� T cell genetically alteredmice, and here the range of differences reaches �100-fold. Itseems clear, therefore, that �� T cells profoundly influence the IgEresponse and that their influence is not limited to the special caseof immunization with OVA-alum.
What might be the significance of the correlation between TCRexpression and IgE-regulatory function? We found that the effectof V�4� cells on the IgE response ranged from none to suppres-sion, whereas that of V�1� cells ranged from enhancement tonone. These nonoverlapping functional effects are reminiscent ofother studies on which the two types of �� T cells also had non-overlapping and opposed functional effects (12). In particular,V�1� cells also enhanced AHR whereas V�4� cells suppressed it(13, 17, 19, 22). Our recent studies have shown that the V�1� ��T cell subset in normal untreated mice contains AHR-enhancingcells (19, 23) but the V�4� subset does not. However, the AHRand IgE-regulatory �� T cell populations do not appear to be iden-tical. Whereas the AHR enhancers seem to be limited to those cellsthat express V�1V�5 (19), IgE enhancers were also found amongV�1�V�5� cells. Alternatively, most of the IgE suppressors ap-pear to express V�4V�5 whereas the AHR suppressors which alsoexpress V�4 might be less biased with regard to V� expression(17). Moreover, we found that the IgE suppressors tend to expressCD8, unlike the AHR suppressors which may be CD8� or CD8�
(22). At least for those functional populations that depend on de-fined V�V� pairs (the AHR enhancers and the IgE suppressors), itseems likely that the TCR is somehow involved with their func-tion, either because TCR-ligand interactions help determine cellu-lar differentiation or because they define function itself throughligand specificity (14, 24). Consistent with the former possibility,a recent study showed that TCR-ligand interactions in the thymusdetermine the ability of peripheral �� T cells to express certaincytokines (15). However, possible ligands for the IgE-regulatory�� T cells remain unknown.
How can airway challenge mediate IgE suppression? Havingexamined both IgE suppressors and enhancers, it is clear that air-way challenge does more than simply change the ratio of these
FIGURE 7. Net effect of total �� T cells on the primary OVA-alum-induced IgE response. Purified �� T cells (3 � 104/inoculum) from non-treated (NT) or OVA-challenged (10N) B6.TCR-��/� mice were adop-tively transferred i.v. to B6.TCR-��/� recipients directly before Agchallenge. Serum total IgE (a) and OVA-specific IgE (b) levels were mea-sured 14 days after the OVA-alum injection. Results for each group areexpressed as the mean � SEM (n � 4 in each group). Significant differ-ences between mice that received cells and those that received no cells areindicated. �, p � 0.05; ��, p � 0.01, ���, p � 0.001. EU, ELISA units.
854 IgE REGULATION BY �� T CELLS
by guest on July 13, 2018http://w
ww
.jimm
unol.org/D
ownloaded from

cells in favor of the suppressors but rather affects the two regula-tory populations separately. The experiments shown in this paperdo not formally rule out that �� T cells migrate from the chal-lenged lung to the spleen. However, it would be difficult for pul-monary �� T cells to change the composition of splenic �� T cellssubstantially because the splenic population of �� T cells is muchlarger (25, 26). More likely perhaps, �� T cells in the spleen areinfluenced by other signal carriers from the lung, e.g., pulmonarydendritic cells (DC). Our preliminary studies with �� T cells ex-posed to transferred non-T cells from challenged mice are consis-tent with such a mechanism (Y. Huang, unpublished observations).Moreover, we found both V�1� and V�4� �� T cells in the spleenin close proximity to CD8� DC in the splenic periarteriolar lym-phoid sheaths (22), a known destination of the peripheral signalcarriers or shuttles. Furthermore, the splenic �� T cells requiredthe presence of CD8� splenic DCs or their functional developmentin another model (22). Therefore, we envision that the splenic ��T cells might be compelled to change their function under theinfluence of CD8� splenic DC, which must have received signalsfrom the challenged lung (27). Indeed, such a mechanism has al-ready been described, where CD8� DC, which are known to re-main in the lymphoid tissues (28), depend on shuttle cells to re-ceive stimulatory signals and Ags from the peripheral tissuesincluding the lung (29–32).
Despite the clear IgE-enhancing effect of purified V�1� cells de-rived from nonchallenged donors, total �� T cells derived from suchdonors enhanced IgE only weakly (detected in the assay for totalserum IgE, but not for OVA-specific IgE). This simply might be dueto the smaller number of transferred V�1� cells in this experiment(only approximately one-half of total splenic �� T cells are V�1�) orit might be caused by interactions among the IgE-regulatory �� T cellsubsets. In any case, the net effect of �� T cells in untreated micetherefore might be only slightly supportive of the primary IgE re-sponse, whereas their effect in challenged mice is clearly suppressive.
