the effect of cations on the specific activities of pyruvate kinase and glucose-6-phosphate...
TRANSCRIPT

Comp. Biochem. PhysioL Vol. 99B. No. 2, pp. 259-264, 1991 0305-0491/91 $3.00 + 0.00 Printed in Great Britain © 1991 Pergamon Press pie
THE EFFECT OF CATIONS ON THE SPECIFIC ACTIVITIES OF PYRUVATE KINASE AND
GLUCOSE-6-PHOSPHATE DEHYDROGENASE OF LUIDIA CLATHRATA (SAY) (ECHINODERMATA: ASTEROIDEA)
JAY R. LEVERONE,*'~ CARL A. LURER* and JOHN M. LAWRENCE'~ *Mote Marine Laboratory, 1600 Thompson Parkway, Sarasota, FL 34236, USA (Tel: 813 388 4441;
Fax: 813 388 4312); and l'Department of Biology, University of South Florida, Tampa, FL 33620, USA (Tel: 813 974 2549)
(Received 1 November 1990)
Abstraet--l. The combined effect of cations (Na ÷, K +, Ca 2+ and Mg 2+ ) on the specific activities of pyruvate kinase (PK) and glucose-6-phosphate dehydrogenase (G-6-PDH) were examined in the pyloric caeca, gonads and tube feet of adult Luidia clathrata.
2. Cation concentrations were based on previously determined intracellular values from individuals that had been exposed to 15, 25 and 35%0 salinity for 1 week.
3. Increases in cationic strength resulted in decreases in specific activity for both enzymes in all tissues. As cationic strength increased, decreases in specific activity were greater for G-6-PDH than PK under similar assay conditions.
4. The relative sensitivity of PK to changes in cation concentration may ensure the stability of the enzyme's catalytic and regulatory functions, while the greater relative sensitivity of G-6-PDH to cations could alter the organism's anabolic capacity.
INTRODUCTION
The asteroid Luidia clathrata is one of several echino- derm species able to inhabit waters of fluctuating salinity (for review see Stickle and Diehl, 1987). During salinity changes, L. clathrata controls cell volume by isosmotic intracellular regulation (Diehl and Lawrence, 1984, 1985), which involves coordi- nated changes in the concentrations of both organic and inorganic osmolytes. As a result, the organism maintains an altered composition and concentration of intracellular solutes as it adjusts to the new salinity.
While the major organic osmolytes are generally compatible with proper enzyme function and struc- ture (Bowlus and Somero, 1979), inorganic ions affect enzyme activity by several mechanisms (Sarkissian and Boatwright, 1974; Clark and Zounes, 1977; Yancey et al., 1982). Altered activities of intermedi- ary metabolic enzymes resulting from changes in ionic concentrations have implications for the func- tioning of catabolic pathways, micromolecular bio- synthesis, and the maintenance of homeostatic mechanisms. The effect of physiological concen- trations of ions on aspects of intermediary metab- olism has received little attention in echinoderms (Ellington, 1982; Marsh and Lawrence, 1985). As the intraceilular ionic composition of cells of L. clathrata has been determined (Diehl and Lawrence, 1984, 1985), an unusual opportunity is provided to ascer- tain the effects of physiological concentrations of ions on enzymatic activity.
Pyruvate kinase, an enzyme involved in the pro- duction of ATP, is a key regulatory enzyme in glycolysis (Mustafa and Hochachka, 1971; Living-
stone, 1975). Glucose-6-phosphate dehydrogenase catalyzes the first step of an alternative route of glucose utilization, the pentose phosphate pathway, that results in the generation of N A D P H for bio- synthetic reactions and the production of pentoses for biosynthesis of nucleotides and nucleosides (Eggleston and Krebs, 1974). This study examines the effects of simulated intracellular cation concen- trations on the specific activities of pyruvate kinase and glucose-6-phosphate dehydrogenase in the pyloric caeca, gonads and tube feet of adult Luidia clathrata.
MATERIALS AND METHODS
Adult Luidia clathrata (65 mm radius) were collected during 1987-1988 from eastern Tampa Bay, Florida, USA (27 ° 43' 30" N, 82 ° 30' 50" W). Individuals were maintained in 38-1 recirculating aquaria (six individuals per aquarium) at room temperature (22-25°C) and salinities representative of field conditions (25-28%0). Each individual was fed 2-3 frozen Donax variabilis every other day until 1 week prior to enzyme analyses, at which time food was withheld to ensure an empty gut.
