the hla genetic constitution of the bushmen (san)
TRANSCRIPT

The HLA Genetic Constitution of the Bushmen (San)
E. D. du Toit, D. G. Taljaard, J. Marshall, C. Ritchie, and M. Oudshoorn
ABSTRACT: The HLA-A. -B, and -C antigens of 290 and the DR antigens of 212 /Kung San individuak were characterized. The most frequent antigens were HLA-A30 [gene frequency (gf) = 0.I93], Bw58 (gf = 0.303), Cw6 (gf - 0.327), DR4 (gf = 0.273), and DQw3 (gf = 0.553). An unexpected finding was the low frequency of the classic African black antigen Bw42 (gf = 0.004). Marked differences as well as similarities in HLA gene frequencies were observed between the San and the South African Negroes, supporting the view that they had a common origin and were then separated for a very long time. During this period difference., developed as a result of selective advantage in the Negroes following the pastoralist-agriculturalist way of life as opposed to the hunter-gatherer way of life. The picture is further complicated by the fact that gene flow, mostly from the San to the southern African Negroes, took place when they met again a few hundred years ago. The data also illustrate HLA haplotypes, linkage disequilibria, and four-locus haplotypes not previously seen in other human populations. The most frequent four-locus haplotype in the San, HLA-Aw43,Cw7,B7,DRw6 was also different from A30, Cw2,Bw42,DR3, the most common among the South African Negroes.
ABBREVIATIONS af antigen frequency gf gene frequency
I N T R O D U C T I O N
The present-day San, or Bushmen, are probably descended from people of the later Stone Age who were living in southern Africa as long as 10,000 years ago. Based on the occurrence of their rock art and other remnants, the population was distributed over most of this area. The interrelationships of the San, Khoikhoi (Hottentots), and the various Negro populations, and the way these may reflect the history of the African continent, is not completely understood. The skeletal evidence suggests that if Khoisan (the name used to refer to the San and Khoikhoi collectively) and Negroes descended from a single common stock, the divergence had already taken place about 30,000 years ago. This divergence, which took place originally outside the southern cul-de-sac of the continent, led to further local differentiation in southern Africa of the Khoisan stock into the Khoikhoi and San subdivisions, both of which became for a time dominant in southern Africa [1]. As a result of the great southern migration of the Bantu-speaking
From the Provincial Laboratory for Tissue Immunology, Cape Town, South Africa CE.D. du T.," D.G.T.: M.O.) and the Ju/wasi Bushman Development Foundation, Windhoek, Namibia (J.M.: C.R.).
Address reprint requests to Professor E.D. du Toit, Provincial Laboratory for Tissue Immunology, Private Bag 4, Observatory 7935, Cape Town, South Africa.
Received October 30, 1989: accepted February 1 l, 1990.
406 Human Immunology 28, 406-415 (1990) 0198-8859/90/$3.50 © American Society for Histocompatibility and lmmunogenetics, 1990

HLA in the Bushmen (San) 407
Negroes 500 or 600 years ago and the arrival of the Dutch in 1652, the Khoisan have been greatly reduced in number. There are now estimated to be only 55,000 San still surviving.
A number of genetic studies using red cell antigens and electrophoretic and anthropometric data have corroborated the already recognized cultural and lin- guistic diversity of the Negro and Khoisan groups. This existing genetic evidence on the past migrations and ancestral relationships is in some respects at variance with the information provided by linguistic data, which suggests clearer divisions or groups [2]. The extensive polymorphism of HLA and the presence of strong linkage disequilibrium between some of the alleles at the HLA loci suggest that the HLA system may provide valuable additional evidence to clarify these relationships.
The present-day San are one of the last remaining hunter-gatherer groups living in isolation, and they are virtually confined to Botswana, Namibia, and Angola. Traditionally they lived in small independent family units and spoke non-Bantu click languages. The San are usually divided on linguistic and geographic grounds into three principal groups: the northern, the largest group; the southern, the smallest group; and the central. However, according to Jenkins, "Irrespective of which genetic distance measurement is used, the evidence of the serogenetic markers (blood groups, serum proteins, and red cell enzyme systems) is over- whelmingly in favour of the close affinities with one another, of all the San populations," regardless of the language they speak [3].
The !Kung San of Botswana, Namibia, and southern Angola number about 6500 and comprise nearly all the remaining northern Bush language speakers. The !Kung San population studied were from the semidesert region around Tsumkwe, near the northeastern boundary of Namibia with Botswana. The Negro influence has been mainly limited to a patron-client relationship with the San, and when miscegenation did take place, the gene flow was almost invariably from the San to the Negroes.