In sum, we found that airway challenge brings about coordi-nated and opposite functional changes of the IgE-enhancing and-suppressive �� T cells. This coordinated change of two antago-nistic cell-types appears to represent an efficient mechanism in theregulation of the primary IgE response.
AcknowledgmentsWe acknowledge the advice and support of Dr. Katja Aviskus and the experthelp of Shirley Sobus and William Townend with cell analysis and sorting.
DisclosuresThe authors have no financial conflict of interest.
References1. Gould, H. J., B. J. Sutton, A. J. Beavil, R. L. Beavil, N. McCloskey, H. A. Coker,
D. Fear, and L. Smurthwaite. 2003. The biology of IgE and the basis of allergicdisease. Annu. Rev. Immunol. 21: 579–628.
2. Gould, H. J., and B. J. Sutton. 2008. IgE in allergy and asthma today. Nat. Rev.Immunol. 8: 205–217.
3. Thornton, C. A., J. A. Holloway, E. J. Popplewell, J. K. Shute, J. Boughton, andJ. O. Warner. 2003. Fetal exposure to intact immunoglobulin E occurs via thegastrointestinal tract. Clin. Exp. Allergy 33: 306–311.
4. Eisenbarth, S. C., O. R. Colegio, W. O’Connor Jr., F. S. Sutterwala, andR. A. Flavell. 2008. Crucial role for the Nalp3 inflammasome in the immunos-timulatory properties of aluminium adjuvants. Nature 453: 1122–1127.
5. Mowat, A. M. 1987. The regulation of immune responses to dietary proteinantigens. Immunol. Today 8: 93–98.
6. Sedgwick, J. D., and P. G. Holt. 1985. Down-regulation of immune responses toinhaled antigen: studis on the mechanism of induced suppression. Immunology56: 635–642.
7. Seymour, B. W. P., L. J. Gershwin, and R. L. Coffman. 1998. Aerosol-inducedimmunoglobulin (Ig)-E unresponsiveness to ovalbumin does not require CD8� or
T cell receptor (TCR)-�/�� T cells or interferon (IFN)-� in a murine model ofallergen sensitization. J. Exp. Med. 187: 721–731.
8. McMenamin, C., C. Pimm, M. McKersey, and P. G. Holt. 1994. Regulation ofIgE responses to inhaled antigen in mice by antigen-specific �� T cells. Science265: 1869–1871.
9. Zuany-Amorim, C., C. Ruffie, S. Haile, B. B. Vargaftig, P. Pereira, andM. Pretolani. 1998. Requirement for �� T cells in allergic airway inflammation.Science 280: 1265–1267.
10. Svenson, L., B. Lilliehook, R. Larsson, and A. Bucht. 2003. �� T cells contributeto the systemic immunoglobulin E response and local B-cell reactivity in allergiceosinophilic airway inflammation. Immunology 108: 98–108.
11. Ferrick, D. A., M. D. Schrenzel, T. Mulvania, B. Hsieh, W. G. Ferlin, and H. Lepper.1995. Differential production of interferon-� and interleukin-4 in response to Th1-and Th2-stimulating pathogens by �� T cells in vivo. Nature 373: 255–257.
12. Huber, S. A., D. Graveline, M. K. Newell, W. K. Born, and R. L. O’Brien. 2000.V�1� T cells suppress and V�4� T cells promote susceptibility to coxsackievirusB3-induced myocarditis in mice. J. Immunol. 165: 4174–4181.
13. Hahn, Y.-S., C. Taube, N. Jin, L. Sharp, J. M. Wands, M. Kemal Aydintug,M. Lahn, S. A. Huber, R. L. O’Brien, E. W. Gelfand, and W. K. Born. 2004.Different potentials of �� T cell subsets in regulating airway responsiveness:V�1� cells, but not V�4� cells, promote airway hyperreactivity: Th2 cytokines,and airway inflammation. J. Immunol. 172: 2894–2902.
14. O’Brien, R. L., C. L. Roark, N. Jin, M. K. Aydintug, J. D. French, J. L. Chain,J. M. Wands, M. Johnston, and W. K. Born. 2007. �� T cell receptors: functionalcorrelations. Immunol. Rev. 215: 77–88.
15. Jensen, K. D. C., X. Su, S. Shin, L. Li, S. Youssef, S. Yamasaki, L. Steinman,T. Saito, R. M. Locksley, M. M. Davis, N. Baumgarth, and Y.-H. Chien. 2008.Thymic selection determines gammadelta T cell effector fate: antigen-naive cellsmake interleukin-17 and antigen-experienced cells make interferon �. Immunity29: 90–100.
16. Heilig, J. S., and S. Tonegawa. 1986. Diversity of murine � genes and expressionin fetal and adult T lymphocytes. Nature 322: 836–840.