Tissues from several individuals were pooled for each enzyme assay. Each individual was weighed to the nearest 0.1 g and the pyloric caeca and gonads (if present) were weighed to the nearest 0.01 g. Condition indices for pyloric caeca and gonad were calculated for each individual as follows:
wet wt (g) ofpyloric caeca or gonad × 100 index =
total wet wt (g) of individual For each enzyme analysis, a 1 : 9 (w/v) tissue preparation
was prepared from the pooled tissues and respective hom- ogenizing buffer (see below) and placed in a 30 ml glass homogenizing tube. The tissue was homogenized with a
259

260 JAY R. LEVERONE et al.
Teflon ~ pestle by a high-speed electric rotor for 2 min on ice. The homogenate was placed in 15-ml tubes and centrifuged at 20,000g for 40 min at 4°C. The lipid layer was penetrated with a 21 gauge x 1--I/2" hypodermic needle and the super- natant extracted. The supernatant was either placed on ice for immediate use or frozen at -20°C for subsequent analysis. Freezing did not lead to any appreciable loss of activity for either enzyme system.
The concentrations of cations in the pyloric caeca and tube feet of Luidia clathrata were calculated by dividing the intracellular amount of each cation (nanomole intracellular cation per milligramme solute-free dry weight) reported by Diehl and Lawrence (1985) by the amount of intracellular water (g H20/g solute-free dry weight) derived from Diehl and Lawrence (1984) for individuals that had been exposed to salinities (S) of 15, 25 and 35%0 for 1 week. Since the weight of intracellular water was derived from the difference between wet tissue weight and freeze-dried tissue weight, it was assumed that 1.0 g water was equivalent to 1.0 ml water. These values were used to simulate the intracellular cation composition in the different tissues of L. clathrata under conditions of high, medium and low salinity. As such, the composition of cations from 35%0 S individuals used in enzyme analyses is designated "High Salinity Cation Concentration" (HSCC). The 250oo S cation composition is designated "Medium Salinity Cation Concentration" (MSCC), and the 15%o S cation composition is "Low Salinity Cation Concentration" (LSCC). The concen- trations of cations obtained from these calculations and utilized in the respective buffers are reported in Table I. Since no values are available for the intracellular amounts of ions in the gonads of L. clathrata, cation concentrations obtained from data on the pyloric caeca were used for assays of the gonads. The osmotic concentration of each cationic buffer was determined using an Osmette Precision Osmometer.
The activity of glucose-6-phosphate dehydrogenase (G-6- PDH; EC 1.1.1.49) was determined by measuring the re- duction of NADP ÷ in the presence of glucose-6-phosphate (Watts and Lawrence, 1990). Tissues were homogenized in 50mM triethanolamine buffer (pH 7.5) and 0.01mM NADP +. The final assay solution contained 50 mM tri- ethanolamine buffer (pH 7.5), 0.3 mM NADP ÷, and the appropriate cation concentrations for each tissue for each of the three salinities in 2.8ml. The addition of 0.1 ml supernatant (10%) indicated endogenous activity. The addition of 0.1 ml of glucose-6-phosphate (0.5mM final concentration) to the above solution indicated total G-6- PDH activity.
The activity of pyruvate kinase (PK; EC 2.7.1.40) was determined in a coupled assay by measuring the oxidation of NADH by lactate dehydrogenase (Watts and Lawrence, 1990). Tissues were homogenized in 50mM Tris buffer (pH 7.5). The final assay solution contained 50 mM Tris buffer (pH 7.5), 3mM ADP, 0.2mM NADH, lactate dehydrogenase (3 Units/ml assay solution), and the appro- priate cation concentrations for each tissue for each of the three salinities in 2.8 ml. The addition of 0.I ml supernatant
Table 2. Osmotic concentrations (mOsm/kg H20) of enzyme assay buffers which simulate tissue intracellular cation concentrations of L.
clathrata exposed to various salinities for 1 week*
Osmotic concentration
Pyloric caeca Enzyme Buffer and gonad Tube feet
Pyruvate kinase LSCC 442 550 MSCC 620 815 HSCC 798 1201
Glucose-6-phosphate LSCC 450 561 dehydrogenase MSCC 635 580
HSCC 820 1470
*LSCC, low salinity cation concentration; MSCC, medium salinity cation concentration; HSCC, high salinity cation concentration.