Limited HLA-A and -B testing was performed on this group of !Kung San in 1971 as part of the Fifth International Histocompatibility Workshop [4]. As the "Negroid" antigens at these loci were not fully defined at the time, the data are therefore of limited value in genetic analyses. We now report on a further study showing more detailed HLA results, including the HLA-C, -DR, and -DQ an- tigens.
MATERIALS AND METHODS
The People
Blood samples were collected from the majority of the families resident at Tsum- kwe and the surrounding region. A total of 290 individuals belonging to 59 nuclear families in 29 extended families were studied. Detailed pedigree informa- tion going back one, or more often two, generations from the HLA-typed individu- als was obtained by two of the authors (J.M. and C.R.), who had been intensively studying the population for a year. The pedigrees were further confirmed by the study of other genetic markers. The gene frequencies were compared with those in the Xhosa, one of the southern African Negro groups [5]. Genetic distances between the San, South African Negroes, Cape Coloureds (a group of mixed ancestry), South African Caucasoids [5], as well as a group of Nigerians studied by Okoye et al. [6] were calculated.

408 E.D. du Toit et al.
HLA Typing
HLA-A, -B, and -C antigens. HLA-A, -B, and -C typing was done according to the standard NIH microlymphocytotoxicity technique [7] modified to use 0.5-/A quantities of serum. Local class I antisera, as well as those obtained by international exchange, were used. HLA antigens A1-Aw74 (with the exception of Aw68 and Aw69), B7-Bw70 (with the exception ofBw54, Bw59, Bw64, Bw65, and Bw67), and Cwl-Cw8 were determined.
HLA-DR and-DQ antigens. The HLA-DR and -DQ typings were performed using the technique agreed upon for the Seventh Workshop [8]. Removal of T cells was done by the nylon wool method [9]- Some of the antisera used to define the class II antigens were of local origin and others were obtained by exchange with colleagues. The antigens DR1-DRwl0 and DQwl -DQw3 were determined.
Statistical Analysis
Since a comparison of the gene frequencies of the whole group and a subgroup of unrelated individuals showed no significant differences, only the results of the whole group are reported. Gene frequencies (gf) were calculated from the antigen frequencies (af) by Bernstein's formula: gf = 1 - k/1 - af. The frequency of the "blank" gene(s) was obtained by subtracting the sum total of defined gene frequencies from 1. The gene frequencies in the San and South African Negroes were compared using the X-' test and Yates correction. Adjustment for multiple comparisons was made by multiplying the p value by the number of specificities (70) studied (Pc).
Delta values (A) and haplotype frequencies were computed from 2 × 2 tables of the phenotype frequencies using the formula of Mattiuz et al. [10], and their significance was evaluated by the X 2 test. Since nearly all individuals could be haplotyped by segregation analysis of HLA in families, haplotype frequencies were also obtained by direct counting. The latter haplotypes were also used to identify the most common four-locus haplotypes.
Genetic distances were calculated by the method of Cavalli-Sforza and Bodmer [11].
RESULTS
HLA-A, -B, and -C antigens were identified in 290 individuals, and of these 212 were HLA-DR typed. A small subset of 20 random individuals was also typed for the DQ antigens. Tables 1 and 2 show the gene frequency distribution in the San for the HLA-A, -B, -C, and -DR antigens. Equivalent data from a previous study in the South African Negroes (Xhosa) is also shown for comparison [5].