17. Jin, N., C. Taube, L. Sharp, Y.-S. Hahn, X. Yin, J. M. Wands, C. L. Roark, R. L.O’Brien, E. W. Gelfand, and W. K. Born. 2005. Mismatched antigen prepares �� Tcells for suppression of airway hyperresponsiveness. J. Immunol. 174: 2671–2679.
18. Lahn, M., A. Kanehiro, K. Takeda, J. Terry, Y.-S. Hahn, M. K. Aydintug,A. Konowal, K. Ikuta, R. L. O’Brien, E. W. Gelfand, and W. K. Born. 2002.MHC class I-dependent V�4� pulmonary T cells regulate �� T cell-independentairway responsiveness. Proc. Natl. Acad. Sci. USA 99: 8850–8855.
19. Jin, N., N. Miyahara, C. L. Roark, J. D. French, M. K. Aydintug, J. L. Matsuda,L. Gapin, R. L. O’Brien, E. W. Gelfand, and W. K. Born. 2007. Airway hyper-responsiveness through synergy of �� T cells and NKT cells. J. Immunol. 179:2961–2968.
20. McMenamin, C., M. McKersey, P. Kuhnlein, T. Hunig, and P. G. Holt. 1995. ��T cells down-regulate primary IgE responses in rats to inhaled soluble proteinantigens. J. Immunol. 154: 4390–4394.
21. Hayday, A., and R. Tigelaar. 2003. Immunoregulation in the tissues by �� T cells.Nat. Rev. Immunol. 3: 233–242.
22. Cook, L., N. Miyahara, N. Jin, J. M. Wands, C. Taube, C. L. Roark, T. A. Potter,E. W. Gelfand, R. L. O’Brien, and W. K. Born. 2008. Evidence that CD8�
dendritic cells enable the development of �� T cells that modulate airway hy-perresponsiveness. J. Immunol. 181: 309–319.
23. Jin, N., C. L. Roark, N. Miyahara, C. Taube, M. K. Aydintug, J. M. Wands,Y. Huang, Y. S. Hahn, E. W. Gelfand, R. L. O’Brien, and W. K. Born. 2009.Allergic airway hyperresponsiveness-enhancing �� T cells develop in normaluntreated mice and fail to produce IL-4/13, unlike Th2 and NKT cells. J. Immu-nol. 182: 2002–2010.
24. O’Brien, R. L., M. Lahn, W. Born, and S. A. Huber. 2001. T cell receptor andfunction cosegregate in �-� T cell subsets. Chem. Immunol. 79: 1–28.
25. Hahn, Y.-S., C. Taube, N. Jin, K. Takeda, J.-W. Park, J. M. Wands,M. K. Aydintug, C. L. Roark, M. Lahn, R. L. O’Brien, E. W. Gelfand, andW. K. Born. 2003. V�4� T cells regulate airway hyperreactivity to methacholinein ovalbumin-sensitized and challenged mice. J. Immunol. 171: 3170–3178.
26. Wands, J. M., C. L. Roark, M. K. Aydintug, N. Jin, Y.-S. Hahn, L. Cook, X. Yin,J. Dalporto, M. Lahn, D. M. Hyde, et al. 2005. Distribution and leukocyte con-tacts of �� T cells in the lung. J. Leukocyte Biol. 78: 1086–1096.
27. Lambrecht, B. N., and H. Hammad. 2003. Taking our breath away: dendritic cellsin the pathogenesis of asthma. Nat. Rev. Immunol. 3: 994–1003.
28. Shortman, K., and S. H. Naik. 2007. Steady-state and inflammatory dendritic celldevelopment. Nat. Rev. Immunol. 7: 19–30.
29. Carbone, F. R., G. T. Belz, and W. R. Heath. 2004. Transfer of antigen betweenmigrating and lymph node-resident DCs in peripheral T-cell tolerance and im-munity. Trends Immunol. 25: 655–658.
30. Allan, R. S., C. M. Smith, G. T. Belz, A. L. van Lint, L. M. Wakim, W. R. Heath,and F. R. Carbone. 2003. Epidermal viral immunity induced by CD8a� dendriticcells but not by Langerhans cells. Science 301: 1925–1928.
31. Mount, A. M., C. M. Smith, F. Kupresanin, K. Stoermer, W. R. Heath, andG. T. Belz. 2008. Multiple dendritic cell populations activate CD4� T cells afterviral stimulation. PLoS ONE 3: 1–10.
32. Belz, G. T., C. M. Smith, L. Kleinert, P. Reading, A. Brooks, K. Shortman,F. R. Carbone, and W. R. Heath. 2004. Distinct migrating and nonmigratingdendritic cell populations are involved in MHC class I-restricted antigen presen-tation after lung infection with virus. Proc. Natl. Acad. Sci. USA 101:8670–8675.
855The Journal of Immunology
by guest on July 13, 2018http://w
ww
.jimm
unol.org/D
ownloaded from