(10%) indicated endogenous activity. The addition of 0.1 ml of PEP (2 mM final concentration) to the above solution indicated total activity.
Enzyme activity was determined spectrophotometrically by the oxidation or reduction of pyridine nucleotides at 340 nm using a Varian DMS 80 u.v.-visible spectro- photometer. Assays were conducted at 25°C. The activities of the enzymes were calculated by the method of Lowry and Passoneau (1972) and expressed as either gramme wet weight or milligramme protein. Commercially prepared enzymes were used to determine whether changes in absorbance were directly dependent on enzyme concentrations. Protein concentrations were measured by the Bio-Rad *: method, with bovine gamma globulin as the standard. All biochemical reagents and chloride salts of each cation were purchased from Sigma Chemical Company, St Louis, Missouri, except potassium chloride, which came from J. T. Baker Company, Phillipsburg, New Jersey.
Specific activities are reported as the mean from three replicates plus or minus one standard error. Significant differences among means for the specific activities of each enzyme in each tissue were calculated by analysis of variance (ANOVA) (P < 0.05) and Duncan's New Multiple Range Test.
RESULTS
The osmotic concentrat ions of enzyme assay buffers are presented in Table 2. Osmotic concen- trations were lowest in LSCC buffers and highest in HSCC buffers for all enzyme systems. Osmotic concentrat ions for tube feet assay buffers were greater than the respective buffers from the pyloric caeca and gonad except for the MSCC buffer from the G-6-PDH assay. The osmotic concentrat ions of buffers of the same cation concentrat ion were similar between PK and G-6-PDH enzyme assay systems except for the MSCC buffers from the tube feet assays.
Table 1. Intracellular concentrations (mM) of selected cations in (A) pyloric caeca/gonads, and (B) tube feet of L. clathrata after 1 week exposure to various
salinities (from Diehl and Lawrence, 1984, 1985)
Salinity
Cation 15%0 25%0 35%0
(A) Pyloric caeca and gonads
(B) Tube feet
Na ÷ 93 t35 215 K ÷ 106 158 164
Ca 2+ 4.4 4.5 5.7 Mg 2+ 7.2 13.8 23.3
Na ÷ 167 267 427 K ÷ 85 112 131
Ca 2÷ 13.3 16.0 19.2 Mg 2+ 16.1 28.1 40.6
Cation effects on enzymes in L. clathrata
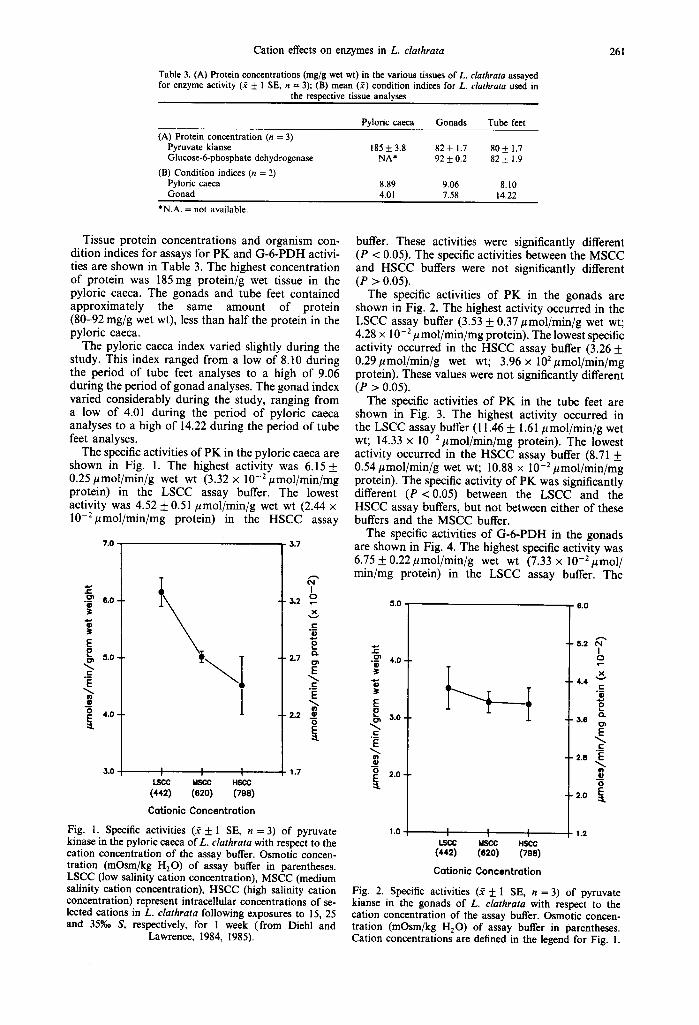
Table 3. (A) Protein concentrations (mg/g wet wt) in the various tissues of L. clathrata assayed for enzyme activity ($ _+ 1 SE, n = 3); (B) mean (~) condition indices for L. clathrata used in
the respective tissue analyses
261
Pyloric caeca Gonads Tube feet (A) Protein concentration (n = 3)
Pyruvate kianse 185 + 3.8 82 + 1.7 80 + 1.7 Glucose-6-phosphate dehydrogenase NA* 92 + 0.2 82 + 1.9
(B) Condition indices (n = 2) Pyloric caeca 8.89 9.06 8.10 Gonad 4•01 7.58 14.22