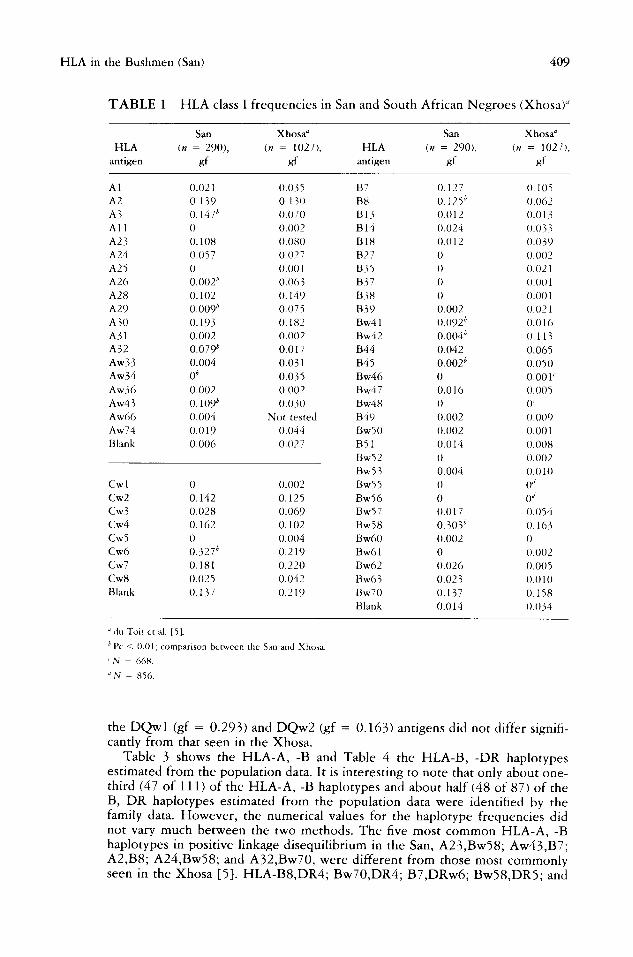
Only three HLA-A antigens and several B-locus antigens were absent in the San (Table 1). Of the A-locus antigens, A2, A3, A23, A28, A30, and Aw43 were the most common (gf > 0.10). At the B locus only B7, B8, Bw58, and Bw70 were seen with gf > 0.10. The classic southern African Negro HLA-B antigens, Bw58 (gf = 0.163) and Bw70 (gf = 0.158), were also very common in the San (gf = 0.303 and gf = 0.137, respectively) (Table 1). HLA-Cwl and -Cw5 were absent in the San and very rare among the South African Negroes. HLA-Cw6 was the most common C-locus antigen in the San and one of the two most common in the Xhosa. All the DR (Table 2) and DQ antigens tested were present in both the San and Xhosa. In the San HLA-DR1, -DR4, -DRw6, and -DQw3 (gf = 0.553) were the most commonly observed class II antigens. The distribution of
HLA in the Bushmen (San) 409
TABLE 1 HLA class I frequencies in San and South African Negroes (XhosaY
San Xhosa ~ San Xhosa ~
HLA (n = 290), (n = 1027), HLA (n = 290), (n = 1027), antigen gf gf antigen gf gf
A l 0.021 0.035 B7 0.127 0.105 A2 0.139 0.130 B8 0.125 t' 0.062 A3 0.147 b 0.070 B 13 0.012 0.01 A l l 0 0.002 B 14 0.024 0.033 A23 0.108 0.080 B l 8 0.012 0.039 A24 0.057 0.027 B27 0 0.002 A 25 0 0.001 B 35 0 (). 021 A26 0.002 tj 0.063 B37 0 0.001 A28 0.102 0.149 B38 0 0.001 A29 0.009 b 0.075 B39 0.002 0.021 A30 0.193 0.182 Bw41 0.()92 ~ 0 .016 A31 0.002 0.002 Bw42 0.()04 ~' 0.113 A32 0.079 b 0.017 B44 0.042 0.065 Aw33 0.004 0.031 B45 0.002 b 0.050 Aw34 0 b 0.035 Bw46 0 0.001' Aw36 0.002 0.002 Bw47 0.016 0.005 Aw43 0.109 b 0.030 Bw48 0 0' Aw66 0.004 Not tested B49 0.002 0.009 Aw74 0.019 0.044 BwS0 0.002 0.001 Blank 0.006 0.027 B51 0.014 0.008
Bw52 0 0.002 Bw53 0.004 0.010
Cw I 0 0.002 Bw55 0 0 '/ Cw2 0.142 0 .125 Bw56 0 0 ~ Cw3 0.028 0.069 Bw57 0.017 0.054 Cw4 0.162 0.102 Bw58 0.303 b 0.163 Cw5 0 0.004 Bw60 0.002 0 Cw6 0.327 b 0.219 Bw61 0 0.002 Cw7 0.181 0 . 2 2 0 Bw62 0.026 0.005 Cw8 0.025 0.042 Bw63 0.023 0.010 Blank 0 .137 0.2 l 9 Bw70 0.137 O. 158
Blank 0.014 0.034
du Toit et ai. [5].
b Pc < 0.01; comparison between the San and Xhosa.
' N 668.
JN 856.
the DQwl (gf = 0.293) and DQw2 (gf = 0.163) antigens did not differ signifi- cantly from that seen in the Xhosa.
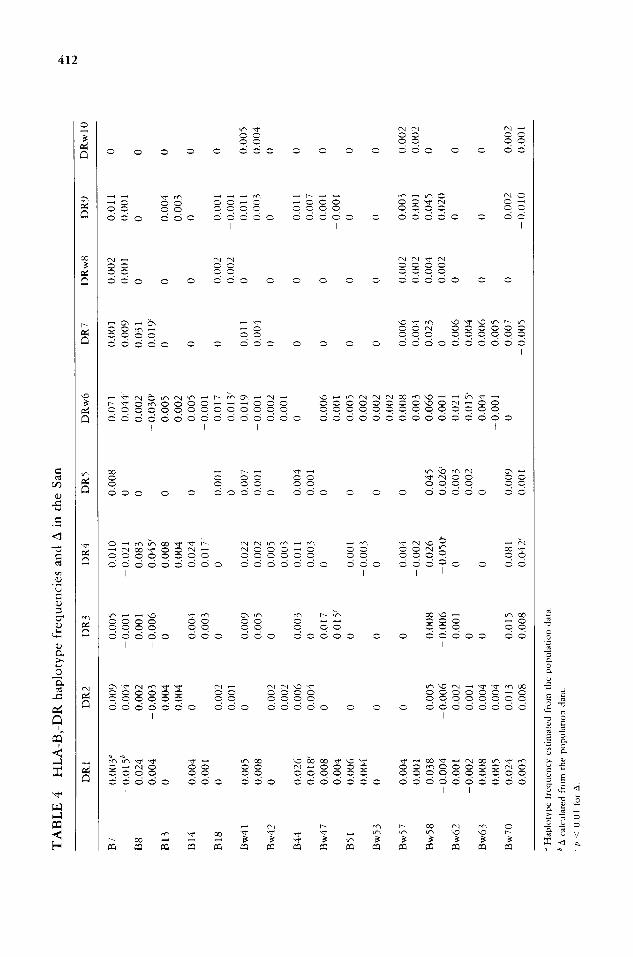
Table 3 shows the HLA-A, -B and Table 4 the HLA-B, -DR haplotypes estimated from the population data. It is interesting to note that only about one- third (47 of 111) of the HLA-A, -B haplotypes and about half (48 of 87) of the B, DR haplotypes estimated from the population data were identified by the family data. However, the numerical values for the haplotype frequencies did not vary much between the two methods. The five most common HLA-A, -B haplotypes in positive linkage disequilibrium in the San, A23,Bw58; Aw43,B7; A2,B8; A24,Bw58; and A32,Bw70, were different from those most commonly seen in the Xhosa [5]. HLA-B8,DR4; Bw70,DR4; B7,DRw6; Bw58,DR5; and

410 E.D. du Toit et al.
TABLE 2 Gene frequencies of HLA class II specificities in the San and South African Negroes (XhosaY
San Xhosa a
HLA (n 212), (n 322),
antigens gf gf
DR1 0.148 b 0.043
DR2 0.039 ~ 0.115 DR3 0.048 b 0.238
DR4 0.273 b 0.069
DR5 0.068 b 0. 172
DRw6 0.232 0.181
DR7 0.08 l 0.074
DRw8 0.007 0.006 DR9 0.089 b 0.008
D R w l 0 0.005 0.028
Blank 0.010 0.066
(n = 110) (n = 245)
DRw52 0.333 b 0.633
DRw53 0.496 b 0.150
du Toit et al. [5].
b Pc < 0.01, comparison between San and Xhosa.
B8,DR7 were the most frequent HLA-B, -DR haplotypes in positive linkage disequilibrium in the San. These were also different from those occurring com- monly in the Xhosa.
The two most common HLA-B, -C haplotypes, Bw58,Cw6 and B7,Cw7, in the San were also the commonest in the Xhosa. Other common B,C haplotypes in the San were B8,Cw2; Bw70,Cw4; and B 14,Cw8. The five most common HLA- A, -C haplotypes in linkage disequilibrium in the San were A23,Cw6; A3,Cw6; A30,Cw4; A2,Cw2; and Aw43,Cw7. Only A3,Cw6 was also commonly seen in the South African Negroes. Table 5 shows the six most common HLA-A, -B, -C, and -DR haplotypes seen in the San families studied.
Based on the genetic distance values, as shown in Table 6, the San appeared to be closest to the South African Negroes and furthest away from the Nigerians.
DISCUSSION
When the HLA-A, -B, -C, and -DR frequencies of the San and the Xhosa were compared with the South African Caucasoids, a typical "southern African" pattern emerged [12]. This is best exemplified by the high frequencies of A30, Aw43, Bw58, Bw70, Cw2, Cw6, and DRw6 and the virtual or complete absence of A 11, B27, Cwl, Cw5, and DRw8. Most of the so-called classic African Negro antigens reached their highest frquencies in the San, thus supporting the suggestion [ 1,13] that the San share a common ancestry with, and may be even more "African" than, the Negroes. These findings are also in agreement with Zoutendyk's work which, according to Mourant, showed that while the Khoisan differed from the Negroes, the two population groups represented a variation on a well-defined African pattern totally distinct from those of other major races [13].