*N.A. = not available.
Tissue protein concentrations and organism con- dition indices for assays for PK and G-6-PDH activi- ties are shown in Table 3. The highest concentration of protein was 185 mg protein/g wet tissue in the pyloric caeca. The gonads and tube feet contained approximately the same amount of protein (80-92 mg/g wet wt), less than half the protein in the pyloric caeca•
The pyloric caeca index varied slightly during the study. This index ranged from a low of 8.10 during the period of tube feet analyses to a high of 9.06 during the period of gonad analyses. The gonad index varied considerably during the study, ranging from a low of 4.01 during the period of pyloric caeca analyses to a high of 14.22 during the period of tube feet analyses.
The specific activities of PK in the pyloric caeca are shown in Fig. 1. The highest activity was 6.15_+ 0.25/~mol/min/g wet wt (3.32 x 10 -2 / lmol /min /mg protein) in the LSCC assay buffer. The lowest activity was 4.52 + 0.51 #mol /min /g wet wt (2.44 x 10-2~mol /min /mg protein) in the HSCC assay
7.0 3.7
• -~ e.o- o
E o ~.0- On
._= E
~ 4 , 0 -
3.0
'\t I I I
LSCC MSCC HSCC (442) (620) (798)
Cationic Concentration
if , I O
3.2
o t~
2.7
E
E
2.2 O
1.7
Fig. 1. Specific activities (~ + 1 SE, n = 3) of pyruvate kinase in the pyloric caeca ofL. clathrata with respect to the cation concentration of the assay buffer. Osmotic concen- tration (mOsm/kg H20 ) of assay buffer in parentheses• LSCC (low salinity cation concentration), MSCC (medium salinity cation concentration), HSCC (high salinity cation concentration) represent intracellular concentrations of se- lected cations in L. clathrata following exposures to 15, 25 and 35%o S, respectively, for I week (from Diehl and
Lawrence, 1984, 1985).
buffer. These activities were significantly different (P < 0.05). The specific activities between the MSCC and HSCC buffers were not significantly different (P > 0.05).
The specific activities of PK in the gonads are shown in Fig. 2. The highest activity occurred in the LSCC assay buffer (3.53 + 0.37 #mol /min /g wet wt; 4.28 x 10-2/zmol/min/mg protein). The lowest specific activity occurred in the HSCC assay buffer (3.26 ___ 0.29/~mol/min/g wet wt; 3.96 x 102/~mol/min/mg protein). These values were not significantly different (P > 0.05).
The specific activities of PK in the tube feet are shown in Fig. 3. The highest activity occurred in the LSCC assay buffer (11.46 __+ 1.61 #mol /min /g wet wt; 14.33 x 10 -2/amol/min/mg protein). The lowest activity occurred in the HSCC assay buffer (8.71 + 0.54 #mol /min/g wet wt; 10.88 x 10 -2 #mol /min /mg protein). The specific activity of PK was significantly different (P <0.05) between the LSCC and the HSCC assay buffers, but not between either of these buffers and the MSCC buffer.
The specific activities of G-6-PDH in the gonads are shown in Fig. 4. The highest specific activity was 6.75__+ 0.22/~mol/min/g wet wt (7.33 x 10-2#mol / min/mg protein) in the LSCC assay buffer. The
5.0
• ~ 4.o
E
3.0
c E
_? .~ 2.0
1.0 I I LSCC MSCC HSCC
(442) (820) (798)
Cationic Concentration
8.0
5.2 I C)
x ,4.4
2 • 3.8
E c
- 2.11 "E
0
- 2.0 ~
1.2
Fig. 2. Specific activities (.~ + I SE, n = 3) of pyruvate kianse in the gonads of L. clathrata with respect to the cation concentration of the assay buffer. Osmotic concen- tration (mOsm/kg H20 ) of assay buffer in parentheses. Cation concentrations are defined in the legend for Fig. 1.

262
14.0 -
• ~ 12.0
E LO 10.0 O~
._c E
~ 8 .0 -
8.0 I I I LSCC MSCC HSCC
(550) (815) (1201)
Cationic Concentrat ion
JAY R. LEVERONE et al.
8.0 17.5
I
o x e.o 15.0
o~
o O. ~ 4.0
12.5 o~ E E p
E ._c E
10.0 ca ~ 2.0 o _~
0.0 7.5
Fig. 3. Specific activities ( ~ _ 1 SE, n = 3) of pyruvate kinase in the tube feet of L. clathrata with respect to the cation concentration of the assay buffer. Osmotic concentration (mOsm/kg H20) of assay buffer in parentheses. Cation concentrations are defined in the legend
for Fig. 1.
10.0
I I I LSCC MSCC HSCC
(s61) (580) (1,~7o)
Cationic Concentration
- 8.0 I 0
x
._c - e . 0
P o.
E • 4.0 ._=
E
0 2.0 ~L
0.0
Fig. 5. Specific activities (,f _ 1 SE, n = 3) of glucose-6- phosphate dehydrogenase in the tube feet of L. clathrata with respect to the cation concentration of the assay buffer. Osmotic concentration (mOsm/kg H20) of assay buffer in parentheses. Cation concentrations are defined in the legend
for Fig. 1.
lowest specific activity was 3.72_ 0.12/~mol/min/g wet wt (4.04 x 10-2#mol/min/mg protein) in the HSCC assay buffer. All specific activities were signifi- cantly different (P > 0.05).
The specific activities of G-6-PDH in the tube feet are shown in Fig. 5. The highest specific activity was 5.89 x 10- ]+0 .43gmol /min /g wet wt (7.18 x 10-3 # mol/min/mg protein) in the LSCC assay buffer. The lowest specific activity was 0.38x 10-1+ 0.01/zmol/min/g wet wt (0.48 × 10-3#mol/min/mg
B.O 8.7
.~ 8 .0 .
E E
.c E
4.0 ¸
0
2.0 I I I LSCC MSCC HSCC
(450) (635) (820)
Cationic Concentrat ion
if, • 7.4 I
O
t- 11.1 ~
E
"g
0 ,3.5 ~L
2.2
Fig. 4. Specific activities (£ _ 1 SE, n = 3) of glucose-6- phosphate dehydrogenase in the gonads of L. clathrata with respect to the cation concentration of the assay buffer. Osmotic concentration (mOsm/kg H20) of assay buffer in parentheses. Cation concentrations are defined in the legend
for Fig. 1.
protein) in the HSCC assay buffer. All specific activi- ties were significantly different (P > 0.05).
DISCUSSION
Pyruvate kinase (PK) and glucose-6-phosphate de- hydrogenase (G-6-PDH) function as rate-limiting enzymes for different points of the glycolytic path- way. Both enzymes show a high level of activity in a number of echinoderm species, including Luidia clathrata (Saito and Watts, 1989; Van der Plas and Oudejans, 1982; Ellington, 1982; Smith and Lawrence, 1987). The activities of these enzymes reflect the flux of substrates through glycolysis and the pentose phosphate pathway.