On the other hand, the San must have evolved for a considerable time in relative isolation, accounting for the differences in HLA frequencies that existed

TA
BL
E 3
H
LA
-A,-
B h
aplo
type
fre
quen
cies
and
A v
alue
s in
the
San
A1
A2
A3
A23
A
24
A
26
A
28
A
29
A
30
A
31
A32
A
w3
3
Aw
36
A
w4
3
Aw
66
A
w7
4
B7
0 0
.02
9 ~
0.00
1 0
.00
2
0.0
06
0
.00
2
0.0
10
0.
001
0.0
12
b -0
.01
8
-0.0
11
-0
.00
1
0.0
02
-0
.00
3
0 B
8 0
0.06
1 0
.00
8
0 0
0 0.
0.30
0
0.0
43
' -0
.01
1
0.0
17
' B
13
0 0
0 0
.00
2
0.00
1 0
0 0
0.00
1 0
B14
0.
003
0.0
06
0
0 0
0 0
0 0.
003
0.0
03
B
l 8
0 0
0.0
08
0
0 0
0.00
1 0
0.0
06
-
0.0
0 l
B3
9
0 0
0.0
02
0
0 0
0 0
0.0
02
B
w41
0
0 0
.01
2
0.0
13
0
.00
6
0 0
.03
7
0 -0
.00
2
0.0
03
0
.00
1
0.0
27
' B
w4
2
0 0
0 0
0 0
0 0
B44
0
.01
9
0.0
08
0
0 0.
01/3
0
0 0
0.01
8'
0.0
02
0.
001
B45
0
0 0
/).0
02
0 0
0 0
0.0
02
B
w4
7
0 0
.00
6
0.0
16
0
0 0
0 0
0.0
03
0
.01
3'
B4
9
0 0
0.0
02
0
0 0
0 0
0.0
02
B
wS
O
0 0
0 0
.00
2
0 0
0.0
02
0
0.0
02
0
.00
2
B51
0
0 0
.00
2
0 0
0 0
.01
4
0 -0
.00
1
0.0
13
' B
w5
3
0 0
.00
4
0.00
1 0
0 0
0 0
0.0
03
0.
001
Bw
57
0.
001
0.0
07
0.
001
0.0
02
0
.00
3
0 0
.00
2
0 0.
001
0.0
05
-
0.0
02
0
0.0
02
0
Bw
58
0.
001
0.0
14
0
.05
7
0.0
77
0
.04
7
0.0
02
0
.00
6
0.0
04
0
.00
6
0.0
28
0
.01
3
0.0
45
' 0
.03
0'
0.00
1 -0
.02
5'
0.0
01
B
w6
0
0 0
.00
2
0.0
02
0
0 0
0 0
0.0
02
0
.00
2
Bw
62
0
0 0
.00
2
0.0
01
0
0 0
.00
3
0.0
02
-0
.00
2
-0.0
02
0
0.0
01
B
w6
3
0.00
1 0
.00
7
0.0
06
0
.00
7
0.0
04
0
0 0
0.0
0l
0.00
.3
0.0
03
0
.00
5
0.0
03
B
w7
0
0.00
1 0
0.03
1 0
.00
1
0 0
0.0
16
0
.00
3
0.00
2
0.0
15
0
0 0
0.0
02
0
.06
5
0 0
.00
1
-0.0
10
11
.002
0
.05
1'
0.0
01
0.
022
(/.0
02
0.0
20
0
0 0
.00
2
0 0
.00
5
-0.0
02
0
.00
2
0.0
10
0
.01
1
0.0
03
0
.01
2
0 0
0 0
0 0
.00
2
0 0
.01
0
0.0
02
0
.00
5
0 0
.02
4
0 0
0 0
0 0
0.0
23
' 0
.00
4
0 0
0 0
0 0
0 0
.00
1
0.0
02
0
0 0
0 0
0 0
0.0
01
0
.01
6
1)
0.0
08
0
0 0
.00
1
0 0
- 0
.00
2
0.00
1 0
.01
0
0.0
04
0
.00
2
0 0
0 0
0 0
0.0
03
0
.00
2
0.0
07
0
0.0
12
0
0 0
.00
1
0 0
- 0.
001
0.0
09
' 0
.00
4
0 0
0 0
0 0
0 0
.00
2
0.0
02
0
0 0.