Studies of the effects of ions on enzyme activities in intracellular isosmotically regulating marine in- vertebrates have shown mixed results. The activity of glutamate dehydrogenase in crustaceans increases with an increase in monovalent cations (Schoffeniels and Gilles, 1963; Schoffeniels, 1964), while activities of isocitrate and malate dehydrogenases decrease (Gilles, 1969). The activity of lactate dehydrogenase in the polychaete Nereis succinea decreases with an increase in anions and cations (Clark and Zounes, 1977). The activities of glucose-6-phosphate dehydro- genase and 6-phosphogluconate dehydrogenase in the hepatopancreas and gill tissue in the mussel Mytilus edulis decrease with a decrease in the salinity (Carri6n et al., 1978). The activity of citrate synthase in the pyloric caeca of Luidia clathrata decreases with an increase in monovalent and divalent cations (Marsh and Lawrence, 1985). Of these studies, only Marsh and Lawrence (1985) considered the effects of physio- logical concentrations of ions on enzyme activity.
A high intracellular cation concentration simulat- ing the conditions in adult Luidia clathrata resulted in a corresponding decrease in the specific activity of

Cation effects on enzymes in L. clathrata 263
PK in all three tissues examined. However, when compared with the specific activity of G-6-PDH, the decreases within each tissue were small and usually insignificant. PK from the holothuroid Parastichopus parvimensis displayed a reduced sensitivity to high KCI and NaCi concentrations relative to PKs from other organisms (Bowlus and Somero, 1979). These differences may relate to the exceptionally high intra- cellular salt concentrations of stenohaline echino- derms (Zanders and Herrara, 1974). Considering the strategic position of PK at the terminus of glycolysis, this property would allow the organism to maintain its capacity for energy production through glucose metabolism. In this way, the salt insensitivity of echinoderm PKs may ensure the stability of their catalytic and regulatory functions, which is of prime importance to those species able to tolerate fluctu- ations in environmental salinity.
The effects of a simulated increase in intracellular cation concentration on the specific activity of G-6- PDH in tissues of adult Luidia clathrata were much greater. In all cases, an increase in assay cation concentration resulted in a significant decrease in the specific activity of this enzyme. In Mytilus edulis, a significant decrease in the specific activity of G-6- PDH was observed in the hepatopancreas (but not in the gills) of individuals held at 50% seawater for 8 days (Carrirn et al., 1978). Watts and Lawrence (1990), however, found no significant differences in G-6-PDH activity from the pyloric caeca of adult L. clathrata after exposure to different tem- perature:salinity combinations for 1 month. The dietary glucose available to the tissues of L. clathrata is apparently phosphorylated by hexokinase and directed into the pentose phosphate pathway (Smith and Lawrence, 1987). A key aspect of this alternate pathway of glucose metabolism is the generation of reducing power for biosynthetic purposes. Watts and Lawrence (1990) showed a seasonality in the activity of this pathway in L. clathrata which was associated with the organism's annual reproductive cycle. In contrast to PK, the relative sensitivity of G-6-PDH in tissues of adult L. clathrata could have major repercussions on an individual's anabolic capacity, The physio- logical Significance of these cation-dependent changes in pentose phosphate pathway fluxes is uncertain.
A number of metabolic regulatory enzymes exist as isozymes in the tissues of marine invert- ebrates (Hoffman and Radeke, 1978; Mustafa and Hochachka, 1971). Homogenates of cardiac stomach and pyloric caeca from 18 species of asteroids showed one to three bands of hexose-6-phosphate dehydro- genase activity on polyacrylamide gels (Mochizuki and Hori, 1976). The induction of isozymes is one mechanism by which seasonal control over metab- olism may be maintained (Livingstone, 1975), as well as assuring that tissue-specific metabolic require- ments involving the same enzymatically catalyzed reactions are met. Since PK activity of the pyloric caeca and gonads responded differently (under identi- cal assay conditions) in L. clathrata, this enzyme may exist in tissue-specific isozymic forms. Although the specific activities of all glycolytic enzymes examined in the pyloric caeca of L. clathrata varied seasonally (Watts and Lawrence, 1990), the mechanisms of regulation have not been determined.