001
0 0
0.0
02
0
0 -0
.00
1
0 0
0 0
.00
2
0 0
0 0
0 0
.00
2
0 0
0 0
0 0
0 0
0.00
1 0
0 0
0 (/
.004
0
0 -
0.0
02
0
.00
3
0 0
0 11
0
0.0
02
0
0 1/
.001
0
.00
2
0 0
0 0
0.0
02
0
0 -0
.00
1
0 0
.07
7
0 0
0.0
04
0
.00
2
0.01
1 0.
001
0.0
04
0
.01
8
0.0
02
0.
001
0.0
22
0
- 0
.00
2
0 0
0 0
0 0
0 0
0.0
20
0
0 0
0 0
.00
1
0 0
0.0
15
' -0
.00
2
0.0
05
0
0 0
0 0
0 0
0.0
01
0
.01
2
0 0
.03
8
0.0
04
0
0.0
28
0
.00
2
0.0
13
0
.02
7'
0.0
03
0
.01
3
0.00
1 0
.01
1'
0.01
1 -0
.01
4
0.0
02
0
.00
2
--0
.01
4
" H
aplo
typ
e fr
equ
ency
est
imat
ed f
rom
the
po
pu
lati
on
dat
a.
b A
cal
cula
ted
fro
m t
he p
op
ula
tio
n d
ata.
'p
< 0.
01 S
igni
fica
nce
leve
ls f
or A
. ,g
-,,
TA
BL
E
4 H
LA
-B,-
DR
h
aplo
typ
e fr
equ
enci
es a
nd A
in
the
San
..,
bO
DR
1
DR
2
DR
3
DR
4
DR
5
DR
w6
D
R7
D
Rw
8
DR
9
DR
w 1
0
B7
0
.00
3"
0.0
09
0
.00
5
0.0
10
0
.00
8
0.0
71
0
.00
1
0.0
02
0
.01
1
0
- 0
.01
5 h
0
.00
4
- 0
.00
1
- 0
.02
1
0 0
.04
4'
0.0
09
0
.00
1
0.0
01
B8
0
.02
4
0.0
02
0
.00
1
0.0
83
0
0.0
02
0
.03
1
0 0
0
0.0
04
-
0.0
03
0
.00
6
0.0
45
c
- 0
.03
0 ~
0
.01
9 c
B 1
3 0
0.0
04
0
0.0
08
0
0.0
05
0
0 0
.00
4
0
0.0
04
0
.00
4
0.0
02
0
.00
3
B 1
4 0
.00
4
0 0
.00
4
0.0
24
0
0.0
05
0
0 0
0
0.0
01
0
.00
3
0.0
17
' -
0.0
01
B 1
8 0
0.0
02
0
0 0
.00
1
0.0
17
0
0.0
02
0
.00
1
0
0.0
01
0
0.0
13
~
0.0
02
0
.00
1
Bw
41
0
.00
5
0 0
.00
9
0.0
22
0
.00
7
0.0
19
0
.01
1
0 0
.01
1
0.0
05
0.0
08
0
.00
5
- 0
.00
2
0.0
01
-
0.0
01
0
.00
4
0.0
03
0
.00
4
Bw
42
0
0.0
02
0
0.0
05
0
0.0
02
0
0 0
0
0.0
02
0
.00
3
0.0
01
B4
4
0.0
26
0
.00
6
0.0
03
0
.01
1
0.0
04
0
0 0
0.0
11
0
0.0
18
' 0
.00
4
0 0
.00
3
0.0
01
0
.00
7
Bw
47
0
.00
8
0 0
.01
7
0 0
0.0
06
0
0 0
.00
1
0
0.0
04
0
.01
5'
0.0
01
-
0.0
01
B51
0
.00
6
0 0
0.0
01
0
0.0
05
0
0 0
0
0.0
04
-
0.0
03
0
.00
2
Bw
53
0
0 0
0 0
0.0
02
0
0 0
0
0.0
02
Bw
57
0.0
04
0
0 0
.00
4
0 0
.00
8
~). 0
06
0
.00
2
0.0
03
0
.00
2
0.0
01
0
.00
2
0.0
03
0
.00
4
0.0
02
0
.00
1
0.0
02
Bw
58
0
.03
8
0.0
05
0
.00
8
0.0
26
0
.04
5
0.0
66
0
.02
3
0.0
04
0
.04
5
0
- 0
.00
4
- 0
.00
6
- 0
.00
6
- 0
.05
0 ~
0
.02
6 '
0.0
01
0
0.0
02
0
.02
0
Bw
62
0
.00
1
0.0
02
0
.00
1
0 0
.00
3
0.0
21
0
.00
6
0 0
0
- 0
.00
2
0.0
01
0
0.0
02
0
.01
5'
0.0
04
Bw
63
0
.00
8
0.0
04
0
0 0
0.0
04
0
.00
6
0 0
0
0.0
05
0
.00
4
-0.0
01
0
.00
5
Bw
T0
0
.02
4
0.0
13
0
.01
5
0.0
81
0
.00
9
0 0
.00
7
0 0
.00
2
0.0
02
0.0
03
0
.00
8
0.0
08
0
.04
2'
- 0
.00
1
- 0
.00
5
- 0
.01
0
0.0
01
Hap
loty
pe f
requ
ency
est
imat
ed f
rom
the
pop
ulat
ion
data
.
b A
cal
cula
ted
from
the
pop
ulat
ion
data
.
'p <
0.
01 f
or-~
.

HLA in the Bushmen (San) 413
T A B L E 5 The six most common HLA-A,-B,-C,-DR haplotypes and their frequencies derived from the San family data
Total number
of haplotypes Frequency A C B DR (n - 360) (%)
w43 w7 7 w6 15 4.2 ~
24 w6 w58 9 13 3.6
23 w6 w58 5 12 3.3
2 w2 8 4 11 3.1
3 w6 w58 w6 10 2.8
32 w8 14 4 9 2.5
" Expressed as a percentage of the total number of haph)types.
between them and the Negroes. Some of the principal HLA features distinguish- ing the San from the Xhosa were the virtual absence of the classic South African Negro antigen Bw42 and the considerably lower frequency of HLA-DR2, -DR3, -DRS, as well as -DRw52 in the San. In addition, HLA-Aw43, -B8, -Bw41, -Bw58, -Cw6, -DR1, -DR4, and -DRw53 were significantly more frequent in the San compared to the Xhosa (Tables 1 and 2). The other striking difference between the San and the Xhosa was seen in the alleles in linkage disequilibrium. The most common HLA-A, -B and B,DR haplotypes in the two groups were different and the six most common four-locus haplotypes in the South African Negroes [ 14], A30,Cw2,Bw42,DR3; A3,Cw6,Bw58,DR3; A29,Cw7,B44,DR5; A2,Cw-,B45,DRw6; A30,Cw6,Bw58,DRS; and A2,Cw7,Bw58,DR5, were en- tirely different from those seen in the San (Table 5). If these haplotypes are primarily due to natural selection, then the data support the suggestion that the two groups have been separated for a very long time.
One of the classic examples of an HLA-A variant apparently confined to southern Africa is HLA-Aw43, originally defined by us in the San [15]. In the present study we found the frequency of Aw43 to be much higher (gf = 0.109) than reported in our earlier studies (gf = 0.050) in the same group of San [15]. This was probably due to a better definition of the antigen with an operationally monospecific antiserum. Jenkins and coworkers suggested that, based on the distribution of the Gm I'1~ haplotype in the various Bantu-speaking chiefdoms, the Xhosa have had the greatest contact with the San [16]. However, from the presence of the classic San antigen HLA-Aw43 at a high frequency in several of the other Negro chiefdoms, i.e., Sotho, Tswana, Zulu (authors' unpublished data), as well as the Xhosa, it follows that all these groups may have had close contact with the San. On the other hand, the presence of Aw43 in all southern African Negroes tested may support the idea of a common stock for the Khoisan and southern African Negroes.
It is interesting to note that A3, B7, and B8, regarded by some as predominantly European Caucasoid antigens [17], are all present at high frequencies in the San, and the frequency of HLA-B8 is even higher than in Caucasoids [5]. It is noteworthy that HLA-B27 was absent in the San and occurred with a low fre- quency in all the other southern African Bantu-speaking groups. There have to our knowledge not been any cases of ankylosing spondylitis reported among the San, and very few have been found among the southern African Negroes [5].
Although the San were closest to the Xhosa based on the genetic distance values, they were, nevertheless, numerically separated by about half (0.024) the

414 E.D. du Toit et al.
T A B L E 6 Genetic distances were calculated from the gene frequencies for all the antigens at the HLA-A,-B,-C and -DR loci for the San, South African Negroes, a Cape Coloureds, a South African Caucasoids, a and Nigerians b
South African San South African Negro Nigerians Cape Coloured Caucasoid
San 0 South African
Negro 0.024 0
Nigerians 0.052 0.025
Cape Coloured 0.030 0.012 South African
Caucasoid 0.045 0.033
0
0.031 0
0.047 0.010
o du Toit et al. [5].
b Okoye et al. [61.
distance usually seen between two major populations. This was in spite of shared "African" genes and of the gene flow that has occurred between the San and Xhosa.