A change in enzyme activity is only one of many biochemical, physiological and behavioral conse- quences for osmoconforming organisms in estuarine environments. The level, content, and caloric equiva- lents of lipid, carbohydrate, and protein change in the major body components of L. clathrata according to season, reproductive state, and nutritional condition (Lawrence, 1973). Low salinity causes a decrease in activity and rate of feeding (Forcucci and Lawrence, 1986), which can lead to a lowering of the organism's metabolism (Lawrence and Lane, 1982)due partially to a decrease in the activity of several metabolic enzymes (Watts and Lawrence, 1982). Periods of low prey availability induce changes in the feeding mode of L. clathrata in Tampa Bay (McClintock et al., 1983), which can provide an alternative food source for metabolism. Seasonal decreases in dissolved oxy- gen (Diehl et al., 1979) depress behavioral activity which can result in lower food intake. Although gonadal development in L. clathrata is correlated with a decrease in water temperature during the fall (Lawrence, 1973), this also coincides with increased salinities following the summer wet season in Tampa Bay (Simon, 1974). Changes in the activity of G-6- PDH as a result of changes in environmental salinity could greatly affect the biosynthetic capacity of the organism. This, in turn, could alter the level, if not the composition, of the gonadal tissue.
Acknowledgements--The authors thank W. Avery, Bay Studies Group, City of Tampa, for collecting L. clathrata, Mote Marine Laboratory for providing facilities and equip- ment, and Drs F. Friedl and B. Cochrane for critically reviewing the manuscript.
REFERENCES
Bowlus R. D. and Somero G. N. (1979) Solute compatibility with enzyme function and structure: rationales for the selection of osmotic agents and end-products of anaerobic metabolism in marine invertebrates. J. exp. Zool. 208, 137-152.
Carrirn A., Silva M., Cienfuegos E. and Ruiz-Amil M. (1978) Influencia de la temperatura y la salinidad sobre la glucosa-6-fosfato deshidrogenasa y 6-fosfogluconato deshidrogenasa del mejillrn (Mytilus edulis L.). Rev. Espanola Fis. 34, 241-246.
Clark M. E. and Zounes M. (1977) The effects of selected cell osmolytes on the activity of lactate dehydrogenase from the euryhaline polychaete, Nereis succinea. BioL Bull. 153, 468-484.
Diehl W. J. and Lawrence J. M. (1984) The effect of salinity on coelomic fluid osmolyte concentration and intracellu- lar water content in Luidia clathrata (Say) (Echinoder- mata: Asteroidea). Comp. Biochem. PhysioL 79A, 119-126.
Diehl W. J. and Lawrence J. M. (1985) Effect of salinity on the intracellular osmolytes in the pyloric caeca and tube feet of Luidia clathrata (Say) (Echinodermata: Astero- idea). Comp. Biochem. PhysioL 82A, 559-566.
Diehl W. J., McEdward L., Proffitt E., Rosenberg V. and Lawrence J. M. (1979) The response of Luidia clathrata (Echinodermata: Asteroidae) to hypoxia. Comp. Biochem. Physiol. 62A, 669471.
Eggleston L. V. and Krebs H. A. (1974) Regulation of the pentose phosphate cycle. Biochem. J. 138, 425-435.
Ellington W. R. (1982) Intermediary metabolism. In Echino- derm Nutrition (Edited by Jangoux M. and Lawrence J. M.), pp. 395-415. Balkema Press, Rotterdam.

264 JAY R. LEVERONE et al.
Forcueci D. and Lawrence J. M. (1986) Effect of low salinity on the activity, feeding, growth and absorption efficiency of Luidia clathrata (Echinodermata: Asteroidea). Mar. Biol. 92, 315-321.
Gilles R. (1969) Effect of various salts on the activity of enzymes implicated in amino-acid metabolism. Arch. Int. Physiol. Biochim. 77, 441-464.
Hoffman K. H. and Radeke U. (1978) Stability of invert- ebrate muscle pyruvate kinases: correlation with enzyme regulatory properties. Comp. Bioehem. Physiol. 61B, 321-325.
Lawrence J. M. (1973) Level, content and caloric equivalent of the lipid, carbohydrate and protein in the body com- ponents of Luidia clathrata (Echinodermata: Asteroidea: Platyasterida) in Tampa Bay. J. exp. Mar. Biol. Ecol. 11, 263-274.