The HLA data presented emphasize the uniqueness of the San people. There is a strong possibility that in the near future the San will no longer be recognizable as a unique entity, and it will be important to extend the present study using new D N A technology. If possible, such studies should also be per formed on other groups of San in order to clarify the linguistic and geographic divisions.
ACKNOWLEDGMENTS
This study was done in collaboration with the MRC Human Ecogenetics Research Unit and the Department of Haematology, School of Pathology, South African Institute for Medical Research and the University of the Witwatersrand. We would also like to thank the San people of Tsumkwe for their kind cooperation, without which this study would have been impossible.
R E F E R E N C E S
1. Nurse GT, Weiner JS, Jenkins T: Research Monographs on Human Population Biol- ogy, No 3: The Peoples of Southern Africa and Their Affinities. Oxford, Clarendon Press, 1985.
2. Westphal EOJ: The linguistic prehistory of Southern Africa: Bush, Kwadi, Hottentot, and Bantu linguistic relationships. Africa 33:237, 1963.
3. Jenkins T: The San and Khoikhoi as recorded in their blood. In Vossen R, Keuthmann K (eds): Contemporary Studies on Khoisan 2. Hamburg, Helmuth Buske Verlag, 1986, p 58.
4. Botha MC, du Toit ED, Jenkins T, van Leeuwen A, D'Amaro J, Meera Khan P, van der Steen G, van Rood JJ, van der DoesJA: The HL-A system in Bushman (San) and Hottentot (Khoikoi) populations of South West Africa. In Dausset J, Colombani J (eds): Histocompatibility Testing 1972. Copenhagen, Munksgaard, 1973, p 421.
5. du Toit ED, MacGregor KJ, Taljaard DG,.Oudshoorn M: HLA-A,B,C,DR, and DQ

HLA in the Bushmen (San) 415
polymorphisms in three South African population groups: South African Negroes, Cape Coloureds, and South African Caucasoids. Tissue Antigens 31:109, 1988.
6. Okoye RC, Williams E, Alonso A, Doyle P, Awad J, Navarrette C, Jaraquemada D, Oilier WER, Festenstein H: HLA polymorphisms in Nigerians. Tissue Antigens 25:142, 1985.
7. Terasaki PI, McClellandJ, Park MS, McCurdy B: Microdroplet lymphocyte cytotoxic- ity test. In Ray JG, Hare DB, Pedersen PD, Kayoe DE (eds): Manual of Tissue Typing Techniques. Bethesda, NIH, 1974.
8. Bodmer JG, Pickbourne P, Richards S: Joint report Ia serology. In Bodmer WF, Batchelor JR, BodmerJG, Festenstein H, Morris PJ (eds): Histocompatibility Testing 1977. Copenhagen, Munksgaard, 1977, p 35.
9. DanilovsJA, Ayoub G, Terasaki PI: B lymphocyte isolation by thrombin-nylon wool. In Terasaki PI (ed): Histocompatibility Testing 1980. Los Angeles, UCLA Press, 1980, p 287.
10. Mattiuz PL, lhde D, Piazza A, Ceppellini R, Bodmer WF: New approaches to the population genetic and segregation analysis of the HL-A system. In Terasaki PI (ed): Histocompatibility Testing 1970. Copenhagen, Munksgaard, 1970, p 193.
11. Cavalli-Sforza LL, Bodmer WF: The genetics of human populations. San Francisco, WH Freeman, 1971, p 704.
12. du Toit ED: HLA polymorphisms in southern African populations. In Mayr WR (ed): Advances in Forensic Haemogenetics 2. Berlin, Heidelberg, Springer-Verlag, 1988, p42.
13. Mourant AE: Blood Relations: Blood Groups and Anthropology. New York, Oxford University Press, 1985.
14. Oudshoorn M: Investigations into the complexity and polymorphism of HLA-D loci in South Africa, PhD thesis, University of Cape Town, 1989.
15. Nurse GT, BodmerJG, Bodmer WF, van Leeuwen A, van Rood JJ, du Toit ED, Botha MC: A reassessment of the HL-A system in Khoisan populations of South West Africa. Tissue Antigens 5:402, 1975.
16. Jenkins T, Zoutendyk A, Steinberg AG: Gammaglobulin groups (Gm and Inv) of various Southern African populations. Am J Phys Anthrop 32:197, 1970.
17. Kostyu D, Dawson D, Ciftan E, Stewart A, Lewis D, Parc F, Laigret J, McCollum R, Amos B: HLA in two islands of French Polynesia. Tissue Antigens 23:217, 1984.