Lawrence J. M. and Lane J. M. (1982) The utilization of nutrients by postmetamorphic echinoderms: respiration, growth and bioenergetics. In Echinoderm Nutrition (Edited by Jangoux M. and Lawrence J. M.), pp. 328-365. Balkema Press, Rotterdam.
Livingstone D. R. (1975) A comparison of the kinetic properties of pyruvate kinase in three populations of Mytilus edulis L. from different environments. In Pro- ceedings of 9th European Marine Biology Symposium (Edited by Barnes H.), pp. 151-164. The Aberdeen Uni- versity Press, Aberdeen.
Lowry O. and Passoneau J. (1972) A Flexible System of Enzymatic Analysis. Academic Press, New York.
Marsh A. and Lawrence J. M. (1985) The effects of cations on the activity of citrate synthase (EC 4.1.3.7) in Luidia clathrata (Say) (Echinodermata: Asteroidea). Comp. Bio- chem. Physiol. 81B, 767-770.
McClintock J. B., Klinger T. S. and Lawrence J. M. (1983) Extraoral feeding in Luidia clathrata (Say) (Echinoder- mata: Asteroidea)., Bull. Mar. Sci. 33, 171-172.
Mochizuki Y. and Hori S. H. (1976) Hexose-6-phosphate dehydrogenase in starfish. Comp. Biochem. Physiol. 54B, 489-494.
Mustafa T. and Hochachka P. W. (1971) Catalytic and regulatory properties of pyruvate kianses in tissues of a marine bivalve. J. biol. Chem. 246, 3169-3203.
Saito N. and Watts S. A. (1989) Activities of hexokinase, phosphofructokinase and pyruvate kinase in the body
wall, pyloric caeca, and tube feet of Asterias vulgaris: evidence of body wall as a major source of glycolytic activity. Comp. Bioehem. Physiol. 94B, 263-267.
Sarkissian I. V. and Boatwright D. T. (1974) Regulation by salt and by Krebs cycle metabolites of citrate synthase from an osmoregulator, white shrimp, Penaeus setiferus, and from a non-osmoregulator, sea anemone, Bunedosoma cavernata. Comp. Bioehem. Physiol. 49B, 325-333.
Schoffeniels E. (1964) Effect of inorganic ions on the activity of L-glutamic acid dehydrogenase. Life Sci. 3, 845-850.
Schoffeniels E. and Gilles R. (1963) Effects of cations on the activity of e-glutamic acid dehydrogenase. Life Sci. 2, 834-839.
Simon J. L. (1974) Tampa Bay estuarine system--a synop- sis. Fla. Sci. 37, 217-245.
Smith L. A. and Lawrence J. M. (1987) Glycolytic activity in the pyloric caeca of Luidia clathrata (Say) (Echinoder- mata: Asteroidea). Comp. Biochem. Physiol. 86B, 693~596.
Stickle W. B. and Diehl W. J. (1987) Effects of salinity on echinoderms. In Echinoderm Studies 2, pp. 235-285. Balkema Press, Rotterdam.
Van der Plas A. J. and Oudejans R. C. H. M. (1982) Changes in the activities of selected enzymes of intermedi- ary metabolism in the pyloric caeca and ovaries of Asterias rubens during the annual reproductive cycle. Comp. Bioehem. Physiol. 71B, 379-385.
Watts S. A. and Lawrence J. M. (1990) The effect of reproductive state, temperature and salinity on DNA and RNA levels and activities of metabolic enzymes of the pyloric caeca in the sea star, Luidia clathrata (Say). Phys. Zool. 63, 1196-1215.
Watts S. A. and Lawrence J. M. (1982) Effect of starvation on enzyme activities and nutrient stores in the pyloric caeca of Echinaster sp. (Echinodermata: Asteroidea). Comp. Biochem. Physiol. 72B, 461-463.
Yancey P. H., Clark M. C., Hand S. C., Bowlus R. D. and Somero G. N. (1982) Living with water stress: evolution of osmolyte systems. Science 217, 1214-1222.
Zanders I. P. and Herrera F. C. (1974) Ionic distribution and fluxes in holothurian tissues. Comp. Biochem. Physiol. 47A, 1153-1170.